
The Feeder Effects of Cultured Rice Cells on the Early Development of Rice Zygotes

1
Department of Biological Sciences, Tokyo Metropolitan University, Minami-Osawa 1-1, Hachioji 192-0397, Tokyo, Japan
2
Department of Chemistry, Tokyo Metropolitan University, Minami-Osawa 1-1, Hachioji 192-0397, Tokyo, Japan
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(22), 16541; https://doi.org/10.3390/ijms242216541
Submission received: 26 October 2023
/
Revised: 13 November 2023
/
Accepted: 16 November 2023
/
Published: 20 November 2023
(This article belongs to the Special Issue State-of-the-Art Molecular Plant Sciences in Japan)
Abstract
:Feeder cells and the synthetic auxin 2,4-dichlorophenoxyacetic acid (2,4-D) in a culture medium promote mitosis and cell division in cultured cells. These are also added to nutrient medium for the cultivation of highly active in mitosis and dividing zygotes, produced in vitro or isolated from pollinated ovaries. In the study, an in vitro fertilization (IVF) system was used to study the precise effects of feeder cells and 2,4-D on the growth and development of rice (Oryza sativa L.) zygote. The elimination of 2,4-D from the culture medium did not affect the early developmental profiles of the zygotes, but decreased the division rates of multicellular embryos. The omission of feeder cells resulted in defective karyogamy, fusion between male and female nuclei, and the subsequent first division of the cultured zygotes. The culture of zygotes in a conditioned medium corrected developmental disorders. Proteome analyses of the conditioned medium revealed the presence of abundant hydrolases possibly released from the feeder cells. Exogenously applied α-amylase ameliorated karyogamy and promoted zygote development. It is suggested that hydrolytic enzymes, including α-amylase, released from feeder cells may be involved in the progression of zygotic development.
Keywords:
amylase; cell wall; feeder cell; fertilization; hydrolytic enzyme; rice; zygote; zygotic development; 2,4-D1. Introduction
In angiosperms, a zygote, a product of the fertilization of an egg cell with a sperm cell, divides into an asymmetric two-celled embryo [1,2,3,4,5]. In rice, the two-celled embryo continues to divide and develop into a multicellular and then a globular and mature embryo [6,7]. As fertilization and subsequent zygotic development occur in the embryo sac deeply embedded in the ovule of plants, an in vitro fertilization (IVF) system, using microtechniques for three stages, gamete isolation from flowers, the electric fusion of gametes, and a culture of the produced zygote, was applied for a direct and precise analysis of zygotic development [8,9]. Such an IVF system has been established in rice, maize, and wheat [7,10,11,12,13]. A synthetic auxin 2,4-dichlorophenoxyacetic acid (2,4-D) and feeder cells, crucial for the developing IVF-produced zygotes in rice, wheat, and maize, are commonly incorporated in the culture media for the cultivation of somatic cells and protoplasts [14,15,16]. It has been reported that auxins exert diverse effects on cultured somatic cells, such as mitotic activity enhancement, cell elongation/growth, and differentiation [17,18]. The mitotic activities of isolated cells/protoplasts depend/related to cell density in culture [15,16,19,20], and feeder cells are often co-cultured with isolated cells to increase cell density as conditioning factors for growth and development [16,21].
Based on intensive studies on somatic cells/protoplasts, 2,4-D and protein/peptide components present in cell cultures are known to trigger the division and development of somatic cells or protoplasts in vitro, although their division efficiency is low, typically 0.1–0.6% [20,22]. In contrast to somatic cells/protoplasts, zygotes possess totipotent potentials and are highly active with a high division efficiency. It has been indicated that more than 80% of rice and wheat zygotes produced by IVF divide and develop into globular embryos [10,13]. Auxin 2,4-D and feeder cells are supposed to support the developmental potential of cultured zygotes. However, the precise effects of feeder cells and 2,4-D on zygote cultures remain unclear. In this study, we examined the effects of 2,4-D and feeder cells on the developmental profiles of rice zygotes produced by the IVF system and identified α-amylase, possibly secreted by feeder cells, as a candidate conditioning factor in the initial development of rice zygotes.
2. Results
2.1. 2,4-Dichlorophenoxyacetic Acid Enhances the Division of Embryonic Cells during or after the Multicellular Stage
In this study, a sperm cell isolated from transformed rice plants expressing histone H2B-green fluorescent protein (H2B-GFP) was fused with a wild-type egg cell to monitor the completion of karyogamy, the fusion between male and female nuclei [23], and the division profiles of zygotes [24]. Then, 128 zygotes were produced using IVF and cultured in an N6Z medium containing 2,4-D. Among these 128 zygotes, karyogamy was detected in 105 (Figure 1(Aa,Ab), Table 1) and 98 of the 105 zygotes divided and developed into two-celled embryos (Figure 1(Ac,Ad), Table 1). Subsequently, the two-celled embryos developed mainly into multicellular embryos (Figure 1(Ae,Af)), globular-like embryos (Figure 1(Ag,Ah)), and cell masses (Figure 1(Ai–An), Table 1). Rice zygotes and their subsequent embryos divided and developed with high efficiencies when cultured in the N6Z medium (Table 1), consistent with previous reports [10,25].
Karyogamy was detected in 53 of the 73 zygotes cultured in 2,4-D free N6Z medium (Figure 1(Ba,Bb), Table 1). In total, 47 and 43 of these 53 zygotes divided and developed into two-celled and multicellular embryos, respectively (Figure 1(Bc–Bf), Table 1). Although 28 of the 43 multicellular embryos developed into cell masses (Figure 1(Bg–Bn), Table 1), the remaining 15 showed developmental arrest at the multicellular stage (Figure 1C). These results suggest that 2,4-D functions in the progression of embryonic cell division during or after the multicellular stage.
2.2. Feeder Cells Promote Karyogamy and Subsequent Cell Division in Zygotes
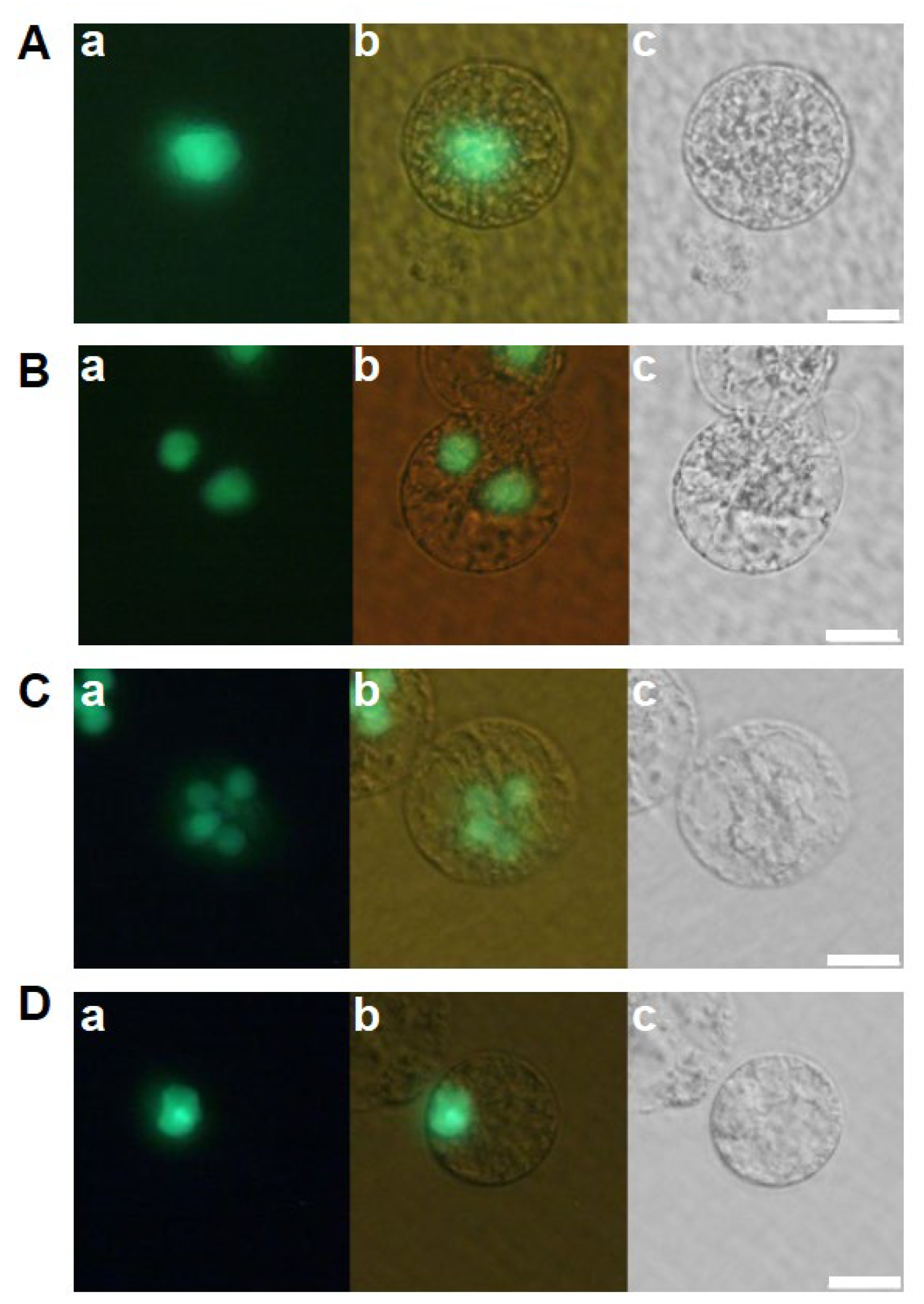
The developmental profiles of zygotes cultured in the feeder cell-free medium were examined to investigate the effects of feeder cells on zygotic development. Among the 103 fertilized eggs cultured in the feeder cell-free N6Z medium, H2B-GFP-labeled zygotic nuclei, an indication of the completion of karyogamy, were detected only in 19 zygotes (Figure 2(Aa,Ab), Table 1). The remaining 84 zygotes appeared to degenerate without karyogamy progression (Figure 2(Ba,Bb)). Of the 19 zygotes with H2B-GFP-labeled nuclei, 5 showed nuclear division but degenerated later (Figure 2(Ca,Cb), Table 1), strongly suggesting that the efficiencies of karyogamy progression and the subsequent first division in zygotes were markedly reduced by the omission of feeder cells from the culture medium. We further tested the culture of zygotes in 2,4-D free N6Z-medium without feeder cells. The developmental profiles of the zygotes were equivalent to those of zygotes cultured in the N6Z medium without feeder cells (Figure 2(Da–Fb), Table 1). Together, these results suggest that the efficiencies of karyogamy progression and zygotic division were reduced in response to feeder cell omission regardless of the presence of 2,4-D. Moreover, karyogamy and subsequent zygotic division are likely promoted by potential substances believed to be secreted by feeder cells.
2.3. Heat-Unstable Proteins in Culture Medium Function as Development-Promoting Substances for Zygotes
A conditioned medium was prepared as described in the Section 4, and IVF-produced zygotes were cultured in the medium to examine whether substances secreted from the feeder cells showed positive feeder effects on the progression of zygotic development. Karyogamy was detected in 87 of 104 zygotes (Figure 3(Aa–Ac), Table 1). In total, 72 and 51 of these 87 zygotes were divided into two-celled and multicellular embryos, respectively (Figure 3(Ad–Ai), Table 1). In the conditioned medium, the frequency of zygotes that completed karyogamy and developed into two-celled/multicellular embryos was greatly increased compared to that of zygotes cultured without feeder cells (Table 1). Although 8 of the 51 multicellular embryos progressed into the cell mass stage (Figure 3(Aj–Ar)), the remaining embryos showed arrested growth (Figure 3B, Table 1). These results suggest that the conditioned medium rescued the efficiencies of karyogamy and subsequent zygotic development and that the conditioned medium contained potential development-promoting substances (DPSs) secreted by the cultured Oc cells.
Next, to determine the molecular nature of DPSs, we tested their heat stability by culturing the zygotes in a preheated or non-heated control-conditioned medium. In the case of zygote culture in the non-heated conditioned medium, karyogamy was detected in 24 of the 39 zygotes tested (Figure 4(Aa–Ac), Table 2). In addition, 11 of these 24 zygotes divided and developed into two-celled embryos, with 3 multicellular embryos being formed (Figure 4(Ba–Bc,Ca–Cc), Table 2). Karyogamy was detected in 16 of the 37 zygotes cultured in the heat-treated conditioned medium (Figure 4(Da–Dc), Table 2). However, none of the 16 zygotes showed a division profile during culture and degenerated without division (Table 2). These results suggest that heat treatment of the conditioned medium decreased the activities of potential DPSs and that potential DPSs are heat-unstable protein components.
2.4. Hydrolytic Enzymes Were Identified as Potential DPSs for Zygotes
Protein fractions prepared from heat-treated and non-heat-conditioned medium were separated using sodium dodecyl sulfate-polyacrylamide gel electrophoresis to identify heat-unstable proteins in the conditioned medium. The proteins were subjected to in-gel digestion, and the subsequent peptides were analyzed using liquid chromatography coupled with tandem mass spectroscopy. A total of 978 proteins were identified using duplicate proteomic analyses. Among these proteins, ubiquitin proteins were frequently detected in both heat-treated and non-heat-treated media at comparable levels, consistent with the heat-stable nature of ubiquitin (Table 3) [26,27]. Moreover, it provided a quantitative control for a comparative analysis of the protein abundance between the two kinds of conditioned media. As candidate DPSs, 21 proteins detected exclusively or highly preferentially in the non-heat-conditioned medium with a total peptide count of >20 were selected (Table 4). Of these proteins, 15 were hydrolytic enzymes possessing a putative signal peptide for secretion, and 12 of these 15 hydrolytic enzymes belonged to the glycosidase family. Among these glycosidases, α-amylase was most abundantly detected (Table 4); therefore, we attempted to examine the effects of α-amylase in promoting zygotic development.
2.5. Exogenously Applied α-Amylase Partially Promoted the Initial Development of Zygotes
IVF-produced zygotes were cultured in feeder cell and 2,4-D free N6Z medium to which α-amylase derived from Bacillus licheniformis was exogenously added at the concentrations of 017, 1.7, or 17 U/mL to evaluate the effects of α-amylase on zygotic development. For the α-amylase treatments at the 0.17 and 17 U/mL concentrations, the developmental efficiencies in karyogamy progression were comparable to that of the control (Figure 5(Aa–Ac,Ba–Bc,Ea–Ec,Fa–Fc), Table 5). However, efficiency in karyogamy progression increased approximately 2 folds when α-amylase was added at the concentration of 1.7 U/mL (Figure 5(Ca–Cc,Da–Dc), Table 5). These results suggest that α-amylase exogenously applied at appropriate concentrations partially functions as a DPS for zygotic development.
3. Discussion
The present study provided the possibility that hydrolases, especially α-amylase, secreted from feeder cells are involved in the initial developmental steps of rice zygotes cultured in vitro. In the somatic embryogenesis of orchard grass, α-amylase is transiently expressed during early embryo development up to the globular stage and localizes on the cell surface and cell-to-cell adhesion zone in microcluster cells, the earliest morphological structures during somatic embryogenesis [28,29]. In addition, the expression of α-amylase in the somatic embryos of orchard grass is speculated to relate to the embryonic potential of cell mass, although the mechanisms underlying the α-amylase-mediated determination of embryonic character remains to be elucidated [28,29,30]. It is likely that the α-amylase detected in our study also degraded the polysaccharides existing in the cell wall or plasma membrane of the rice zygotes. The determination of the target substance of α-amylase and the responsive machineries in rice zygotes against α-amylase will be required for understanding how the enzyme functions as DPS on rice zygotes.
Hydrolases have been reported as possible conditioning factors [21,28,31,32]. In the present study, classIII chitinase RCB4 was also identified as possible hydrolytic enzyme possessing DPS function (Table 4). The chitinase RCB4 has been purified from rice (O. sativa, Nakdong) cell suspension and characterized as endochitinase with an optimum pH around 4 [33]. Van Hengel et al. (2001) reported that endochitinase acts on the arabinogalactan proteins (AGPs) of the cell wall [22]. Somatic embryogenesis-incompetent cells regain their potential when chitinase-treated AGPs are added to the culture medium [22,34,35], suggesting that AGPs degraded by endochitinase act as signaling molecules to trigger somatic embryogenesis. Immunological assays using antibodies against AGPs have shown that AGPs are abundantly localized in the cells of the rice embryo sac, such as antipodal cells, egg cells, and synergids [36]. In tobacco, Qin and Zhao (2006) reported abundant AGP localization in egg cells and a subsequent APG decrease in zygotes after fertilization [37], suggesting the involvement of AGPs in embryonic development. These suggest that chitinase RCB4 secreted from feeder cells enhances the development of rice zygotes through the possible degradation of APGs in the cell walls around rice zygotes.
Cell wall proteins are not abundant in quantity but are believed to have crucial functions in cell defense, cell wall structure and modification, interactions with plasma membrane proteins, and signal transduction [28]. Zygotes start forming cell walls rapidly after gamete fusion because female and male gametes lack the cell walls to facilitate gamete fusion [38,39]. In addition, it has been reported the cell wall of tobacco zygotes possesses essential functions for maintaining cell polarity, the apical–basal axis, and typical suspensor formation [40]. Certain hydrolytic enzymes, including α-amylase, may be involved in the proper cell wall formation in plant zygotes and subsequent zygotic development.
4. Materials and Methods
4.1. Plant Materials
Oryza sativa L. cv Nipponbare plants were grown in an environmental chamber (K30-7248; Koito Industries, Yokohama, Japan) at 26 °C under a 13 h light/11 h dark photoperiod. Transformed rice plants expressing the histone H2B-green fluorescent protein (H2B-GFP) fusion protein were prepared [41].
4.2. Isolation of Gametes, Electro-Fusion of Gametes, and Subsequent Culture of Zygotes
The isolation of egg and sperm cells from rice flowers and the subsequent electro-fusion of isolated gametes for zygote production were conducted [10,42]. The resulting zygotes were cultured as previously reported [10], with a few modifications. For examining the effects of feeder cells on zygotic development, the zygotes were cultured in the N6Z medium [10,12], with or without feeder cells (rice suspension cell culture, line Oc, provided by Riken Bio-Resource Center, Tsukuba, Japan; https://plant.rtc.riken.jp/resource/cell_line/web_documents/cell_lines/rpc00031.html, 15 November 2023). For analyzing the effects of 2,4-dichlorophenoxyacetic acid (2,4-D) on zygotic development, zygotes were cultured in N6Z medium with or without 2,4-D. For examining the effects of α-amylase on zygotic development, zygotes were cultured in feeder cell- and 2,4-D-free N6Z medium containing 0.17, 1.7, or 17 U/mL of Bacillus licheniformis α-amylase purchased from Sigma, St. Louis, MO, USA.
4.3. Microscopic Observations
Zygotes expressing H2B-GFP were observed under an IX-71 inverted fluorescence microscope (Olympus, Tokyo, Japan) with 460–490 nm excitation and 510–550 nm emission wavelengths (U-MWIBA2 mirror unit; Olympus, Tokyo, Japan). Digital images of the zygotes and their resulting embryos were obtained using a cooled charge-coupled device camera (DP73; Olympus) with cellSens software (Standard version 1.6, Olympus).
4.4. Preparation of Conditioned Medium
The Oc cells (feeder cells) in culture were subcultured every week according to the protocol provided by Riken Bio-Resource Center (https://plant.rtc.riken.jp/resource/cell_line/web_documents/cell_lines/rpc00031.html, 15 November 2023). The Oc cells at 4 or 6 days after subculture were weighed (0.2–0.6 g) after rinsing them with 2,4-D-free MS medium and removing the medium using pipettes. Subsequently, the cells were suspended in 10 mL of 2,4-D-free N6Z medium and cultured overnight in 6 cm diameter Petri dishes at 26 °C in darkness with rotation at 40 rpm. The cultured medium was roughly separated from feeder cells and filtered through a filter (0.45 µm pore size, Millex-HV Filter Unit). The filtrate was used as the conditioned medium.
4.5. Preparation of Heat-Treated Conditioned Medium
The conditioned medium was dispensed into two glass vials of 5 mL each. One aliquot was placed in boiling water at 100 °C for 15 min and then cooled to room temperature to be used as the heat-treated conditioned medium. Another aliquot of the conditioned medium was left at room temperature while preparing the heat-treated medium and used as the non-heat-treated conditioned medium.
4.6. Isolation of Protein Samples from the Conditioned Medium for Proteome Analysis
The conditioned medium with or without heat treatment was centrifuged at 15,000× g for 15 min, and 300 µL of methanol was added to 100 µL of the supernatant. After centrifugation at 15,000× g for 3 min, the precipitate was dried under vacuum and used as the protein fraction from the conditioned medium. The dried precipitate dissolved in 20 µL sample buffer solution (2-mercaptoethanol+; FUJIFILM Wako Chemicals, Oosaka, Japan) was subjected to 100 °C for 5 min and applied to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE).
4.7. SDS-PAGE and Silver Staining
According to Laemmli (1970) [43], 10–20% SDS-PAGE gels were prepared in a small mold (e-PAGEL HR her-T1020L, 90 mm × 83 mm × 1 mm; Atto, Tokyo, Japan), and 20 µL protein sample solutions were applied. After electrophoresis, proteins in the gels were detected using conventional silver staining [44]. When used for subsequent liquid chromatography coupled with tandem mass spectroscopy (LC-MS/MS) analyses, the proteins in the gel were visualized with modified silver staining using Silver Stain MS Kit (FUJIFILM Wako Chemicals).
4.8. Identification of Proteins by Tandem Mass Spectrometry
SDS-PAGE gels were cut into five pieces. Proteins in each piece were in-gel-digested with trypsin [45] and identified with LC-MS/MS analyses using a direct nanoflow LC-MS system equipped with an Orbitrap mass spectrometer (Q Exactive; Thermo Scientific, Waltham, MA, USA), as described elsewhere [46]. The dataset of protein sequences obtained from the Rice Annotation Project Database (Tsukuba, Japan; http://rapdb.dna.affrc.go.jp/download/irgsp1.html, 19 November 2019) was searched using Mascot software (ver. 2.3.2; Matrix Science, Boston, MA, USA) with the following parameters. The variable modification parameters were pyro-Glu, acetylation (protein N-terminus), and oxidation (Met). The maximum missed cleavage was set at 3 with a peptide mass tolerance of ±15 ppm. Peptide charges from +2 to +4 states and MS/MS tolerances of ±0.8 Da were allowed. The criteria for peptide identification were based on the vendor’s definitions (expectation value < 0.05; Matrix Science), and we assigned the protein as “identified” when at least two peptides were identified from the protein.
Author Contributions
Y.W. and T.O. designed the experiments; Y.W. performed most of the experiments; M.T. and Y.N. performed the proteomic analysis; Y.W. and T.O. conceived the project and wrote the article. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported in part by JSPS KAKENHI (Grant-in-Aid for Scientific Research(B), Grant No. 22H02315 to T.O.), NEDO Moonshot Program (Grant No. 22101490-0 to T.O.), JST-Mirai Program (Grant No. 23-231038606 to TO) and by the Joint Research Program of Arid Land Research Center, Tottori University (Grant No. 04B2020 to T.O.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
Acknowledgments
We thank T. Mochizuki (Tokyo Metropolitan University) for isolating rice egg cells and the RIKEN Bio Resource Center (Tsukuba, Japan) for providing cultured rice cells (Oc line).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pritchard, H.N. A cytochemical study of embryo development in Stellaria medis. Botany 1964, 51, 472–479. [Google Scholar] [CrossRef]
- Schulz, R.; Jensen, W.A. Capsella embryogenesis: The egg, zygote, and young embryo. Am. J. Bot. 1968, 55, 807–819. [Google Scholar] [CrossRef]
- Tykarska, T. Rape embryogenesis: I. The proembryo development. Acta Soc. Bot. Pol. 1976, 45, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Schel, J.H.N.; Kieft, H.; van Lammeren, A.A.M. Interactions between embryo and endosperm during early developmental stage of maize caryopses (Zea mays). Can. J. Bot. 1984, 62, 2842–2853. [Google Scholar] [CrossRef]
- Lindsey, K.; Topping, J.E. Embryogenesis: A question of pattern. J. Exp. Bot. 1993, 259, 359–374. [Google Scholar] [CrossRef]
- Itoh, J.; Nonomura, K.; Ikeda, K.; Yamaki, S.; Inukai, Y.; Yamagishi, H.; Kitano, H.; Nagato, Y. Rice Plant development: From zygote to spikelet. Plant Cell Physiol. 2005, 46, 23–47. [Google Scholar] [CrossRef] [PubMed]
- Sato, A.; Toyooka, K.; Okamoto, T. Asymmetric cell division of rice zygotes located in embryo sac and produced by in vitro fertilization. Sex Plant Reprod. 2010, 23, 211–217. [Google Scholar] [CrossRef]
- Kranz, E.; Bautor, J.; Lörz, H. In vitro fertilization of single, isolated gametes of maize mediated by electrofusion. Sex. Plant Reprod. 1991, 4, 12–16. [Google Scholar] [CrossRef]
- Okamoto, T. Gamete fusion site on the egg cell and autonomous establishment of cell polarity in the zygote. Plant Signal. Behav. 2010, 5, 1464–1467. [Google Scholar] [CrossRef]
- Uchiumi, T.; Uemura, I.; Okamoto, T. Establishment of an in vitro fertilization system in rice (Oryza sativa L.). Planta 2007, 226, 581–589. [Google Scholar] [CrossRef]
- Kranz, E.; Lörz, H. In vitro fertilization with isolated, single gametes results in zygotic embryogenesis and fertile maize plants. Plant Cell 1993, 5, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Kumlehn, J.; Lörz, H.; Kranz, E. Differentiation of isolated wheat zygotes into embryos and normal plants. Planta 1998, 205, 327–333. [Google Scholar] [CrossRef]
- Maryenti, T.; Kato, N.; Ichikawa, M.; Okamoto, T. Establishment of an in vitro fertilization system in wheat (Triticum aestivum L.). Plant Cell Physiol. 2019, 60, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Moloney, M.M.; Hall, J.F.; Robinson, G.M.; Elliott, M.C. Auxin Requirements of Sycamore Cells in Suspension Culture. Plant Physiol. 1983, 71, 927–931. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Bird, S.; Kemble, R.; Simmonds, D.; Keller, W.; Miki, B. Effects of culture density, conditioned medium and feeder cultures on microspore embryogenesis in Brassica napus L. cv. Topas. Plant Cell Rep. 1990, 8, 594–597. [Google Scholar] [CrossRef] [PubMed]
- Folling, M.; Madsen, S.; Olesen, A. Effect of nurse culture and conditioned medium on colony formation and plant regeneration from Lolium perenne protoplasts. Plant Sci. 1995, 108, 229–239. [Google Scholar] [CrossRef]
- Campanoni, P.; Nick, P. Auxin-dependent cell division and cell elongation. 1-naphthaleneacetic acid and 2,4-dichlorophenoxyacetic acid activate different pathways. Plant Physiol. 2005, 137, 939–948. [Google Scholar] [CrossRef]
- Woójcik, A.M.; Woójcikowska, B.; Gaj, M.D. Current Perspectives on the Auxin-Mediated Genetic Network that Controls the Induction of Somatic Embryogenesis in Plants. Int. J. Mol. Sci. 2020, 21, 1333. [Google Scholar] [CrossRef]
- Imbrie-Milligan, C.W.; Hodges, T.K. Microcallus formation from maize protoplasts prepared from embryogenic callus. Planta 1986, 168, 395–401. [Google Scholar] [CrossRef]
- Jain, R.K.; Khehra, G.S.; Lee, S.-H.; Blackhal, N.W.; Marehant, L.R.; Davey, M.R.; Power, J.B.; Cocking, E.C.; Gosal, S.S. An improved procedure for plant regeneration from indica and japonica rice protoplasts. Plant Cell Rep. 1995, 14, 515–519. [Google Scholar] [CrossRef]
- Matthys-Rochon, E. Secreted molecules and their role in embryo formation in plants: A mini-review. Acta Biol. Crac. Ser. Bot. 2005, 47, 23–29. Available online: https://hal.science/hal-00188834 (accessed on 4 January 2023).
- Van Hengel, A.J.; Tadesse, Z.; Immerzeel, P.; Schols, H.; Van Kammen, A.; de Vries, S.C. N-acetylglucosamine and glucosamine-containing arabinogalactan proteins control somatic embryogenesis. Plant Physiol. 2001, 125, 1880–1890. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, Y.; Hoshino, R.; Okamoto, T. Dynamics of male and female chromatin during karyogamy in rice zygotes. Plant Physiol. 2014, 165, 1533–1543. [Google Scholar] [CrossRef]
- Toda, E.; Ohnishi, Y.; Okamoto, T. Development of polyspermic rice zygotes. Plant Physiol. 2016, 171, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Deushi, R.; Toda, E.; Koshimizu, S.; Yano, K.; Okamoto, T. Effect of paternal genome excess on the developmental and gene expression profiles of polyspermic zygotes in rice. Plants 2021, 10, 255. [Google Scholar] [CrossRef]
- Kitahara, R.; Akasaka, K. Close identity of a pressure-stabilized intermediate with a kinetic intermediate in protein folding. Proc. Natl. Acad. Sci. USA 2003, 100, 3167–3172. [Google Scholar] [CrossRef]
- Kitahara, R.; Yokoyama, S.; Akasaka, K. NMR snapshots of a fluctuating protein structure: Ubiquitin at 30 bar-3 kbar. J. Mol. Biol. 2005, 347, 277–285. [Google Scholar] [CrossRef]
- Rakleova, G.; Keightley, A.; Pantchev, I.; Tsacheva, I.; Tchorbadjieva, M. Identification, molecular cloning, and recombinant gene expression of an extracellular a-amylase from Dactylis Glomerata L. embryogenic suspension cultures. Biotech. Biotechnol. Eq. 2012, 26, 3192–3200. [Google Scholar] [CrossRef]
- Tchorbadjieva, M.I.; Kalmukova, R.I.; Pantchev, I.Y.; Kyurkchiev, S.D. Monoclonal antibody against a cell wall marker protein for embryogenic potential of Dactylis glomerata L. suspension cultures. Planta 2005, 222, 811–819. [Google Scholar] [CrossRef]
- Pernis, M.; Salaj, T.; Bellová, J.; Danchenko, M.; Baráth, P.; Klubicová, K. Secretome analysis revealed that cell wall remodeling and starch catabolism underlie the early stages of somatic embryogenesis in Pinus nigra. Front. Plant Sci. 2023, 14, 1225424. [Google Scholar] [CrossRef]
- De Jong, A.J.; Cordewener, J.; Lo Schiavo, F.; Terzi, M.; Vandekerckhove, J.; Van Kammen, A.; De Vries, S.C. A carrot somatic embryo mutant is rescued by chitinase. Plant Cell 1992, 4, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Helleboid, S.; Bauw, G.; Belingheri, L.; Vasseur, J.; Hilbert, J.L. Extracellular b-1,3-glucanases are induced during early somatic embryogenesis in Cichorium. Planta 1998, 205, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Pan, C.H.; So, M.Y.; Ah, J.H.; Jo, D.H.; Kim, S.I. Purification, characterization, and cDNA cloning of rice class III chitinase. Mol. Cells 2002, 3, 69–76. [Google Scholar]
- De Jong, A.J.; Heidstra, R.; Spaink, H.P.; Hartog, M.V.; Meijer, E.A.; Hendriks, T.; Lo Schiavo, F.; Terzi, M.; Bisseling, T.; Van Kammen, A.; et al. Rhizobium lipooligosaccharides rescue a carrot somatic embryo mutant. Plant Cell 1993, 5, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Quiroz-Figueroa, F.R.; Rojas-Herrera, R.; Galaz-Avalos, R.M.; Loyola-Vargas, V.M. Embryo production through somatic embryogenesis can be used to study cell differentiation in plants. Plant Cell Tissue Organ Cult. 2006, 86, 285–301. [Google Scholar] [CrossRef]
- Ma, T.; Dong, F.; Luan, D.; Hu, H.; Zhao, J. Gene expression and localization of arabinogalactan proteins during the development of anther, ovule, and embryo in rice. Protoplasma 2019, 256, 909–922. [Google Scholar] [CrossRef]
- Qin, Y.; Zhao, J. Localization of arabinogalactan proteins in egg cells, zygotes, and two-celled proembryos and effects of b-D-glucosyl Yariv reagent on egg cell fertilization and zygote division in Nicotiana tabacum L. J. Exp. Bot. 2006, 57, 2061–2074. [Google Scholar] [CrossRef]
- Kranz, E.; von Wiegen, P.; Lörz, H. Early cytological events after induction of cell division in egg cells and zygote development following in vitro fertilization with angiosperm gametes. Plant J. 1995, 8, 9–23. [Google Scholar] [CrossRef]
- Ohnishi, Y.; Kokubu, I.; Kinoshita, T.; Okamoto, T. Sperm Entry into the Egg Cell Induces the Progression of Karyogamy in Rice Zygotes. Plant Cell Physiol. 2019, 60, 1656–1665. [Google Scholar] [CrossRef]
- He, Y.C.; He, Y.Q.; Qu, L.H.; Sun, M.X.; Yang, H.Y. Tobacco zygotic embryogenesis in vitro: The original cell wall of the zygote is essential for maintenance of cell polarity, the apical–basal axis and typical suspensor formation. Plant J. 2007, 49, 515–527. [Google Scholar] [CrossRef]
- Abiko, M.; Furuta, K.; Yamauchi, Y.; Fujita, C.; Taoka, M.; Isobe, T.; Okamoto, T. Identification of proteins enriched in rice egg or sperm cells by single-cell proteomics. PLoS ONE 2013, 8, e69578. [Google Scholar] [CrossRef]
- Uchiumi, T.; Komatsu, S.; Koshiba, T.; Okamoto, T. Isolation of gametes and central cells from Oryza sativa L. Sex. Plant Reprod. 2006, 19, 37–45. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Oakley, B.R.; Kirsch, D.R.; Morris, N.R. A simplified ultrasensitive silver stain for detecting proteins in polyacrylamide gels. Anal. Biochem. 1980, 105, 361–363. [Google Scholar] [CrossRef]
- Taoka, M.; Ichimura, T.; Wakamiya-Tsuruta, A.; Kubota, Y.; Araki, T.; Obinata, T.; Isobe, T. V-1, a protein expressed transiently during murine cerebellar development, regulates actin polymerization via interaction with capping protein. J. Biol. Chem. 2003, 278, 5864–5870. [Google Scholar] [CrossRef] [PubMed]
- Taoka, M.; Morofuji, N.; Yamauchi, Y.; Ojima, H.; Kubota, D.; Terukina, G. Global PROTOMAP profiling to search for biomarkers of early-recurrent hepatocellular carcinoma. J. Proteome Res. 2014, 13, 4847–4858. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Developmental profiles of zygotes cultured in N6Z medium (A) and 2,4-D free N6Z medium (B,C). (A) Developmental profiles of the zygotes cultured in N6Z medium. After completion of karyogamy in zygote (Aa,Ab), the zygote developed into a two-celled embryo (Ac,Ad), multicellular embryo (Ae,Af), a globular-like embryo (Ag,Ah), and cell mass (Ai–An). (B) Developmental profiles of the zygotes cultured in 2,4-D free N6Z medium. After completion of karyogamy in zygote (Ba,Bb), the zygote developed into a two-celled embryo (Bc,Bd), multicellular embryo (Be,Bf), a globular-like embryo (Bg,Bh), and cell mass (Bi–Bn). (C) Developmentally arrested embryos cultured in 2,4-D free N6Z medium. Upper and lower panels are fluorescent and bright-field images, respectively. Bars, 20 µm.
Figure 1.
Developmental profiles of zygotes cultured in N6Z medium (A) and 2,4-D free N6Z medium (B,C). (A) Developmental profiles of the zygotes cultured in N6Z medium. After completion of karyogamy in zygote (Aa,Ab), the zygote developed into a two-celled embryo (Ac,Ad), multicellular embryo (Ae,Af), a globular-like embryo (Ag,Ah), and cell mass (Ai–An). (B) Developmental profiles of the zygotes cultured in 2,4-D free N6Z medium. After completion of karyogamy in zygote (Ba,Bb), the zygote developed into a two-celled embryo (Bc,Bd), multicellular embryo (Be,Bf), a globular-like embryo (Bg,Bh), and cell mass (Bi–Bn). (C) Developmentally arrested embryos cultured in 2,4-D free N6Z medium. Upper and lower panels are fluorescent and bright-field images, respectively. Bars, 20 µm.

Figure 2.
Developmental profiles of zygotes cultured in feeder cell free N6Z medium. Zygotes cultured in feeder-cell free N6Z medium (A–C) and in feeder cell and 2,4-D free N6Z medium (D–F) were observed 1 day after fusion. Left and right panels are fluorescent and bright-field images, respectively. Bars, 20 µm.
Figure 2.
Developmental profiles of zygotes cultured in feeder cell free N6Z medium. Zygotes cultured in feeder-cell free N6Z medium (A–C) and in feeder cell and 2,4-D free N6Z medium (D–F) were observed 1 day after fusion. Left and right panels are fluorescent and bright-field images, respectively. Bars, 20 µm.

Figure 3.
Developmental profiles of the zygotes cultured in conditioned medium. (A) After completion of karyogamy in zygote (Aa–Ac), the zygote developed into a two-celled embryo (Ad–Af), multicellular embryo (Ag–Ai), a globular-like embryo (Aj–Ao), and cell mass (Ap–Ar). Bars, 20 µm (Aa–Ao), 100 µm (Ap–Ar). (B) The zygote arrested development. The zygote developed into a multicellular embryo (Ba–Bc) 3 days after fusion and arrested development thereafter (Bd–Bi). Upper, lower, and middle panels are fluorescent, bright-field, and merged images, respectively. Bars, 20 µm.
Figure 3.
Developmental profiles of the zygotes cultured in conditioned medium. (A) After completion of karyogamy in zygote (Aa–Ac), the zygote developed into a two-celled embryo (Ad–Af), multicellular embryo (Ag–Ai), a globular-like embryo (Aj–Ao), and cell mass (Ap–Ar). Bars, 20 µm (Aa–Ao), 100 µm (Ap–Ar). (B) The zygote arrested development. The zygote developed into a multicellular embryo (Ba–Bc) 3 days after fusion and arrested development thereafter (Bd–Bi). Upper, lower, and middle panels are fluorescent, bright-field, and merged images, respectively. Bars, 20 µm.

Figure 4.
Developmental profiles of the zygotes cultured in conditioned medium without heat treatment (A–C) and in heat-treated conditioned medium (D). After completion of karyogamy in zygote cultured in non-treated conditioned medium (Aa–Ac), the zygote developed into a two-celled embryos (Bc–Bc) and multicellular embryos (Ca–Cc). In case of zygotes cultured in heat-treated conditioned medium, zygotes were degenerated after completion of karyogamy (Da–Dc). Left, right, and middle panels are fluorescent, bright-field, and merged images, respectively. Bars, 20 µm.
Figure 4.
Developmental profiles of the zygotes cultured in conditioned medium without heat treatment (A–C) and in heat-treated conditioned medium (D). After completion of karyogamy in zygote cultured in non-treated conditioned medium (Aa–Ac), the zygote developed into a two-celled embryos (Bc–Bc) and multicellular embryos (Ca–Cc). In case of zygotes cultured in heat-treated conditioned medium, zygotes were degenerated after completion of karyogamy (Da–Dc). Left, right, and middle panels are fluorescent, bright-field, and merged images, respectively. Bars, 20 µm.

Figure 5.
Effect of exogeneously applied α-amylase on the development of rice zygotes. Zygotes were cultured in 2,4-D free N6Z medium containing α-amylase at the concentration of 0.17 U/mL (A,B), 1.7 U/mL (C,D), and 17 U/mL (E,F). Left, right, and middle panels are fluorescent, bright-field, and merged images, respectively. Bars, 20 µm.
Figure 5.
Effect of exogeneously applied α-amylase on the development of rice zygotes. Zygotes were cultured in 2,4-D free N6Z medium containing α-amylase at the concentration of 0.17 U/mL (A,B), 1.7 U/mL (C,D), and 17 U/mL (E,F). Left, right, and middle panels are fluorescent, bright-field, and merged images, respectively. Bars, 20 µm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Effects of 2,4-D and feeder cells on developmental profiles of rice zygotes.
| Medium | No. of Zygotes Cultured | No. of Zygotes That Developed to Specific Growth Stages | |||||
|---|---|---|---|---|---|---|---|
| Feeder Cell | 2,4-D | Karyogamy | Two-Celled Embryo | Multicellular Embryo | Globular-LikeEmbryo | Cell Mass | |
| + | + | 128 | 105 | 98 | 97 | 88 | 74 |
| + | − | 73 | 53 | 47 | 43 | 31 | 28 |
| − | + | 103 | 19 | 5 | 0 | 0 | 0 |
| − | − | 83 | 27 | 5 | 0 | 0 | 0 |
| Conditioned medium | 104 | 8 | 72 | 51 | 12 | 8 | |
Table 2.
Developmental profiles of zygotes cultured in conditioned medium pretreated with or without heat.
Table 2.
Developmental profiles of zygotes cultured in conditioned medium pretreated with or without heat.
| Heat Treatment a | No. of Zygotes Cultured | No. of Zygotes That Developed to Specific Growth Stages | ||
|---|---|---|---|---|
| Karyogamy | Two-Celled Embryo | Multicellular Embryo | ||
| − | 39 | 24 | 11 | 3 |
| + | 37 | 16 | 0 | 0 |
a Conditioned medium with or without heat-treatment at 100 °C for 15 min were used for zygote culture.
Table 3.
Proteins detected both in non-heat and heated conditioned medium with >20 spectra.
| Gene Locus | Protein a | Number of Identified Spectra b | |
|---|---|---|---|
| Non-Heat Conditioned Medium | Heated Conditioned Medium | ||
| Os06g0681400 | Ubiquitin domain-containing protein. | 368 | 310 |
| Os02g0161900 | Similar to polyubiquitin containing seven ubiquitin monomers. | 317 | 268 |
| Os04g0628100 | Similar to polyubiquitin. | 250 | 207 |
| Os06g0673500 | Similar to polyubiquitin containing seven ubiquitin monomers. | 93 | 79 |
| Os03g0770775 | Hypothetical protein. | 61 | 78 |
| Os01g0328400 | Ubiquitin. | 62 | 58 |
| Os05g0160200 | Ubiquitin. | 62 | 58 |
| Os06g0650100 | Similar to the ubiquitin-NEDD8-like protein RUB2. | 62 | 58 |
| Os07g0489500 | Ubiquitin domain-containing protein. | 62 | 58 |
| Os09g0420800 | Similar to ubiquitin. | 62 | 58 |
| Os09g0452700 | Ubiquitin. | 62 | 58 |
| Os01g0687400 | Similar to chitinase. | 157 | 47 |
| Os03g0429000 | Proteinase inhibitor I25, cystatin domain-containing protein. | 45 | 41 |
| Os10g0359200 | Similar to UPF0496 protein 1. | 30 | 36 |
| Os01g0853000 | Conserved hypothetical protein. | 29 | 35 |
| Os02g0525900 | Similar to acetyl-coenzyme A synthetase 2 (acetate–CoA ligase 2) (acyl-activating enzyme 2). | 43 | 29 |
| Os11g0427700 | Xanthine/uracil/vitamin C permease family protein. | 25 | 29 |
| Os01g0582400 | Similar to multidomain cyclophilin type peptidyl-prolyl cis-trans isomerase. | 28 | 26 |
| Os08g0434100 | Similar to S-like ribonuclease (RNase PD2) (fragment). | 34 | 22 |
| Os06g0208800 | Lysin motif-containing protein, pattern recognition receptor, roles in peptidoglycan and chitin perception in innate immunity | 36 | 21 |
| Os03g0786100 | Similar to glycolate oxidase (fragment). | 24 | 20 |
a Protein annotations were obtained from The Rice Annotation Project Database (RAP-DB, Tsukuba, Japan http://rapdb.dna.affrc.go.jp/download/irgsp1.html, 19 November 2019). b The number of spectra is the sum of analyses performed twice. Abbreviations: RUB2, Related to Ubiquitin 2; NEDD8, neural precursor cell-expressed, developmentally downregulated 8.
Table 4.
Proteins that were preferentially detected from the non-heat-conditioned medium with >20 spectra.
Table 4.
Proteins that were preferentially detected from the non-heat-conditioned medium with >20 spectra.
| Gene Locus | Protein a | Number of Identified Spectra b | Signal Peptide c | |
|---|---|---|---|---|
| Non-Heat-Conditioned Medium | Heated Conditioned Medium | |||
| Os08g0473900 | Similar to alpha amylase isozyme 3D. | 531 | 2 | + |
| Os08g0473600 | Alpha-amylase isozyme 3E precursor. | 259 | 0 | + |
| Os09g0457800 | Alpha-amylase isozyme 3C precursor. | 135 | 0 | + |
| Os09g0457600 | Alpha-amylase isozyme 3B precursor. | 131 | 0 | + |
| Os04g0574200 | FAS1 domain-containing protein | 130 | 13 | + |
| Os06g0104300 | Similar to pectinesterase-like protein. | 107 | 0 | + |
| Os03g0761500 | Similar to subtilisin-like protease (fragment). | 73 | 0 | + |
| Os03g0603600 | PLC-like phosphodiesterase, TIM beta/alpha-barrel domain-containing protein. | 66 | 0 | + |
| Os02g0121300 | Cyclophilin, peptidyl-prolyl cis-trans isomerase, auxin signal transduction, lateral root initiation, stress tolerance | 48 | 0 | − |
| Os11g0525600 | Similar to alpha-mannosidase. | 46 | 0 | + |
| Os01g0739700 | Glycoside hydrolase, family 17 protein. | 43 | 0 | + |
| Os10g0493600 | Alpha-galactosidase precursor. | 43 | 0 | + |
| Os02g0765400 | Similar to alpha-amylase. | 36 | 0 | + |
| Os02g0765600 | Alpha-amylase glycoprotein, degradation of starch granule | 36 | 0 | + |
| Os06g0546500 | Similar to class III peroxidase GvPx2b (fragment). | 32 | 0 | + |
| Os02g0139300 | Glycoside hydrolase, family 17 protein. | 28 | 0 | + |
| Os10g0416100 | Class III chitinase RCB4. | 26 | 0 | + |
| Os03g0703100 | Similar to beta-glucosidase. | 25 | 0 | + |
| Os03g0327600 | Ricin B-related lectin domain-containing protein. | 24 | 0 | − |
| Os03g0285700 | Similar to L-ascorbate peroxidase. | 21 | 1 | − |
| Os09g0548200 | Peptidoglycan-binding lysin subgroup domain-containing protein. | 21 | 2 | + |
a Protein annotations were obtained from The Rice Annotation Project Database (RAP-DB, Tsukuba, Japan http://rapdb.dna.affrc.go.jp/download/irgsp1.html, 19 November 2019). b The number of spectra is the sum of analyses performed twice. c Existence of signal peptides for protein secretion on the identified proteins was estimated using TargetP-2.0 (https://services.healthtech.dtu.dk/services/TargetP-2.0/, 24 July 2023). Abbreviations: FAS1, fasciclin-1; PLC, phospholipase C; TIM, triose-phosphate isomerase.
Table 5.
Effects of exogenously applied α-amylase on developmental profiles of rice zygotes.
| α-Amylase Concentration (Unit/mL) | No. of Zygotes Produced | No. of Zygotes That Developed to Specific Growth Stages | |
|---|---|---|---|
| Karyogamy | Two-Celled Embryo | ||
| 0 | 13 | 3 | 0 |
| 0.17 | 7 | 2 | 0 |
| 1.7 | 18 | 12 | 0 |
| 17 | 21 | 5 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Watanabe, Y.; Nobe, Y.; Taoka, M.; Okamoto, T. The Feeder Effects of Cultured Rice Cells on the Early Development of Rice Zygotes. Int. J. Mol. Sci. 2023, 24, 16541. https://doi.org/10.3390/ijms242216541
AMA Style
Watanabe Y, Nobe Y, Taoka M, Okamoto T. The Feeder Effects of Cultured Rice Cells on the Early Development of Rice Zygotes. International Journal of Molecular Sciences. 2023; 24(22):16541. https://doi.org/10.3390/ijms242216541
Chicago/Turabian StyleWatanabe, Yoriko, Yuko Nobe, Masato Taoka, and Takashi Okamoto. 2023. "The Feeder Effects of Cultured Rice Cells on the Early Development of Rice Zygotes" International Journal of Molecular Sciences 24, no. 22: 16541. https://doi.org/10.3390/ijms242216541
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.