
Searching for Virulence Factors among Staphylococcus lugdunensis Isolates from Orthopedic Infections: Correlation of β-hemolysin, hemolysin III, and slush Genes with Hemolytic Activity and Synergistic Hemolytic Activity
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. PCR
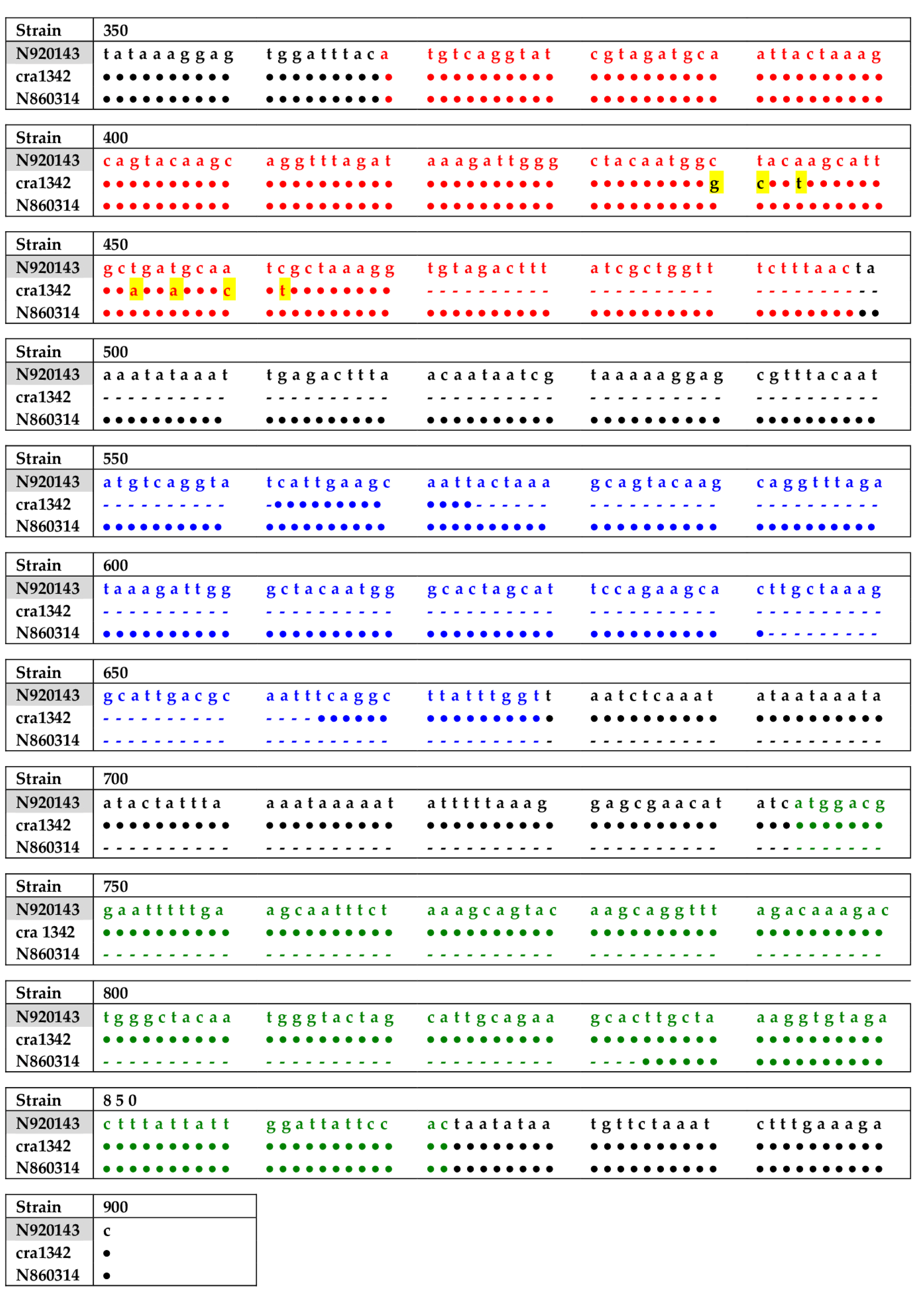
2.2. PCR and Sequencing of Slush Gene
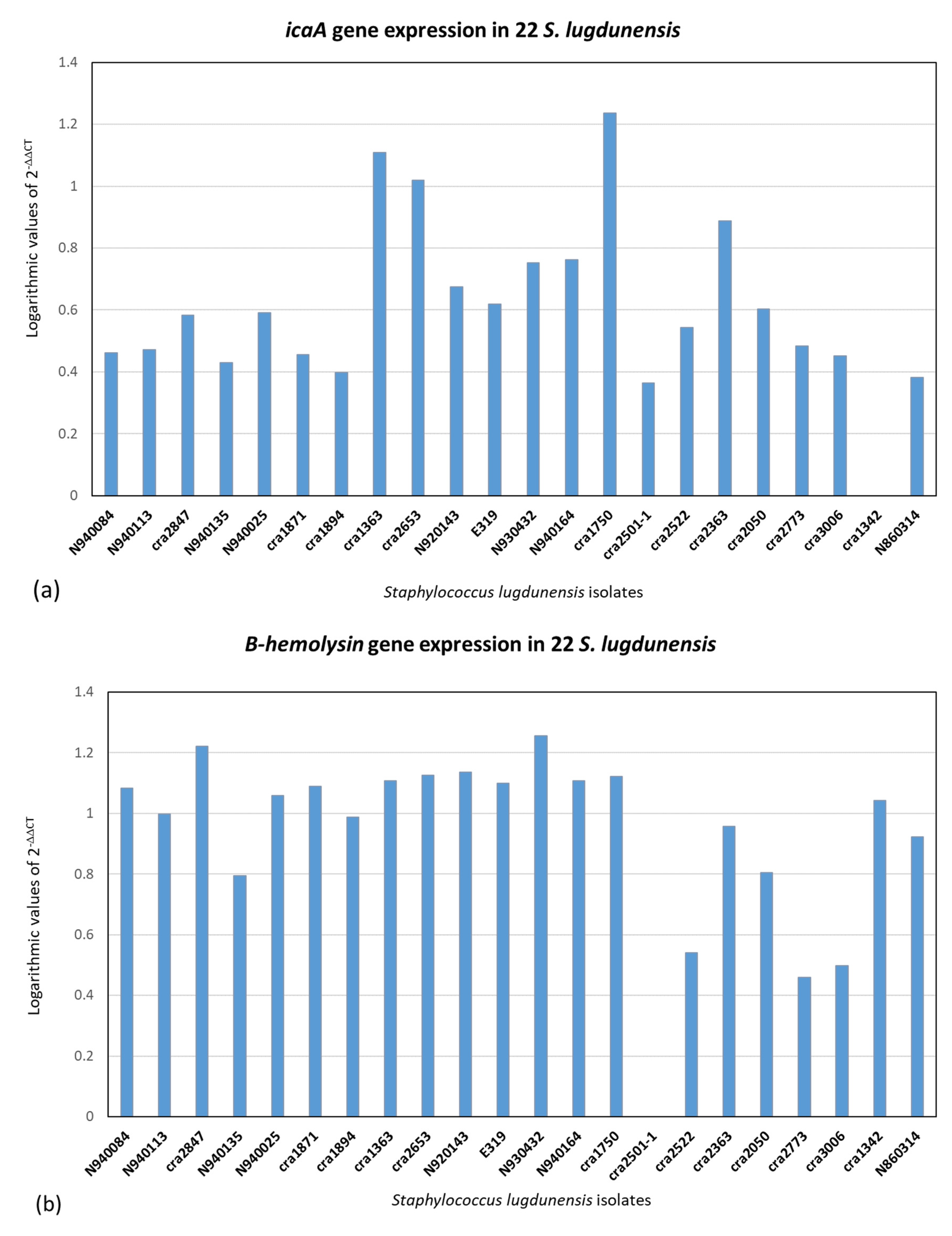
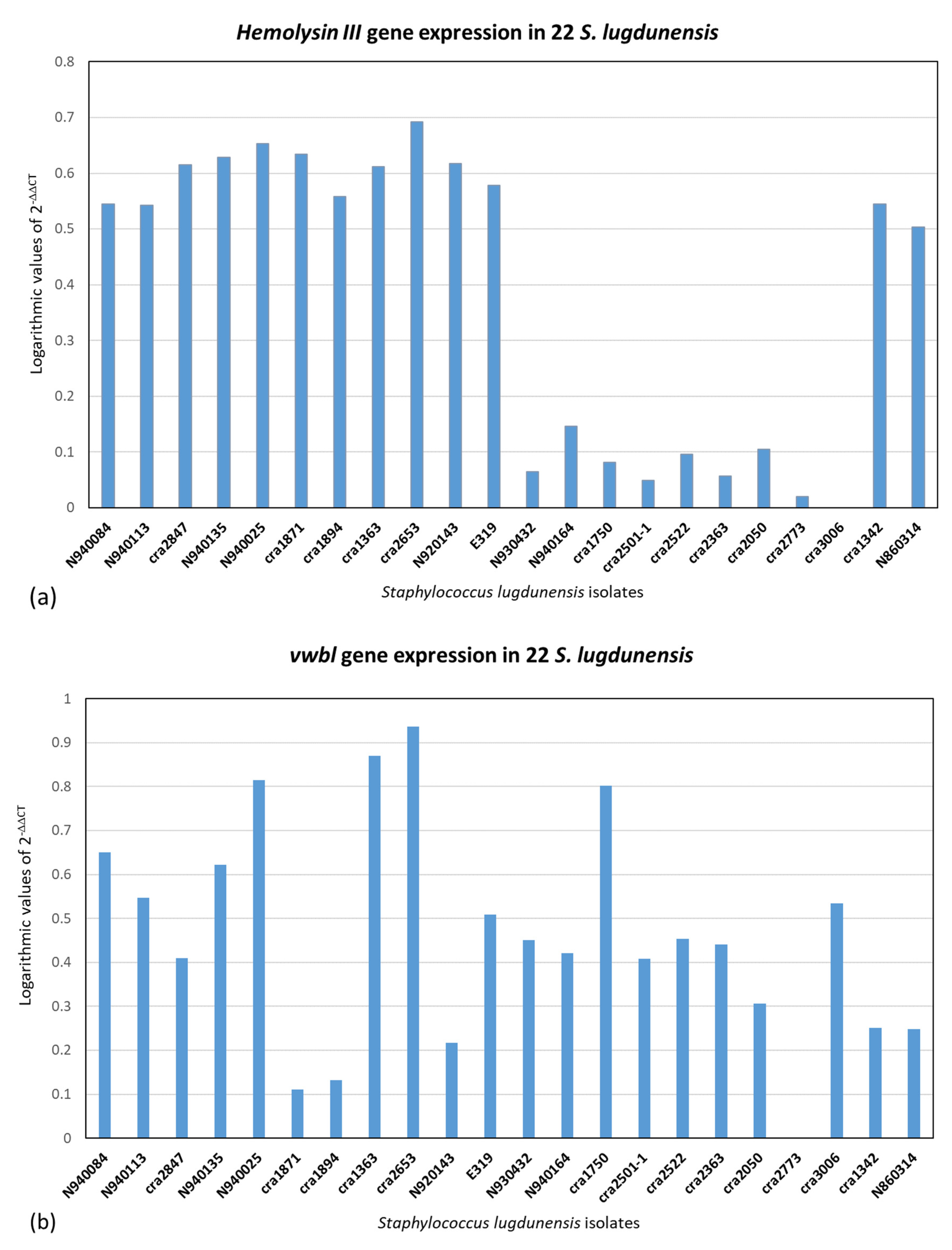
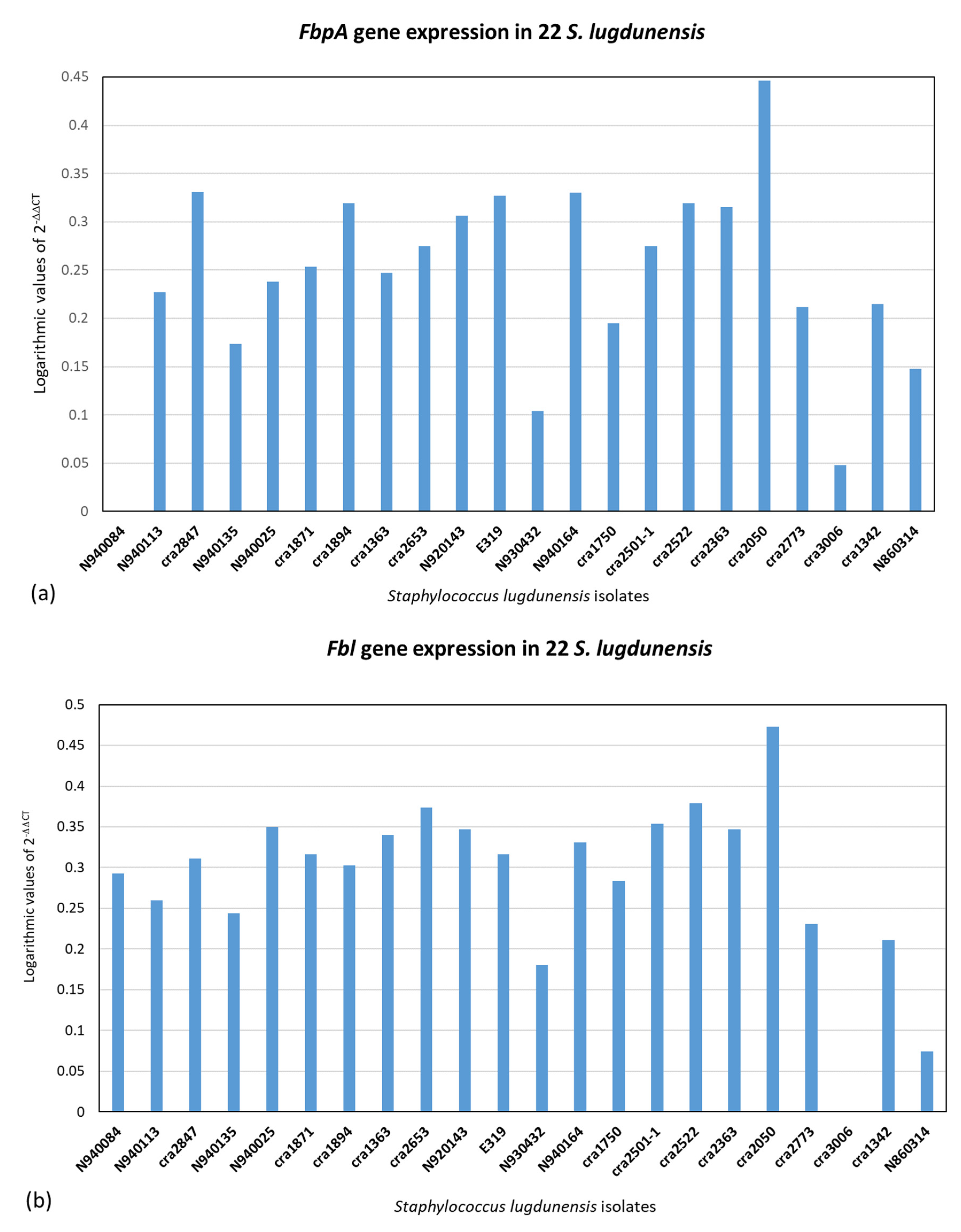
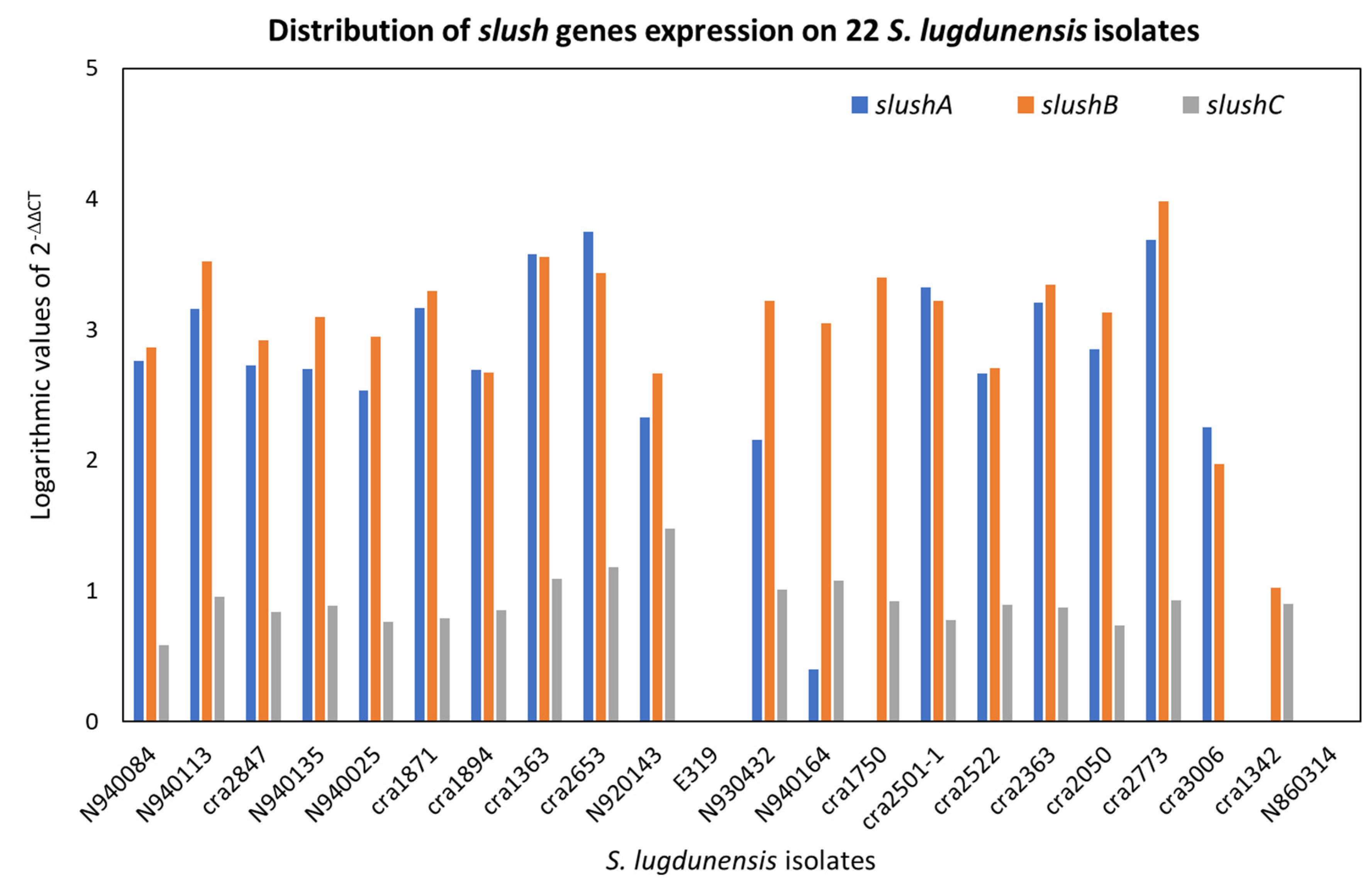
2.3. Quantitative Real-Time PCR (qPCR) of icaA, fbl, vwbl, fbpA, β-Hemolysin and Hemolysin III and (qPCR) of Slush A, B and C Genes
2.4. Hemolytic Test and Synergistic Activity
3. Discussion
4. Materials and Methods
4.1. Species Identification and Storage
4.2. Staphylococcus Lugdunensis Subtyping and Biofilm Characterization
4.3. Bacterial DNA Isolation
4.4. PCR
4.5. Slush Gene Amplicons Sequencing
4.6. Quantitative Real-Time PCR of icaA, fbl, vwbl, fbpA, β-Hemolysin and Hemolysin III
4.7. Quantitative Real-Time PCR (qPCR) of Slush A, B and C Genes
4.7.1. Primers and Probe Design
4.7.2. Quantitative Real-Time PCR Conditions
4.8. Hemolysis Test
4.9. Synergistic Hemolysis Growth Test
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Freney, J.; Brun, Y.; Bes, M.; Meugnier, H.; Grimont, F.; Grimont, P.; Nervi, C.; Fleurette, J. Staphylococcus lugdunensis sp. nov and Staphylococcus schleiferi sp. nov., two species from human clinical specimens. Int. J. Syst. Evol. Microbiol. 1988, 38, 168–172. [Google Scholar] [CrossRef]
- Argemi, X.; Martin, V.; Loux, V.; Dahyot, S.; Lebeurre, J.; Guffroy, A.; Martin, M.; Velay, A.; Keller, D.; Riegel, P.; et al. Whole-Genome Sequencing of Seven Strains of Staphylococcus lugdunensis Allows Identification of Mobile Genetic Elements. Genome Biol. Evol. 2017, 9, 1183–1189. [Google Scholar]
- Hagstrand Aldman, M.; Thompson, O.; Påhlman, L.I. Biofilm formation is associated with poor outcome in prosthetic joint infections caused by Staphylococcus lugdunensis. Infect. Dis. 2023, 55, 328–332. [Google Scholar] [CrossRef] [PubMed]
- García-Malinis, A.J.; Milagro, A.; Torres Sopena, L.; Gilaberte, Y. Staphylococcus lugdunensis Skin Infection: Report of 16 Cases. Actas Dermo-Sifiliográficas (Engl. Ed.) 2021, 112, 261–265, (In English, Spanish). [Google Scholar] [CrossRef]
- Parthasarathy, S.; Shah, S.; Raja Sager, A.; Rangan, A.; Durugu, S. Staphylococcus lugdunensis: Review of epidemiology, complications, and treatment. Cureus 2020, 12, e8801. [Google Scholar] [CrossRef]
- Non, L.R.; Santos, C.A. The occurrence of infective endocarditis with Staphylococcus lugdunensis bacteremia: A retrospective cohort study and systematic review. J. Infect. 2017, 74, 179–186. [Google Scholar] [CrossRef]
- Douiri, N.; Hansmann, Y.; Lefebvre, N.; Riegel, P.; Martin, M.; Baldeyrou, M.; Christmann, D.; Prevost, G.; Argemi, X. Staphylococcus lugdunensis: A virulent pathogen causing bone and joint infections. Clin. Microbiol. Infect. 2016, 22, 747–748. [Google Scholar] [CrossRef]
- Lourtet-Hascoët, J.; Bicart-See, A.; Félicé, M.P.; Giordano, G.; Bonnet, E. Staphylococcus lugdunensis, a serious pathogen in periprosthetic joint infections: Comparison to Staphylococcus aureus and Staphylococcus epidermidis. Int. J. Infect. Dis. 2016, 51, 56–61. [Google Scholar] [CrossRef]
- Frank, K.L.; Del Pozo, J.L.; Patel, R. From clinical microbiology to infection pathogenesis: How daring to be different works for Staphylococcus lugdunensis. Clin. Microbiol. Rev. 2008, 21, 111–133. [Google Scholar] [CrossRef]
- Ebright, J.R.; Penugonda, N.; Brown, W. Clinical experience with Staphylococcus lugdunensis bacteremia: A retrospective analysis. Diagn. Microbiol. Infect. Dis. 2004, 48, 17–21. [Google Scholar] [CrossRef]
- Fernández-Fernández, R.; Lozano, C.; Ruiz-Ripa, L.; Robredo, B.; Azcona-Gutiérrez, J.M.; Alonso, C.A.; Aspiroz, C.; Zarazaga, M.; Torres, C. Antimicrobial Resistance and Antimicrobial Activity of Staphylococcus lugdunensis Obtained from Two Spanish Hospitals. Microorganisms 2022, 10, 1480. [Google Scholar] [CrossRef]
- Betrán, A.; Cebollada, R.; Lavilla, M.J.; Torres, L. Infección por Staphylococcus lugdunensis: Descripción de 44 casos [Staphylococcus lugdunensis infection: Report of 44 cases]. Rev. Chil. De Infectología 2022, 39, 254–259. (In Spanish) [Google Scholar] [CrossRef] [PubMed]
- Kachrimanidou, M.; Malliou, P.; Meletis, G.; Netsika, F.; Mavrovouniotis, I.; Protonotariou, E.; Skoura, L. Epidemiology and antimicrobial susceptibility of Staphylococcus lugdunensis in a Greek tertiary-care hospital. New Microbiol. 2020, 43, 133–135. [Google Scholar]
- Taha, L.; Stegger, M.; Söderquist, B. Staphylococcus lugdunensis: Antimicrobial susceptibility and optimal treatment options. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1449–1455. [Google Scholar] [CrossRef] [PubMed]
- Fleurette, J.; Bès, M.; Brun, Y.; Freney, J.; Forey, F.; Coulet, M.; Reverdy, M.E.; Etienne, J. Clinical isolates of Staphylococcus lugdunensis and S. schleiferi: Bacteriological characteristics and susceptibility to antimicrobial agents. Res. Microbiol. 1989, 140, 107–118. [Google Scholar] [CrossRef]
- Geoghegan, J.A.; Ganesh, V.K.; Smeds, E.; Liang, X.; Hook, M.; Foster, T.J. Molecular characterization of the interaction of staphylococcal microbial surface components recognizing adhesive matrix molecules (MSCRAMM) ClfA and Fbl with fibrinogen. J. Biol. Chem. 2010, 285, 6208–6216. [Google Scholar] [CrossRef] [PubMed]
- Szabados, F.; Nowotny, Y.; Marlinghaus, L.; Korte, M.; Neumann, S.; Kaase, M.; Gatermann, S.G. Occurrence of genes of putative fibrinogen binding proteins and hemolysins, as well as of their phenotypic correlates in isolates of S. lugdunensis of different origins. BMC Res. Notes 2011, 4, 113. [Google Scholar] [CrossRef]
- Lebeurre, J.; Dahyot, S.; Diene, S.; Paulay, A.; Aubourg, M.; Argemi, X.; Giard, J.C.; Tournier, I.; François, P.; Pestel-Caron, M. Comparative Genome Analysis of Staphylococcus lugdunensis Shows Clonal Complex-Dependent Diversity of the Putative Virulence Factor, ess/Type VII Locus. Front. Microbiol. 2019, 10, 2479. [Google Scholar] [CrossRef]
- Heilbronner, S.; Holden, M.T.; van Tonder, A.; Geoghegan, J.A.; Foster, T.J.; Parkhill, J.; Bentley, S.D. Genome sequence of Staphylococcus lugdunensis N920143 allows identification of putative colonization and virulence factors. FEMS Microbiol. Lett. 2011, 322, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Weidenmaier, C.; Peschel, A.; Kempf, V.A.; Lucindo, N.; Yeaman, M.R.; Bayer, A.S. DltABCD- and MprF-mediated cell envelope modifications of Staphylococcus aureus confer resistance to platelet microbicidal proteins and contribute to virulence in a rabbit endocarditis model. Infect. Immun. 2005, 73, 8033–8038. [Google Scholar] [CrossRef]
- Donvito, B.; Etienne, J.; Denoroy, L.; Greenland, T.; Benito, Y.; Vandenesch, F. Synergistic hemolytic activity of Staphylococcus lugdunensis is mediated by three peptides encoded by a non-agr genetic locus. Infect. Immun. 1997, 65, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Hébert, G.A. Hemolysins and other characteristics that help differentiate and biotype Staphylococcus lugdunensis and Staphylococcus schleiferi. J. Clin. Microbiol. 1990, 28, 2425–2431. [Google Scholar] [CrossRef] [PubMed]
- Ravaioli, S.; Campoccia, D.; Speziale, P.; Pietrocola, G.; Zatorska, B.; Maso, A.; Presterl, E.; Montanaro, L.; Arciola, C.R. Various biofilm matrices of the emerging pathogen Staphylococcus lugdunensis: Exopolysaccharides, proteins, eDNA and their correlation with biofilm mass. Biofouling 2020, 36, 86–100. [Google Scholar] [CrossRef] [PubMed]
- Didi, J.; Lemée, L.; Gibert, L.; Pons, J.L.; Pestel-Caron, M. Multi-virulence-locus sequence typing of Staphylococcus lugdunensis generates results consistent with a clonal population structure and is reliable for epidemiological typing. J. Clin. Microbiol. 2014, 52, 3624–3632. [Google Scholar] [CrossRef]
- Lin, L.C.; Cheng, C.W.; Chang, S.C.; Lu, J.J. Molecular Epidemiological Survey of Staphylococcus lugdunensis Isolates with Variable Number of Repeats in the von Willebrand Factor-Binding Protein Gene. Front. Cell. Infect. Microbiol. 2021, 11, 748640. [Google Scholar] [CrossRef]
- Chang, S.C.; Lin, L.C.; Lu, J.J. Comparative Genomic Analyses Reveal Potential Factors Responsible for the ST6 Oxacillin-Resistant Staphylococcus lugdunensis Endemic in a Hospital. Front. Microbiol. 2021, 12, 765437. [Google Scholar] [CrossRef]
- Arciola, C.R.; Campoccia, D.; Gamberini, S.; Rizzi, S.; Donati, M.E.; Baldassarri, L.; Montanaro, L. Search for the insertion element IS256 within the ica locus of Staphylococcus epidermidis clinical isolates collected from biomaterial-associated infections. Biomaterials 2004, 25, 4117–4125. [Google Scholar] [CrossRef]
- Ji, G.; Beavis, R.; Novick, R.P. Bacterial interference caused by autoinducing peptide variants. Science 1997, 276, 2027–2030. [Google Scholar] [CrossRef]
- Chin, D.; Flannagan, R.S.; Tuffs, S.W.; Chan, J.K.; McCormick, J.K.; Heinrichs, D.E. Staphylococcus lugdunensis Uses the Agr Regulatory System to Resist Killing by Host Innate Immune Effectors. Infect. Immun. 2022, 90, e0009922. [Google Scholar] [CrossRef]
- Ortega-Peña, S.; Vargas-Mendoza, C.F.; Franco-Cendejas, R.; Aquino-Andrade, A.; Vazquez-Rosas, G.J.; Betanzos-Cabrera, G.; Guerrero-Barajas, C.; Jan-Roblero, J.; Rodríguez-Martínez, S.; Cancino-Diaz, M.E.; et al. sesA, sesB, sesC, sesD, sesE, sesG, sesH, and embp genes are genetic markers that differentiate commensal isolates of Staphylococcus epidermidis from isolates that cause prosthetic joint infection. Infect. Dis. 2019, 51, 435–445. [Google Scholar] [CrossRef]
- Sánchez, A.; Benito, N.; Rivera, A.; García, L.; Miró, E.; Mur, I.; González, Y.; Gutiérrez, C.; Horcajada, J.P.; Espinal, P.; et al. Pathogenesis of Staphylococcus epidermidis in prosthetic joint infections: Can identification of virulence genes differentiate between infecting and commensal strains? J. Hosp. Infect. 2020, 105, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Chassain, B.; Lemée, L.; Didi, J.; Thiberge, J.M.; Brisse, S.; Pons, J.L.; Pestel-Caron, M. Multilocus sequence typing analysis of Staphylococcus lugdunensis implies a clonal population structure. J. Clin. Microbiol. 2012, 50, 3003–3009. [Google Scholar] [CrossRef]
- Ravaioli, S.; Campoccia, D.; Ruppitsch, W.; Allerberger, F.; Poggi, A.; Chisari, E.; Montanaro, L.; Arciola, C.R. Comparison of Automated Ribotyping, spa Typing, and MLST in 108 Clinical Isolates of Staphylococcus aureus from Orthopedic Infections. Int. J. Mol. Sci. 2022, 23, 1660. [Google Scholar] [CrossRef] [PubMed]
- Campoccia, D.; Baldassarri, L.; Pirini, V.; Ravaioli, S.; Montanaro, L.; Arciola, C.R. Molecular epidemiology of Staphylococcus aureus from implant orthopaedic infections: Ribotypes, agr polymorphism, leukocidal toxins and antibiotic resistance. Biomaterials 2008, 29, 4108–4116. [Google Scholar] [CrossRef]
- Campoccia, D.; Speziale, P.; Ravaioli, S.; Cangini, I.; Rindi, S.; Pirini, V.; Montanaro, L.; Arciola, C.R. The presence of both bone sialoprotein-binding protein gene and collagen adhesin gene as a typical virulence trait of the major epidemic cluster in isolates from orthopedic implant infections. Biomaterials 2009, 30, 6621–6628. [Google Scholar] [CrossRef]
- Cochis, A.; Azzimonti, B.; Della Valle, C.; Chiesa, R.; Arciola, C.R.; Rimondini, L. Biofilm formation on titanium implants counteracted by grafting gallium and silver ions. J. Biomed. Mater. Res. A 2015, 103, 1176–1187. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A.; Shakoor, A. Recent Strategies and Future Recommendations for the Fabrication of Antimicrobial, Antibiofilm, and Antibiofouling Biomaterials. Int. J. Nanomed. 2023, 21, 3377–3405. [Google Scholar] [CrossRef]
- Poggio, C.; Colombo, M.; Arciola, C.R.; Greggi, T.; Scribante, A.; Dagna, A. Copper-Alloy Surfaces and Cleaning Regimens against the Spread of SARS-CoV-2 in Dentistry and Orthopedics. From Fomites to Anti-Infective Nanocoatings. Materials 2020, 13, 3244. [Google Scholar] [CrossRef]
- Campoccia, D.; Montanaro, L.; Ravaioli, S.; Mariani, V.; Bottau, G.; De Donno, A.; Arciola, C.R. Antibacterial Activity on Orthopedic Clinical Isolates and Cytotoxicity of the Antimicrobial Peptide Dadapin-1. Int. J. Mol. Sci. 2023, 24, 779. [Google Scholar] [CrossRef]
- Hussain, Y.; Alam, W.; Ullah, H.; Dacrema, M.; Daglia, M.; Khan, H.; Arciola, C.R. Antimicrobial Potential of Curcumin: Therapeutic Potential and Challenges to Clinical Applications. Antibiotics 2022, 11, 322. [Google Scholar] [CrossRef] [PubMed]
- Baldan, R.; Sendi, P. Precision Medicine in the Diagnosis and Management of Orthopedic Biofilm Infections. Front. Med. 2020, 7, 580671. [Google Scholar] [CrossRef] [PubMed]
- Arciola, C.R.; Campoccia, D.; Montanaro, L. Implant infections: Adhesion, biofilm formation and immune evasion. Nat. Rev. Microbiol. 2018, 16, 397–409. [Google Scholar] [CrossRef]
- Jochim, A.; Adolf, L.; Belikova, D.; Schilling, N.A.; Setyawati, I.; Chin, D.; Meyers, S.; Verhamme, P.; Heinrichs, D.E.; Slotboom, D.J.; et al. An ECFtype transporter scavenges heme to overcome iron-limitation in Staphylococcus lugdunensis. eLife 2020, 9, e57322. [Google Scholar] [CrossRef] [PubMed]
- Aubourg, M.; Gravey, F.; Dhalluin, A.; Giard, J.C. Identification of the iron-limitation stimulon in Staphylococcus lugdunensis. Arch. Microbiol. 2021, 203, 3687–3694. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2−ΔΔCT method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinform. Biomath. 2013, 3, 71–85. [Google Scholar]
- Hébert, G.A.; Hancock, G.A. Synergistic hemolysis exhibited by species of staphylococci. J. Clin. Microbiol. 1985, 22, 409–415. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Ribogroup | 24 h Hemolysis | 48 h Hemolysis | 72 h Hemolysis | S. aureus Synergy | Log SlushA | Log SlushB | Log SlushC |
|---|---|---|---|---|---|---|---|---|
| N940084 | cra-62-S1 | positive | positive | 2.757 | 2.867 | 0.587 | ||
| N940113 | cra-62-S1 | positive | positive | 3.161 | 3.522 | 0.955 | ||
| cra2847 | cra-62-S1 | negative | positive | positive | 2.728 | 2.916 | 0.842 | |
| N940025 | cra-62-S1 | positive | positive | 2.535 | 2.949 | 0.763 | ||
| N940135 | cra-62-S1 | negative | positive | positive | 2.696 | 3.095 | 0.890 | |
| cra1871 | cra-62-S1 | positive | positive | 3.167 | 3.298 | 0.790 | ||
| cra1894 | cra-62-S1 | positive | positive | 2.691 | 2.673 | 0.850 | ||
| cra1363 | cra-62-S1 | positive | positive | 3.574 | 3.558 | 1.091 | ||
| cra2653 | cra-62-S1 | negative | positive | positive | 3.747 | 3.435 | 1.184 | |
| N920143 | cra-62-S1 | negative | negative | weak | negative | 2.328 | 2.666 | 1.479 |
| E319 | cra-62-S1 | negative | negative | weak | negative | 0 | 0 | 0 |
| N930432 | cra-193-S3 | negative | negative | weak | negative | 2.154 | 3.219 | 1.009 |
| N940164 | cra-74-S5 | negative | negative | weak | negative | 0.403 | 3.047 | 1.080 |
| cra1750 | cra-74-S5 | positive | positive | 0 | 3.401 | 0.923 | ||
| cra2501–1 | cra-64-S8 | positive | positive | 3.325 | 3.220 | 0.779 | ||
| cra2522 | cra-64-S8 | negative | positive | positive | 2.667 | 2.707 | 0.891 | |
| cra2363 | cra-64-S8 | negative | positive | positive | 3.207 | 3.347 | 0.875 | |
| cra2050 | cra-64-S8 | negative | positive | positive | 2.848 | 3.132 | 0.737 | |
| cra2773 | cra-64-S8 | negative | positive | negative | 3.688 | 3.979 | 0.929 | |
| cra3006 | cra-64-S8 | negative | negative | weak | positive | 2.250 | 1.970 | 0 |
| cra1342 | cra-129-S8 | positive | Positive | 0 | 1.027 | 0.903 | ||
| N860314 | cra-129-S8 | negative | positive | Positive | 0 | 0 | 0 |
| Primer/Probe Sequence (5′–3′) | Target Name | Amplicon Size |
|---|---|---|
| CAG GTA TCG TAG ATG CAA TTT CAA | slushA fw | 126 bp |
| AGT TAA AGA AAC CAG CGA TAA AGT C | slushA rev | |
| /56-FAM/AA GCA TTG C + T + G A + TG CA + A TC/3IABkFQ/ | Probe slushA | |
| TGT CAG GTA TCA TTG AAG CAA TTA C | slushB fw | 119 bp |
| GCC TGA AAT TGC GTC AAT GC | slushB rev | |
| /56-FAM/AA + TGG + G + C + A CT + AGCA/3IABkFQ/ | Probe slushB | |
| ATG GAC GGA ATT TTT GAA GCA | slushC fw | 129 bp |
| GTG GAA TAA TCC AAT AAT AAA GTC TAC AC | slushC rev | |
| /56-FAM/AG + A + C + A AAG + A+C + T GGG C/3IABkFQ/ | Probe slushC | |
| TCT AAA CTT GGT GGC GCT AAA | GMK fw | |
| CGA TGG AAG CTG GAC ATG AT | GMK rev | |
| /56-FAM/AG TGC GTC C/ZEN/G GGA ATT TCT TCC TT/3IABkFQ/ | Probe GMK |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ravaioli, S.; Campoccia, D.; Mirzaei, R.; Mariani, V.; Bottau, G.; De Donno, A.; Montanaro, L.; Speziale, P.; Arciola, C.R. Searching for Virulence Factors among Staphylococcus lugdunensis Isolates from Orthopedic Infections: Correlation of β-hemolysin, hemolysin III, and slush Genes with Hemolytic Activity and Synergistic Hemolytic Activity. Int. J. Mol. Sci. 2023, 24, 15724. https://doi.org/10.3390/ijms242115724
Ravaioli S, Campoccia D, Mirzaei R, Mariani V, Bottau G, De Donno A, Montanaro L, Speziale P, Arciola CR. Searching for Virulence Factors among Staphylococcus lugdunensis Isolates from Orthopedic Infections: Correlation of β-hemolysin, hemolysin III, and slush Genes with Hemolytic Activity and Synergistic Hemolytic Activity. International Journal of Molecular Sciences. 2023; 24(21):15724. https://doi.org/10.3390/ijms242115724
Chicago/Turabian StyleRavaioli, Stefano, Davide Campoccia, Rasoul Mirzaei, Valentina Mariani, Giulia Bottau, Andrea De Donno, Lucio Montanaro, Pietro Speziale, and Carla Renata Arciola. 2023. "Searching for Virulence Factors among Staphylococcus lugdunensis Isolates from Orthopedic Infections: Correlation of β-hemolysin, hemolysin III, and slush Genes with Hemolytic Activity and Synergistic Hemolytic Activity" International Journal of Molecular Sciences 24, no. 21: 15724. https://doi.org/10.3390/ijms242115724