
Investigating the Impact of Selective Modulators on the Renin–Angiotensin–Aldosterone System: Unraveling Their Off-Target Perturbations of Transmembrane Ionic Currents

Abstract
:1. Introduction
2. Apocynin (Acetovanillone, 4′-Hydroxy-3′-methoxyacetophenone)
3. Esaxerenone (7α-[(2R,4R,5S,7S)-7-(2-Carboxyethyl)-2,3-dihydroxy-5,6-dimethyl-4-(2-oxo-1-pyrrolidinyl)oxytetrahydro-2H-pyran-4-yl]-5β,6β-dihydrospiro[naphthalene-1(2H),2′-pyrrolizine]-3′,5′-dione)
4. Dexamethasone (9α-Fluoro-11β,17α,21-trihydroxy-16α-methylpregna-1,4-diene-3,20-dione)
5. Sparsentan (N-(4-((5-(4-(2-Carboxyethyl)-1-piperazinyl)-2-methylphenyl)sulfonyl)-2,5-dimethylphenyl)-4-(2-methylphenyl)-1-piperazinecarboxamide)
6. Telmisartan (4′-[[4-Methyl-6-(1-methylbenzimidazol-2-yl)-2-propylbenzimidazol-1-yl]methyl]biphenyl-2-carboxylic acid)
7. Tolvaptan (N-[4-(7-Chloro-2,3,4,5-tetrahydro-5-hydroxy-1H-1-benzazepin-1-yl)-3-methylbenzoyl]benzene sulfonamide)
8. Conclusions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Hall, J.E.; Coleman, T.G.; Guyton, A.C. The renin-angiotensin system. Normal physiology and changes in older hypertensives. J. Am. Geriatr. Soc. 1989, 37, 801–813. [Google Scholar] [CrossRef] [PubMed]
- Ferrario, C.M. Role of angiotensin II in cardiovascular disease therapeutic implications of more than a century of research. J. Renin Angiotensin Aldosterone Syst. 2006, 7, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, R.M.; Satou, R.; Zhuo, J.L.; Nishiyama, A. The Renin-Angiotensin-Aldosterone System in Metabolic Diseases and Other Pathologies. Int. J. Mol. Sci. 2023, 24, 7413. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, S.; Mehta, S.; Khwaja, A.; Cleland, J.G.F.; Ives, N.; Brettell, E.; Chadburn, M.; Cockwell, P. Renin–Angiotensin System Inhibition in Advanced Chronic Kidney Disease. N. Engl. J. Med. 2022, 387, 2021–2032. [Google Scholar] [CrossRef] [PubMed]
- Miura, S.-I. The renin-angiotensin-aldosterone system: A new look at an old system. Hypertens. Res. 2023, 46, 932–933. [Google Scholar] [CrossRef] [PubMed]
- Ferrario, C.M.; Mullick, A.E. Renin angiotensin aldosterone inhibition in the treatment of cardiovascular disease. Pharmacol. Res. 2017, 125 Pt A, 57–71. [Google Scholar] [CrossRef]
- Chuang, T.H.; Cho, H.Y.; Wu, S.N. Effective Accentuation of Voltage-Gated Sodium Current Caused by Apocynin (4′-Hydroxy-3′-methoxyacetophenone), a Known NADPH-Oxidase Inhibitor. Biomedicines 2021, 9, 1146. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.T.; Wu, S.N. Characterization of Direct Perturbations on Voltage-Gated Sodium Current by Esaxerenone, a Nonsteroidal Mineralocorticoid Receptor Blocker. Biomedicines 2021, 9, 549. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.-H.; So, E.C.; Liu, Y.-C.; Wu, S.-N. Glucocorticoids stimulate the activity of large-conductance Ca2+-activated K+ channels in pituitary GH3 and AtT-20 cells via a non-genomic mechanism. Steroids 2006, 71, 129–140. [Google Scholar] [CrossRef]
- Chuang, T.H.; Cho, H.Y.; Wu, S.N. The Evidence for Sparsentan-Mediated Inhibition of INa and IK(erg): Possibly Unlinked to Its Antagonism of Angiotensin II or Endothelin Type a Receptor. Biomedicines 2021, 10, 86. [Google Scholar] [CrossRef]
- Kim, H.K.; Youm, J.B.; Lee, S.R.; Lim, S.E.; Lee, S.Y.; Ko, T.H.; Long le, T.; Nilius, B.; Won du, N.; Noh, J.H.; et al. The angiotensin receptor blocker and PPAR-γ agonist, telmisartan, delays inactivation of voltage-gated sodium channel in rat heart: Novel mechanism of drug action. Pflug. Arch. 2012, 464, 631–643. [Google Scholar] [CrossRef]
- Chang, T.T.; Yang, C.J.; Lee, Y.C.; Wu, S.N. Stimulatory Action of Telmisartan, an Antagonist of Angiotensin II Receptor, on Voltage-Gated Na+ Current: Experimental and Theoretical Studies. Chin. J. Physiol. 2018, 61, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.T.; Wu, S.N. Activation of voltage-gated sodium current and inhibition of erg-mediated potassium current caused by telmisartan, an antagonist of angiotensin II type-1 receptor, in HL-1 atrial cardiomyocytes. Clin. Exp. Pharmacol. Physiol. 2018, 45, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.C.; Wu, S.N.; Huang, C.W. Telmisartan, an Antagonist of Angiotensin II Receptors, Accentuates Voltage-Gated Na+ Currents and Hippocampal Neuronal Excitability. Front. Neurosci. 2020, 14, 902. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.-L.; Chang, W.-T.; Chan, C.-H.; Wu, S.-N. Evidence for Effective Multiple K+-Current Inhibitions by Tolvaptan, a Non-peptide Antagonist of Vasopressin V2 Receptor. Front. Pharmacol. 2019, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Zang, D.W.; Shan, W.; Guo, A.C.; Wu, J.P.; Wang, Y.J.; Wang, Q. Synthesis and Evaluations of Novel Apocynin Derivatives as Anti-Glioma Agents. Front. Pharmacol. 2019, 10, 951. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.D.; Yu, S.; Qi, Y.; Qu, T.F.; He, L.; Wei, W.; Liu, K.; Gong, S.S. NADPH oxidase inhibitor apocynin decreases mitochondrial dysfunction and apoptosis in the ventral cochlear nucleus of D-galactose-induced aging model in rats. Neurochem. Int. 2019, 124, 31–40. [Google Scholar] [CrossRef]
- Ilatovskaya, D.V.; Pavlov, T.S.; Levchenko, V.; Staruschenko, A. ROS production as a common mechanism of ENaC regulation by EGF, insulin, and IGF-1. Am. J. Physiol. Cell Physiol. 2013, 304, C102–C111. [Google Scholar] [CrossRef]
- Downs, C.A.; Johnson, N.M.; Coca, C.; Helms, M.N. Angiotensin II regulates δ-ENaC in human umbilical vein endothelial cells. Microvasc. Res. 2018, 116, 26–33. [Google Scholar] [CrossRef]
- Liu, F.; Fan, L.M.; Michael, N.; Li, J.M. In vivo and in silico characterization of apocynin in reducing organ oxidative stress: A pharmacokinetic and pharmacodynamic study. Pharmacol. Res. Perspect. 2020, 8, e00635. [Google Scholar] [CrossRef]
- Duggan, S. Esaxerenone: First Global Approval. Drugs 2019, 79, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Itoh, H.; Rakugi, H.; Okuda, Y.; Yoshimura, M.; Yamakawa, S. Double-Blind Randomized Phase 3 Study Comparing Esaxerenone (CS-3150) and Eplerenone in Patients With Essential Hypertension (ESAX-HTN Study). Hypertension 2020, 75, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Sawano, T.; Sen, A.; Hossain, A.; Jahan, N.; Kobara, H.; Masaki, T.; Kosaka, S.; Kitada, K.; Nakano, D.; et al. Cardioprotective Effects of a Nonsteroidal Mineralocorticoid Receptor Blocker, Esaxerenone, in Dahl Salt-Sensitive Hypertensive Rats. Int. J. Mol. Sci. 2021, 22, 2069. [Google Scholar] [CrossRef] [PubMed]
- Wan, N.; Rahman, A.; Nishiyama, A. Esaxerenone, a novel nonsteroidal mineralocorticoid receptor blocker (MRB) in hypertension and chronic kidney disease. J. Hum. Hypertens. 2021, 35, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Janković, S.M.; Janković, S.V. Clinical Pharmacokinetics and Pharmacodynamics of Esaxerenone, a Novel Mineralocorticoid Receptor Antagonist: A Review. Eur. J. Drug Metab. Pharmacokinet. 2022, 47, 291–308. [Google Scholar] [CrossRef] [PubMed]
- Ojha, U.; Ruddaraju, S.; Sabapathy, N.; Ravindran, V.; Worapongsatitaya, P.; Haq, J.; Mohammed, R.; Patel, V. Current and Emerging Classes of Pharmacological Agents for the Management of Hypertension. Am. J. Cardiovasc. Drugs 2022, 22, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.N.; Wu, Y.H.; Chen, B.S.; Lo, Y.C.; Liu, Y.C. Underlying mechanism of actions of tefluthrin, a pyrethroid insecticide, on voltage-gated ion currents and on action currents in pituitary tumor (GH3) cells and GnRH-secreting (GT1-7) neurons. Toxicology 2009, 258, 70–77. [Google Scholar] [CrossRef] [PubMed]
- So, E.C.; Wu, S.N.; Lo, Y.C.; Su, K. Differential regulation of tefluthrin and telmisartan on the gating charges of INa activation and inactivation as well as on resurgent and persistent INa in a pituitary cell line (GH3). Toxicol. Lett. 2018, 285, 104–112. [Google Scholar] [CrossRef]
- Kato, M.; Furuie, H.; Shimizu, T.; Miyazaki, A.; Kobayashi, F.; Ishizuka, H. Single- and multiple-dose escalation study to assess pharmacokinetics, pharmacodynamics and safety of oral esaxerenone in healthy Japanese subjects. Br. J. Clin. Pharmacol. 2018, 84, 1821–1829. [Google Scholar] [CrossRef]
- Wu, S.N. Large-conductance Ca2+- activated K+ channels:physiological role and pharmacology. Curr. Med. Chem. 2003, 10, 649–661. [Google Scholar] [CrossRef]
- Zang, K.; Zhang, Y.; Hu, J.; Wang, Y. The Large Conductance Calcium- and Voltage-activated Potassium Channel (BK) and Epilepsy. CNS Neurol. Disord. Drug Targets 2018, 17, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Orfali, R.; Albanyan, N. Ca(2+)-Sensitive Potassium Channels. Molecules 2023, 28, 885. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Duncan, R.R.; Hammond, M.S.; Coghill, L.S.; Wen, H.; Rusinova, R.; Clark, A.G.; Levitan, I.B.; Shipston, M.J. Alternative splicing switches potassium channel sensitivity to protein phosphorylation. J. Biol. Chem. 2001, 276, 7717–7720. [Google Scholar] [CrossRef] [PubMed]
- Hyde, G.N.; Seale, A.P.; Grau, E.G.; Borski, R.J. Cortisol rapidly suppresses intracellular calcium and voltage-gated calcium channel activity in prolactin cells of the tilapia (Oreochromis mossambicus). Am. J. Physiol. Endocrinol. Metab. 2004, 286, E626–E633. [Google Scholar] [CrossRef] [PubMed]
- Shipston, M.J.; Duncan, R.R.; Clark, A.G.; Antoni, F.A.; Tian, L. Molecular components of large conductance calcium-activated potassium (BK) channels in mouse pituitary corticotropes. Mol. Endocrinol. 1999, 13, 1728–1737. [Google Scholar] [CrossRef] [PubMed]
- Armanini, D.; Zennaro, C.M.; Martella, L.; Pratesi, C.; Scali, M.; Zampollo, V. Regulation of aldosterone receptors in hypertension. Steroids 1993, 58, 611–613. [Google Scholar] [CrossRef] [PubMed]
- Kazama, I.; Shoji, M. Targeting colonic BK channels: A novel therapeutic strategy against hyperkalemia in chronic kidney disease. Nefrología, 2019; in press. [Google Scholar] [CrossRef]
- Pofi, R.; Caratti, G.; Ray, D.W.; Tomlinson, J.W. Treating the side effects of exogenous glucocorticoids; Can we separate the good from the bad? Endocr. Rev. 2023; online ahead of print. [Google Scholar] [CrossRef]
- Loechner, K.J.; Knox, R.J.; McLaughlin, J.T.; Dunlap, K. Dexamethasone-mediated inhibition of calcium transients and ACTH release in a pituitary cell line (AtT-20). Steroids 1999, 64, 404–412. [Google Scholar] [CrossRef]
- Compagnone, N.A.; Mellon, S.H. Neurosteroids: Biosynthesis and Function of These Novel Neuromodulators. Front. Neuroendocrinol. 2000, 21, 1–56. [Google Scholar] [CrossRef]
- Gulyaeva, N.V. Glucocorticoids Orchestrate Adult Hippocampal Plasticity: Growth Points and Translational Aspects. Biochemistry 2023, 88, 565–589. [Google Scholar] [CrossRef]
- Lisakovska, O.; Labudzynskyi, D.; Khomenko, A.; Isaev, D.; Savotchenko, A.; Kasatkina, L.; Savosko, S.; Veliky, M.; Shymanskyi, I. Brain vitamin D(3)-auto/paracrine system in relation to structural, neurophysiological, and behavioral disturbances associated with glucocorticoid-induced neurotoxicity. Front. Cell. Neurosci. 2023, 17, 1133400. [Google Scholar] [CrossRef]
- Baylis, E.M.; Williams, I.A.; English, J.; Marks, V.; Chakraborty, J. High dose intravenous methylprednisolone “pulse” therapy in patients with rheumatoid disease. Plasma methylprednisolone levels and adrenal function. Eur. J. Clin. Pharmacol. 1982, 21, 385–388. [Google Scholar] [CrossRef]
- Morphy, R.; Rankovic, Z. Designed Multiple Ligands. An Emerging Drug Discovery Paradigm. J. Med. Chem. 2005, 48, 6523–6543. [Google Scholar] [CrossRef] [PubMed]
- Komers, R.; Gipson, D.S.; Nelson, P.; Adler, S.; Srivastava, T.; Derebail, V.K.; Meyers, K.E.; Pergola, P.; MacNally, M.E.; Hunt, J.L.; et al. Efficacy and Safety of Sparsentan Compared With Irbesartan in Patients With Primary Focal Segmental Glomerulosclerosis: Randomized, Controlled Trial Design (DUET). Kidney Int. Rep. 2017, 2, 654–664. [Google Scholar] [CrossRef]
- Syed, Y.Y. Sparsentan: First Approval. Drugs 2023, 83, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Heerspink, H.J.L.; Radhakrishnan, J.; Alpers, C.E.; Barratt, J.; Bieler, S.; Diva, U.; Inrig, J.; Komers, R.; Mercer, A.; Noronha, I.L.; et al. Sparsentan in patients with IgA nephropathy: A prespecified interim analysis from a randomised, double-blind, active-controlled clinical trial. Lancet 2023, 401, 1584–1594. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xue, S.; Hou, J.; Chen, G.; Xu, Z.G. Endothelin receptor antagonists for the treatment of diabetic nephropathy: A meta-analysis and systematic review. World J. Diabetes 2020, 11, 553–566. [Google Scholar] [CrossRef] [PubMed]
- Vega, A.V.; Espinosa, J.L.; López-Domínguez, A.M.; López-Santiago, L.F.; Navarrete, A.; Cota, G. L-type calcium channel activation up-regulates the mRNAs for two different sodium channel alpha subunits (Nav1.2 and Nav1.3) in rat pituitary GH3 cells. Brain Res. Mol. Brain Res. 2003, 116, 115–125. [Google Scholar] [CrossRef]
- Weber, M.A. Telmisartan in high-risk cardiovascular patients. Am. J. Cardiol. 2010, 105, 36a–43a. [Google Scholar] [CrossRef]
- Maejima, Y.; Okada, H.; Haraguchi, G.; Onai, Y.; Kosuge, H.; Suzuki, J.; Isobe, M. Telmisartan, a unique ARB, improves left ventricular remodeling of infarcted heart by activating PPAR gamma. Lab. Investig. 2011, 91, 932–944. [Google Scholar] [CrossRef]
- Chang, W.-T.; Cheng, J.-T.; Chen, Z.-C. Telmisartan improves cardiac fibrosis in diabetes through peroxisome proliferator activated receptor δ (PPARδ): From bedside to bench. Cardiovasc. Diabetol. 2016, 15, 113. [Google Scholar] [CrossRef]
- Caballero, R.; Delpón, E.; Valenzuela, C.; Longobardo, M.; Tamargo, J. Losartan and its metabolite E3174 modify cardiac delayed rectifier K(+) currents. Circulation 2000, 101, 1199–1205. [Google Scholar] [CrossRef] [PubMed]
- Stangier, J.; Schmid, J.; Türck, D.; Switek, H.; Verhagen, A.; Peeters, P.A.M.; Marle, S.P.; Tamminga, W.J.; Sollie, F.A.E.; Jonkman, J.H.G. Absorption, Metabolism, and Excretion of Intravenously and Orally Administered [14C]Telmisartan in Healthy Volunteers. J. Clin. Pharmacol. 2000, 40, 1312–1322. [Google Scholar] [CrossRef] [PubMed]
- Cosarderelioglu, C.; Nidadavolu, L.S.; George, C.J.; Oh, E.S.; Bennett, D.A.; Walston, J.D.; Abadir, P.M. Brain Renin-Angiotensin System at the Intersect of Physical and Cognitive Frailty. Front. Neurosci. 2020, 14, 586314. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.Z.-Y.; Loh, S.-H.; Cheng, T.-H.; Lu, H.-H.; Lin, C.-I. Antiarrhythmic effects of (−)-epicatechin-3-gallate, a novel sodium channel agonist in cultured neonatal rat ventricular myocytes. Biochem. Pharmacol. 2013, 85, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Mechaly, I.; Scamps, F.; Chabbert, C.; Sans, A.; Valmier, J. Molecular diversity of voltage-gated sodium channel alpha subunits expressed in neuronal and non-neuronal excitable cells. Neuroscience 2005, 130, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Al-kuraishy, H.M.; Al-Gareeb, A.I.; Alkhuriji, A.F.; Al-Megrin, W.A.I.; Elekhnawy, E.; Negm, W.A.; De Waard, M.; Batiha, G.E.-S. Investigation of the impact of rosuvastatin and telmisartan in doxorubicin-induced acute cardiotoxicity. Biomed. Pharmacother. 2022, 154, 113673. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, M.; Fabrès, V.; Dumont, R.; Chetboul, V.; Chahory, S.; Saponaro, V.; Trehiou, E.; Poissonnier, C.; Passavin, P.; Jondeau, C.; et al. Prospective evaluation of a telmisartan suppression test as a diagnostic tool for primary hyperaldosteronism in cats. J. Vet. Intern. Med. 2023, 37, 1348–1357. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Hou, Y.; Tu, W.; Campbell, N.; Pieper, A.A.; Leverenz, J.B.; Gao, S.; Cummings, J.; Cheng, F. Population-based discovery and Mendelian randomization analysis identify telmisartan as a candidate medicine for Alzheimer’s disease in African Americans. Alzheimer’s Dement. 2023, 19, 1876–1887. [Google Scholar] [CrossRef]
- Takimura, H.; Hada, T.; Kawano, M.; Yabe, T.; Takimura, Y.; Nishio, S.; Nakano, M.; Tsukahara, R.; Muramatsu, T. A novel validated method for predicting the risk of re-hospitalization for worsening heart failure and the effectiveness of the diuretic upgrading therapy with tolvaptan. PLoS ONE 2018, 13, e0207481. [Google Scholar] [CrossRef]
- Huang, J.-H.; Chen, Y.-C.; Lu, Y.-Y.; Lin, Y.-K.; Chen, S.-A.; Chen, Y.-J. Arginine vasopressin modulates electrical activity and calcium homeostasis in pulmonary vein cardiomyocytes. J. Biomed. Sci. 2019, 26, 71. [Google Scholar] [CrossRef]
- Vidic, A.; Shuster, J.E.; Goff, Z.D.; Godishala, A.; Joseph, S.M.; Chibnall, J.T.; Hauptman, P.J. Vasopressin antagonism for decompensated right-sided heart failure. Int. J. Cardiol. 2019, 274, 245–247. [Google Scholar] [CrossRef] [PubMed]
- Alukal, J.J.; John, S.; Thuluvath, P.J. Hyponatremia in Cirrhosis: An Update. Am. J. Gastroenterol. 2020, 115, 1775–1785. [Google Scholar] [CrossRef] [PubMed]
- Berardi, R.; Antonuzzo, A.; Blasi, L.; Buosi, R.; Lorusso, V.; Migliorino, M.R.; Montesarchio, V.; Zilembo, N.; Sabbatini, R.; Peri, A. Practical issues for the management of hyponatremia in oncology. Endocrine 2018, 61, 158–164. [Google Scholar] [CrossRef]
- Casis, O.; Gallego, M.; Sánchez-Chapula, J.A. Imipramine, mianserine and maprotiline block delayed rectifier potassium current in ventricular myocytes. Pharmacol. Res. 2002, 45, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Quintero, J.L.; Arenas, M.I.; García, D.E. The antidepressant imipramine inhibits M current by activating a phosphatidylinositol 4,5-bisphosphate (PIP2)-dependent pathway in rat sympathetic neurones. Br. J. Pharmacol. 2005, 145, 837–843. [Google Scholar] [CrossRef] [PubMed]
- Iijima, M.; Yoshimizu, T.; Shimazaki, T.; Tokugawa, K.; Fukumoto, K.; Kurosu, S.; Kuwada, T.; Sekiguchi, Y.; Chaki, S. Antidepressant and anxiolytic profiles of newly synthesized arginine vasopressin V1B receptor antagonists: TASP0233278 and TASP0390325. Br. J. Pharmacol. 2014, 171, 3511–3525. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.-S.; Lo, Y.-C.; Peng, H.; Hsu, T.-I.; Wu, S.-N. Effects of Ranolazine, a Novel Anti-anginal Drug, on Ion Currents and Membrane Potential in Pituitary Tumor GH3 Cells and NG108-15 Neuronal Cells. J. Pharmacol. Sci. 2009, 110, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Ayyasamy, L.; Bagepally, B.S. Cost-utility of Ranolazine for Chronic Stable Angina Pectoris: Systematic Review and Meta-analysis. Clin. Ther. 2023, 45, 458–465. [Google Scholar] [CrossRef]
- Emathinger, J.M.; Nelson, J.W.; Gurley, S.B. Advances in use of mouse models to study the renin-angiotensin system. Mol. Cell. Endocrinol. 2021, 529, 111255. [Google Scholar] [CrossRef]
- Li, A.; Shi, W.; Wang, J.; Wang, X.; Zhang, Y.; Lei, Z.; Jiao, X.Y. The gene knockout of angiotensin II type 1a receptor improves high-fat diet-induced obesity in rat via promoting adipose lipolysis. PLoS ONE 2022, 17, e0267331. [Google Scholar] [CrossRef]


{kind=link}
| Drug or Compound | Chemical Structure |
|---|---|
| Apocynin |  |
| Esaxerenone |  |
| Dexamethasone | 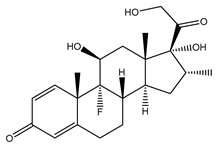 |
| Sparsentan |  |
| Telmisartan | 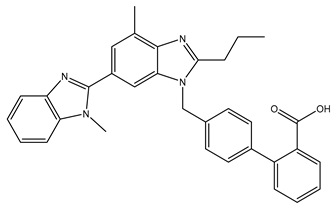 |
| Tolvaptan |  |
| Drug or Compound | Abbreviation | Off-Target Effect on Ionic Currents | References |
|---|---|---|---|
| Apocynin | aPO | ↑INa | [7] |
| Esaxerenone | ESAX | ↓INa | [8] |
| Dexamethasone | DEX | ↑IK(Ca) | [9] |
| Sparsentan | RE-021 * | ↓INa, ↓IK(erg), ↓IK(DR) | [10] |
| Telmisartan | TEL | ↑INa, ↓IK(erg) | [11,12,13,14] |
| Tolvaptan | TLV | ↓IK(DR), ↓IK(M) | [15] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, T.-L.; Wu, S.-N. Investigating the Impact of Selective Modulators on the Renin–Angiotensin–Aldosterone System: Unraveling Their Off-Target Perturbations of Transmembrane Ionic Currents. Int. J. Mol. Sci. 2023, 24, 14007. https://doi.org/10.3390/ijms241814007
Lu T-L, Wu S-N. Investigating the Impact of Selective Modulators on the Renin–Angiotensin–Aldosterone System: Unraveling Their Off-Target Perturbations of Transmembrane Ionic Currents. International Journal of Molecular Sciences. 2023; 24(18):14007. https://doi.org/10.3390/ijms241814007
Chicago/Turabian StyleLu, Te-Ling, and Sheng-Nan Wu. 2023. "Investigating the Impact of Selective Modulators on the Renin–Angiotensin–Aldosterone System: Unraveling Their Off-Target Perturbations of Transmembrane Ionic Currents" International Journal of Molecular Sciences 24, no. 18: 14007. https://doi.org/10.3390/ijms241814007