
Fibrin-Rhamnogalacturonan I Composite Gel for Therapeutic Enzyme Delivery to Intestinal Tumors
 , , and
, , and
Abstract
:1. Introduction
2. Results
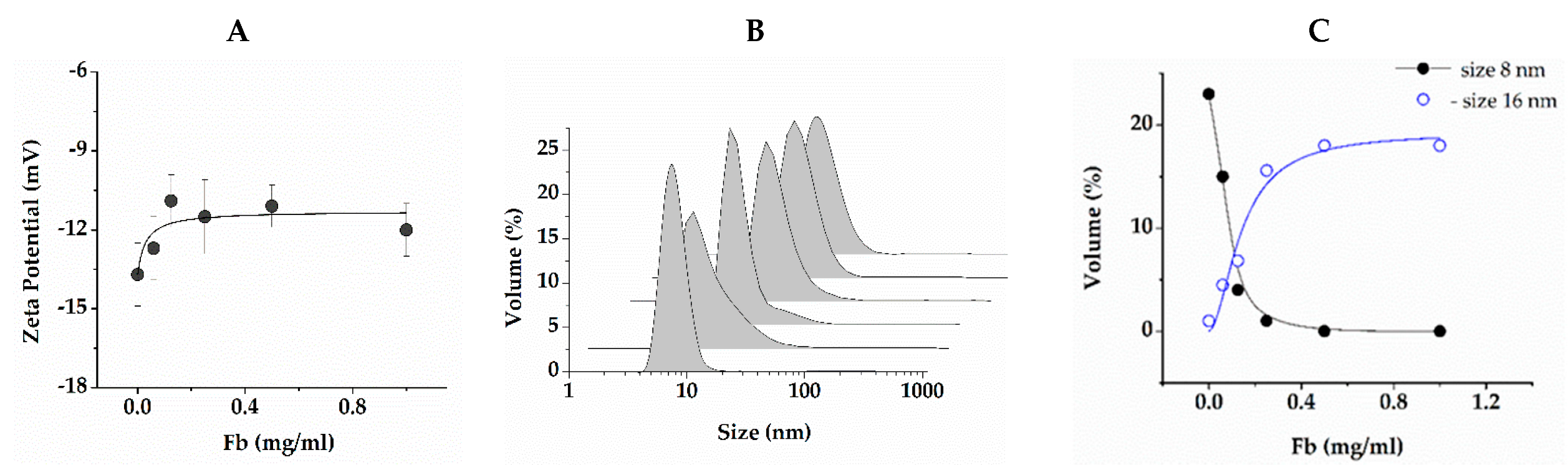
2.1. Morphology of Composite Gels
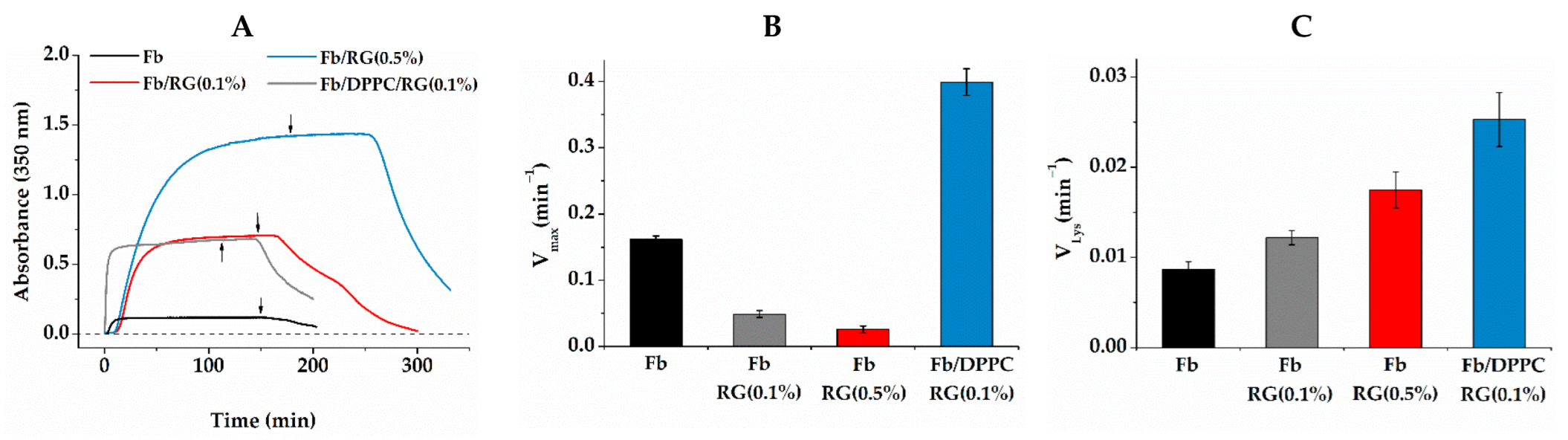
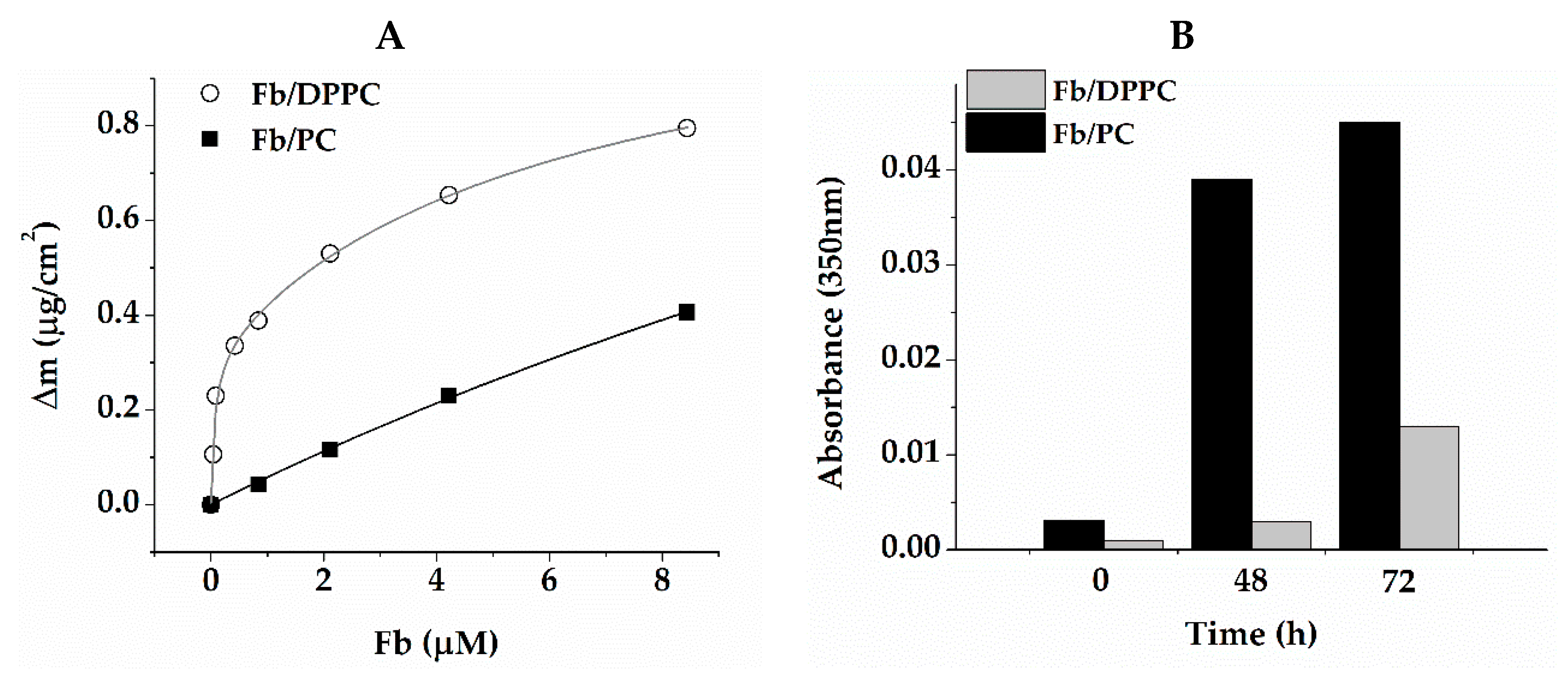
2.2. Fibrin Formation and Fibrinolysis in the Presence of Liposomes and RG
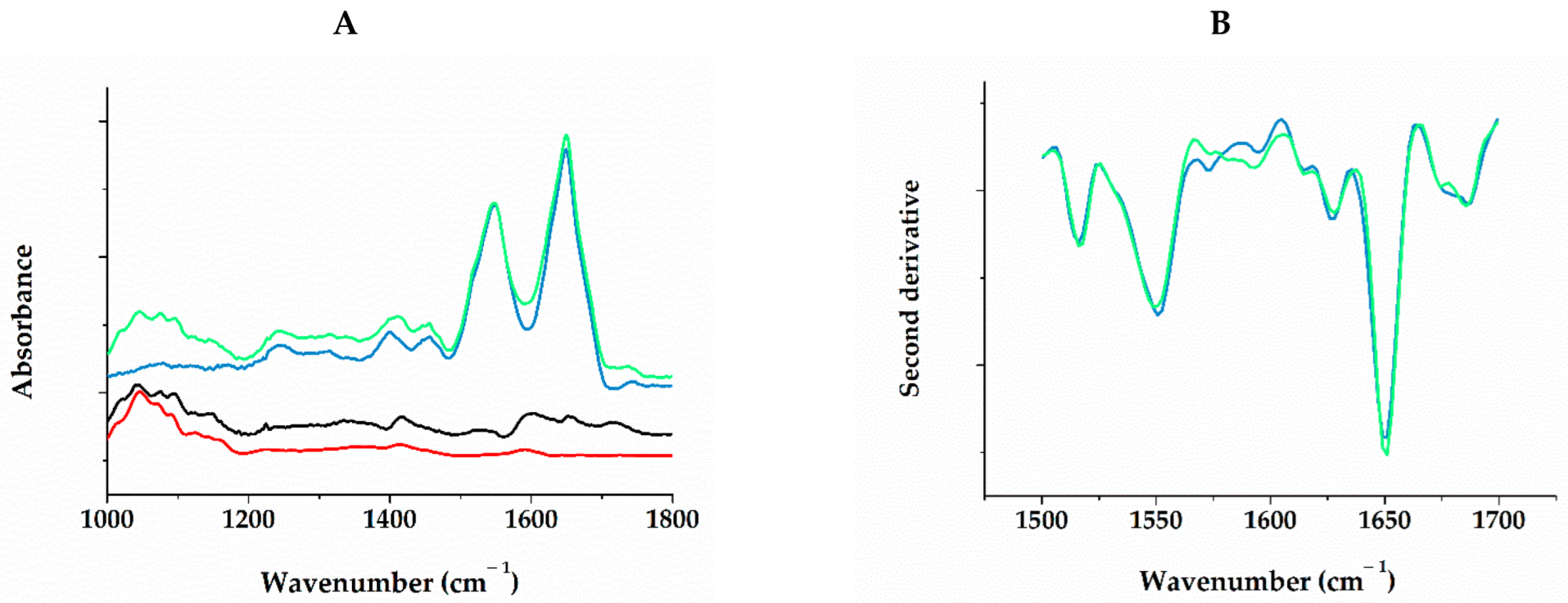
2.3. Interaction of Gel Components
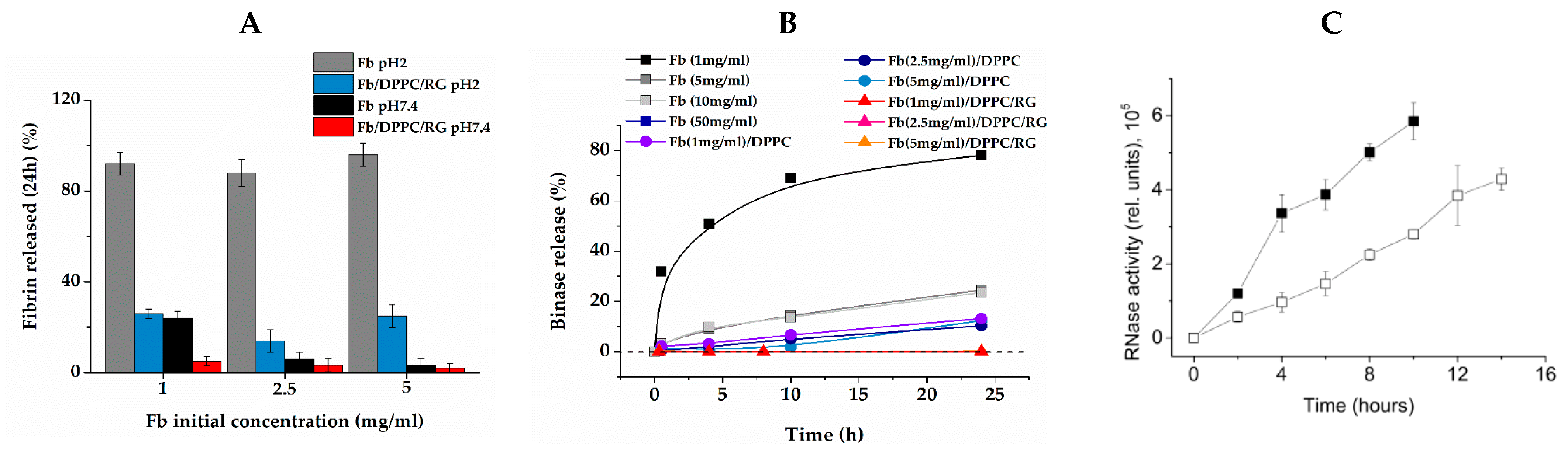
2.4. Resistance of Composite Gel to pH
2.5. Release of Binase from Composite Gel
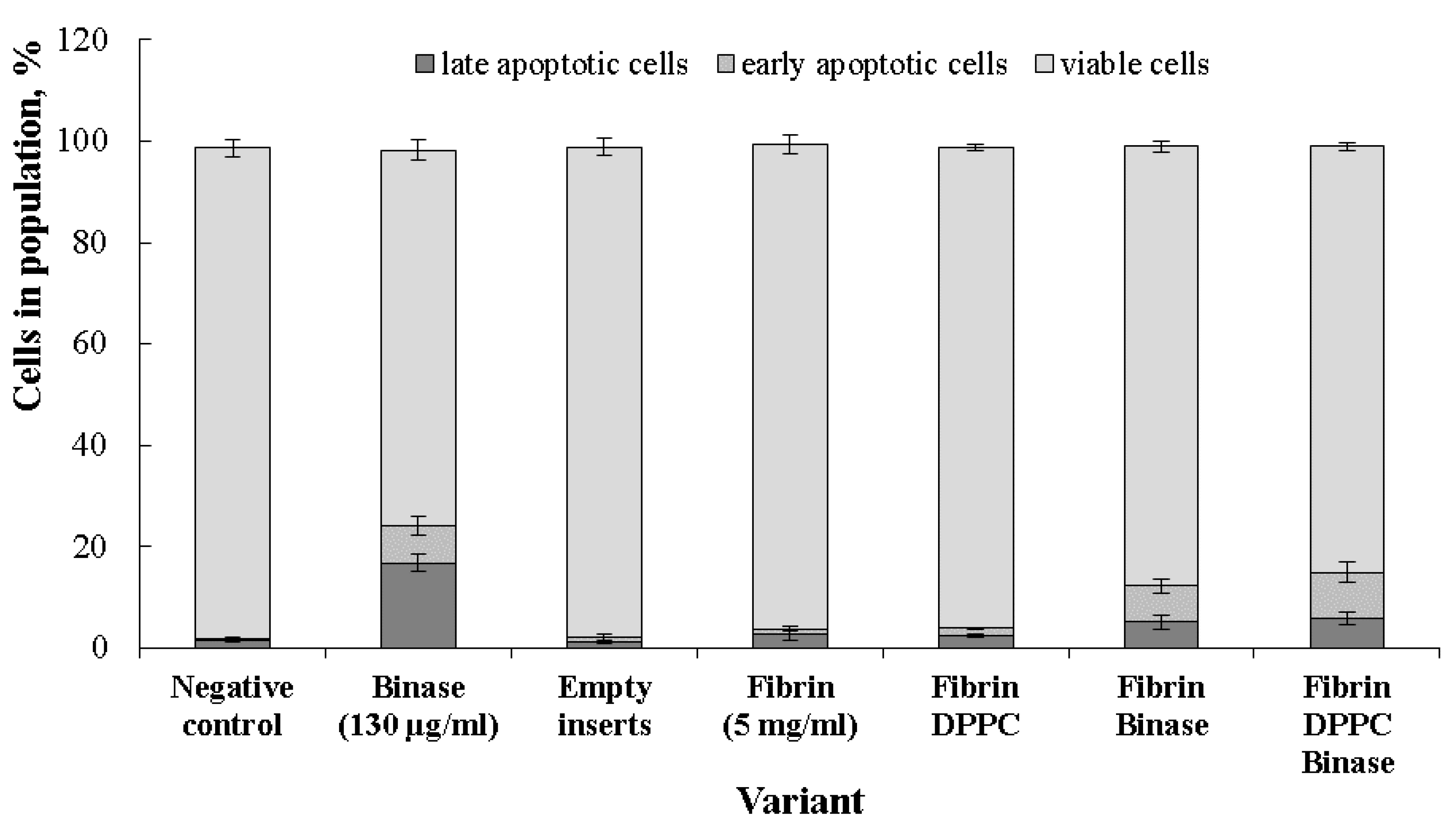
2.6. Cytotoxic Effect of Binase towards Cancer Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Enzyme
4.3. Preparation of Solutions
4.4. Preparation of Liposomes
4.5. Microgravimetry
4.6. Turbidimetry
4.7. Analysis of Gels and Their Components Interaction
- (i)
- The resistance of gels to pH was studied by placing 1 mL of the formed gels without binase into a test tube with 10 mL of buffer 1 at pH 7.4 and pH 2 and measuring the optical density of supernatant at 280 nm after the incubation time.
- (ii)
- The interaction of lipid with binase was determined turbidimetrically measuring the half-transition temperature of a DPPC suspension with or without encapsulated binase. For these, an optical density at 350 nm was recorded upon continuous temperature scanning from 35 °C to 45 °C at a rate of 0.1 °C/min. The accuracy of temperature setting was not worse than 0.05 °C with a Pelkin Elmer Peltier thermostat. The dependence of optical density on temperature was approximated by the Vant-Hoff two-state model equation from the set of functions of the OriginPro 2015 package, obtaining the half-transition temperature.
- (iii)
- The release of binase from the composite gel was determined by subtracting the optical density in the supernatant at 280 nm in the control from the optical density in the experiment. To do this, 1 mL of the formed gel of various compositions with binase (experiment) and without binase (control) were placed in test tubes with 10 mL of buffer 1 at pH 7.4 and incubated for a specified time.
- (iv)
- Adsorption of Fb on lipid bilayers was measured using the microgravimetry technique.
4.8. Dynamic Light Scattering
4.9. IR Spectroscopy
4.10. Scanning Electron Microscopy
4.11. Cytotoxicity towards Cancer Cells
4.12. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- ICBP-SURVMARK-2. Available online: https://gco.iarc.fr/survival/survmark/visualizations/viz7/?mode=%22bar%22&sub_mode=%22all%22&groupby=%22country%22&period=%225%22&cancer=%22COLO%22&country=%22Australia%22&gender=%220%22&stage=%22SEER%22&age_group=%2215-99%22&show_ci=%22%22. (accessed on 20 December 2022).
- Müller, C.; Ulyanova, V.; Ilinskaya, O.; Pleschka, S.; Shah Mahmud, R. A Novel Antiviral Strategy against MERS-CoV and HCoV-229E Using Binase to Target Viral Genome Replication. Bionanoscience 2017, 7, 294–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah Mahmud, R.; Mostafa, A.; Müller, C.; Kanrai, P.; Ulyanova, V.; Sokurenko, Y.; Dzieciolowski, J.; Kuznetsova, I.; Ilinskaya, O.; Pleschka, S. Bacterial Ribonuclease Binase Exerts an Intra-Cellular Anti-Viral Mode of Action Targeting Viral RNAs in Influenza a Virus-Infected MDCK-II Cells. Virol. J. 2018, 15, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah Mahmud, R.; Efimova, M.A.; Ulyanova, V.; Ravilov, R.K.; Shuralev, E.A.; Kolpakov, A.; Ilinskaya, O. Bacillus Pumilus Ribonuclease Rescues Mice Infected by Double-Stranded RNA-Containing Reovirus Serotype 1. Virus Res. 2020, 286, 198086. [Google Scholar] [CrossRef]
- Makarov, A.A.; Ilinskaya, O.N. Cytotoxic Ribonucleases: Molecular Weapons and Their Targets. FEBS Lett. 2003, 540, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Ilinskaya, O.; Decker, K.; Koschinski, A.; Dreyer, F.; Repp, H. Bacillus Intermedius Ribonuclease as Inhibitor of Cell Proliferation and Membrane Current. Toxicology 2001, 156, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Ilinskaya, O.N.; Singh, I.; Dudkina, E.; Ulyanova, V.; Kayumov, A.; Barreto, G. Direct Inhibition of Oncogenic KRAS by Bacillus Pumilus Ribonuclease (Binase). Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2016, 1863, 1559–1567. [Google Scholar] [CrossRef]
- Mironova, N.; Petrushanko, I.; Patutina, O.; Sen’kova, A.V.; Simonenko, O.; Mitkevich, V.; Markov, O.; Zenkova, M.; Makarov, A.A. Ribonuclease Binase Inhibits Primary Tumor Growth and Metastases via Apoptosis Induction in Tumor Cells. Cell Cycle 2013, 12, 2120–2131. [Google Scholar] [CrossRef]
- Bogdanova, L.R.; Zelenikhin, P.V.; Makarova, A.O.; Zueva, O.S.; Salnikov, V.V.; Zuev, Y.F.; Ilinskaya, O.N. Alginate-Based Hydrogel as Delivery System for Therapeutic Bacterial RNase. Polymers 2022, 14, 2461. [Google Scholar] [CrossRef]
- Sinicropi, M.S.; Amantea, D.; Caruso, A.; Saturnino, C. Chemical and Biological Properties of Toxic Metals and Use of Chelating Agents for the Pharmacological Treatment of Metal Poisoning. Arch. Toxicol. 2010, 84, 501–520. [Google Scholar] [CrossRef]
- Khotimchenko, M. Pectin Polymers for Colon-Targeted Antitumor Drug Delivery. Int. J. Biol. Macromol. 2020, 158, 1110–1124. [Google Scholar] [CrossRef]
- Alvarez-Lorenzo, C.; Blanco-Fernandez, B.; Puga, A.M.; Concheiro, A. Crosslinked Ionic Polysaccharides for Stimuli-Sensitive Drug Delivery. Adv. Drug Deliv. Rev. 2013, 65, 1148–1171. [Google Scholar] [CrossRef] [PubMed]
- Assifaoui, A.; Bouyer, F.; Chambin, O.; Cayot, P. Silica-Coated Calcium Pectinate Beads for Colonic Drug Delivery. Acta Biomater. 2013, 9, 6218–6225. [Google Scholar] [CrossRef] [PubMed]
- Tønnesen, H.H.; Karlsen, J. Alginate in Drug Delivery Systems. Drug Dev. Ind. Pharm. 2002, 28, 621–630. [Google Scholar] [CrossRef]
- Gubaidullin, A.T.; Makarova, A.O.; Derkach, S.R.; Voron’ko, N.G.; Kadyirov, A.I.; Ziganshina, S.A.; Salnikov, V.V.; Zueva, O.S.; Zuev, Y.F. Modulation of Molecular Structure and Mechanical Properties of κ-Carrageenan-Gelatin Hydrogel with Multi-Walled Carbon Nanotubes. Polymers 2022, 14, 2346. [Google Scholar] [CrossRef]
- Makarova, A.O.; Derkach, S.R.; Kadyirov, A.I.; Ziganshina, S.A.; Kazantseva, M.A.; Zueva, O.S.; Gubaidullin, A.T.; Zuev, Y.F. Supramolecular Structure and Mechanical Performance of κ-Carrageenan–Gelatin Gel. Polymers 2022, 14, 4347. [Google Scholar] [CrossRef]
- Faizullin, D.; Valiullina, Y.; Salnikov, V.; Zuev, Y. Direct Interaction of Fibrinogen with Lipid Microparticles Modulates Clotting Kinetics and Clot Structure. Nanomed. Nanotechnol. Biol. Med. 2020, 23, 102098. [Google Scholar] [CrossRef]
- Makshakova, O.N.; Zuev, Y.F. Interaction-Induced Structural Transformations in Polysaccharide and Protein-Polysaccharide Gels as Functional Basis for Novel Soft-Matter: A Case of Carrageenans. Gels 2022, 8, 287. [Google Scholar] [CrossRef]
- Zhang, W.; Jin, X.; Li, H.; Zhang, R.; Wu, C. Injectable and Body Temperature Sensitive Hydrogels Based on Chitosan and Hyaluronic Acid for PH Sensitive Drug Release. Carbohydr. Polym. 2018, 186, 82–90. [Google Scholar] [CrossRef]
- Makshakova, O.N.; Bogdanova, L.R.; Makarova, A.O.; Kusova, A.M.; Ermakova, E.A.; Kazantseva, M.A.; Zuev, Y.F. κ-Carrageenan Hydrogel as a Matrix for Therapeutic Enzyme Immobilization. Polymers 2022, 14, 4071. [Google Scholar] [CrossRef]
- Ahadian, S.; Sadeghian, R.B.; Salehi, S.; Ostrovidov, S.; Bae, H.; Ramalingam, M.; Khademhosseini, A. Bioconjugated Hydrogels for Tissue Engineering and Regenerative Medicine. Bioconjugate Chem. 2015, 26, 1984–2001. [Google Scholar] [CrossRef]
- Wang, Y.; Liang, Z.; Su, Z.; Zhang, K.; Ren, J.; Sun, R.; Wang, X. All-Biomass Fluorescent Hydrogels Based on Biomass Carbon Dots and Alginate/Nanocellulose for Biosensing. ACS Appl. Bio Mater. 2018, 1, 1398–1407. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Sun, P.; Li, P.; Xue, A.; Zhang, X.; Zhang, H.; Jin, X. A Magnetic Chitosan Hydrogel for Sustained and Prolonged Delivery of Bacillus Calmette–Guérin in the Treatment of Bladder Cancer. Biomaterials 2013, 34, 10258–10266. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, E.G.; Colquhoun, I.J.; Chau, H.K.; Hotchkiss, A.T.; Waldron, K.W.; Morris, V.J.; Belshaw, N.J. Rhamnogalacturonan I Containing Homogalacturonan Inhibits Colon Cancer Cell Proliferation by Decreasing ICAM1 Expression. Carbohydr. Polym. 2015, 132, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.; Grant, L.; Gidley, M.; Mikkelsen, D. Gut Fermentation of Dietary Fibres: Physico-Chemistry of Plant Cell Walls and Implications for Health. Int. J. Mol. Sci. 2017, 18, 2203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.; Zheng, J.; Mao, G.; Hu, W.; Ye, X.; Linhardt, R.J.; Chen, S. Rethinking the Impact of RG-I Mainly from Fruits and Vegetables on Dietary Health. Crit. Rev. Food Sci. Nutr. 2020, 60, 2938–2960. [Google Scholar] [CrossRef] [PubMed]
- Manderson, K.; Pinart, M.; Tuohy, K.M.; Grace, W.E.; Hotchkiss, A.T.; Widmer, W.; Yadhav, M.P.; Gibson, G.R.; Rastall, R.A. In Vitro Determination of Prebiotic Properties of Oligosaccharides Derived from an Orange Juice Manufacturing By-Product Stream. Appl. Environ. Microbiol. 2005, 71, 8383–8389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khodaei, N.; Fernandez, B.; Fliss, I.; Karboune, S. Digestibility and Prebiotic Properties of Potato Rhamnogalacturonan I Polysaccharide and Its Galactose-Rich Oligosaccharides/Oligomers. Carbohydr. Polym. 2016, 136, 1074–1084. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Zhi, Y.; Sun, L.; Peng, X.; Zhang, T.; Xue, H.; Tai, G.; Zhou, Y. The Inhibitory Effects of a Rhamnogalacturonan I (RG-I) Domain from Ginseng Pectin on Galectin-3 and Its Structure-Activity Relationship. J. Biol. Chem. 2013, 288, 33953–33965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khodaei, N.; Karboune, S.; Orsat, V. Microwave-Assisted Alkaline Extraction of Galactan-Rich Rhamnogalacturonan I from Potato Cell Wall by-Product. Food Chem. 2016, 190, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Dubnika, A.; Egle, K.; Skrinda-Melne, M.; Skadins, I.; Rajadas, J.; Salma, I. Development of Vancomycin Delivery Systems Based on Autologous 3D Platelet-Rich Fibrin Matrices for Bone Tissue Engineering. Biomedicines 2021, 9, 814. [Google Scholar] [CrossRef] [PubMed]
- Heher, P.; Mühleder, S.; Mittermayr, R.; Redl, H.; Slezak, P. Fibrin-Based Delivery Strategies for Acute and Chronic Wound Healing. Adv. Drug Deliv. Rev. 2018, 129, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-S.; Yang, M.-C.; Chung, T.-W. Liposomes/Chitosan Scaffold/Human Fibrin Gel Composite Systems for Delivering Hydrophilic Drugs—Release Behaviors of Tirofiban In Vitro. Drug Deliv. 2008, 15, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Bujoli, B.; Scimeca, J.-C.; Verron, E. Fibrin as a Multipurpose Physiological Platform for Bone Tissue Engineering and Targeted Delivery of Bioactive Compounds. Pharmaceutics 2019, 11, 556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, F.H.; Byg, I.; Damager, I.; Diaz, J.; Engelsen, S.B.; Ulvskov, P. Residue Specific Hydration of Primary Cell Wall Potato Pectin Identified by Solid-State 13 C Single-Pulse MAS and CP/MAS NMR Spectroscopy. Biomacromolecules 2011, 12, 1844–1850. [Google Scholar] [CrossRef]
- Makshakova, O.N.; Faizullin, D.A.; Mikshina, P.V.; Gorshkova, T.A.; Zuev, Y.F. Spatial Structures of Rhamnogalacturonan I in Gel and Colloidal Solution Identified by 1D and 2D-FTIR Spectroscopy. Carbohydr. Polym. 2018, 192, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Mikshina, P.V.; Idiyatullin, B.Z.; Petrova, A.A.; Shashkov, A.S.; Zuev, Y.F.; Gorshkova, T.A. Physicochemical Properties of Complex Rhamnogalacturonan I from Gelatinous Cell Walls of Flax Fibers. Carbohydr. Polym. 2015, 117, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Svagan, A.J.; Kusic, A.; De Gobba, C.; Larsen, F.H.; Sassene, P.; Zhou, Q.; van de Weert, M.; Mullertz, A.; Jørgensen, B.; Ulvskov, P. Rhamnogalacturonan-I Based Microcapsules for Targeted Drug Release. PLoS ONE 2016, 11, e0168050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weisel, J.W.; Nagaswami, C. Computer Modeling of Fibrin Polymerization Kinetics Correlated with Electron Microscope and Turbidity Observations: Clot Structure and Assembly Are Kinetically Controlled. Biophys. J. 1992, 63, 111–128. [Google Scholar] [CrossRef] [Green Version]
- Weisel, J.W.; Litvinov, R.I. Fibrin Formation, Structure and Properties. In Fibrous Proteins: Structures and Mechanisms; Parry, D.A.D., Squire, J.M., Eds.; Subcellular Biochemistry; Springer International Publishing: Cham, Switzerland, 2017; Volume 82, pp. 405–456. ISBN 978-3-319-49672-6. [Google Scholar]
- Zuev, Y.F.; Litvinov, R.I.; Sitnitsky, A.E.; Idiyatullin, B.Z.; Bakirova, D.R.; Galanakis, D.K.; Zhmurov, A.; Barsegov, V.; Weisel, J.W. Conformational Flexibility and Self-Association of Fibrinogen in Concentrated Solutions. J. Phys. Chem. B 2017, 121, 7833–7843. [Google Scholar] [CrossRef]
- Wedgwood, J.; Freemont, A.J.; Tirelli, N. Rheological and Turbidity Study of Fibrin Hydrogels. Macromol. Symp. 2013, 334, 117–125. [Google Scholar] [CrossRef]
- Jurkiewicz, P.; Cwiklik, L.; Jungwirth, P.; Hof, M. Lipid Hydration and Mobility: An Interplay between Fluorescence Solvent Relaxation Experiments and Molecular Dynamics Simulations. Biochimie 2012, 94, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Huang, X.; Nakamura, A.; Burchard, W.; Hallett, F.R. Molecular Characterisation of Soybean Polysaccharides: An Approach by Size Exclusion Chromatography, Dynamic and Static Light Scattering Methods. Carbohydr. Res. 2005, 340, 2637–2644. [Google Scholar] [CrossRef] [PubMed]
- Kavitake, D.; Techi, M.; Abid, U.K.; Kandasamy, S.; Devi, P.B.; Shetty, P.H. Effect of γ-Irradiation on Physico-Chemical and Antioxidant Properties of Galactan Exopolysaccharide from Weissella Confusa KR780676. J. Food Sci. Technol. 2019, 56, 1766–1774. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.-R.; Fu, Y.; Wu, D.-T.; Huang, T.-T.; Jiang, Q.; Zhao, L.; Zhang, Q.; Lin, D.-R.; Chen, H.; Qin, W. Ultrasonic-Assisted Extraction, Structural Characterization, Chain Conformation, and Biological Activities of a Pectic-Polysaccharide from Okra (Abelmoschus esculentus). Molecules 2020, 25, 1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamczyk, Z.; Bratek-Skicki, A.; Dąbrowska, P.; Nattich-Rak, M. Mechanisms of Fibrinogen Adsorption on Latex Particles Determined by Zeta Potential and AFM Measurements. Langmuir 2012, 28, 474–485. [Google Scholar] [CrossRef]
- Makshakova, O.N.; Faizullin, D.A.; Zuev, Y.F. Interplay between Secondary Structure and Ion Binding upon Thermoreversible Gelation of κ-Carrageenan. Carbohydr. Polym. 2020, 227, 115342. [Google Scholar] [CrossRef]
- Madni, A.; Sarfraz, M.; Rehman, M.; Ahmad, M.; Akhtar, N.; Ahmad, S.; Tahir, N.; Ijaz, S.; Al-Kassas, R.; Löbenberg, R. Liposomal Drug Delivery: A Versatile Platform for Challenging Clinical Applications. J. Pharm. Pharm. Sci. 2014, 17, 401. [Google Scholar] [CrossRef] [Green Version]
- Walde, P.; Ichikawa, S. Enzymes inside Lipid Vesicles: Preparation, Reactivity and Applications. Biomol. Eng. 2001, 18, 143–177. [Google Scholar] [CrossRef]
- Xu, X.; Costa, A.; Burgess, D.J. Protein Encapsulation in Unilamellar Liposomes: High Encapsulation Efficiency and A Novel Technique to Assess Lipid-Protein Interaction. Pharm. Res. 2012, 29, 1919–1931. [Google Scholar] [CrossRef]
- Koynova, R.; Caffrey, M. Phases and Phase Transitions of the Phosphatidylcholines. Biochim. Biophys. Acta (BBA)-Rev. Biomembr. 1998, 1376, 91–145. [Google Scholar] [CrossRef]
- Takegawa, Y.; Takao, T.; Sakaguchi, H.; Nakai, T.; Takeo, K.; Morita, Y.; Toyonaga, T.; Kodama, Y. The Importance of PH Adjustment for Preventing Fibrin Glue Dissolution in the Stomach: An in Vitro Study. Sci. Rep. 2022, 12, 6986. [Google Scholar] [CrossRef] [PubMed]
- Okumura, N.; Gorkun, O.V.; Lord, S.T. Severely impaired polymerization of recombinant fibrinogen. J. Biol. Chem. 1997, 272, 29596–29601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelenikhin, P.V.; Ead Mohamed, I.S.; Nadyrova, A.I.; Sirotkina, A.A.; Ulyanova, V.V.; Mironova, N.L.; Mitkevich, V.A.; Makarov, A.A.; Zenkova, M.A.; Ilinskaya, O.N. [Bacillus pumilus Ribonuclease Inhibits Migration of Human Duodenum Adenocarcinoma HuTu 80 Cells]. Mol. Biol. (Mosk) 2020, 54, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Shirshikov, F.V.; Cherepnev, G.V.; Ilinskaya, O.N.; Kalacheva, N.V. A Hydrophobic Segment of Some Cytotoxic Ribonucleases. Med. Hypotheses 2013, 81, 328–334. [Google Scholar] [CrossRef]
- Takahashi, K.; Moore, S. 13 Ribonuclease T1. In The Enzymes; Elsevier: Amsterdam, The Netherlands, 1982; Volume 15, pp. 435–468. ISBN 978-0-12-122715-9. [Google Scholar]
- Patelli, G.; Tosi, F.; Amatu, A.; Mauri, G.; Curaba, A.; Patanè, D.A.; Pani, A.; Scaglione, F.; Siena, S.; Sartore-Bianchi, A. Strategies to Tackle RAS-Mutated Metastatic Colorectal Cancer. ESMO Open 2021, 6, 100156. [Google Scholar] [CrossRef]
- Ilinskaya, O.N.; Kharitonova, M.A.; Doynikova, A.N.; Zelenikhin, P.V.; Gataullin, B.I. Analysis of RAS-Mutation and Microbiom of Patients with Colorectal Cancer. Int. J. Pharm. Res. 2018, 10, 307–313. [Google Scholar]
- Dudkina, E.; Ulyanova, V.; Shah Mahmud, R.; Khodzhaeva, V.; Dao, L.; Vershinina, V.; Kolpakov, A.; Ilinskaya, O. Three-step Procedure for Preparation of Pure Bacillus altitudinis Ribonuclease. FEBS Open Bio. 2016, 6, 24–32. [Google Scholar] [CrossRef]
- Yakovlev, G.I.; Moiseyev, G.P.; Struminskaya, N.K.; Borzykh, O.A.; Kipenskaya, L.V.; Znamenskaya, L.V.; Leschinskaya, I.B.; Chernokalskaya, E.B.; Hartley, R.W. Mutational Analysis of the Active Site of RNase of Bacillus intermedius (BINASE). FEBS Lett. 1994, 354, 305–306. [Google Scholar] [CrossRef] [Green Version]
- Ilinskaya, O. SOS-Inducing Ability of Native and Mutant Microbial Ribonucleases. Mutat. Res./Fundam. Mol. Mech. Mutagen. 1996, 354, 203–209. [Google Scholar] [CrossRef]
- Kolpakov, A.I.; Il’inskaia, O.N. The optimization of a method for determining RNAse activity by using high-polymer RNA. Klin Lab Diagn 1999, 5, 14–16. [Google Scholar]
- Mosher, D.F.; Blout, E.R. Heterogeneity of Bovine Fibrinogen and Fibrin. J. Biol. Chem. 1973, 248, 6896–6903. [Google Scholar] [CrossRef] [PubMed]
- Pace, C.N.; Vajdos, F.; Fee, L.; Grimsley, G.; Gray, T. How to Measure and Predict the Molar Absorption Coefficient of a Protein. Protein Sci. 1995, 4, 2411–2423. [Google Scholar] [CrossRef] [PubMed]
- Castanho, M.; Santos, N.; Loura, L. Separating the turbidity spectra of vesicles from the absorption spectra of membrane probes and other chromophores. Eur. Biophys. J. 1997, 26, 253–259. [Google Scholar] [CrossRef]
- Wakamatsu, K.; Hosoda, K.; Mitomo, H.; Ohya, M.; Okahata, Y.; Yasunaga, K. Direct in Situ Measurement of Phospholipid Hydration in an Aqueous Environment Using a Quartz Crystal Microbalance. Anal. Chem. 1995, 67, 3336–3341. [Google Scholar] [CrossRef]
- Hantgan, R.R.; Hermans, J. Assembly of Fibrin. A Light Scattering Study. J. Biol. Chem. 1979, 254, 11272–11281. [Google Scholar] [CrossRef] [PubMed]
- Tutwiler, V.; Litvinov, R.I.; Lozhkin, A.P.; Peshkova, A.D.; Lebedeva, T.; Ataullakhanov, F.I.; Spiller, K.L.; Cines, D.B.; Weisel, J.W. Kinetics and mechanics of clot contraction are governed by the molecular and cellular composition of the blood. Blood 2016, 127, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Litvinov, R.I.; Faizullin, D.A.; Zuev, Y.F.; Weisel, J.W. The α-Helix to β-Sheet Transition in Stretched and Compressed Hydrated Fibrin Clots. Biophys. J. 2012, 103, 1020–1027. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | Ttr (°C) |
|---|---|
| DPPC | 40.29 ± 0.05 |
| DPPC + binase | 39.45 ± 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faizullin, D.; Valiullina, Y.; Salnikov, V.; Zelenikhin, P.; Zuev, Y.; Ilinskaya, O. Fibrin-Rhamnogalacturonan I Composite Gel for Therapeutic Enzyme Delivery to Intestinal Tumors. Int. J. Mol. Sci. 2023, 24, 926. https://doi.org/10.3390/ijms24020926
Faizullin D, Valiullina Y, Salnikov V, Zelenikhin P, Zuev Y, Ilinskaya O. Fibrin-Rhamnogalacturonan I Composite Gel for Therapeutic Enzyme Delivery to Intestinal Tumors. International Journal of Molecular Sciences. 2023; 24(2):926. https://doi.org/10.3390/ijms24020926
Chicago/Turabian StyleFaizullin, Dzhigangir, Yuliya Valiullina, Vadim Salnikov, Pavel Zelenikhin, Yuriy Zuev, and Olga Ilinskaya. 2023. "Fibrin-Rhamnogalacturonan I Composite Gel for Therapeutic Enzyme Delivery to Intestinal Tumors" International Journal of Molecular Sciences 24, no. 2: 926. https://doi.org/10.3390/ijms24020926