
IHF and Fis as Escherichia coli Cell Cycle Regulators: Activation of the Replication Origin oriC and the Regulatory Cycle of the DnaA Initiator

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
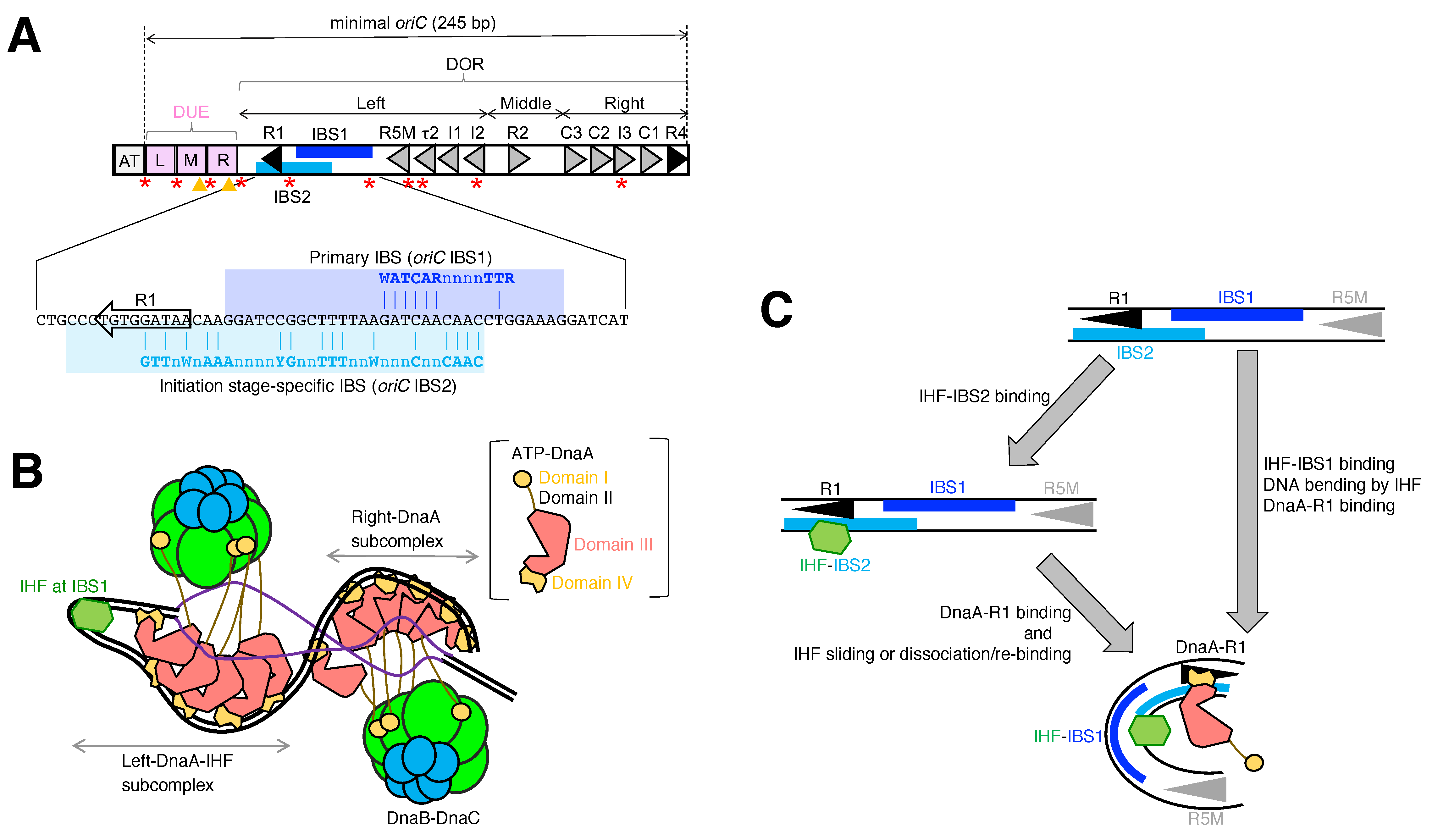
2. Initiation Complex at oriC
3. Regulations of oriC and DnaA
3.1. Replication-Coupled Negative Regulation of oriC: The Sequestration System
3.2. DnaA Cycle: RIDA, DDAH, DARS1/2
3.2.1. Replication-Coupled Negative Regulation of DnaA: The RIDA System
3.2.2. IHF-Dependent Negative Regulation of DnaA: The DDAH System
3.2.3. IHF- and Fis-Dependent Positive Regulation of DnaA: The DARS System
4. Cell Cycle-Coordinated Regulation of IHF/Fis Binding and Dissociation
4.1. Regulation of oriC-IHF Binding
4.2. Regulation of DARS2-Fis Binding and Dissociation
4.3. Regulation of DARS2-IHF Binding
4.4. Regulation of datA-IHF Binding
5. Perspectives
5.1. Regulatory Mechanism for oriC-IHF Dissociation
5.2. Possible Regulatory Mechanisms for IHF/Fis Binding and Dissociation at DARS2 and datA
5.3. Similar Mechanism of Other DNA Bending Proteins among Species
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Molan, K.; Žgur Bertok, D. Small Prokaryotic DNA-Binding Proteins Protect Genome Integrity throughout the Life Cycle. Int. J. Mol. Sci. 2022, 23, 4008. [Google Scholar] [CrossRef]
- Hołówka, J.; Zakrzewska-Czerwińska, J. Nucleoid Associated Proteins: The Small Organizers that Help to Cope with Stress. Front. Microbiol. 2020, 11, 590. [Google Scholar] [CrossRef]
- Dillon, S.C.; Dorman, C.J. Bacterial Nucleoid-Associated Proteins, Nucleoid Structure and Gene Expression. Nat. Rev. Microbiol. 2010, 8, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Katayama, T.; Kasho, K.; Kawakami, H. The DnaA Cycle in Escherichia coli: Activation, Function and Inactivation of the Initiator Protein. Front. Microbiol. 2017, 8, 2496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frimodt-Møller, J.; Charbon, G.; Løbner-Olesen, A. Control of Bacterial Chromosome Replication by Non-Coding Regions Outside the Origin. Curr. Genet. 2017, 63, 607–611. [Google Scholar] [CrossRef]
- Hansen, F.G.; Atlung, T. The DnaA Tale. Front. Microbiol. 2018, 9, 319. [Google Scholar] [CrossRef] [Green Version]
- Grimwade, J.E.; Leonard, A.C. Blocking, Bending, and Binding: Regulation of Initiation of Chromosome Replication during the Escherichia coli Cell Cycle by Transcriptional Modulators that Interact with Origin DNA. Front. Microbiol. 2021, 12, 732270. [Google Scholar] [CrossRef]
- Ali Azam, T.; Iwata, A.; Nishimura, A.; Ueda, S.; Ishihama, A. Growth Phase-Dependent Variation in Protein Composition of the Escherichia coli Nucleoid. J. Bacteriol. 1999, 181, 6361–6370. [Google Scholar] [CrossRef]
- Aeling, K.A.; Opel, M.L.; Steffen, N.R.; Tretyachenko-Ladokhina, V.; Hatfield, G.W.; Lathrop, R.H.; Senear, D.F. Indirect Recognition in Sequence-Specific DNA Binding by Escherichia coli Integration Host Factor: The Role of DNA Deformation Energy. J. Biol. Chem. 2006, 281, 39236–39248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, G.D.; Chan, E.W.; Leake, M.C.; Noy, A. Structural Interplay between DNA-Shape Protein Recognition and Supercoiling: The Case of IHF. Comput. Struct. Biotechnol. J. 2022, 20, 5264–5274. [Google Scholar] [CrossRef] [PubMed]
- Ball, C.A.; Osuna, R.; Ferguson, K.C.; Johnson, R.C. Dramatic Changes in Fis Levels upon Nutrient Upshift in Escherichia coli. J. Bacteriol. 1992, 174, 8043–8056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ninnemann, O.; Koch, C.; Kahmann, R. The E. coli Fis Promoter Is Subject to Stringent Control and Autoregulation. EMBO J. 1992, 11, 1075–1083. [Google Scholar] [CrossRef] [PubMed]
- Gerganova, V.; Maurer, S.; Stoliar, L.; Japaridze, A.; Dietler, G.; Nasser, W.; Kutateladze, T.; Travers, A.; Muskhelishvili, G. Upstream Binding of Idling RNA Polymerase Modulates Transcription Initiation from a Nearby Promoter. J. Biol. Chem. 2015, 290, 8095–8109. [Google Scholar] [CrossRef] [Green Version]
- Aiyar, S.E.; McLeod, S.M.; Ross, W.; Hirvonen, C.A.; Thomas, M.S.; Johnson, R.C.; Gourse, R.L. Architecture of Fis-Activated Transcription Complexes at the Escherichia coli rrnB P1 and rrnE P1 Promoters. J. Mol. Biol. 2002, 316, 501–516. [Google Scholar] [CrossRef]
- Stella, S.; Cascio, D.; Johnson, R.C. The Shape of the DNA Minor Groove Directs Binding by the DNA-Bending Protein Fis. Genes Dev. 2010, 24, 814–826. [Google Scholar] [CrossRef] [Green Version]
- Cassler, M.R.; Grimwade, J.E.; Leonard, A.C. Cell Cycle-Specific Changes in Nucleoprotein Complexes at a Chromosomal Replication Origin. EMBO J. 1995, 14, 5833–5841. [Google Scholar] [CrossRef] [PubMed]
- Sakiyama, Y.; Kasho, K.; Noguchi, Y.; Kawakami, H.; Katayama, T. Regulatory Dynamics in the Ternary DnaA Complex for Initiation of Chromosomal Replication in Escherichia coli. Nucleic Acids Res. 2017, 45, 12354–12373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasho, K.; Oshima, T.; Chumsakul, O.; Nakamura, K.; Fukamachi, K.; Katayama, T. Whole-Genome Analysis Reveals That the Nucleoid Protein IHF Predominantly Binds to the Replication Origin oriC Specifically at the Time of Initiation. Front. Microbiol. 2021, 12, 697712. [Google Scholar] [CrossRef]
- Ryan, V.T.; Grimwade, J.E.; Nievera, C.J.; Leonard, A.C. IHF and HU Stimulate Assembly of Pre-Replication Complexes at Escherichia coli oriC by Two Different Mechanisms. Mol. Microbiol. 2002, 46, 113–124. [Google Scholar] [CrossRef]
- Weigel, C.; Messer, W.; Preiss, S.; Welzeck, M.; Morigen; Boye, E. The Sequence Requirements for a Functional Escherichia coli Replication Origin Are Different for the Chromosome and a Minichromosome. Mol. Microbiol. 2001, 40, 498–507. [Google Scholar] [CrossRef]
- Kasho, K.; Fujimitsu, K.; Matoba, T.; Oshima, T.; Katayama, T. Timely Binding of IHF and Fis to DARS2 Regulates ATP-DnaA Production and Replication Initiation. Nucleic Acids Res. 2014, 42, 13134–13149. [Google Scholar] [CrossRef] [Green Version]
- Kasho, K.; Katayama, T. DnaA Binding Locus datA Promotes DnaA-ATP Hydrolysis to Enable Cell Cycle-Coordinated Replication Initiation. Proc. Natl. Acad. Sci. USA 2013, 110, 936–941. [Google Scholar] [CrossRef] [PubMed]
- Sakiyama, Y.; Nagata, M.; Yoshida, R.; Kasho, K.; Ozaki, S.; Katayama, T. Concerted Actions of DnaA Complexes with DNA-Unwinding Sequences within and Flanking Replication Origin oriC Promote DnaB Helicase Loading. J. Biol. Chem. 2022, 298, 102051. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, S.; Kawakami, H.; Nakamura, K.; Fujikawa, N.; Kagawa, W.; Park, S.-Y.; Yokoyama, S.; Kurumizaka, H.; Katayama, T. A Common Mechanism for the ATP-DnaA-Dependent Formation of Open Complexes at the Replication Origin. J. Biol. Chem. 2008, 283, 8351–8362. [Google Scholar] [CrossRef] [Green Version]
- Riber, L.; Frimodt-Møller, J.; Charbon, G.; Løbner-Olesen, A. Multiple DNA Binding Proteins Contribute to Timing of Chromosome Replication in E. coli. Front. Mol. Biosci. 2016, 3, 29. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, C.; Miyazaki, E.; Ozaki, S.; Abe, Y.; Katayama, T. DnaB Helicase Is Recruited to the Replication Initiation Complex via Binding of DnaA Domain I to the Lateral Surface of the DnaB N-Terminal Domain. J. Biol. Chem. 2020, 295, 1131–1143. [Google Scholar] [CrossRef]
- Sakiyama, Y.; Nishimura, M.; Hayashi, C.; Akama, Y.; Ozaki, S.; Katayama, T. The DnaA AAA+ Domain His136 Residue Directs DnaB Replicative Helicase to the Unwound Region of the Replication Origin, oriC. Front. Microbiol. 2018, 9, 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, L.; Davey, M.J.; O’Donnell, M. Replisome Assembly at oriC, the Replication Origin of E. coli, Reveals an Explanation for Initiation Sites Outside an Origin. Mol. Cell 1999, 4, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Katayama, T.; Ozaki, S.; Keyamura, K.; Fujimitsu, K. Regulation of the Replication Cycle: Conserved and Diverse Regulatory Systems for DnaA and oriC. Nat. Rev. Microbiol. 2010, 8, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Chodavarapu, S.; Kaguni, J.M. Replication Initiation in Bacteria. Enzym. 2016, 39, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Abe, Y.; Jo, T.; Matsuda, Y.; Matsunaga, C.; Katayama, T.; Ueda, T. Structure and Function of DnaA N-Terminal Domains: Specific Sites and Mechanisms in Inter-DnaA Interaction and in DnaB Helicase Loading on oriC. J. Biol. Chem. 2007, 282, 17816–17827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keyamura, K.; Abe, Y.; Higashi, M.; Ueda, T.; Katayama, T. DiaA Dynamics Are Coupled with Changes in Initial Origin Complexes Leading to Helicase Loading. J. Biol. Chem. 2009, 284, 25038–25050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zawilak-Pawlik, A.; Nowaczyk, M.; Zakrzewska-Czerwińska, J. The Role of the N-Terminal Domains of Bacterial Initiator DnaA in the Assembly and Regulation of the Bacterial Replication Initiation Complex. Genes 2017, 8, 136. [Google Scholar] [CrossRef] [Green Version]
- Keyamura, K.; Fujikawa, N.; Ishida, T.; Ozaki, S.; Su’etsugu, M.; Fujimitsu, K.; Kagawa, W.; Yokoyama, S.; Kurumizaka, H.; Katayama, T. The Interaction of DiaA and DnaA Regulates the Replication Cycle in E. coli by Directly Promoting ATP—DnaA-Specific Initiation Complexes. Genes Dev. 2007, 21, 2083–2099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutton, M.D.; Carr, K.M.; Vicente, M.; Kaguni, J.M. Escherichia coli DnaA Protein. The N-Terminal Domain and Loading of DnaB Helicase at the E. coli Chromosomal Origin. J. Biol. Chem. 1998, 273, 34255–34262. [Google Scholar] [CrossRef] [Green Version]
- Nozaki, S.; Ogawa, T. Determination of the Minimum Domain II Size of Escherichia coli DnaA Protein Essential for Cell Viability. Microbiology 2008, 154, 3379–3384. [Google Scholar] [CrossRef] [Green Version]
- Erzberger, J.P.; Mott, M.L.; Berger, J.M. Structural Basis for ATP-Dependent DnaA Assembly and Replication-Origin Remodeling. Nat. Struct. Mol. Biol. 2006, 13, 676–683. [Google Scholar] [CrossRef]
- Kawakami, H.; Keyamura, K.; Katayama, T. Formation of an ATP-DnaA-Specific Initiation Complex Requires DnaA Arginine 285, a Conserved Motif in the AAA+ Protein Family. J. Biol. Chem. 2005, 280, 27420–27430. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, Y.; Sakiyama, Y.; Kawakami, H.; Katayama, T. The Arg Fingers of Key DnaA Protomers Are Oriented Inward within the Replication Origin oriC and Stimulate DnaA Subcomplexes in the Initiation Complex. J. Biol. Chem. 2015, 290, 20295–20312. [Google Scholar] [CrossRef] [Green Version]
- Saxena, R.; Vasudevan, S.; Patil, D.; Ashoura, N.; Grimwade, J.E.; Crooke, E. Nucleotide-Induced Conformational Changes in Escherichia coli DnaA Protein Are Required for Bacterial ORC to Pre-RC Conversion at the Chromosomal Origin. Int. J. Mol. Sci. 2015, 16, 27897–27911. [Google Scholar] [CrossRef] [Green Version]
- Duderstadt, K.E.; Chuang, K.; Berger, J.M. DNA Stretching by Bacterial Initiators Promotes Replication Origin Opening. Nature 2011, 478, 209–213. [Google Scholar] [CrossRef]
- Hou, Y.; Kumar, P.; Aggarwal, M.; Sarkari, F.; Wolcott, K.M.; Chattoraj, D.K.; Crooke, E.; Saxena, R. The Linker Domain of the Initiator DnaA Contributes to Its ATP Binding and Membrane Association in E. coli Chromosomal Replication. Sci. Adv. 2022, 8, eabq6657. [Google Scholar] [CrossRef]
- Fujikawa, N.; Kurumizaka, H.; Nureki, O.; Terada, T.; Shirouzu, M.; Katayama, T.; Shigeyuki, Y. Structural Basis of Replication Origin Recognition by the DnaA Protein. Nucleic Acids Res. 2003, 31, 2077–2086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonard, A.C.; Grimwade, J.E. The Orisome: Structure and Function. Front. Microbiol. 2015, 6, 545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, M.; Noguchi, Y.; Sakiyama, Y.; Kawakami, H.; Katayama, T.; Takada, S. Near-Atomic Structural Model for Bacterial DNA Replication Initiation Complex and Its Functional Insights. Proc. Natl. Acad. Sci. USA 2016, 113, E8021–E8030. [Google Scholar] [CrossRef] [PubMed]
- Rozgaja, T.A.; Grimwade, J.E.; Iqbal, M.; Czerwonka, C.; Vora, M.; Leonard, A.C. Two Oppositely Oriented Arrays of Low-Affinity Recognition Sites in oriC Guide Progressive Binding of DnaA during Escherichia coli Pre-RC Assembly. Mol. Microbiol. 2011, 82, 475–488. [Google Scholar] [CrossRef] [Green Version]
- Ozaki, S.; Noguchi, Y.; Hayashi, Y.; Miyazaki, E.; Katayama, T. Differentiation of the DnaA-oriC Subcomplex for DNA Unwinding in a Replication Initiation Complex. J. Biol. Chem. 2012, 287, 37458–37471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozaki, S.; Katayama, T. Highly Organized DnaA-oriC Complexes Recruit the Single-Stranded DNA for Replication Initiation. Nucleic Acids Res. 2012, 40, 1648–1665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, R.; Ozaki, S.; Kawakami, H.; Katayama, T. Single-Stranded DNA Recruitment Mechanism in Replication Origin Unwinding by DnaA Initiator Protein and HU, an Evolutionary Ubiquitous Nucleoid Protein. Nucleic Acids Res. 2023, 51, 6286–6306. [Google Scholar] [CrossRef] [PubMed]
- Margulies, C.; Kaguni, J.M. The FIS Protein Fails to Block the Binding of DnaA Protein to oriC, the Escherichia coli Chromosomal Origin. Nucleic Acids Res. 1998, 26, 5170–5175. [Google Scholar] [CrossRef] [Green Version]
- Wold, S.; Crooke, E.; Skarstad, K. The Escherichia coli Fis Protein Prevents Initiation of DNA Replication from oriC in vitro. Nucleic Acids Res. 1996, 24, 3527–3532. [Google Scholar] [CrossRef] [Green Version]
- Ryan, V.T.; Grimwade, J.E.; Camara, J.E.; Crooke, E.; Leonard, A.C. Escherichia coli Prereplication Complex Assembly Is Regulated by Dynamic Interplay among Fis, IHF and DnaA. Mol. Microbiol. 2004, 51, 1347–1359. [Google Scholar] [CrossRef]
- Flåtten, I.; Skarstad, K. The Fis Protein Has a Stimulating Role in Initiation of Replication in Escherichia coli in vivo. PLoS ONE 2013, 8, e83562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnefoy, E.; Takahashi, M.; Yaniv, J.R. DNA-Binding Parameters of the HU Protein of Escherichia coli to Cruciform DNA. J. Mol. Biol. 1994, 242, 116–129. [Google Scholar] [CrossRef] [PubMed]
- Jaworski, P.; Donczew, R.; Mielke, T.; Weigel, C.; Stingl, K.; Zawilak-Pawlik, A. Structure and Function of the Campylobacter jejuni Chromosome Replication Origin. Front. Microbiol. 2018, 9, 1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaworski, P.; Zyla-Uklejewicz, D.; Nowaczyk-Cieszewska, M.; Donczew, R.; Mielke, T.; Weigel, C.; Zawilak-Pawlik, A. Putative Cooperative ATP-DnaA Binding to Double-Stranded DnaA Box and Single-Stranded DnaA-Trio Motif upon Helicobacter pylori Replication Initiation Complex Assembly. Int. J. Mol. Sci. 2021, 22, 6643. [Google Scholar] [CrossRef] [PubMed]
- Richardson, T.T.; Stevens, D.; Pelliciari, S.; Harran, O.; Sperlea, T.; Murray, H. Identification of a Basal System for Unwinding a Bacterial Chromosome Origin. EMBO J. 2019, 38, e101649. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Yoshida, R.; Katayama, T.; Ozaki, S. Thermotoga maritima oriC Involves a DNA Unwinding Element with Distinct Modules and a DnaA-Oligomerizing Region with a Novel Directional Binding Mode. J. Biol. Chem. 2023, 299, 104888. [Google Scholar] [CrossRef]
- Karaboja, X.; Wang, X. HBsu Is Required for the Initiation of DNA Replication in Bacillus subtilis. J. Bacteriol. 2022, 204, e0011922. [Google Scholar] [CrossRef]
- Krause, M.; Ru, B.; Lurz, R.; Messer, W. Complexes at the Replication Origin of Bacillus subtilis with Homologous and Heterologous DnaA Protein. J. Mol. Biol. 1997, 274, 365–380. [Google Scholar] [CrossRef]
- Stepankiw, N.; Kaidow, A.; Boye, E.; Bates, D. The Right Half of the Escherichia coli Replication Origin Is Not Essential for Viability, but Facilitates Multi-Forked Replication. Mol. Microbiol. 2009, 74, 467–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldminghaus, T.; Skarstad, K. The Escherichia coli SeqA Protein. Plasmid 2009, 61, 141–150. [Google Scholar] [CrossRef]
- Skarstad, K.; Katayama, T. Regulating DNA Replication in Bacteria. Cold Spring Harb. Perspect. Biol. 2013, 5, a012922. [Google Scholar] [CrossRef] [Green Version]
- Jha, J.K.; Chattoraj, D.K. Inactivation of Individual SeqA Binding Sites of the E. coli Origin Reveals Robustness of Replication Initiation Synchrony. PLoS ONE 2016, 11, e0166722. [Google Scholar] [CrossRef] [Green Version]
- Slater, S.; Wold, S.; Lu, M.; Boye, E.; Skarstad, K.; Kleckner, N. E. coli SeqA Protein Binds oriC in Two Different Methyl-Modulated Reactions Appropriate to Its Roles in DNA Replication Initiation and Origin Sequestration. Cell 1995, 82, 927–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujikawa, N.; Kurumizaka, H.; Nureki, O.; Tanaka, Y.; Yamazoe, M.; Hiraga, S.; Yokoyama, S. Structural and Biochemical Analyses of Hemimethylated DNA Binding by the SeqA Protein. Nucleic Acids Res. 2004, 32, 82–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guarné, A.; Brendler, T.; Zhao, Q.; Ghirlando, R.; Austin, S.; Yang, W. Crystal Structure of a SeqA-N Filament: Implications for DNA Replication and Chromosome Organization. EMBO J. 2005, 24, 1502–1511. [Google Scholar] [CrossRef] [Green Version]
- Torheim, N.K.; Skarstad, K. Escherichia coli SeqA Protein Affects DNA Topology and Inhibits Open Complex Formation at oriC. EMBO J. 1999, 18, 4882–4888. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Kim, H.K.; Kang, S.; Hong, C.B.; Yim, J.; Hwang, D.S. Expression of the SeqA Gene Is Negatively Modulated by the HU Protein in Escherichia coli. Mol. Gen. Genet. 2001, 264, 931–935. [Google Scholar] [CrossRef]
- Nievera, C.; Torgue, J.J.-C.; Grimwade, J.E.; Leonard, A.C. SeqA Blocking of DnaA-oriC Interactions Ensures Staged Assembly of the E. coli Pre-RC. Mol. Cell 2006, 24, 581–592. [Google Scholar] [CrossRef] [Green Version]
- Bach, T.; Skarstad, K. An oriC-like Distribution of GATC Sites Mediates Full Sequestration of Non-Origin Sequences in Escherichia coli. J. Mol. Biol. 2005, 350, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Campbell, J.L.; Boye, E.; Kleckner, N. SeqA: A Negative Modulator of Replication Initiation in E. coli. Cell 1994, 77, 413–426. [Google Scholar] [CrossRef]
- Katayama, T.; Kubota, T.; Kurokawa, K.; Crooke, E.; Sekimizu, K. The Initiator Function of DnaA Protein Is Negatively Regulated by the Sliding Clamp of the E. coli Chromosomal Replicase. Cell 1998, 94, 61–71. [Google Scholar] [CrossRef] [Green Version]
- Pomerantz, R.T.; O’Donnell, M. Replisome Mechanics: Insights into a Twin DNA Polymerase Machine. Trends Microbiol. 2007, 15, 156–164. [Google Scholar] [CrossRef]
- Kato, J.; Katayama, T. Hda, a Novel DnaA-Related Protein, Regulates the Replication Cycle in Escherichia coli. EMBO J. 2001, 20, 4253–4262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su’etsugu, M.; Nakamura, K.; Keyamura, K.; Kudo, Y.; Katayama, T. Hda Monomerization by ADP Binding Promotes Replicase. Mol. Microbiol. 2008, 283, 36118–36131. [Google Scholar] [CrossRef] [Green Version]
- Baxter, J.C.; Sutton, M.D. Evidence for Roles of the Escherichia coli Hda Protein beyond Regulatory Inactivation of DnaA. Mol. Microbiol. 2012, 85, 648–668. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.S.; Nanfara, M.T.; Chodavarapu, S.; Jin, K.S.; Babu, V.M.P.; Ghazy, M.A.; Chung, S.; Kaguni, J.M.; Sutton, M.D.; Cho, Y. Dynamic Assembly of Hda and the Sliding Clamp in the Regulation of Replication Licensing. Nucleic Acids Res. 2017, 45, 3888–3905. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Katayama, T. Novel Essential Residues of Hda for Interaction with DnaA in the Regulatory Inactivation of DnaA: Unique Roles for Hda AAA Box VI and VII Motifs. Mol. Microbiol. 2010, 76, 302–317. [Google Scholar] [CrossRef]
- Collier, J.; Shapiro, L. Feedback Control of DnaA-Mediated Replication Initiation by Replisome-Associated HdaA Protein in Caulobacter. J. Bacteriol. 2009, 191, 5706–5716. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H. Regulation of DNA Replication Licensing and Re-Replication by Cdt1. Int. J. Mol. Sci. 2021, 22, 5195. [Google Scholar] [CrossRef]
- Noirot-Gros, M.-F.; Velten, M.; Yoshimura, M.; McGovern, S.; Morimoto, T.; Ehrlich, S.D.; Ogasawara, N.; Polard, P.; Noirot, P. Functional Dissection of YabA, a Negative Regulator of DNA Replication Initiation in Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2006, 103, 2368–2373. [Google Scholar] [CrossRef] [PubMed]
- Kasho, K.; Tanaka, H.; Sakai, R.; Katayama, T. Cooperative DnaA Binding to the Negatively Supercoiled datA Locus Stimulates DnaA-ATP Hydrolysis. J. Biol. Chem. 2017, 292, 1251–1266. [Google Scholar] [CrossRef] [Green Version]
- Kitagawa, R.; Ozaki, T.; Moriya, S.; Ogawa, T. Negative Control of Replication Initiation by a Novel Chromosomal Locus Exhibiting Exceptional Affinity for Escherichia coli DnaA Protein. Genes Dev. 1998, 12, 3032–3043. [Google Scholar] [CrossRef] [Green Version]
- Kitagawa, R.; Mitsuki, H.; Okazaki, T.; Ogawa, T. A Novel DnaA Protein-Binding Site at 94.7 Min on the Escherichia coli Chromosome. Mol. Microbiol. 1996, 19, 1137–1147. [Google Scholar] [CrossRef] [PubMed]
- Nozaki, S.; Yamada, Y.; Ogawa, T. Initiator Titration Complex Formed at datA with the Aid of IHF Regulates Replication Timing in Escherichia coli. Genes cells 2009, 14, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Frimodt-Møller, J.; Charbon, G.; Krogfelt, K.A.; Løbner-Olesen, A. Control Regions for Chromosome Replication Are Conserved with Respect to Sequence and Location among Escherichia coli Strains. Front. Microbiol. 2015, 6, 1011. [Google Scholar] [CrossRef] [Green Version]
- Cagliero, C.; Grand, R.S.; Jones, M.B.; Jin, D.J.; O’Sullivan, J.M. Genome Conformation Capture Reveals That the Escherichia coli Chromosome Is Organized by Replication and Transcription. Nucleic Acids Res. 2013, 41, 6058–6071. [Google Scholar] [CrossRef] [Green Version]
- Valens, M.; Thiel, A.; Boccard, F. The MaoP/MaoS Site-Specific System Organizes the Ori Region of the E. coli Chromosome into a Macrodomain. PLoS Genet. 2016, 12, e1006309. [Google Scholar] [CrossRef] [Green Version]
- Nozaki, S.; Niki, H.; Ogawa, T. Replication Initiator DnaA of Escherichia coli Changes Its Assembly Form on the Replication Origin during the Cell Cycle. J. Bacteriol. 2009, 191, 4807–4814. [Google Scholar] [CrossRef] [Green Version]
- Fujimitsu, K.; Senriuchi, T.; Katayama, T. Specific Genomic Sequences of E. coli Promote Replicational Initiation by Directly Reactivating ADP-DnaA. Genes Dev. 2009, 23, 1221–1233. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, R.; Kasho, K.; Miyoshi, K.; Ozaki, S.; Kagawa, W.; Kurumizaka, H.; Katayama, T. A Novel Mode of DnaA-DnaA Interaction Promotes ADP Dissociation for Reactivation of Replication Initiation Activity. Nucleic Acids Res. 2019, 47, 11209–11224. [Google Scholar] [CrossRef]
- Miyoshi, K.; Tatsumoto, Y.; Ozaki, S.; Katayama, T. Negative Feedback for DARS2–Fis Complex by ATP–DnaA Supports the Cell Cycle-Coordinated Regulation for Chromosome Replication. Nucleic Acids Res. 2021, 49, 12820–12835. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Tanaka, H.; Kasho, K.; Fujimitsu, K.; Oshima, T.; Katayama, T. Chromosomal Location of the DnaA-Reactivating Sequence DARS2 Is Important to Regulate Timely Initiation of DNA Replication in Escherichia coli. Genes Cells 2016, 21, 1015–1023. [Google Scholar] [CrossRef] [PubMed]
- Saxena, R.; Fingland, N.; Patil, D.; Sharma, A.K.; Crooke, E. Crosstalk between DnaA Protein, the Initiator of Escherichia coli Chromosomal Replication, and Acidic Phospholipids Present in Bacterial Membranes. Int. J. Mol. Sci. 2013, 14, 8517–8537. [Google Scholar] [CrossRef] [Green Version]
- Chumsakul, O.; Nakamura, K.; Kurata, T.; Sakamoto, T.; Hobman, J.L.; Ogasawara, N.; Oshima, T.; Ishikawa, S. High-Resolution Mapping of in vivo Genomic Transcription Factor Binding Sites Using in situ DNase I Footprinting and ChIP-Seq. DNA Res. 2013, 20, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Sugimura, S.; Crothers, D.M. Stepwise Binding and Bending of DNA by Escherichia coli Integration Host Factor. Proc. Natl. Acad. Sci. USA 2006, 103, 18510–18514. [Google Scholar] [CrossRef]
- Cho, B.; Knight, E.M.; Barrett, C.L.; Palsson, B.Ø. Genome-Wide Analysis of Fis Binding in Escherichia coli Indicates a Causative Role for A-/AT-Tracts. Genome Res. 2008, 18, 900–910. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Ma, Q.; Xu, Y. Physical Properties of DNA May Direct the Binding of Nucleoid-Associated Proteins along the E. coli Genome. Math. Biosci. 2018, 301, 50–58. [Google Scholar] [CrossRef]
- Polaczek, P.; Kwan, K.; Liberles, D.A.; Campbell, J.L. Role of Architectural Elements in Combinatorial Regulation of Initiation of DNA Replication in Escherichia coli. Mol. Microbiol. 1997, 26, 261–275. [Google Scholar] [CrossRef]
- Su’etsugu, M.; Emoto, A.; Fujimitsu, K.; Keyamura, K.; Katayama, T. Transcriptional Control for Initiation of Chromosomal Replication in Escherichia coli: Fluctuation of the Level of Origin Transcription Ensures Timely Initiation. Genes Cells 2003, 8, 731–745. [Google Scholar] [CrossRef]
- Theisen, P.W.; Grimwade, J.E.; Leonard, A.C.; Bogan, J.A.; Helmstetter, C.E. Correlation of Gene Transcription with the Time of Initiation of Chromosome Replication in Escherichia coli. Mol. Microbiol. 1993, 10, 575–584. [Google Scholar] [CrossRef]
- Monteiro, L.M.O.; Sanches-Medeiros, A.; Westmann, C.A.; Silva-Rocha, R. Unraveling the Complex Interplay of Fis and IHF through Synthetic Promoter Engineering. Front. Bioeng. Biotechnol. 2020, 8, 510. [Google Scholar] [CrossRef] [PubMed]
- Kamar, R.I.; Banigan, E.J.; Erbas, A.; Giuntoli, R.D.; De La Cruz, M.O.; Johnson, R.C.; Marko, J.F. Facilitated Dissociation of Transcription Factors from Single DNA Binding Sites. Proc. Natl. Acad. Sci. USA 2017, 114, E3251–E3257. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.Y.; Zheng, W.; Chen, M.; Wolynes, P.G. Multiple Binding Configurations of Fis Protein Pairs on DNA: Facilitated Dissociation versus Cooperative Dissociation. J. Am. Chem. Soc. 2019, 141, 18113–18126. [Google Scholar] [CrossRef] [PubMed]
- Hołówka, J.; Trojanowski, D.; Janczak, M.; Jakimowicz, D.; Zakrzewska-Czerwińska, J. The Origin of Chromosomal Replication Is Asymmetrically Positioned on the Mycobacterial Nucleoid, and the Timing of Its Firing Depends on HupB. J. Bacteriol. 2018, 200. [Google Scholar] [CrossRef] [Green Version]
- Miyake, T.; Loch, C.M.; Li, R. Identification of a Multifunctional Domain in Autonomously Replicating Sequence-Binding Factor 1 Required for Transcriptional Activation, DNA Replication, and Gene Silencing. Mol. Cell. Biol. 2002, 22, 505–516. [Google Scholar] [CrossRef] [Green Version]
- Farge, G.É.R.; Falkenberg, M. Organization of DNA in Mammalian Mitochondria. Int. J. Mol. Sci. 2019, 20, 2770. [Google Scholar] [CrossRef] [Green Version]
- Zelenaya-Troitskaya, O.; Newman, S.M.; Okamoto, K.; Perlman, P.S.; Butow, R.A. Functions of the High Mobility Group Protein, Abf2p, in Mitochondrial DNA Segregation, Recombination and Copy Number in Saccharomyces cerevisiae. Genetics 1998, 148, 1763–1776. [Google Scholar] [CrossRef]
- Hillen, H.S.; Morozov, Y.I.; Sarfallah, A.; Temiakov, D.; Cramer, P. Structural Basis of Mitochondrial Transcription Initiation. Cell 2017, 171, 1072–1081.e10. [Google Scholar] [CrossRef] [Green Version]
- Loth, K.; Largillière, J.; Coste, F.; Culard, F.; Landon, C.; Castaing, B.; Delmas, A.F.; Paquet, F. New Protein-DNA Complexes in Archaea: A Small Monomeric Protein Induces a Sharp V-Turn DNA Structure. Sci. Rep. 2019, 9, 14253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatre, L.; Ricchetti, M. Prevalent Coordination of Mitochondrial DNA Transcription and Initiation of Replication with the Cell Cycle. Nucleic Acids Res. 2013, 41, 3068–3078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greci, M.D.; Bell, S.D. Archaeal DNA Replication. Annu. Rev. Microbiol. 2020, 74, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.S.; Johnson, R.C.; Marko, J.F. Concentration-Dependent Exchange Accelerates Turnover of Proteins Bound to Double-Stranded DNA. Nucleic Acids Res. 2011, 39, 2249–2259. [Google Scholar] [CrossRef] [Green Version]
- Strzałka, A.; Kois-Ostrowska, A.; Kędra, M.; Łebkowski, T.; Bieniarz, G.; Szafran, M.J.; Jakimowicz, D. Enhanced Binding of an HU Homologue under Increased DNA Supercoiling Preserves Chromosome Organisation and Sustains Streptomyces Hyphal Growth. Nucleic Acids Res. 2022, 50, 12202–12216. [Google Scholar] [CrossRef]
- Ahmed, S.M.; Dröge, P. Chromatin Architectural Factors as Safeguards against Excessive Supercoiling during DNA Replication. Int. J. Mol. Sci. 2020, 21, 4504. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.S.; Haakonsen, D.L.; Zeng, W.; Schumacher, M.A.; Laub, M.T. A Bacterial Chromosome Structuring Protein Binds Overtwisted DNA to Stimulate Type II Topoisomerases and Enable DNA Replication. Cell 2018, 175, 583–597.e23. [Google Scholar] [CrossRef] [Green Version]
- Datta, C.; Jha, R.K.; Ganguly, S.; Nagaraja, V. NapA (Rv0430), a Novel Nucleoid-Associated Protein That Regulates a Virulence Operon in Mycobacterium tuberculosis in a Supercoiling-Dependent Manner. J. Mol. Biol. 2019, 431, 1576–1591. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, A.; Sokunbi, A.O.; Grove, A. DNA Protection by Histone-like Protein HU from the Hyperthermophilic Eubacterium Thermotoga maritima. Nucleic Acids Res. 2008, 36, 3956–3968. [Google Scholar] [CrossRef] [Green Version]
- Parekh, V.J.; Wien, F.; Grange, W.; De Long, T.A.; Arluison, V.; Sinden, R.R. Crucial Role of the C-Terminal Domain of Hfq Protein in Genomic Instability. Microorganisms 2020, 8, 1598. [Google Scholar] [CrossRef] [PubMed]
- Hoshina, S.; Yura, K.; Teranishi, H.; Kiyasu, N.; Tominaga, A.; Kadoma, H.; Nakatsuka, A.; Kunichika, T.; Obuse, C.; Waga, S. Human Origin Recognition Complex Binds Preferentially to G-Quadruplex-Preferable RNA and Single-Stranded DNA. J. Biol. Chem. 2013, 288, 30161–30171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remus, D.; Beall, E.L.; Botchan, M.R. DNA Topology, Not DNA Sequence, Is a Critical Determinant for Drosophila ORC–DNA Binding. EMBO J. 2004, 23, 897–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasho, K.; Ozaki, S.; Katayama, T. IHF and Fis as Escherichia coli Cell Cycle Regulators: Activation of the Replication Origin oriC and the Regulatory Cycle of the DnaA Initiator. Int. J. Mol. Sci. 2023, 24, 11572. https://doi.org/10.3390/ijms241411572
Kasho K, Ozaki S, Katayama T. IHF and Fis as Escherichia coli Cell Cycle Regulators: Activation of the Replication Origin oriC and the Regulatory Cycle of the DnaA Initiator. International Journal of Molecular Sciences. 2023; 24(14):11572. https://doi.org/10.3390/ijms241411572
Chicago/Turabian StyleKasho, Kazutoshi, Shogo Ozaki, and Tsutomu Katayama. 2023. "IHF and Fis as Escherichia coli Cell Cycle Regulators: Activation of the Replication Origin oriC and the Regulatory Cycle of the DnaA Initiator" International Journal of Molecular Sciences 24, no. 14: 11572. https://doi.org/10.3390/ijms241411572