
Natural Inhibitors of P-glycoprotein in Acute Myeloid Leukemia

Department of Biological, Chemical and Pharmaceutical Science and Technology (STEBICEF), University of Palermo, 90128 Palermo, Italy
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(4), 4140; https://doi.org/10.3390/ijms24044140
Submission received: 22 December 2022
/
Revised: 15 February 2023
/
Accepted: 16 February 2023
/
Published: 18 February 2023
(This article belongs to the Special Issue Latest Review Papers in Molecular Pathology, Diagnostics, and Therapeutics 2023)
Abstract
:Acute myeloid leukemia (AML) remains an insidious neoplasm due to the percentage of patients who develop resistance to both classic chemotherapy and emerging drugs. Multidrug resistance (MDR) is a complex process determined by multiple mechanisms, and it is often caused by the overexpression of efflux pumps, the most important of which is P-glycoprotein (P-gp). This mini-review aims to examine the advantages of using natural substances as P-gp inhibitors, focusing on four molecules: phytol, curcumin, lupeol, and heptacosane, and their mechanism of action in AML.
1. Introduction
The new therapeutic strategies available today for cancer treatment include targeted therapies, which have significantly increased patient survival [1]. Despite the current and emerging therapeutic landscape for the treatment of cancer and an improved understanding of mechanisms of resistance to conventional therapies [2], primary and secondary drug resistance remains a substantial problem for most patients. Acquired resistance is often determined by the overexpression of efflux pumps and develops rapidly. Acute myeloid leukemia is no exception in this sense because a subgroup of patients often develops resistance and becomes refractory to all available therapies. Among the efflux pumps, P-glycoprotein (P-gp) is certainly the most studied as a pharmacological target. However, the inhibitors tested so far have shown toxicity incompatible with use in the clinic [3,4,5]; therefore, especially recently, the attention has shifted towards natural molecules, characterized by low toxicity. These substances can act specifically on the overexpression of the pump, and while they do not show toxicity on normal cells, their effects on the physiological function of the P-gp are unknown. In this case, targeting a specific action on the P-gp, expressed by the neoplastic cell, could represent a good strategy.
In particular, we present the mechanism of action of four natural substances belonging to different categories: terpenoids, polyphenols, and hydrocarbons. Phytol is acyclic hydrogenated diterpene alcohol, lupeol is a pentacyclic triterpenoid, curcumin is a polyphenol and heptacosane is a straight-chain alkane with 27 carbon atoms. All these compounds are capable of targeting P-gp by inhibiting its expression at the transcriptional level and/or by acting as substrate inhibitors of the same protein, yet they do so through different mechanisms.
2. Acute Myeloid Leukemia
Acute myeloid leukemia is the most common acute form of leukemia, the onset of which is related to age, with an incidence of 3–4 cases per 100,000 people a year. It is a heterogeneous neoplastic disease, characterized by the acquisition of a broad spectrum of molecular alterations, with a higher frequency of occurrence in the elderly population (60–65 years) [6]. However, a good percentage also occurs in pediatric ages and in young people (15–20% of leukemias occur in children and approximately 33% in adolescents and young adults), with a 5-year overall survival (OS) rate over the last 20 years of 60–75% in children and only 50–60% in young adults (15–39 years) [7]. Regarding the therapeutic possibilities, the standard “7 + 3” chemotherapy (continuous infusion of cytarabine over 7 days with the addition of an anthracycline, given daily for the first 3 days) often has a weak outcome, with only about 40% complete remission rate and a median OS of 12–18 months. Contextually, knowledge of the great molecular heterogeneity and genetic diversity of AML has led to the discovery of several new promising therapies for the treatment of this disease. Recently, new therapeutic agents have been approved by the FDA: CPX 351, a liposomal formulation of fixed-dose cytarabine and daunorubicin; venetoclax, an oral highly selective inhibitor of the anti-apoptotic protein, BCL-2, in combination with a hypomethylating agent, decitabine or azacitidine; FLT3 (Fms-like tyrosine kinase 3) inhibitors, midostaurin, and gilteritinib, either alone or in combination with conventional chemotherapies; IDH1/2 (isocitrate dehydrogenase 1 and 2) inhibitors, ivosidenib, and enasidenib; the conjugated anti-CD33-antibody, gemtuzumab ozogamicin; glasdegib, a selective inhibitor of hedgehog signaling [8,9,10]. These new targeted therapies, compared to standard chemotherapy, have improved response rates and outcomes for selected AML patients in different clinical scenarios. Furthermore, sorafenib, a multi-kinase inhibitor of RAF, KIT, and FLT3 kinases could represent a useful drug in AML. Recently, it has been evaluated in a phase II trial (SORMAIN) as a maintenance therapy and in an open-label randomized phase III trial in adult patients with FLT3-ITD (internal duplications in tandem) mutated AML, who had already undergone allogeneic hematopoietic stem cell transplantations. Both studies showed a good survival benefit for AML patients [11,12].
Many types of cancer express ABC transporters, such as colorectal cancer, liver cancer, acute myeloid leukemia (AML), and breast cancer. In approximately 35% of AML patients, the overexpression of P-gp causes the failure of anticancer therapies. Indeed, P-g expression has been detected in approximately one-third of patients with AML at diagnosis and in more than half of the patients in remission. Furthermore, higher levels have also been observed in certain subtypes, including secondary leukemia. In AML, P-gp expression is associated with a lower rate of complete remission, disease-free survival, and overall survival [13,14,15].
Unfortunately, even with the most innovative therapies, we are still observing the development of drug resistance, which remains one of the key events affecting therapeutic failure. In fact, there are refractory or resistant forms, in which the targeted therapies can be made ineffective by genetic mutations. Multidrug resistance (MDR) concerns most AML patients, which can develop promptly, and in these cases, allogeneic stem cell transplantation becomes the only choice. The research of molecules able to overcome this condition is therefore urgent.
3. The Multidrug Resistance Mediated by Efflux Pumps
The phenomenon of multidrug resistance is complex and can spring from different mechanisms, such as the overexpression of members of the adenosine triphosphate binding cassette (ABC) transporters, which includes P-gp, a multidrug resistance-1 protein-coding gene (MDR1 or ABCB1), the multidrug resistance associated protein (MRP1 or ABCC1), and the breast cancer resistance protein (BCRP or ABCG2). These transporters are implied in the genesis of the MDR phenotype in various tumor models [13].
To date, the most studied strategy to limit MDR is by blocking the efflux of the chemotherapy drug through the inhibition and modulation of the efflux pump functions, by either chemosensitizing, revertant inhibitors of MDR, or modulators of P-gp. As mentioned, the main pharmacological strategy of preclinical and clinical research, to overcome MDR and revert the resistant phenotype, is represented by the co-administration of an anticancer chemotherapy drug and P-gp substrate-inhibitor compounds, to restore the effective therapeutic concentration of the drug in the tumor-resistant cells. Among the many compounds tested, a classification was made over time between first-, second-, and third-generation inhibitors. Moreover, these compounds have different mechanisms of action since some act as competitive antagonist substrates, others as modulators, and some as pure inhibitors. The first-generation modulators include very different molecules as well as calcium channel blockers (e.g., verapamil), calmodulin antagonists (trifluoperazine), antiarrhythmic agents, and immunomodulators (e.g., quinidine and cyclosporine A). The second-generation modulators are characterized by greater potency and efficacy, yet remain very toxic; they are often structural derivatives of first-generation compounds such as dexverapamil, the R-enantiomer of verapamil. The third-generation modulators are derived from quantitative structure–activity studies and combinatorial chemistry techniques. These compounds (e.g., laniquidar, ONT-093, zosuquidar, and tariquidar) cause fewer pharmacokinetic and pharmacodynamic interactions because they only minimally interfere with the cytochrome P-450 system. In addition, due to their high affinity for P-gp, they are used at very low concentrations and are, therefore, not very toxic [16]. It is important to consider that P-gp is normally present in healthy cells because it performs many physiological functions, including providing protection against lipophilic and cytotoxic xenobiotic molecules. Inhibition of the physiological function of P-gp increases the danger of intoxication due to increased drug absorption and the reduced excretion of metabolites. However, despite all the progress that has been made in designing strategies to overcome MDR in humans, none of the currently known P-gp inhibitors can be considered an ideal inhibitor, capable of safely and completely reversing MDR in cancer cells [17]. Moreover, none have ever made it to the clinic due to multiple factors, including numerous drug interactions, dose-limiting toxicity, and a lack of specificity for P-gp [3,6,18]. In the case of acquired drug resistance, P-gp is typically expressed in response to chemotherapy and, thus, is induced by the administration of anticancer chemotherapy drugs; however, non-toxic ABC transporter modulators possessing synergistic activity with conventional chemotherapy drugs can be used prophylactically to prevent the incidence of the MDR phenotype.
4. Natural Inhibitors of P-glycoprotein in Leukemia
Many researchers are focusing on products of natural origin that have been shown to be more effective and less toxic in nature, which could provide new sources of novel P-gp inhibitors [19,20,21]. The natural molecules most recently identified as potential inhibitors of P-gp are part of the fourth generation of inhibitors. These compounds such as flavonoids, alkaloids, terpenoids, and saponins [22] are able to perform a double mode of action on P-gp: they can interfere with its transcriptional expression and/or directly inhibit its function. These compounds can be substrates for P-gp and compete with anticancer agents to bind to the active site of the transporter (drug-binding pocket, DBP) and reduce drug efflux. Furthermore, they can modulate the ATPase activity of the pump through interaction with the nucleotide-binding pocket (NBP) and, finally, they can interact with membrane lipids and alter the membrane environment. Fluidity and lipid density are two characteristics of the plasma membrane that can interfere with MDR. It is known how some natural molecules can modulate the activity of the proteins incorporated in the membrane through global perturbations of the lipid environment [23]. This alteration in the membrane fluidity could also influence the correct interaction of different substrates with the efflux systems, such as P-gp. Phenolic phytochemicals are promiscuous modifiers of membrane protein functions, suggesting that some of their actions may be due to a common membrane bilayer-mediated mechanism. The incorporation of pentacyclic triterpenes into the phospholipid film significantly changes its morphology. They are structurally similar to steroids, therefore, analogously to cholesterol and plant sterols, they are easily included in biomembranes, although they exert a more fluidizing effect [24]. P-gp inhibitors of plant origin, belonging to various classes of secondary metabolites, were identified. Several published reviews summarize the most relevant advances in plant-focused P-gp inhibition research in different tumor models [22,25,26,27]. There are many examples of the ability of natural molecules, as fourth-generation inhibitors, to modulate P-gp in leukemia tumor models (Table 1). These studies provide examples of the potential use of natural substances in combination with chemotherapy, as a P-gp inhibitor, to increase the bioavailability of anticancer drugs in leukemia. While it is true that inhibition of the efflux transporter is essential to enhance the activity of natural compounds to reverse MDR, it is also true that non-specific inhibition can produce undesirable adverse effects on other essential cellular functions. Sometimes, inhibition of P-gp leads to excessive accumulation of cytotoxic drugs and decreased excretion rates, which in turn, produces toxicity to normal cell functions. Naturally occurring molecules have structural diversity, which provides a valuable tool in the search for highly targeted P-gp inhibitors. Many naturally occurring P-gp inhibitors have been observed to be very non-specific, although less toxic in nature. However, the use of natural molecules, instead of conventional synthetic molecules due to their structural diversity and non-specific binding to targets, can lead to undesirable pharmacokinetic changes, meaning extensive research is needed to establish the pharmacological characteristics of these molecules [28]. Indeed, there is no shortage of contradictory claims adding to the challenges of using natural products, such as quercetin, which, according to some studies, stimulates P-gp-mediated efflux and increases resistance to anticancer drugs in MDR cells [29,30]. Contrastingly, other studies have shown that quercetin is able to inhibit P-gp and decrease the resistance of anticancer drugs [31]; thus, it is necessary to evaluate all-natural molecules using the same standard methods, while all research needs to be more specific and focused to avoid such contradictions.
5. Four Natural Molecules Active in AML: Our Experience
This review will focus on the potential anticancer properties of specific compounds on which many research groups, including ours, have previously directed their studies: phytol, curcumin, lupeol, and heptacosane. In the present review, we have chosen to report data on four compounds that has exerted inhibitory effects on P-gp through various mechanisms of action. These compounds can inhibit the function of the efflux pump and/or inhibit its expression, yet they are all able to restore the sensitivity of the MDR cancer cells to chemotherapeutic drugs, exhibiting strong MDR reversal activity in an in vitro resistant AML model.
These four molecules have the common characteristic of being present in natural foods, such as phytol, which is in the chlorophylls of vegetables, curcumin is in curry, lupeol is in both vegetables and fruit, and heptacosane is in fruits; moreover, they can be assumed to be nutraceutical substances [50,51,52,53]. The list of therapeutic properties is certainly very long for curcumin and lupeol, whose anti-inflammatory, antioxidant, antimicrobial, antitumor, cardioprotective, antipyretic, analgesic, wound healing, anticonvulsant, antiarthritic, and antidiabetic activities are well known [54,55,56,57], whereas for phytol and heptacosane, the knowledge of those is limited. Furthermore, it is known that the dysfunction of the intestinal microbiome is associated with both the outcome of chemotherapy and adverse events, in many types of neoplastic tumors as well as in AML [58,59]. Many natural compounds are able to modify the response to therapy by modulating the microbiota [60,61,62]. It has been documented, for example, that there is a bidirectional regulation between the curcumin, which implements the intestinal bacterial flora, and itself which can biotransform the curcumin. Furthermore, in studies using AML animal models, curcumin seems to sensitize the response to Ara-C (cytarabine) by regulating the microbiota and controlling the level of cholesterol, which improves chemoresistance [63,64].
The choice of these four compounds, in particular, was made, above all, because they share a common target, whose prominent role in the resistance to antiblastic agents has been studied extensively by our group: NF-κB. Many natural substances, P-gp inhibitors, modulate its expression, through NF-κB, which includes P-gp as a target at the transcriptional level. Here, we present a few studies that have been performed in vitro on cell lines of different tumor types, which have been made resistant to the most common chemotherapeutic substrates of P-gp. S-Adenosylmethionine, a natural compound and a nutritional supplement, in addition to the numerous anticancer actions observed in different tumor models, is able to reverse multi-drug resistance, reducing the overexpression of P-gp by inhibiting the activation of NF-κB in colorectal cancer cell lines [65].
Among the natural polyphenols, the grape seed, procyanidin, seems to affect MDR mediated by P-gp overexpression, by acting on the NF-κB and MAPK/ERK pathways in ovarian cancer cell lines [66]. A similar result was obtained by Sun J. and colleagues who show that the natural compound clitocine is able to reverse P-gp-associated MDR via the downregulation of NF-κB in hepatoma and uterine cancer cell lines [67]. Feroniellin A, a furanocoumarin, reduces the expression of NF-κB and inhibits P-glycoprotein expression in lung cancer cell lines [68]. Moreover, in a breast cancer cell line, Zhu L. et al. demonstrate that Oroxylin A, a natural monoflavonoid, suppresses P-gp expression via the Chk2/P53/NF-κB signaling pathway [69].
With regards to the chosen natural substances, the mechanisms of action on NF-κB for both curcumin and lupeol are well known. However, phytol and heptacosane are major compounds in the essential oil of Euphorbia intisy, while in AML cells they have been shown previously to be able to reverse the MDR induced by P-gp, by acting both on its expression, through the inhibition of NF-κB, and on its efflux function [70].
6. Phytol
One of the most documented actions of phytol is its ability to interfere with the NF-κB pathway. This transcription factor has a preponderant role in all diseases characterized by a chronic inflammatory matrix, and its hyperactivation is found in many neoplastic diseases. Phytol has the ability to reduce inflammation by acting on important mediators, such as proinflammatory cytokines, and on the NF-κB pathway [71]. We have previously demonstrated a strong inhibition of NF-κB DNA-binding activity by phytol in a model of multi-drug resistant AML, characterized by overactivation of this factor [46]. Phytol is a precursor to produce synthetic forms of vitamin E and vitamin K1, and it is a component of many essential oils. There are several pieces of evidence that demonstrate how vitamins K1, K2, and K3 can potentially modulate important cellular responses in metabolic, inflammatory, and tumor diseases, such as free radical species of oxygen (ROS)-induced apoptosis, radical, and oxidative stress, alongside synthesis of metalloproteinases [72,73]. Similarly, essential oils with a high content of phytol have been shown to have therapeutic effects, for example, by showing antiproliferative effects on triple-negative breast cancer cell lines and on AML MDR cells [74,75,76,77,78,79]. In recent years, it has been proven that the phytol molecule is responsible for the numerous therapeutic effects observed [80,81]. In particular, a systematic review of the different properties of phytol underlines how this is characterized by anxiolytic, metabolism-modulating, cytotoxic, antioxidant, autophagy- and apoptosis-inducing, antinociceptive, anti-inflammatory, immune-modulating, and antimicrobial effects [51]. Although Bobe et al. indicates, in a systematic review, that 10 µM is the physiological concentration of phytol, and anything above this, phytol appears to cause toxicity in normal cells and mortality in animals [82]; however, our group did not observe toxicity in two normal-like cell lines, hTERT RPE-1 and 1-7HB2, until a concentration of 100 µg/mL (340 µM) [46]. Among the signal pathways, in which phytol intervenes, NF-κB is redundant. Labbozzetta et al. [46] show that in a study conducted in the AML MDR cell line, HL-60R, which is derived from HL-60 cells by selecting for doxorubicin resistance, phytol produces a strong decrease in the expression of P-gp at 25 µg/mL (84 µM), at both the mRNA and protein levels. This MDR cell line is characterized by overexpression of P-gp, hyperactivation of NF-κB, and pharmacological resistance to some antiblastic drugs, such as doxorubicin [83]. Phytol was able to reduce P-gp expression levels through inhibition of the NF-κB factor. Conversely, although molecular docking analysis showed the ability of phytol to interact with both sites of the P-gp drug-binding pocket (DBP) and the nucleotide-binding pocket (NBP), the doxorubicin accumulation analysis conducted in the HL-60R cell line, demonstrates how phytol does not seem to modulate the function of the efflux pump [46].
7. Curcumin
Curcumin is a phytochemical extracted from the dried rhizomes of Curcuma longa L. and is part of traditional herbal medicines. Much research has been done on curcumin. Its innumerable biological properties are widely known. Even if curcumin has the advantage of not being toxic (doses up to 3600–8000 mg daily for 4 months did not result in evident toxicities, except for mild nausea and diarrhea), unfortunately, it has the great disadvantage of not being very bioavailable after oral administration, with a systemic ineffectiveness. On the contrary, oral application can be useful for therapeutic action at the intestinal level, or when it is applied topically for local actions in tissues, such as skin and oral mucosa [84]. Mirzaei et al. summarize the results of several pharmacokinetic studies on curcumin. Here, it can be seen that oral administration determines the achievement of very low plasma concentrations, while intraperitoneal administration in mice and rats improves this aspect. Oral administration in humans, up to 10–12 mg/kg, determines the achievement of approximately 50/51.2 ng/mL [85]. The different formulations studied over the years have certainly led to an increase in the bioavailability of curcumin [55], as well as all vehicle systems into which curcumin has been incorporated. Tomeh et al. [86] examined all the delivery systems that have been analyzed in recent years to improve the stability and also cellular uptake of curcumin. Undoubtedly a system that transports the substance in a specific tissue or, in the case of liquid neoplasms such as leukemia, towards the tumor cells, could be a winning weapon. Many research groups, including ours, have analyzed many analogs with the aim of finding compounds that are equally or more effective, yet with higher in vivo stability [87,88,89]. Curcumin is endowed with different actions, which may explain its anti-inflammatory and antitumor properties [90]. In particular, it may interfere with both NF-κB activation and Akt kinase signaling. The pathway PI3K (phosphoinositide 3-kinases)/Akt/NF-κB seems to be the target of curcumin, from which its ability to inhibit the expression of P-gp derives [91]. The effects of curcumin on P-gp expression, both at the mRNA and protein levels, are documented in many MDR tumor models [34], although its action on the pump function is more interesting. The action of curcumin on P-gp function, in fact, was demonstrated by studies on rhodamine 123 (Rh123) accumulation and efflux in different tumor models, as well as retinoblastoma, breast cancer, human cervical carcinoma, and chronic myeloid leukemia cell lines [35,92,93,94]. In particular, Sreenivasan et al. [92] showed that curcumin-inhibited verapamil-stimulated ATPase activity of P-gp at higher concentrations and interacted at the substrate binding site of P-gp and not at the nucleotide-binding region. Analysis of the modulation of MDR by curcumin derivatives provided some evidence that confirms an inhibitory action on P-gp, both on its expression, mediated by three natural curcuminoids present in turmeric [95], and on its function, modulating P-gp mediated efflux through synthetic analogs of curcumin [35,96,97,98]. To date, no evidence has been reported on a possible modulation of the P-gp pump function by curcumin or its analogs in AML. In our preliminary study, conducted in an MDR AML cell line HL-60R and using a flow cytometry assay, we observed that curcumin causes an increase in the intracellular accumulation of doxorubicin, similar to the P-gp inhibitor verapamil. This led us to hypothesize that the modulating action of the function of P-gp occurs also in AML (data presented at the 41st National Congress of the Italian Society of Pharmacology, 16–19 November 2022).
8. Lupeol
AlQathama et al. [99] evaluated the antiproliferative and antimigratory abilities of 27 popular herbal infusions, in addition to the reverse P-gp efflux, in many types of tumor cell lines. Among the bioactive phytochemicals contained in these plants, which possess a better activity in terms of reversing MDR, lupeol is also present. Lupeol exhibits cytotoxic effects, particularly in AML cell lines [100,101]. These properties, together with the anti-inflammatory ones, seem to depend on its specific action on the NF-κB pathway, as shown in many studies concerning neoplastic and chronic inflammatory diseases [102,103,104].
Recently, Fontana et al. [44] confirmed this mechanism of action in two AML cell lines, one of which was responsive and the other exhibited MDR. They showed that in the MDR cell line, the strong inhibition of NF-κB activation led to a reduction in P-gp expression. However, the action of lupeol appears to be complex, involving multiple signaling pathways, which leads to the induction of apoptosis and cell growth inhibition.
Lupeol significantly reduces the expression of the Ras oncoprotein and modulates the protein expression of various signaling molecules involved in PKCα (protein kinase C alpha)/ODC (ornithine decarboxylase), PI3K/Akt and MAPKs (mitogen-activated protein kinase) pathways, in addition to strong reductions in the activation of the NF-κB signaling pathway [105,106,107] in some solid tumors. In analysis of a specific action of the P-gp function, in AML cell lines with an MDR phenotype, we observed a minimal increase in the accumulation of its substrate following pretreatment with lupeol. This result leads us to hypothesize that its action is also directed to the pump function, although with lower efficacy than previously observed in the P-gp expression (data presented on 41st National Congress of the Italian Society of Pharmacology, 16–19 November 2022).
Unfortunately, pharmacokinetic studies on lupeol in rats, mainly after oral administration, demonstrated a bioavailability of < 1% [108,109].
Different formulations able to enhance the solubility and oral bioavailability of lupeol, as well as more potent derivatives [45], are needed.
9. Heptacosane
There is little information regarding the biological properties of heptacosane, except those that report actions referable to phytocomplexes, for which it is one of the major constituents [110,111,112,113]. Heptacosane is one of the major components (8%) of the Centaurea baseri essential oil, Köse et al. have previously described its multiple activities, including its cytotoxic properties against MCF-7, PANC-1, A549, and C6 glioma cell lines [114]. Antitumor properties of the Euphorbia intisy essential oil on AML cells were likewise investigated, and the specific actions of its two major components phytol and heptacosane were evaluated [57]. In contrast to phytol, heptacosane does not modulate the NF-κB pathway but reduces the expression levels of some of the NF-κB targets. As suggested by a docking study on P-gp, heptacosane appears to have the most favorable interaction with the drug binding pocket site (DBP), whereby it assumes a conformation that could close the efflux pump. The compound also binds to the nucleotide-binding pocket (NBP) site, similar to verapamil, the classic reference drug used in P-gp functionality studies [115,116].
Heptacosane acts both as an ATP-dependent drug efflux transporter stimulator and as a substrate for P-gp transport. It was shown to stimulate the activity of the P-gp ATPase and concurrently inhibit the efflux function of the P-gp by acting in a competitive manner with another substrate [46].
10. Conclusions
First-, second-, and third-generation P-gp inhibitors have, for different reasons, proven unsuccessful in the clinic. Their main problem is owing to the high toxicity produced through the inhibition of the physiological functionality of P-gp in healthy tissues, in addition to the interference of these inhibitors with the metabolism of numerous substances. In this context, it could be interesting to intervene with low-toxic molecules able to modulate the overexpression of efflux pumps present in cells with the MDR phenotype. Another interesting strategy could be to use the delivery of efflux pump inhibitors, by administering nanoformulations to cells that overexpress them. This approach would allow us to avoid the undesirable effects on the physiological function of P-gp, providing natural inhibitors in nanosystems targeting cancer cells and resistant leukemic stem cells that overexpress efflux pumps, i.e., using this possibility in AML to target CD33 positive cells [117]. Here, it would be possible to specifically eradicate resistant clones.
Several recent papers indicate that natural substances such as phytol, curcumin, lupeol, and heptacosane, acting on P-gp expression and function, could be fully included among the fourth-generation inhibitors as they offer lower toxicity (Figure 1). These molecules deserve to be evaluated in vivo, as they could have implications in the clinic as adjuvants in chemotherapy regimens, especially in the niche category of patients with relapsing and resistant AML.
Author Contributions
Writing—original draft preparation, P.P.; writing—review and editing, M.L.; M.N.; P.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lee, Y.T.; Tan, Y.J.; Oon, C.E. Molecular targeted therapy: Treating cancer with specificity. Eur. J. Pharmacol. 2018, 834, 188–196. [Google Scholar] [CrossRef]
- Haider, T.; Pandey, V.; Banjare, N.; Gupta, P.N.; Soni, V. Drug resistance in cancer: Mechanisms and tackling strategies. Pharmacol. Rep. 2020, 72, 1125–1151. [Google Scholar] [CrossRef] [PubMed]
- Gottesman, M.M.; Fojo, T.; Bates, S.E. Multidrug resistance in cancer: Role of ATP–dependent transporters. Nat. Rev. Cancer 2002, 2, 48–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mollazadeh, S.; Sahebkar, A.; Hadizadeh, F.; Behravan, J.; Arabzadeh, S. Structural and functional aspects of P-glycoprotein and its inhibitors. Life Sci. 2018, 214, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Seelig, A. P-Glycoprotein: One Mechanism, Many Tasks and the Consequences for Pharmacotherapy of Cancers. Front. Oncol. 2020, 10, 576559. [Google Scholar] [CrossRef] [PubMed]
- Shallis, R.M.; Wang, R.; Davidoff, A.; Ma, X.; Zeidan, A.M. Epidemiology of acute myeloid leukemia: Recent progress and enduring challenges. Blood Rev. 2019, 36, 70–87. [Google Scholar] [CrossRef]
- Creutzig, U.; Kutny, M.A.; Barr, R.; Schlenk, R.F.; Ribeiro, R.C. Acute myelogenous leukemia in adolescents and young adults. Pediatr. Blood Cancer 2018, 65, e27089. [Google Scholar] [CrossRef]
- Pelcovits, A.; Niroula, R. Acute Myeloid Leukemia: A Review. Rhode Isl. Med. J. 2020, 103, 38–40. [Google Scholar]
- Castaigne, S.; Pautas, C.; Terré, C.; Raffoux, E.; Bordessoule, D.; Bastie, J.N.; Legrand, O.; Thomas, X.; Turlure, P.; Reman, O.; et al. Acute Leukemia French Association. Effect of gemtuzumab ozogamicin on survival of adult patients with de-novo acute myeloid leukaemia (ALFA-0701): Aandomizedd, open-label, phase 3 study. Lancet 2012, 379, 1508–1516. [Google Scholar] [CrossRef]
- Cortes, J.E.; Heidel, F.H.; Hellmann, A.; Fiedler, W.; Smith, B.D.; Robak, T.; Montesinos, P.; Pollyea, D.A.; DesJardins, P.; Ottmann, O.; et al. Randomized comparison of low dose cytarabine with or without glasdegib in patients with newly diagnosed acute myeloid leukemia or high-risk myelodysplastic syndrome. Leukemia 2018, 33, 379–389. [Google Scholar] [CrossRef] [Green Version]
- Burchert, A.; Bug, G.; Fritz, L.V.; Finke, J.; Stelljes, M.; Röllig, C.; Wollmer, E.; Wäsch, R.; Bornhäuser, M.; Berg, T.; et al. Sorafenib Maintenance After Allogeneic Hematopoietic Stem Cell Transplantation for Acute Myeloid Leukemia with FLT3–Internal Tandem Duplication Mutation (SORMAIN). J. Clin. Oncol. 2020, 38, 2993–3002. [Google Scholar] [CrossRef] [PubMed]
- Xuan, L.; Wang, Y.; Huang, F.; Fan, Z.; Xu, Y.; Sun, J.; Xu, N.; Deng, L.; Li, X.; Liang, X.; et al. Sorafenib maintenance in patients with FLT3-ITD acute myeloid leukaemia undergoing allogeneic haematopoietic stem-cell transplantation: An open-label, multicentre, randomizedd phase 3 trial. Lancet Oncol. 2020, 21, 1201–1212. [Google Scholar] [CrossRef] [PubMed]
- Amawi, H.; Sim, H.-M.; Tiwari, A.K.; Ambudkar, S.V.; Shukla, S. ABC Transporter-Mediated Multidrug-Resistant Cancer. Drug Transp. Drug Dispos. Eff. Toxic. 2019, 1141, 549–580. [Google Scholar] [CrossRef]
- Vasconcelos, F.C.; de Souza, P.S.; Hancio, T.; de Faria, F.C.C.; Maia, R.C. Update on drug transporter proteins in acute myeloid leukemia: Pathological implication and clinical setting. Crit. Rev. Oncol. 2021, 160, 103281. [Google Scholar] [CrossRef]
- Gao, F.; Dong, W.; Yang, W.; Liu, J.; Zheng, Z.; Sun, K. Expression of P-gp in acute myeloid leukemia and the reversal function of As2O3 on drug resistance. Oncol. Lett. 2014, 9, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Poma, P.; Labbozzetta, M.; Notarbartolo, M. Patterns of Innate or Acquired Resistance to Anticancer Drugs: Our Experience to Overcome It. Crit. Rev. Oncog. 2021, 26, 27–37. [Google Scholar] [CrossRef]
- Palmeira, A.; Sousa, E.; Vasconcelos, M.H.; Pinto, M.M. Three Decades of P-gp Inhibitors: Skimming Through Several Generations and Scaffolds. Curr. Med. Chem. 2012, 19, 1946–2025. [Google Scholar] [CrossRef]
- Robey, R.W.; Pluchino, K.M.; Hall, M.D.; Fojo, A.T.; Bates, S.E.; Gottesman, M.M. Revisiting the role of ABC transporters in multidrug-resistant cancer. Nat. Rev. Cancer 2018, 18, 452–464. [Google Scholar] [CrossRef]
- Dewanjee, S.; Dua, T.K.; Bhattacharjee, N.; Das, A.; Gangopadhyay, M.; Khanra, R.; Joardar, S.; Riaz, M.; De Feo, V.; Zia-Ul-Haq, M. Natural Products as Alternative Choices for P-Glycoprotein (P-gp) Inhibition. Molecules 2017, 22, 871. [Google Scholar] [CrossRef]
- Kumar, A.; Jaitak, V. Natural products as multidrug resistance modulators in cancer. Eur. J. Med. Chem. 2019, 176, 268–291. [Google Scholar] [CrossRef]
- Honda, Y.; Ushigome, F.; Koyabu, N.; Morimoto, S.; Shoyama, Y.; Uchiumi, T.; Kuwano, M.; Ohtani, H.; Sawada, Y. Effects of grapefruit juice and orange juice components on P-glycoprotein- and MRP2-mediated drug efflux. Br. J. Pharmacol. 2004, 143, 856–864. [Google Scholar] [CrossRef] [PubMed]
- Shah, D.; Ajazuddin; Bhattacharya, S. Role of natural P-gp inhibitor in the effective delivery for chemotherapeutic agents. J. Cancer Res. Clin. Oncol. 2022, 149, 367–391. [Google Scholar] [CrossRef] [PubMed]
- Lebecque, S.; Lins, L.; Dayan, F.E.; Fauconnier, M.-L.; Deleu, M. Interactions between Natural Herbicides and Lipid Bilayers Mimicking the Plant Plasma Membrane. Front. Plant Sci. 2019, 10, 329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broniatowski, M.; Flasiński, M.; Wydro, P. Investigation of the interactions of lupane type pentacyclic triterpenes with outer leaflet membrane phospholipids–Langmuir monolayer and synchrotron X-ray scattering study. J. Colloid Interface Sci. 2012, 381, 116–124. [Google Scholar] [CrossRef]
- Silva, N.; Salgueiro, L.; Fortuna, A.; Cavaleiro, C. P-glycoprotein Mediated Efflux Modulators of Plant Origin: A Short Review. Nat. Prod. Commun. 2016, 11, 1934578X1601100538. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, N.; Takagi, A.; Kitazawa, H.; Kawakami, J.; Adachi, I. Inhibition of P-glycoprotein-mediated transport by extracts of and monoterpenoids contained in Zanthoxyli Fructus. Toxicol. Appl. Pharmacol. 2005, 209, 167–173. [Google Scholar] [CrossRef]
- Ferreira, A.; Pousinho, S.; Fortuna, A.; Falcão, A.; Alves, G. Flavonoid compounds as reversal agents of the P-glycoprotein-mediated multidrug resistance: Biology, chemistry and pharmacology. Phytochem. Rev. 2014, 14, 233–272. [Google Scholar] [CrossRef]
- Yu, J.; Zhou, P.; Asenso, J.; Yang, X.-D.; Wang, C.; Wei, W. Advances in plant-based inhibitors of P-glycoprotein. J. Enzym. Inhib. Med. Chem. 2016, 31, 867–881. [Google Scholar] [CrossRef] [Green Version]
- Phang, J.M.; Poore, C.M.; Lopaczynska, J.; Yeh, G.C. Flavonol stimulated efflux of 7,12-dimethylbenz(a)-anthracene in multi-drug-resistant breast cancer cells. Cancer Res. 1993, 53, 5977–5981. [Google Scholar]
- Critchfield, J.W.; Welsh, C.J.; Phang, J.M.; Yeh, G.C. Modulation of adriamycin® accumulation and efflux by flavonoids in HCT-15 colon cells: Activation of P-glycoprotein as a putative mechanism. Biochem. Pharmacol. 1994, 48, 1437–1445. [Google Scholar] [CrossRef]
- Chen, F.-Y.; Cao, L.-F.; Wan, H.-X.; Zhang, M.-Y.; Cai, J.-Y.; Shen, L.-J.; Zhong, J.-H.; Zhong, H. Quercetin enhances adriamycin cytotoxicity through induction of apoptosis and regulation of mitogen-activated protein kinase/extracellular signal-regulated kinase/c-Jun N-terminal kinase signaling in multidrug-resistant leukemia K562 cells. Mol. Med. Rep. 2014, 11, 341–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.; Enomoto, R.; Koshiba, C.; Hirano, H. Inhibition of P-Glycoprotein by Wogonin Is Involved with the Potentiation of Etoposide-Induced Apoptosis in Cancer Cells. Ann. N. Y. Acad. Sci. 2009, 1171, 132–136. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.-N.; Jiang, Y.-L.; Zhang, S.-G.; Huang, S.-Y.; Li, H. Grape seed proanthocyanidin extract reverses multidrug resistance in HL-60/ADR cells via inhibition of the PI3K/Akt signaling pathway. Biomed. Pharmacother. 2020, 125, 109885. [Google Scholar] [CrossRef] [PubMed]
- Lopes-Rodrigues, V.; Sousa, M.E.; Vasconcelos, M.H. Curcumin as a Modulator of P-Glycoprotein in Cancer: Challenges and Perspectives. Pharmaceuticals 2016, 9, 71. [Google Scholar] [CrossRef] [Green Version]
- Lopes-Rodrigues, V.; Oliveira, A.; Correia-Da-Silva, M.; Pinto, M.; Lima, R.T.; Sousa, E.; Vasconcelos, M.H. A novel curcumin derivative which inhibits P-glycoprotein, arrests cell cycle and induces apoptosis in multidrug resistance cells. Bioorganic Med. Chem. 2017, 25, 581–596. [Google Scholar] [CrossRef]
- Mi, Q.; Cui, B.; Silva, G.; Lantvit, D.; Lim, E.; Chai, H.; You, M.; Hollingshead, M.G.; Mayo, J.G.; Kinghorn, A.D.; et al. Pervilleine A, a novel tropane alkaloid that reverses the multidrug-resistance phenotype. Cancer Res 2001, 61, 4030–4037. [Google Scholar]
- Ma, Y.; Wink, M. Lobeline, a piperidine alkaloid from Lobelia can reverse P-gp dependent multidrug resistance in tumor cells. Phytomedicine 2008, 15, 754–758. [Google Scholar] [CrossRef]
- Eid, S.Y.; El-Readi, M.Z.; Eldin, E.E.M.N.; Fatani, S.H.; Wink, M. Influence of combinations of digitonin with selected phenolics, terpenoids, and alkaloids on the expression and activity of P-glycoprotein in leukaemia and colon cancer cells. Phytomedicine 2013, 21, 47–61. [Google Scholar] [CrossRef]
- Wei, Y.-L.; Xu, L.; Liang, Y.; Xu, X.-H.; Zhao, X.-Y. Berbamine exhibits potent antitumor effects on imatinib-resistant CML cells in vitro and in vivo. Acta Pharmacol. Sin. 2009, 30, 451–457. [Google Scholar] [CrossRef] [Green Version]
- Pearce, H.L.; Safa, A.R.; Bach, N.J.; Winter, M.A.; Cirtain, M.C.; Beck, W.T. Essential features of the P-glycoprotein pharmacophore as defined by a series of reserpine analogs that modulate multidrug resistance. Proc. Natl. Acad. Sci. USA 1989, 86, 5128–5132. [Google Scholar] [CrossRef] [Green Version]
- Shiraki, N.; Okamura, K.; Tokunaga, J.; Ohmura, T.; Yasuda, K.; Kawaguchi, T.; Hamada, A.; Nakano, M. Bromocriptine Reverses P-Glycoprotein-mediated Multidrug Resistance in Tumor Cells. Jpn. J. Cancer Res. 2002, 93, 209–215. [Google Scholar] [CrossRef] [PubMed]
- El-Readi, M.Z.; Hamdan, D.; Farrag, N.; El-Shazly, A.; Wink, M. Inhibition of P-glycoprotein activity by limonin and other secondary metabolites from Citrus species in human colon and leukaemia cell lines. Eur. J. Pharmacol. 2010, 626, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Corea, G.; Di Pietro, A.; Dumontet, C.; Fattorusso, E.; Lanzotti, V. Jatrophane diterpenes from Euphorbia spp. as modulators of multidrug resistance in cancer therapy. Phytochem. Rev. 2009, 8, 431–447. [Google Scholar] [CrossRef]
- Fontana, G.; Badalamenti, N.; Bruno, M.; Castiglione, D.; Notarbartolo, M.; Poma, P.; Spinella, A.; Tutone, M.; Labbozzetta, M. Synthesis, In Vitro and In Silico Analysis of New Oleanolic Acid and Lupeol Derivatives against Leukemia Cell Lines: Involvement of the NF-κB Pathway. Int. J. Mol. Sci. 2022, 23, 6594. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Zhang, X.; Xie, L.; Deng, M.; Chen, H.; Song, J.; Long, J.; Li, X.; Luo, J. Lupeol and its derivatives as anticancer and anti-inflammatory agents: Molecular mechanisms and therapeutic efficacy. Pharmacol. Res. 2020, 164, 105373. [Google Scholar] [CrossRef] [PubMed]
- Labbozzetta, M.; Poma, P.; Tutone, M.; McCubrey, J.A.; Sajeva, M.; Notarbartolo, M. Phytol and Heptacosane Are Possible Tools to Overcome Multidrug Resistance in an In Vitro Model of Acute Myeloid Leukemia. Pharmaceuticals 2022, 15, 356. [Google Scholar] [CrossRef]
- Choi, C.-H.; Kang, G.; Min, Y.-D. Reversal of P-Glycoprotein-Mediated Multidrug Resistance by Protopanaxatriol Ginsenosides from Korean Red Ginseng. Planta Medica 2003, 69, 235–240. [Google Scholar] [CrossRef]
- Nguyen, V.T.B.; Darbour, N.; Bayet, C.; Doreau, A.; Raad, I.; Phung, B.H.; Dumontet, C.; Di Pietro, A.; Dijoux-Franca, M.-G.; Guilet, D. Selective modulation of P-glycoprotein activity by steroidal saponines from Paris polyphylla. Fitoterapia 2009, 80, 39–42. [Google Scholar] [CrossRef] [Green Version]
- Duarte, N.; Járdánházy, A.; Molnár, J.; Hilgeroth, A.; Ferreira, M.-J.U. Synergistic interaction between p-glycoprotein modulators and epirubicine on resistant cancer cells. Bioorganic Med. Chem. 2008, 16, 9323–9330. [Google Scholar] [CrossRef]
- Suhett, L.G.; Santos, R.D.M.M.; Silveira, B.K.S.; Leal, A.C.G.; de Brito, A.D.M.; de Novaes, J.F.; Della Lucia, C.M. Effects of curcumin supplementation on sport and physical exercise: A systematic review. Crit. Rev. Food Sci. Nutr. 2020, 61, 946–958. [Google Scholar] [CrossRef]
- Islam, M.T.; Ali, E.S.; Uddin, S.J.; Shaw, S.; Islam, A.; Ahmed, I.; Shill, M.C.; Karmakar, U.K.; Yarla, N.S.; Khan, I.N.; et al. Phytol: A review of biomedical activities. Food Chem. Toxicol. 2018, 121, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Lenucci, M.S.; Tornese, R.; Mita, G.; Durante, M. Bioactive Compounds and Antioxidant Activities in Different Fractions of Mango Fruits (Mangifera indica L., Cultivar Tommy Atkins and Keitt). Antioxidants 2022, 11, 484. [Google Scholar] [CrossRef] [PubMed]
- Zito, P.; Sajeva, M.; Bruno, M.; Maggio, A.M.; Rosselli, S.; Senatore, F.; Formisano, C. Essential oil composition of the fruits ofPeriploca laevigataAiton subsp.angustifolia(Labill.) Markgraf (Apocynaceae-Periplocoideae). Nat. Prod. Res. 2011, 25, 1339–1346. [Google Scholar] [CrossRef] [PubMed]
- Dhatwalia, J.; Kumari, A.; Verma, R.; Upadhyay, N.; Guleria, I.; Lal, S.; Thakur, S.; Gudeta, K.; Kumar, V.; Chao, J.C.-J.; et al. Phytochemistry, Pharmacology, and Nutraceutical Profile of Carissa Species: An Updated Review. Molecules 2021, 26, 7010. [Google Scholar] [CrossRef] [PubMed]
- Kotha, R.R.; Luthria, D.L. Curcumin: Biological, Pharmaceutical, Nutraceutical, and Analytical Aspects. Molecules 2019, 24, 2930. [Google Scholar] [CrossRef] [Green Version]
- Urošević, M.; Nikolić, L.; Gajić, I.; Nikolić, V.; Dinić, A.; Miljković, V. Curcumin: Biological Activities and Modern Pharmaceutical Forms. Antibiotics 2022, 11, 135. [Google Scholar] [CrossRef]
- Sohag, A.A.M.; Hossain, T.; Rahaman, A.; Rahman, P.; Hasan, M.S.; Das, R.C.; Khan, K.; Sikder, M.H.; Alam, M.; Uddin, J.; et al. Molecular pharmacology and therapeutic advances of the pentacyclic triterpene lupeol. Phytomedicine 2022, 99, 154012. [Google Scholar] [CrossRef]
- Oh, B.; Boyle, F.; Pavlakis, N.; Clarke, S.; Guminski, A.; Eade, T.; Lamoury, G.; Carroll, S.; Morgia, M.; Kneebone, A.; et al. Emerging Evidence of the Gut Microbiome in Chemotherapy: A Clinical Review. Front. Oncol. 2021, 11, 706331. [Google Scholar] [CrossRef]
- Rattanathammethee, T.; Tuitemwong, P.; Thiennimitr, P.; Sarichai, P.; Na Pombejra, S.; Piriyakhuntorn, P.; Hantrakool, S.; Chai-Adisaksopha, C.; Rattarittamrong, E.; Tantiworawit, A.; et al. Gut microbiota profiles of treatment-naïve adult acute myeloid leukemia patients with neutropenic fever during intensive chemotherapy. PLoS ONE 2020, 15, e0236460. [Google Scholar] [CrossRef]
- Lai, Y.; Masatoshi, H.; Ma, Y.; Guo, Y.; Zhang, B. Role of Vitamin K in Intestinal Health. Front. Immunol. 2022, 12, 5491. [Google Scholar] [CrossRef]
- Tsai, F.-S.; Lin, L.-W.; Wu, C.-R. Lupeol and Its Role in Chronic Diseases. Drug Discov. Mother Nat. 2016, 929, 145–175. [Google Scholar] [CrossRef]
- Arnoriaga-Rodríguez, M.; Mayneris-Perxachs, J.; Burokas, A.; Pérez-Brocal, V.; Moya, A.; Portero-Otin, M.; Ricart, W.; Maldonado, R.; Fernández-Real, J.M. Gut bacterial ClpB-like gene function is associated with decreased body weight and a characteristic microbiota profile. Microbiome 2020, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Luo, W.; Chen, Q.; Chen, X.; Zhou, G.; Sun, H. Curcumin sensitizes response to cytarabine in acute myeloid leukemia by regulating intestinal microbiota. Cancer Chemother. Pharmacol. 2022, 89, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Jabczyk, M.; Nowak, J.; Hudzik, B.; Zubelewicz-Szkodzińska, B. Curcumin and Its Potential Impact on Microbiota. Nutrients 2021, 13, 2004. [Google Scholar] [CrossRef] [PubMed]
- Mosca, L.; Pagano, M.; Borzacchiello, L.; Mele, L.; Russo, A.; Russo, G.; Cacciapuoti, G.; Porcelli, M. S-Adenosylmethionine Increases the Sensitivity of Human Colorectal Cancer Cells to 5-Fluorouracil by Inhibiting P-Glycoprotein Expression and NF-κB Activation. Int. J. Mol. Sci. 2021, 22, 9286. [Google Scholar] [CrossRef]
- Zhao, B.-X.; Sun, Y.-B.; Wang, S.-Q.; Duan, L.; Huo, Q.-L.; Ren, F.; Li, G.-F. Grape Seed Procyanidin Reversal of P-glycoprotein Associated Multi-Drug Resistance via Down-regulation of NF-κB and MAPK/ERK Mediated YB-1 Activity in A2780/T Cells. PLoS ONE 2013, 8, e71071. [Google Scholar] [CrossRef]
- Sun, J.; Yeung, C.A.; Na Co, N.; Tsang, T.Y.; Yau, E.; Luo, K.; Wu, P.; Wa, J.C.Y.; Fung, K.-P.; Kwok, T.-T.; et al. Clitocine Reversal of P-Glycoprotein Associated Multi-Drug Resistance through Down-Regulation of Transcription Factor NF-κB in R-HepG2 Cell Line. PLoS ONE 2012, 7, e40720. [Google Scholar] [CrossRef]
- Kaewpiboon, C.; Surapinit, S.; Malilas, W.; Moon, J.; Phuwapraisirisan, P.; Tip-Pyang, S.; Johnston, R.N.; Koh, S.S.; Assavalapsakul, W.; Chung, Y.-H. Feroniellin A-induced autophagy causes apoptosis in multidrug-resistant human A549 lung cancer cells. Int. J. Oncol. 2014, 44, 1233–1242. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Zhao, L.; Wang, H.; Wang, Y.; Pan, D.; Yao, J.; Li, Z.; Wu, G.; Guo, Q. Oroxylin A reverses P-glycoprotein-mediated multidrug resistance of MCF7/ADR cells by G2/M arrest. Toxicol. Lett. 2013, 219, 107–115. [Google Scholar] [CrossRef]
- Poma, P.; Labbozzetta, M.; Ramarosandratana, A.; Rosselli, S.; Tutone, M.; Sajeva, M.; Notarbartolo, M. In Vitro Modulation of P-Glycoprotein Activity by Euphorbia intisy Essential Oil on Acute Myeloid Leukemia Cell Line HL-60R. Pharmaceuticals 2021, 14, 111. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.M.S.; Heimfarth, L.; Pereira, E.W.M.; Oliveira, F.S.; Menezes, I.R.A.; Coutinho, H.D.M.; Picot, L.; Antoniolli, A.R.; Quintans, J.S.S.; Quintans-Júnior, L.J. Phytol, a Chlorophyll Component, Produces Antihyperalgesic, Anti-inflammatory, and Antiarthritic Effects: Possible NFκB Pathway Involvement and Reduced Levels of the Proinflammatory Cytokines TNF-α and IL-6. J. Nat. Prod. 2020, 83, 1107–1117. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.E.; Lo, C.R.; Liu, H.; Pradhan, Z.; Garcia, L.; Srinivasan, A.; Valentino, K.; Czaja, M.J. Role of caspases and NF-kappaB signaling in hydrogen peroxide- and superoxide-induced hepatocyte apoptosis. Am. J. Physiol.-Gastrointest. Liver Physiol. 2000, 278, G693–G699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozaki, I.; Ide, Y.; Zhang, H.; Hamajima, H.; Kawaguchi, Y.; Eguchi, Y.; Mizuta, T.; Yamamoto, K.; Fujimoto, K. Inhibition of matrix metalloproteinase expression by menatetrenone, a vitamin K2 analogue. Oncol. Rep. 2009, 22, 599–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozaki, I.; Zhang, H.; Mizuta, T.; Ide, Y.; Eguchi, Y.; Yasutake, T.; Sakamaki, T.; Pestell, R.G.; Yamamoto, K. Menatetrenone, a Vitamin K2 Analogue, Inhibits Hepatocellular Carcinoma Cell Growth by Suppressing Cyclin D1 Expression through Inhibition of Nuclear Factor κB Activation. Clin. Cancer Res. 2007, 13, 2236–2245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, K.-R.; Choi, J.-Y.; Chung, S.; Kim, D.-H. Anti-scratching Behavioral Effect of the Essential Oil and Phytol Isolated from Artemisia princeps Pamp. in Mice. Planta Medica 2010, 77, 22–26. [Google Scholar] [CrossRef]
- Chandan, G.; Kumar, C.; Verma, M.K.; Satti, N.K.; Saini, A.K.; Saini, R.V. Datura stramonium essential oil composition and it’s immunostimulatory potential against colon cancer cells. 3 Biotech 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Pavithra, P.; Mehta, A.; Verma, R.S. Induction of apoptosis by essential oil from P. missionis in skin epidermoid cancer cells. Phytomedicine 2018, 50, 184–195. [Google Scholar] [CrossRef]
- Zito, P.; Labbozzetta, M.; Notarbartolo, M.; Sajeva, M.; Poma, P. Essential oil of Cyphostemma juttae (Vitaceae): Chemical composition and antitumor mechanism in triple negative breast cancer cells. PLoS ONE 2019, 14, e0214594. [Google Scholar] [CrossRef]
- Poma, P.; Labbozzetta, M.; McCubrey, J.A.; Ramarosandratana, A.V.; Sajeva, M.; Zito, P.; Notarbartolo, M. Antitumor Mechanism of the Essential Oils from Two Succulent Plants in Multidrug Resistance Leukemia Cell. Pharmaceuticals 2019, 12, 124. [Google Scholar] [CrossRef] [Green Version]
- Soltanian, S.; Sheikhbahaei, M.; Ziasistani, M. Phytol Down-Regulates Expression of Some Cancer Stem Cell Markers and Decreases Side Population Proportion in Human Embryonic Carcinoma NCCIT Cells. Nutr. Cancer 2020, 73, 1520–1533. [Google Scholar] [CrossRef]
- Costa, J.; Oliveira, J.S.; Junior, L.M.R.; De Freitas, R.M. Phytol a Natural Diterpenoid with Pharmacological Applications on Central Nervous System: A Review. Recent Patents Biotechnol. 2015, 8, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Bobe, G.; Zhang, Z.; Kopp, R.; Garzotto, M.; Shannon, J.; Takata, Y. Phytol and its metabolites phytanic and pristanic acids for risk of cancer: Current evidence and future directions. Eur. J. Cancer Prev. 2020, 29, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Notarbartolo, M.; Cervello, M.; Poma, P.; Dusonchet, L.; Meli, M.; D’Alessandro, N. Expression of the IAPs in multidrug resistant tumor cells. Oncol. Rep. 2004, 11, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-H.; Cheng, A.-L. CLINICAL STUDIES WITH CURCUMIN. Mol. Targets Ther. Uses Curcumin Health Dis. 2007, 595, 471–480. [Google Scholar] [CrossRef]
- Mirzaei, H.; Shakeri, A.; Rashidi, B.; Jalili, A.; Banikazemi, Z.; Sahebkar, A. Phytosomal curcumin: A review of pharmacokinetic, experimental and clinical studies. Biomed. Pharmacother. 2017, 85, 102–112. [Google Scholar] [CrossRef]
- Tomeh, M.A.; Hadianamrei, R.; Zhao, X. A Review of Curcumin and Its Derivatives as Anticancer Agents. Int. J. Mol. Sci. 2019, 20, 1033. [Google Scholar] [CrossRef] [Green Version]
- Vyas, A.; Dandawate, P.; Padhye, S.; Ahmad, A.; Sarkar, F. Perspectives on new synthetic curcumin analogs and their potential anticancer properties. Curr. Pharm. Des. 2013, 19, 2047–2069. [Google Scholar]
- Hosseini-Zare, M.S.; Sarhadi, M.; Zarei, M.; Thilagavathi, R.; Selvam, C. Synergistic effects of curcumin and its analogs with other bioactive compounds: A comprehensive review. Eur. J. Med. Chem. 2020, 210, 113072. [Google Scholar] [CrossRef]
- Bonaccorsi, P.M.; Labbozzetta, M.; Barattucci, A.; Salerno, T.M.G.; Poma, P.; Notarbartolo, M. Synthesis of Curcumin Derivatives and Analysis of Their Antitumor Effects in Triple Negative Breast Cancer (TNBC) Cell Lines. Pharmaceuticals 2019, 12, 161. [Google Scholar] [CrossRef] [Green Version]
- Joshi, P.; Joshi, S.; Semwal, D.; Bisht, A.; Paliwal, S.; Dwivedi, J.; Sharma, S. Curcumin: An Insight into Molecular Pathways Involved in Anticancer Activity. Mini-Reviews Med. Chem. 2021, 21, 2420–2457. [Google Scholar] [CrossRef]
- Choi, B.H.; Kim, C.G.; Lim, Y.; Shin, S.Y.; Lee, Y.H. Curcumin down-regulates the multidrug-resistance mdr1b gene by inhibiting the PI3K/Akt/NFκB pathway. Cancer Lett. 2008, 259, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasan, S.; Ravichandran, S.; Vetrivel, U.; Krishnakumar, S. Modulation of multidrug resistance 1 expression and function in retinoblastoma cells by curcumin. J. Pharmacol. Pharmacother. 2013, 4, 103–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.-K.; Ki, S.-H.; Choi, J.-S. Effects of oral curcumin on the pharmacokinetics of intravenous and oral etoposide in rats: Possible role of intestinal CYP3A and P-gp inhibition by curcumin. Biopharm. Drug Dispos. 2011, 32, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Anuchapreeda, S.; Leechanachai, P.; Smith, M.M.; Ambudkar, S.V.; Limtrakul, P.-N. Modulation of P-glycoprotein expression and function by curcumin in multidrug-resistant human KB cells. Biochem. Pharmacol. 2002, 64, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Tian, W.; Shen, H. P-gp upregulation may be blocked by natural curcuminoids, a novel class of chemoresistance-preventing agent. Mol. Med. Rep. 2012, 7, 115–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, L.; Zhao, P.; Li, Y.; Yang, D.; Hu, P.; Li, L.; Cheng, Y.; Yao, H. Reversal of P-glycoprotein-mediated multidrug resistance by novel curcumin analogues in paclitaxel-resistant human breast cancer cells. Biochem. Cell Biol. 2020, 98, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Mapoung, S.; Pitchakarn, P.; Yodkeeree, S.; Ovatlarnporn, C.; Sakorn, N.; Limtrakul, P. Chemosensitizing effects of synthetic curcumin analogs on human multi-drug resistance leukemic cells. Chem. Interactions 2016, 244, 140–148. [Google Scholar] [CrossRef]
- Revalde, J.L.; Li, Y.; Hawkins, B.C.; Rosengren, R.J.; Paxton, J.W. Heterocyclic cyclohexanone monocarbonyl analogs of curcumin can inhibit the activity of ATP-binding cassette transporters in cancer multidrug resistance. Biochem. Pharmacol. 2015, 93, 305–317. [Google Scholar] [CrossRef]
- Alqathama, A.; Ezuruike, U.F.; Mazzari, A.L.D.A.; Yonbawi, A.; Chieli, E.; Prieto, J.M. Effects of Selected Nigerian Medicinal Plants on the Viability, Mobility, and Multidrug-Resistant Mechanisms in Liver, Colon, and Skin Cancer Cell Lines. Front. Pharmacol. 2020, 11. [Google Scholar] [CrossRef]
- Machado, V.R.; Jacques, A.V.; Marceli, N.S.; Biavatti, M.W.; Santos-Silva, M.C. Anti-leukemic activity of semisynthetic derivatives of lupeol. Nat. Prod. Res. 2020, 35, 4494–4501. [Google Scholar] [CrossRef]
- Anuchapreeda, S.; Chueahongthong, F.; Viriyaadhammaa, N.; Panyajai, P.; Anzawa, R.; Tima, S.; Ampasavate, C.; Saiai, A.; Rungrojsakul, M.; Usuki, T.; et al. Antileukemic Cell Proliferation of Active Compounds from Kaffir Lime (Citrus hystrix) Leaves. Molecules 2020, 25, 1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buakaew, W.; Sranujit, R.P.; Noysang, C.; Thongsri, Y.; Potup, P.; Nuengchamnong, N.; Suphrom, N.; Usuwanthim, K. Phytochemical Constituents of Citrus hystrix DC. Leaves Attenuate Inflammation via NF-κB Signaling and NLRP3 Inflammasome Activity in Macrophages. Biomolecules 2021, 11, 105. [Google Scholar] [CrossRef]
- Beserra, F.P.; Gushiken, L.F.S.; Vieira, A.J.; Bérgamo, D.A.; Bérgamo, P.L.; De Souza, M.O.; Hussni, C.A.; Takahira, R.K.; Nóbrega, R.H.; Martinez, E.R.M.; et al. From Inflammation to Cutaneous Repair: Topical Application of Lupeol Improves Skin Wound Healing in Rats by Modulating the Cytokine Levels, NF-κB, Ki-67, Growth Factor Expression, and Distribution of Collagen Fibers. Int. J. Mol. Sci. 2020, 21, 4952. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.K.; Poon, R.T.; Wo, J.Y.; Ma, S.; Guan, X.-Y.; Myers, J.N.; Altevogt, P.; Yuen, A.P. Lupeol Suppresses Cisplatin-Induced Nuclear Factor-κB Activation in Head and Neck Squamous Cell Carcinoma and Inhibits Local Invasion and Nodal Metastasis in an Orthotopic Nude Mouse Model. Cancer Res. 2007, 67, 8800–8809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleem, M.; Kaur, S.; Kweon, M.-H.; Adhami, V.M.; Afaq, F.; Mukhtar, H. Lupeol, a fruit and vegetable based triterpene, induces apoptotic death of human pancreatic adenocarcinoma cells via inhibition of Ras signaling pathway. Carcinog. 2005, 26, 1956–1964. [Google Scholar] [CrossRef] [Green Version]
- Saleem, M.; Afaq, F.; Adhami, V.M.; Mukhtar, H. Lupeol modulates NF-κB and PI3K/Akt pathways and inhibits skin cancer in CD-1 mice. Oncogene 2004, 23, 5203–5214. [Google Scholar] [CrossRef] [Green Version]
- Prasad, S.; Madan, E.; Nigam, N.; Roy, P.; George, J.; Shukla, Y. Induction of apoptosis by lupeol in human epidermoid carcinoma A431 cells through regulation of mitochondrial, Akt/PKB and NF-kappaB signaling pathways. Cancer Biol. Ther. 2009, 8, 1632–1639. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wang, Y.; Yu, T.; Hu, Z.; Wang, Y. An LC–ESI/MS/MS method for the determination of lupeol via precolumn derivatization and its application to pharmacokinetic studies in rat plasma. Biomed. Chromatogr. 2020, 35, e5005. [Google Scholar] [CrossRef]
- Khatal, L.; More, H. Development and validation of a liquid chromatography-tandem mass spectrometry method for quantification of Lupeol in plasma and its application to pharmacokinetic study in rats. J. Chromatogr. B 2019, 1121, 58–65. [Google Scholar] [CrossRef]
- Hanif, S.; Jabeen, K.; Akhtar, N.; Iqbal, S. GC-MS analysis & antifungal activity of Datura metel L. against Rhizoctonia solani Kuhn. Anais da Academia Brasileira de Ciências 2022, 94. [Google Scholar] [CrossRef]
- Balikci, N.; Sarimahmut, M.; Ari, F.; Aztopal, N.; Özel, M.Z.; Ulukaya, E.; Celikler, S. Toxicity assessment of Hypericum olympicum subsp. olympicum L. on human lymphocytes and breast cancer cell lines. J. Appl. Biomed. 2020, 18, 18–25. [Google Scholar] [CrossRef] [Green Version]
- Kuiate, J.-R.; Bessière, J.M.; Zollo, P.H.A.; Kuate, S.P. Chemical composition and antidermatophytic properties of volatile fractions of hexanic extract from leaves of Cupressus lusitanica Mill. from Cameroon. J. Ethnopharmacol. 2006, 103, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Dunkić, V.; Kosalec, I.; Košir, I.; Potočnik, T.; Čerenak, A.; Končič, M.; Vitali, D.; Muller, I.; Kopricanec, M.; Bezić, N.; et al. Antioxidant and antimicrobial properties of Veronica spicata L. (Plantaginaceae). Curr. Drug Targets 2015, 16, 1660–1670. [Google Scholar] [CrossRef] [PubMed]
- Köse, Y.B.; Işcan, G.; Göger, F.; Akalın, G.; Demirci, B.; Başer, K.H.C. Chemical Composition and Biological Activity of Centaurea baseri: New Species from Turkey. Chem. Biodivers. 2016, 13, 1369–1379. [Google Scholar] [CrossRef] [PubMed]
- Ambudkar, S.V.; Dey, S.; Hrycyna, C.A.; Ramachandra, M.; Pastan, I.; Gottesman, M.M. Biochemical, Cellular, and Pharmacological Aspects of the Multidrug Transporter. Annu. Rev. Pharmacol. Toxicol. 1999, 39, 361–398. [Google Scholar] [CrossRef] [Green Version]
- Litman, T.; Skovsgaard, T.; Stein, W.D. Pumping of Drugs by P-Glycoprotein: A Two-Step Process? Experiment 2003, 307, 846–853. [Google Scholar] [CrossRef] [Green Version]
- Haubner, S.; Perna, F.; Köhnke, T.; Schmidt, C.; Berman, S.; Augsberger, C.; Schnorfeil, F.M.; Krupka, C.; Lichtenegger, F.S.; Liu, X.; et al. Coexpression profile of leukemic stem cell markers for combinatorial targeted therapy in AML. Leukemia 2018, 33, 64–74. [Google Scholar] [CrossRef]
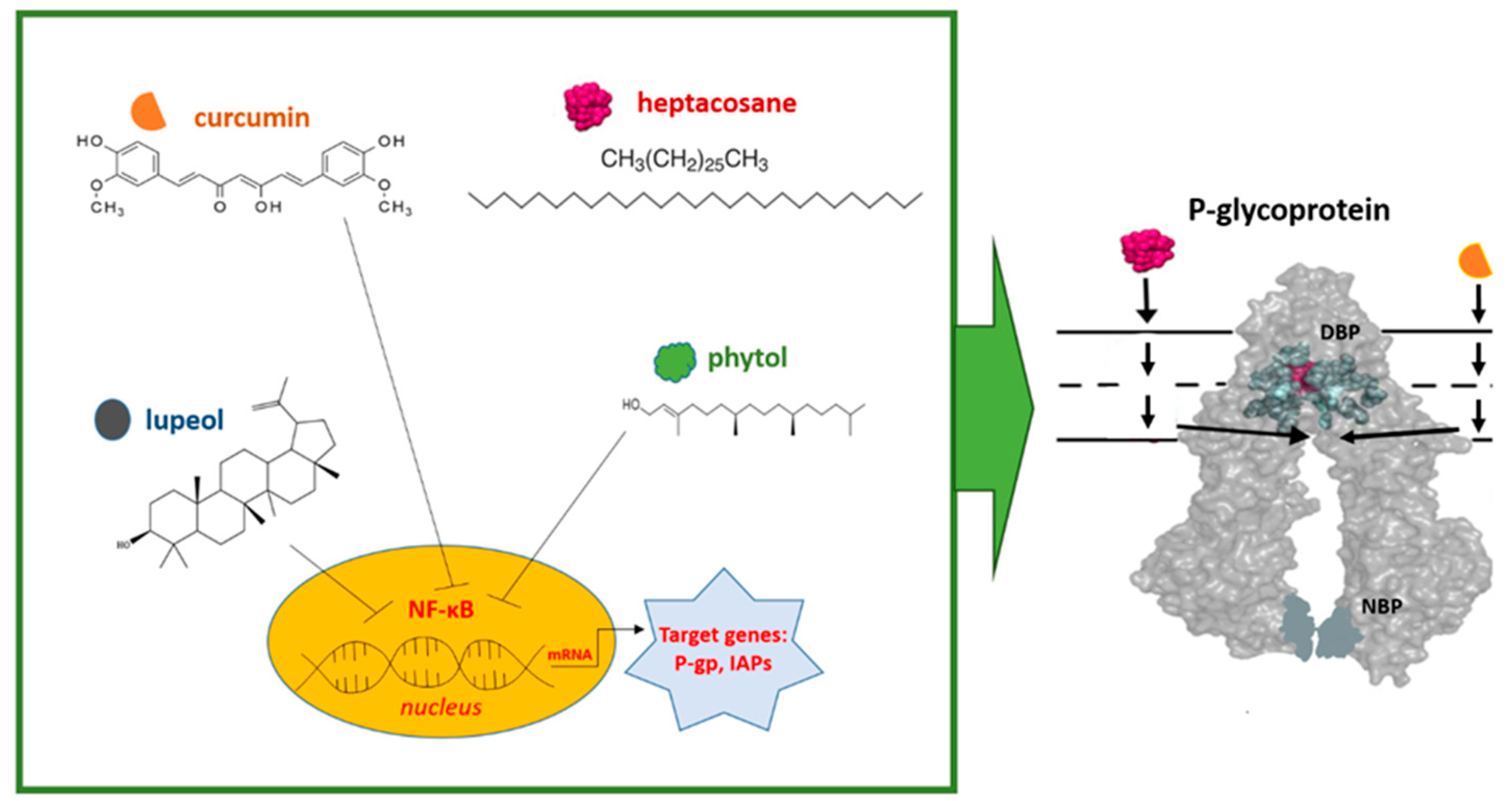
Figure 1.
Graphical representation of the observed mechanisms of action of phytol, curcumin, lupeol, and heptacosane on the expression and function of P-gp in AML. Curcumin, phytol, and lupeol act by modulating the expression of P-gp through the inhibition of NF-κB. Curcumin is also able to modulate the function of P-gp through interaction with the DBP site. The modulation mechanism of heptacosane is instead linked to its interaction with the DBP and NBP sites (see text for explanation). (IAPs: apoptosis inhibitory proteins; DBP: drug-binding pocket; NBP: nucleotide-binding pocket).
Figure 1.
Graphical representation of the observed mechanisms of action of phytol, curcumin, lupeol, and heptacosane on the expression and function of P-gp in AML. Curcumin, phytol, and lupeol act by modulating the expression of P-gp through the inhibition of NF-κB. Curcumin is also able to modulate the function of P-gp through interaction with the DBP site. The modulation mechanism of heptacosane is instead linked to its interaction with the DBP and NBP sites (see text for explanation). (IAPs: apoptosis inhibitory proteins; DBP: drug-binding pocket; NBP: nucleotide-binding pocket).


{kind=link}
Table 1.
Some examples of natural inhibitors of P-glycoprotein in leukemia cell lines.
| P-gp Natural Inhibitors “Fourth Generation” | Mechanism of Action |
|---|---|
| Flavonoids | |
| Quercetin | Inhibition of P-gp gene expression in adryamicin-resistant human chronic myeloid leukemia cell line (K562/ADR) [31]. |
| Wogonin | Inhibition of P-gp expression in a human acute myeloid leukemia cell line (HL-60) [32]. |
| Grape seed proanthocyanidin extract (GSPE) | GSPE inhibits P-gp expression via the PI3K / Akt signal transduction pathway in adryamicin-resistant human acute myeloid leukemia cell line (HL-60/ADR) [33]. |
| Curcumin | Inhibition of P-gp mediated efflux in a doxorubicin-resistant human chronic myeloid leukemia cell line (K562/Dox) [34,35]. |
| Alkaloids | |
| Pervilleine A | Inhibition of P-gp gene expression in a multidrug-resistant human T-cell childhood acute lymphocytic leukemia cell line (CEM/VLB100) [36]. |
| Lobeline | Inhibition of P-gp mediated efflux probably by substrate competition in a multidrug-resistant human T-cell childhood acute lymphocytic leukemia cell line (CEM/ADR5000) [37]. |
| harmine, and sanguinarine | Inhibition of P-gp gene expression in a multidrug-resistant human T-cell childhood acute lymphocytic leukemia cell line (CEM/ADR5000) [38]. |
| Berbamine | Inhibition of mRNA and P-gp protein expression in an imatinib-resistant BCR-ABL-positive chronic myeloid leukemia cell line (K562-r) [39]. |
| Reserpine and yohimbine | Inhibition of P-gp mediated efflux in a multidrug-resistant human T-cell childhood acute lymphocytic leukemia cell line (CEM/VLB100) [40]. |
| Bromocriptine | Inhibition of the overexpressed P-gp protein in a vinblastine-resistant human chronic myeloid leukemia cell line (K562/R10) [41]. |
| Isoquinoline alkaloid chelidonine | Inhibition of P-gp mediated efflux in multidrug-resistant human T-cell childhood acute lymphocytic leukemia cell line (CEM/DOX5000) [20]. |
| Terpenoids | |
| Limonin | Inhibition of P-gp mediated efflux in a multidrug-resistant human T-cell childhood acute lymphocytic leukemia cell lines (CCRF-CEM and CEM/ADR5000) [42]. |
| Euphodendroidin D and Pepluanin A | Inhibition of P-gp mediated efflux via binding with its active sites in a daunomycin-resistant human chronic myeloid leukemia cell line (K562/R7) [43]. |
| Lupeol | Inhibition of mRNA and P-gp protein expression in a doxorubicin-resistant human acute myeloid leukemia cell line (HL-60R) [44,45]. |
| Phytol and heptacosane | Inhibition of P-gp mediated efflux via binding with the DBP site, in a similar way to verapamil, in a doxorubicin-resistant human acute myeloid leukemia cell line (HL-60R) [46]. |
| Saponins | |
| Ginsenoside F1 | Inhibition of P-gp mediated efflux in daunorubicin- and doxorubicin-resistant acute myeloid leukemia sublines (AML-2/D100 and AML-2/DX100) [47]. |
| Gracillin | Inhibition of P-gp mediated efflux via direct interaction with active binding sites in a daunorubicin-resistant human chronic myeloid leukemia cell line (K562/R7) [48]. |
| Pinnatasterone | Inhibition of P-gp mediated efflux via binding with its active sites in a daunomycin-resistant human chronic myeloid leukemia cell line (K562/R7) [49]. |
CEM/VLB100 derived from CCRF-CEM cells with selection for vinblastine resistance; CEM/DOX5000 derived from CCRF-CEM cells with selection for doxorubicin resistance; CEM/ADR5000 derived from CCRF-CEM cells with selection for adriamycin resistance; AML-2/D100 and AML-2/DX100 derived from acute myeloid leukemia (OCI/ AML-2) cells with selection for daunorubicin and for doxorubicin resistances, respectively.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Labbozzetta, M.; Poma, P.; Notarbartolo, M. Natural Inhibitors of P-glycoprotein in Acute Myeloid Leukemia. Int. J. Mol. Sci. 2023, 24, 4140. https://doi.org/10.3390/ijms24044140
AMA Style
Labbozzetta M, Poma P, Notarbartolo M. Natural Inhibitors of P-glycoprotein in Acute Myeloid Leukemia. International Journal of Molecular Sciences. 2023; 24(4):4140. https://doi.org/10.3390/ijms24044140
Chicago/Turabian StyleLabbozzetta, Manuela, Paola Poma, and Monica Notarbartolo. 2023. "Natural Inhibitors of P-glycoprotein in Acute Myeloid Leukemia" International Journal of Molecular Sciences 24, no. 4: 4140. https://doi.org/10.3390/ijms24044140
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.