
Approach to Functions of BHLHE41/DEC2 in Non-Small Lung Cancer Development

 ,
,
Abstract
:1. Introduction
2. The Functions of BHLHE41/DEC2 in Differentiation
3. Oncogenic and Tumor-Suppressive Functions of BHLHE40/DEC1 and BHLHE41/DEC2
4. BHLHE41/DEC2 as a Tumor Suppressor Protein in NSCLC Development
5. Post-Translational Modifications Regulate the Functions of BHLHE40/DEC1 and BHLHE41/DEC2
6. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Honma, S.; Kawamoto, T.; Takagi, Y.; Fujimoto, K.; Sato, F.; Noshiro, M.; Kato, Y.; Honma, K. Dec1 and Dec2 are regulators of the mammalian molecular clock. Nature 2002, 419, 841–844. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Jones, C.R.; Fujiki, N.; Xu, Y.; Guo, B.; Holder, J.L., Jr.; Rossner, M.J.; Nishino, S.; Fu, Y.H. The transcriptional repressor DEC2 regulates sleep length in mammals. Science 2009, 325, 866–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegrino, R.; Kavakli, I.H.; Goel, N.; Cardinale, C.J.; Dinges, D.F.; Kuna, S.T.; Maislin, G.; Van Dongen, H.P.; Tufik, S.; Hogenesch, J.B.; et al. A novel BHLHE41 variant is associated with short sleep and resistance to sleep deprivation in humans. Sleep 2014, 37, 1327–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirano, A.; Hsu, P.K.; Zhang, L.; Xing, L.; McMahon, T.; Yamazaki, M.; Ptacek, L.J.; Fu, Y.H. DEC2 modulates orexin expression and regulates sleep. Proc. Natl. Acad. Sci. USA 2018, 115, 3434–3439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.O.; Angkasekwinai, P.; Zhu, J.; Peng, J.; Liu, Z.; Nurieva, R.; Liu, X.; Chung, Y.; Chang, S.H.; Sun, B.; et al. Requirement for the basic helix-loop-helix transcription factor Dec2 in initial TH2 lineage commitment. Nat. Immunol. 2009, 10, 1260–1266. [Google Scholar] [CrossRef] [Green Version]
- Kreslavsky, T.; Vilagos, B.; Tagoh, H.; Poliakova, D.K.; Schwickert, T.A.; Wohner, M.; Jaritz, M.; Weiss, S.; Taneja, R.; Rossner, M.J.; et al. Essential role for the transcription factor Bhlhe41 in regulating the development, self-renewal and BCR repertoire of B-1a cells. Nat. Immunol. 2017, 18, 442–455. [Google Scholar] [CrossRef] [Green Version]
- Rauschmeier, R.; Gustafsson, C.; Reinhardt, A.; A-Gonzalez, N.; Tortola, L.; Cansever, D.; Subramanian, S.; Taneja, R.; Rossner, M.J.; Sieweke, M.H.; et al. Bhlhe40 and Bhlhe41 transcription factors regulate alveolar macrophage self-renewal and identity. EMBO J. 2019, 38, e101233. [Google Scholar] [CrossRef]
- Azmi, S.; Ozog, A.; Taneja, R. Sharp-1/DEC2 inhibits skeletal muscle differentiation through repression of myogenic transcription factors. J. Biol. Chem. 2004, 279, 52643–52652. [Google Scholar] [CrossRef] [Green Version]
- Morosetti, R.; Mirabella, M.; Gliubizzi, C.; Broccolini, A.; De Angelis, L.; Tagliafico, E.; Sampaolesi, M.; Gidaro, T.; Papacci, M.; Roncaglia, E.; et al. MyoD expression restores defective myogenic differentiation of human mesoangioblasts from inclusion-body myositis muscle. Proc. Natl. Acad. Sci. USA 2006, 103, 16995–17000. [Google Scholar] [CrossRef]
- Gulbagci, N.T.; Li, L.; Ling, B.; Gopinadhan, S.; Walsh, M.; Rossner, M.; Nave, K.A.; Taneja, R. SHARP1/DEC2 inhibits adipogenic differentiation by regulating the activity of C/EBP. EMBO Rep. 2009, 10, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Sasamoto, T.; Fujimoto, K.; Kanawa, M.; Kimura, J.; Takeuchi, J.; Harada, N.; Goto, N.; Kawamoto, T.; Noshiro, M.; Suardita, K.; et al. DEC2 is a negative regulator for the proliferation and differentiation of chondrocyte lineage-committed mesenchymal stem cells. Int. J. Mol. Med. 2016, 38, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Azim, S.T. Reshma, Embryonic expression of mSharp-1/mDEC2, which encodes a basic helix–loop–helix transcription factor. Mech. Dev. 2002, 114, 181–185. [Google Scholar] [CrossRef]
- Ornitz, D.M.; Marie, P.J. Fibroblast growth factor signaling in skeletal development and disease. Genes Dev. 2015, 29, 1463–1486. [Google Scholar] [CrossRef] [Green Version]
- Falvella, F.S.; Colombo, F.; Spinola, M.; Campiglio, M.; Pastorino, U.; Dragani, T.A. BHLHB3: A candidate tumor suppressor in lung cancer. Oncogene 2008, 27, 3761–3764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bigot, P.; Colli, L.M.; Machiela, M.J.; Jessop, L.; Myers, T.A.; Carrouget, J.; Wagner, S.; Roberson, D.; Eymerit, C.; Henrion, D.; et al. Functional characterization of the 12p12.1 renal cancer-susceptibility locus implicates BHLHE41. Nat. Commun. 2016, 7, 12098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semenza, G.L. The hypoxic tumor microenvironment: A driving force for breast cancer progression. Biochim. Biophys. Acta 2016, 1863, 382–391. [Google Scholar] [CrossRef]
- Miyazaki, K.; Kawamoto, T.; Tanimoto, K.; Nishiyama, M.; Honda, H.; Kato, Y. Identification of functional hypoxia response elements in the promoter region of the DEC1 and DEC2 genes. J. Biol. Chem. 2002, 277, 47014–47021. [Google Scholar] [CrossRef] [Green Version]
- Montagner, M.; Enzo, E.; Forcato, M.; Zanconato, F.; Parenti, A.; Rampazzo, E.; Basso, G.; Leo, G.; Rosato, A.; Bicciato, S.; et al. SHARP1 suppresses breast cancer metastasis by promoting degradation of hypoxia-inducible factors. Nature 2012, 487, 380–384. [Google Scholar] [CrossRef]
- Chen, X.; Iliopoulos, D.; Zhang, Q.; Tang, Q.; Greenblatt, M.B.; Hatziapostolou, M.; Lim, E.; Tam, W.L.; Ni, M.; Chen, Y.; et al. XBP1 promotes triple-negative breast cancer by controlling the HIF1α pathway. Nature 2014, 508, 103–107. [Google Scholar] [CrossRef] [Green Version]
- Sato, F.; Bhawal, U.K.; Kawamoto, T.; Fujimoto, K.; Imaizumi, T.; Imanaka, T.; Kondo, J.; Koyanagi, S.; Noshiro, M.; Yoshida, H.; et al. Basic-helix-loop-helix (bHLH) transcription factor DEC2 negatively regulates vascular endothelial growth factor expression. Genes Cells 2008, 13, 131–144. [Google Scholar] [CrossRef]
- Zhou, Z.H.; Wang, B.; Cheng, X.B.; Zhang, X.E.; Tang, J.; Tang, W.J.; Gu, L. Roles of SHARP1 in thyroid cancer. Mol. Med. Rep. 2016, 13, 5365–5371. [Google Scholar] [CrossRef] [PubMed]
- Mandriota, S.J.; Turner, K.J.; Davies, D.R.; Murray, P.G.; Morgan, N.V.; Sowter, H.M.; Wykoff, C.C.; Maher, E.R.; Harris, A.L.; Ratcliffe, P.J.; et al. HIF activation identifies early lesions in VHL kidneys: Evidence for site-specific tumor suppressor function in the nephron. Cancer Cell 2002, 1, 459–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazumder, S.; Higgins, P.J.; Samarakoon, R. Downstream Targets of VHL/HIF-α Signaling in Renal Clear Cell Carcinoma Progression: Mechanisms and Therapeutic Relevance. Cancers 2023, 15, 1316. [Google Scholar] [CrossRef] [PubMed]
- Grampp, S.; Schmid, V.; Salama, R.; Lauer, V.; Kranz, F.; Platt, J.L.; Smythies, J.; Choudhry, H.; Goppelt-Struebe, M.; Ratcliffe, P.J.; et al. Multiple renal cancer susceptibility polymorphisms modulate the HIF pathway. PLoS Genet. 2017, 13, e1006872. [Google Scholar] [CrossRef] [Green Version]
- Nassar, A.H.; Abou Alaiwi, S.; Baca, S.C.; Adib, E.; Corona, R.I.; Seo, J.H.; Fonseca, M.A.S.; Spisak, S.; El Zarif, T.; Tisza, V.; et al. Epigenomic charting and functional annotation of risk loci in renal cell carcinoma. Nat. Commun. 2023, 14, 346. [Google Scholar] [CrossRef]
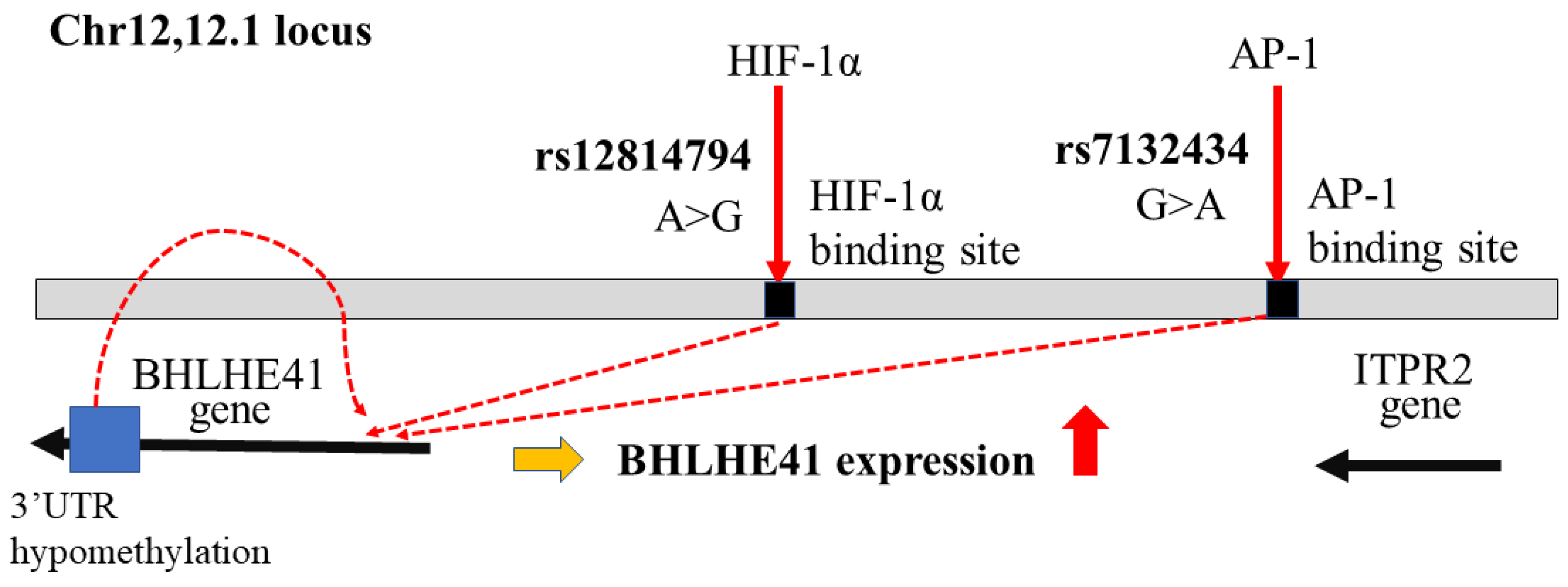
- Shen, Z.; Zhu, L.; Zhang, C.; Cui, X.; Lu, J. Overexpression of BHLHE41, correlated with DNA hypomethylation in 3′UTR region, promotes the growth of human clear cell renal cell carcinoma. Oncol. Rep. 2019, 41, 2137–2147. [Google Scholar] [CrossRef] [Green Version]
- Winters, A.C.; Bernt, K.M. MLL-Rearranged Leukemias-An Update on Science and Clinical Approaches. Front. Pediatr. 2017, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Deshpande, A.J.; Chen, L.; Fazio, M.; Sinha, A.U.; Bernt, K.M.; Banka, D.; Dias, S.; Chang, J.; Olhava, E.J.; Daigle, S.R.; et al. Leukemic transformation by the MLL-AF6 fusion oncogene requires the H3K79 methyltransferase Dot1l. Blood 2013, 121, 2533–2541. [Google Scholar] [CrossRef] [Green Version]
- Manara, E.; Baron, E.; Tregnago, C.; Aveic, S.; Bisio, V.; Bresolin, S.; Masetti, R.; Locatelli, F.; Basso, G.; Pigazzi, M. MLL-AF6 fusion oncogene sequesters AF6 into the nucleus to trigger RAS activation in myeloid leukemia. Blood 2014, 124, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Numata, A.; Kwok, H.S.; Kawasaki, A.; Li, J.; Zhou, Q.L.; Kerry, J.; Benoukraf, T.; Bararia, D.; Li, F.; Ballabio, E.; et al. The basic helix-loop-helix transcription factor SHARP1 is an oncogenic driver in MLL-AF6 acute myelogenous leukemia. Nat. Commun. 2018, 9, 1622. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Shen, Q.; Kim, H.T.; Bissonnette, R.P.; Lamph, W.W.; Yan, B.; Brown, P.H. The rexinoid bexarotene represses cyclin D1 transcription by inducing the DEC2 transcriptional repressor. Breast Cancer Res. Treat. 2011, 128, 667–677. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Sato, H.; Suzuki, T.; Yoshizawa, T.; Morohashi, S.; Seino, H.; Kawamoto, T.; Fujimoto, K.; Kato, Y.; Kijima, H. Involvement of c-Myc in the proliferation of MCF-7 human breast cancer cells induced by bHLH transcription factor DEC2. Int. J. Mol. Med. 2015, 35, 815–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Zhao, N.; Zheng, Q.; Zhang, D.; Liu, Y. BHLHE41 promotes U87 and U251 cell proliferation via ERK/cyclinD1 signaling pathway. Cancer Manag. Res. 2019, 11, 7657–7672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Settleman, J. EMT, cancer stem cells and drug resistance: An emerging axis of evil in the war on cancer. Oncogene 2010, 29, 4741–4751. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Tilló, E.; Liu, Y.; de Barrios, O.; Siles, L.; Fanlo, L.; Cuatrecasas, M.; Darling, D.S.; Dean, D.C.; Castells, A.; Postigo, A. EMT-activating transcription factors in cancer: Beyond EMT and tumor invasiveness. Cell Mol. Life Sci. 2012, 69, 3429–3456. [Google Scholar] [CrossRef]
- Sato, F.; Kawamura, H.; Wu, Y.; Sato, H.; Jin, D.; Bhawal, U.K.; Kawamoto, T.; Fujimoto, K.; Noshiro, M.; Seino, H.; et al. The basic helix-loop-helix transcription factor DEC2 inhibits TGF-β-induced tumor progression in human pancreatic cancer BxPC-3 cells. Int. J. Mol. Med. 2012, 30, 495–501. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Dong, Q.J.; Wan, Z.A.; Gao, S.; Tu, S.L.; Chai, R. BHLHE41 Overexpression Alleviates the Malignant Behavior of Colon Cancer Cells Induced by Hypoxia via Modulating HIF-1α/EMT Pathway. Gastroenterol. Res. Pract. 2022, 2022, 6972331. [Google Scholar] [CrossRef]
- Asanoma, K.; Liu, G.; Yamane, T.; Miyanari, Y.; Takao, T.; Yagi, H.; Ohgami, T.; Ichinoe, A.; Sonoda, K.; Wake, N.; et al. Regulation of the Mechanism of TWIST1 Transcription by BHLHE40 and BHLHE41 in Cancer Cells. Mol. Cell Biol. 2015, 35, 4096–4109. [Google Scholar] [CrossRef] [Green Version]
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef]
- Nagata, T.; Minami, K.; Yamamoto, M.; Hiraki, T.; Idogawa, M.; Fujimoto, K.; Kageyama, S.; Tabata, K.; Kawahara, K.; Ueda, K.; et al. BHLHE41/DEC2 Expression Induces Autophagic Cell Death in Lung Cancer Cells and Is Associated with Favorable Prognosis for Patients with Lung Adenocarcinoma. Int. J. Mol. Sci. 2021, 22, 11509. [Google Scholar] [CrossRef]
- Kunz, K.; Wagner, K.; Mendler, L.; Holper, S.; Dehne, N.; Muller, S. SUMO Signaling by Hypoxic Inactivation of SUMO-Specific Isopeptidases. Cell Rep. 2016, 16, 3075–3086. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.; Xing, X.; Li, S.; Bi, H.; Yang, C.; Zhao, F.; Liu, Y.; Ao, X.; Chang, A.K.; Wu, H. SUMOylation of DEC1 protein regulates its transcriptional activity and enhances its stability. PLoS ONE 2011, 6, e23046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Rao, V.K.; Kok, W.K.; Roy, D.N.; Sethi, S.; Ling, B.M.; Lee, M.B.; Taneja, R. SUMO modification of Stra13 is required for repression of cyclin D1 expression and cellular growth arrest. PLoS ONE 2012, 7, e43137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Shankar, S.R.; Kher, D.; Ling, B.M.; Taneja, R. Sumoylation of the basic helix-loop-helix transcription factor sharp-1 regulates recruitment of the histone methyltransferase G9a and function in myogenesis. J. Biol. Chem. 2013, 288, 17654–17662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Wang, T.; Mei, W.; Li, D.; Cai, R.; Zuo, Y.; Cheng, J. Small ubiquitin-like modifier (SUMO) protein-specific protease 1 de-SUMOylates Sharp-1 protein and controls adipocyte differentiation. J. Biol. Chem. 2014, 289, 22358–22364. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; D’Annibale, S.; Magliozzi, R.; Low, T.Y.; Jansen, P.; Shaltiel, I.A.; Mohammed, S.; Heck, A.J.; Medema, R.H.; Guardavaccaro, D. USP17- and SCFβTrCP-regulated degradation of DEC1 controls the DNA damage response. Mol. Cell. Biol. 2014, 34, 4177–4185. [Google Scholar] [CrossRef]
- Jones, S. An overview of the basic helix-loop-helix proteins. Genome Biol. 2004, 5, 226. [Google Scholar] [CrossRef] [Green Version]
- Sato, F.; Bhawal, U.K.; Yoshimura, T.; Muragaki, Y. DEC1 and DEC2 Crosstalk between Circadian Rhythm and Tumor Progression. J. Cancer 2016, 7, 153–159. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, K.; Hamaguchi, H.; Hashiba, T.; Nakamura, T.; Kawamoto, T.; Sato, F.; Noshiro, M.; Bhawal, U.K.; Suardita, K.; Kato, Y. Transcriptional repression by the basic helix-loop-helix protein Dec2: Multiple mechanisms through E-box elements. Int. J. Mol. Med. 2007, 19, 925–932. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Xie, M.; Yang, J.; Yang, D.; Deng, R.; Wan, Y.; Yan, B. The expression of antiapoptotic protein survivin is transcriptionally upregulated by DEC1 primarily through multiple sp1 binding sites in the proximal promoter. Oncogene 2006, 25, 3296–3306. [Google Scholar] [CrossRef] [Green Version]
- Qi, H.; Cao, Q.; Liu, Q. MicroRNA-16 directly binds to DEC2 and inactivates the TLR4 signaling pathway to inhibit lupus nephritis-induced kidney tissue hyperplasia and mesangial cell proliferation. Int. Immunopharmacol. 2020, 88, 106859. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Pan, Y.; Zhou, Q.; Liang, C.; Wong, C.C.; Zhou, Y.; Huang, D.; Liu, W.; Zhai, J.; Gou, H.; et al. METTL3 Inhibits Antitumor Immunity by Targeting m(6)A-BHLHE41-CXCL1/CXCR2 Axis to Promote Colorectal Cancer. Gastroenterology 2022, 163, 891–907. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Phenotype | Cells | Spices | Model | Effect | Gegens Which Receive Direct and/or Indirect Effects | REF |
|---|---|---|---|---|---|---|
| Th2 differentiation promotion | Th2 | mouse | Bhlhe41 knockout mouse | induce | JunB, Gata3 | [5] |
| B-1a cell differentiation promotion | B-1a | mouse | Bhlhe41 knockout mouse | suppress | Ccnh, Cdkl1, Cks2, Usp28 et al. | [6] |
| induce | Ighm, Dusp1, cytokine signal receptor component et al. | |||||
| alveolar macrophage differentiation promotion | Alveolar macrophages | mouse | Bhlhe40 and Bhlhe41 double knockout mice | suppress | specific genes of other subtypes of macrophage | [7] |
| induce | Epcam | |||||
| myogenesis inhibition | C2C12 | mouse | in vitro induction | suppress | MyoD, E47 | [8] |
| myogenesis inhibition | Mesoangioblasts of inclusion body myositis | human | in vitro induction | suppress | MyoD | [9] |
| adipose cell differentiation inhibition | 3T3L1 | mouse | in vitro induction | suppress | C/EBPα, PPAR γ | [10] |
| chondrogenic differentiation inhibition | human mesenchymal stem cells | human | in vitro induction | suppress | Cyclin D1, several chondrocyte-related genes possibly thorough Fgf18 | [11] |
| induce | p16INK4, p21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Furukawa, T.; Mimami, K.; Nagata, T.; Yamamoto, M.; Sato, M.; Tanimoto, A. Approach to Functions of BHLHE41/DEC2 in Non-Small Lung Cancer Development. Int. J. Mol. Sci. 2023, 24, 11731. https://doi.org/10.3390/ijms241411731
Furukawa T, Mimami K, Nagata T, Yamamoto M, Sato M, Tanimoto A. Approach to Functions of BHLHE41/DEC2 in Non-Small Lung Cancer Development. International Journal of Molecular Sciences. 2023; 24(14):11731. https://doi.org/10.3390/ijms241411731
Chicago/Turabian StyleFurukawa, Tatsuhiko, Kentaro Mimami, Toshiyuki Nagata, Masatasu Yamamoto, Masami Sato, and Akihide Tanimoto. 2023. "Approach to Functions of BHLHE41/DEC2 in Non-Small Lung Cancer Development" International Journal of Molecular Sciences 24, no. 14: 11731. https://doi.org/10.3390/ijms241411731