
Epigenetic Regulation of Driver Genes in Testicular Tumorigenesis
by
 ,
,
Finn E. von Eyben
1,*,
Karsten Kristiansen
2,3,4,
Daniel S. Kapp
5,
Rong Hu
6,
Ovidiu Preda
7 and
Francisco F. Nogales
8 1
Center for Tobacco Control Research, Birkevej 17, 5230 Odense, Denmark
2
Laboratory of Genomics and Molecular Biomedicine, August Krogh Building Department of Biology, University of Copenhagen, Universitetsparken 13, 2100 Copenhagen, Denmark
3
BGI-Research, BGI-Shenzhen, Shenzhen 518120, China
4
Institute of Metagenomics, Qingdao-Europe Advanced Institute for Life Sciences, BGI-Qingdao, Qingdao 166555, China
5
Department of Radiation Oncology, Stanford University, Stanford, CA 94305, USA
6
Department of Pathology, Laboratory Medicine, University of Wisconsin Hospital and Clinics, Madison, WI 53792, USA
7
Department of Pathology, San Cecilio University Hospital, 18071 Granada, CP, Spain
8
Department of Pathology, School of Medicine, University Granada, 18071 Granada, CP, Spain
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(4), 4148; https://doi.org/10.3390/ijms24044148
Submission received: 19 December 2022
/
Revised: 15 February 2023
/
Accepted: 15 February 2023
/
Published: 19 February 2023
(This article belongs to the Special Issue New Evidence and Emerging Strategies in the Management of Testicular Cancer In Vivo and In Vitro)
Abstract
:In testicular germ cell tumor type II (TGCT), a seminoma subtype expresses an induced pluripotent stem cell (iPSC) panel with four upregulated genes, OCT4/POU5F1, SOX17, KLF4, and MYC, and embryonal carcinoma (EC) has four upregulated genes, OCT4/POU5F1, SOX2, LIN28, and NANOG. The EC panel can reprogram cells into iPSC, and both iPSC and EC can differentiate into teratoma. This review summarizes the literature on epigenetic regulation of the genes. Epigenetic mechanisms, such as methylations of cytosines on the DNA string and methylations and acetylations of histone 3 lysines, regulate expression of these driver genes between the TGCT subtypes. In TGCT, the driver genes contribute to well-known clinical characteristics and the driver genes are also important for aggressive subtypes of many other malignancies. In conclusion, epigenetic regulation of the driver genes are important for TGCT and for oncology in general.
1. Introduction: Epigenetics, Differentiation, and Testicular Germ Cell Tumors
In 1957, Waddington described an epigenetic landscape of differentiation where a one-way road leads totipotent stem cells over pluripotent stem cells to unipotent differentiated cells (Figure 1a) [1]. Later studies show that gene panels are able to reprogram unipotent differentiated cells into induced pluripotent stem cells (iPSC) [2,3]. An iPSC panel includes POU5F1 and SOX2, combined with LIN28 and NANOG (the Thomson OSLN panel) [2]. A second iPSC panel includes POU5F1 and SOX2, combined with KLF4 and MYC (the Yamanaka OSKM panel) [3]. iPSC can differentiate into teratoma (TER) [4]. The iPSC concept represents a major paradigm shift which points to a bi-directional two-way road for differentiation in the Waddington landscape.
Two precursor lesions for testicular germ cell tumors type II, germ cell neoplasia in situ (GCNIS) and microinvasive germ cell tumor (MGCT) (Figure 1b) [5,6], and seminoma (SE) also express an OSKM panel, with SOX17 as substitute for SOX2. In SE, SOX17 stimulates genes that inhibit differentiation [7] but allows SE to be transformed into embryonal carcinoma (EC) [8]. EC is the undifferentiated nonseminomatous (NST) subtype of testicular germ cell tumors type II. The transformation includes a shift from a highly expressed SOX17 in SE to a highly expressed SOX2 in EC and a shift from the OSKM to the OSLN panel [9]. The iPSC panels are driver genes in the tumorigenesis of testicular germ cell tumors type II.
EC highly expresses the OSLN panel (Figure 1c) [6]. EC can differentiate both into an embryonic subtype, TER, and into two extra-embryonic subtypes, yolk sac tumor (YST), and choriocarcinoma (CC) (Figure 2a) [6]. As EC differentiates into TER, the OSLN panel is silenced (Figure 1a). As proof of principle for the Waddington landscape has a two-way road of differentiation, Eldar-Geva et al. reported that fibroblasts from a 32-year-old SE patient were reprogrammed into iPSC [10]. A circuit of OCT4, SOX2, and NANOG is crucial for the tumorigenesis of testicular germ cell tumors type II [11]. One of the authors of this review (FFN) was the first to argue that iPSC is crucial for some patients with EC [12].
In this review of the tumorigenesis of TGCT, we examine whether DNA methylations and histone modifications modify the expression of driver genes between the subtypes of testicular germ cell tumors type II, and whether epigenetic writers, readers, and erasers modulate the modifiers.
2. Testicular Germ Cell Tumors Type II
2.1. Pathology
There are seven types of testicular germ cell tumors [13]. Testicular germ cell tumors type I are tumors in early infancy. Testicular germ cell tumors type II (denoted TGCT in our review) have two main groups where patients with SE are diagnosed at a median age of 35 years and patients with NST are diagnosed at a median age of 25 years. Type III consists of spermatocytic tumor (ST). Patients with ST had a median age > 50 years. Type IV and V are mainly female germ cell tumors. Type VI are germ cell tumors that arise due to reprogramming for iPSC.
TGCT is unique in oncology. Of the seven germ cell tumor types, only TGCT originates from GCNIS, first described by Wilms in 1896 [14]. In oncology, only EC develops into TER. TGCT has a pathognomonic genetic abnormality, an isochromosome of the short arm of chromosome 12, i (12p) [15,16,17,18,19]. TGCT has a unique network with downregulated CDKN2A (p16 (INK4), CDNKN2C (p18INK4C), CDKN2D (p19INK4D)), and CDKN1A (p21, CIP1, WAF1), upregulated CCND2, and downregulated RB1 [20]. The genes synergistically inhibit suppression of proliferation.
Compared with other malignancies, patients with TGCT have a unique isoenzyme pattern for serum lactate dehydrogenase (S-LDH) with an unusually high LDH isoenzyme 1 activity (S-LDH-1) [21,22,23]. LDH-1 is a tetramer of four LDHB subunits, generated from LDHB on the short arm of chromosome 12, 12p [24]. Of all malignancies, the international tumor, nodes, and metastases (TNM) classification incorporates serum tumor markers only for TGCT [25]. The serum tumor markers are serum alpha fetoprotein (S-AFP), serum human chorionic gonadotropin (S-hCG), and S-LDH. The classification was adopted from a study by the International Germ Cell Consensus Classification Group [26]. Metastatic TGCT responds extraordinarily well to platin-based chemotherapy, so Einhorn called TGCT a model of a curable solid neoplasm [27].
For young adult Caucasian men, TGCT is the most frequent malignancy, and the incidence has increased worldwide in many recent decades [28,29,30].
The TGCT subtypes differ in frequency. For 78% of testes with TGCT, GCNIS is present in the testis besides the TGCT [31]. A Danish Testis Cancer (DATECA) group study reported a five-year nation-wide cohort of patients with TGCT where a central review reevaluated the TGCT histology according to a World Health Organization (WHO) classification [32]. The cohort included 1058 patients. Half of the patients had SE, slightly less than half of the patients had NST (including those with both SE and NST), and one percent of the patients had ST. Of the NST patients, three quarters of the patients had an EC component in the tumors, half the patients had a TER component, and less than a quarter of the patients had a YST component or a CC component. The patients were followed with regular monitoring of S-AFP and S-hCG [33]. The serum tumor markers were important for staging and outcome of treatment.
2.2. Tumorigenesis
As background for TGCT, endogenous androgen levels during puberty and early adulthood are inversely associated with the risk of TGCT [34]. Severe acne reduced the risk of TGCT with 50%. TGCT develops from normal germ cells through precancer lesions and subclinical precursor lesions to macroscopically overt tumors (Figure 2a) [35,36,37,38,39]. On ultrastructural level, testicular malignant germ cells resemble spermatogonia type A more than gonocytes [40].
2.3. Cytogenetics
The TGCT tumorigenesis is governed by cytogenetic abnormalities. To illustrate the cytogenetic abnormalities, long-term cultures of primordial germ cells gradually increase the copy number of chromosome 12, 17, and X, similar to the pattern in TGCT [41,42,43,44,45].
Walt et al. found that tetraploidization is the first step in the TGCT tumorigenesis [46]. Testicular germ cells have several ways to tetraploidization. Cells that are deficient in CDKN1A (p21/Cip1) can bypass a G2 block and conduct a second S phase before they undergo mitosis [47]. TGCT has deregulated AURKA, AURKB, and AURKC, that may lead to abnormal chromosomal segregation and cytogenesis during cell divisions [48,49]. Later, malignant tetraploid germ cells loose chromosomal material [37]. In consequence, SE consists of hypertriploid cells and NS of hypotriploid cells [36]. GCNIS and SE have a higher ploidy than NST [50,51].
Comparative genomic hybridization of TGCT shows the tumors have gains of parts of chromosome 7, 8, 12p, 14, and X, and losses of parts of chromosomes 3, 4, 5, 10, 11, 12q, 16, 18, 22, and Y [52].
2.4. Genetics
The TGCT subtypes differ in gene expression and the differences contribute to the TGCT tumorigenesis [55]. Two regions of chromosome 12p are important for the tumorigenesis (Figure 2b) as the regions have gene loci for NANOG, CCND2, LDHB, and KRAS2. DNA methylation increases in the progression from GCNIS to differentiated NST subtypes [56,57,58,59].
The gene expression of the OSLN/OSKM panels changes dramatically as SE develops into EC and TER (Figure 2c) [6]. Thus, POU5F1/OCT4, SOX17, SOX2, KLF4, MYC, LIN28, and NANOG are driver genes for TGCT. Other driver genes in undifferentiated TGCT subtypes are upregulated oncogenes CCND2 [60], KLF4 [61], and MYC [62] and a downregulated tumor suppressor gene RB1 [63].
Many candidate genes for TGCT have their gene locus on chromosomes other than the short arm of chromosome 12 (12p). AFP has gene locus on 4q13.3, TET2 on chromosome 4q24, SOX17 on 8q11.23, MYC on 8q14.13, POU5F1 on 8q24,21, KLF4 on 9q31.2, RB1 on 13q14.2, CCGB3 on 19q13.33, and DNMT3B on 20q11.1. Similarities and differences in expression of the driver genes between the TGCT subtypes form a biologic chain for the TGCT tumorigenesis.
The OSLN/OSK panels change dramatically in expression as SE progresses to EC and TER (Figure 2b). Environmental factors regulate gene expressions in TGCT through a KIT/KITL signal transduction pathway [64].
Normal germ cells express PRAME and LDHC, the gene for the subunit of a testis specific lactate dehydrogenase isoenzyme, LDH-C [65,66,67], but not KIT and NANOG. Normal germ cells, GCNIS, and SE express PRAME but not NST [68,69,70]. GCNIS expresses PRAME, KIT, and NANOG but not LDHC. Normal male germ cells express RBMY1A, whereas the gene is silenced in SE and EC [71].
GCNIS and embryonic stem cells have similar gene expressions [72,73,74]. In GCNIS, POU5F1, NANOG, SOX17, LIN28, and KIT were among the twenty genes with the highest gene expression. Invasion as GCNIS progresses to MGCT is associated with i (12p) [75,76], downregulated CDKN1A (p21), and upregulated MDM2 [77]. SE highly expresses POU5F1 [78,79,80]. Microenvironment factors such as TGF-beta, EGF, and FGF4 support the transition from TCam-2 to EC [81].
The development from the SE expression of SOX17 to the EC expression of SOX2 is part of the iPSC changes between the TGCT subtypes (Figure 2c) [82]. The changes show TGCT has a cellular plasticity. SE has OCT4 and SOX17 as partners, and EC has OCT4 and SOX2 as partners [7]. NANOG is not expressed in normal testicular germ cells, highly expressed in SE and EC, and silenced in YST, CC, and TER. Moreover, SE and EC overexpress POU5F1 and NANOG whereas TER does not [82].
EC and YST highly express the NODAL co-receptor CRIPTO and GCNIS, SE, and CC have a lower expression [83]. Immunohistochemistry links AFP in NST in patients to S-AFP [84] and links hCG in TGCT in patients to S-hCG [85].
Retinoic acid stimulates EC to differentiate into TER. The differentiation is associated with downregulated POU5F1 leading to downregulated PUMA and NOXA and reduced apoptosis [86].
3. Epigenetics and TGCT
3.1. Tumorigenesis
The TGCT tumorigenesis is also governed by epigenetics. Epigenetic regulation of genes in iPSC panels is important for the similarity between embryonic stem cells (ESC) and EC [87]. Normal germ cell tissue and TGCT subtypes differ in epigenetics [57]. Normal tissues have imprinted genes and methylated LINE1 and Alu elements in the DNA string whereas both SE and NST have unmethylated LINE1. SE has unmethylated Alu elements whereas NST has methylated Alu elements. TGCT expresses TET1 and the thymine DNA glycosylase TDG supporting that TGCT demethylate methylated DNA using an oxidative pathway [88]. MiR-223-3p regulates TGCT growth and apoptosis [89].
Of the TGCT precursors, GCNIS expresses DNMT1 and has generally hypomethylated DNA. GCNIS had elevated H2A.Z, mono-, di-, and trimethylated histone 3 lysine 4 (H3K4me1, H3K4me2, and H3K4me3) and H3K9me2, and acetylated histone 3 lysine 9 (H3K9ac), and low H3Kme2 and H3K29me2 [90,91]. In GCNIS, hypomethylation is not solely due to lack of demethylation. Regarding demethylation, GCNIS does not have the methylcytosine dioxygenases TET1 and TET2, so GCNIS uses an AID/APOBEC1 pathway for the demethylation [90].
Of TGCT subtypes, SE had elevated H3K4me1 and H3K9me2 and low H3K4me2, H3K4me3, H3K9ac, and H3K27me3. In SE, an OCT4/SOX17 complex binds to a compressed motif CTTTGTATAAAT [92]. The SOX2 promoter has H3K27me3 and polycomb complex 2, and suppresses SOX2 [93]. Further in SE, the compressed motif together with other OSKM genes hinder differentiation of SE and gives SE a poised pluripotency. In malignant germ cells, the canonical motif is located in enhancers of genes for pluripotency whereas the compressed motif is located in regulatory regions of genes for differentiation into endodermal structures. TGCT expresses HDAC1, HDAC2, and HDAC3, where CC has a high HDAC2 [94].
TCam-2, a SE cell type, does not express SOX2 [93,95]. TCam-2 has elevated H3K4me3 and H3K27ac for SOX17 and low H3K4me3 and H3K27ac for SOX2 whereas NCCIT, an EC cell line derived from a mediastinal germ cell tumor, has elevated H3K4me3 and H3K27ac for SOX2 and low H3K4me2 and H3K27ac for SOX17 [96,97]. TCam-2 expressed SOX17 more than NCCIT. In TCam-2, elimination of an upstream region of SOX17 downregulated POU5F1, NANOG, and LIN28 [7].
TCam-2 xenotransplanted to the abdominal flank in mice upregulate NODAL to express EC genes [97]. The upregulation of NODAL was mediated through a hypomethylated DNMT3B, GDF3, DPPA3, SOX2, LIN28, and ZIC3.
SE had <1% methylations of CpG/CpH sites whereas EC had frequent CpH methylations [51]. Comparing SE and EC, DNMT3B and TET2 were the genes that differed most in expression [98]. EC has moderate levels of H3K4me2 and H3K4me3 and no H3K9me2 and H3K27me3.
In EC, an OCT4/SOX2 complex binds to a canonical motif CTTTGTCATGCAAT [7,92]. It is located in enhancers of pluripotency genes whereas the compressed motif is located in regulatory regions of genes for differentiation into endoderm structures.
In TCam-2, SOX17 bound more to the compressed motif of OCT4 and SOX2 than to the canonical motif whereas in 2012p, a NST cell type, SOX2 bound equally well to the canonical and compressed motifs. The canonical motif activates GDF3, POU5F1, SALLA, SOX2, and TP53.
3.2. DNA Methylations
The 5-C methylation of cytosines in the DNA string changes the chromatin structure in DNA from heterochromatin to euchromatin and opens it for gene expression (Supplementary Figure S1). The level of DNA methylation increases as GCNIS progresses through undifferentiated TGCT into differentiated NST. Comparing the TGCT subtypes, the expression of TET was inversely associated with DNA methylation [82]. GCNIS and SE did not stain for 5-mC, whereas EC had a moderate staining, and YST, CC, TER, drug-resistant SE, TCam-2, and NS cell lines NT2, 2012p, and NCCIT stained homogenously positive [102]. In TGCT, a methylated gene body increases expression of the gene whereas a methylated promoter of the gene inhibits expression of the gene [103].
GCNIS expresses both DNMT1, DNMT3A, and DNMT3B [90]. For restitution, GCNIS expresses a base excision repair (BER) protein that makes cells reestablish cytosines at abasic sites where the cytosines previously had been eliminated [104]. GCNIS has an extremely low level of 5-mC [20].
SE overexpresses TET1, has low proportions of 5-mC and 5-mhC, and no CpG methylations at CpG sites [82]. SE upregulated TET1, TET2, and TET3 [82,88]. SE and EC had median 17% and 31% complete methylations of CpG sites in the upstream region for POU5F1, whereas YST and TER had the methylations in three quarters of the CpG sites [105]. SE and differentiated NST expressed MYC, but differed in gene networks and miRNA [106].
TCam-2 expressed TET2 more than the NST cell lines NT2, 2012p, and NCCIT [98]. Compared with SE, EC had significantly more methylations of promoters of the tumor suppressor genes MCAM (p < 0.0005, chi2 test), MGMT (p < 0.0005), MLH1 (p = 0.004), S100 (p < 0.0005), VGF (p = 0.011, and FKBP4 (p < 0.0005) [107]. EC expresses DNMT3B and EHMT2 more than SE [105]. EC has hypermethylated promoters in 40 downregulated genes [71]. Transfection of micro-RNA 630 into a NT2/D2 EC cell line targeting the 3′UTR of POU5F1, SOX2, and NANOG inhibited the expression of the genes [108]. TGCT has methylated repetitive elements in the DNA strand [57]. Differentiated NST had CpG methylation similar to solid cancers [82].
3.3. Histone Modifications and Modulations
Methylations and acetylations of histone 3 lysines change DNA structures from hetrochromatin to euchromatin and opens them for gene expression (Supplementary Figure S2a,b). Histone modifications also contribute to the TGCT tumorigenesis. In normal germ cells, upregulation of E2F1 in the CCND2/RB1 signal transduction pathway gave rise to atypical mitosis and cell atypia like that in GCNIS [109].
EC had H3K4 and H3K9 methylations [110]. EZH2, enhancer of Zeste-2, is a histone methyltransferase (“writer”) and a catalytic subunit in the polycomb repressor complex 2 (PRC2). EZH2 methylates H3K27, condenses chromatin structure, and silences tumor suppressor genes CDKN2A (p16(INK4)) and RB1 (Figure 3a). Silencing EZH2 with siRNA reduced cellular DNA replication and cell growth. A lysine specific demethylase 1A, KDM1A/LSD1, demethylates H3K4me2 [111].
H3K modifications differ by their binding sites on the genes as they regulate expression of genes. H3K4me3 and H3K27ac activate enhancers whereas H3K4me1 poises enhancers (Figure 3a). Akt-phosphorylated OCT4 stabilized OCT4, facilitated its nuclear localization, and its interaction with SOX2 [112].
Normal testicular tissue highly expresses EZH2 whereas the expression is lower in GCNIS, and further reduced in invasive TGCT [113]. In EC cells, a H3K36 methyltransferase SETD2 trimethylated H3K9, H3K27, and H3K36 on sites in OCT4, SOX2, and NANOG [114]. In 2102p, knockdown of DNMTB increased H3K27me3, EZH2 expression, and sensitivity to cisplatin [114]. TGCT has an antagonism between the regulation with H3K4me3 and H3K27me3. The antagonism gives TGCT a cellular plasticity and supports pluripotency. In POU5F1- and SOX2-positive EC cells, inhibition of LSD1 reduced proliferation whereas LSD1 inhibition of POU5F1- and SOX2-negative EC cells did not change the proliferation [111].
TER moderately expresses H3K4me2, H3K4me3, H3K9me1, H4K9me2, and H3K9me3, whereas YST strongly express the histone marks.
Combined, H3K4me3 and H3K27me3 in promoters of genes generate poised/bivalent signals for expression of genes [115].
3.4. Driver Genes
Epigenetics regulate driver genes in TGCT (Table 1). TCam-2 have low levels of H3K4me3 and H3K27ac whereas NST cell lines have high levels [96]. The driver genes POU5F1, SOX2, LIN28, and NANOG form a network of genes [116]. Upstream of the gene body, POU5F1 has four conserved regions that can bind SOX2, NANOG, and HIF2A (HIF-2ꬰ, EPAS1) [117]. Dimethylated promoter for POU5F1 increased expression of POU5F1, whereas reduced methylation reduced expression of the gene [110]. OCT4 binds to the ATTTTGCAT motif.
POU5F1 is expressed in most GCNIS and all SE and EC [118]. NST cell lines have a network between DNMT3B and POU5F1 [119]. Many transcription factors may be involved with the SOX2-OCT4 connection.
TCam-2 has low H3K4me3 and H3K27ac whereas NST cell lines have elevated levels. In EC cells, overexpression of POU5F1 inhibited the expression of NANOG but not the expression of FGF4 and UTF1 [120]. In malignant germ cells, ACT-driven phosphorylation of OCT4 stabilizes OCT4, stimulates its interaction with SOX2, and increases cell survival [112]. Many transcription factors may be involved in the SOX2-OCT4 connection. In EC cell lines, LSD1 inhibition downregulated OCT4 and SOX2 [121].
In NCCIT cells, siRNA downregulation of POU5F1 reduced expression of SOX2, LIN28, and NANOG, and upregulated genes for differentiation, such as OTX1, HAND1, and LAMB1 [123]. In EC cell lines, miRNA-27 inhibited expression of POU5F1 and made the cells differentiate [124]. Downregulation of OCT4 increased miR-27, and the increase of miR-27 down-regulated NANOG [125]. Treatment of an EC cell line with an LSD1 inhibitor downregulated OCT4 and SOX2 [121].
SOX17 is hypomethylated in EC [126]. Deletion of SOX17 reduced expression of POU5F1 [7]. In malignant germ cells, the switch from SOX17 to SOX2 determines whether cells function as SE or as EC [127].
Deletion of SOX17 reduced expression of POU5F1. TCam-2 had an elevated H3K4me1 in SOX17 whereas NCCIT had a low level [128]. Methylation of the SOX17 promoter downregulated expression of SOX17 [129]. The SOX17 promoter has 420 base pairs and 48 GpC sites [130].
SOX2 is suppressed in SE by the polycomb repressive complex and H3K27me3 [93]. SE and EC has a hypomethylated SOX2 whereas SOX2 in TER has a higher methylation [131]. TCam-2 needs expression of SOX2 for being reprogrammed into EC [7].
NST cell lines have a network between DNMT3B and POU5F1.
EC expressed SOX2 three times more than TCam-2. In NCCIT cultures, silencing of SOX2 gave prominent cell death within three days [123].
LIN28 has a role in the TGCT tumorigenesis [132]. A distal enhancer of LIN28 has three binding sites for OCT4 [133]. LIN28 mediates effects by downregulated LET-7 microRNA [134]. SiRNA downregulated LIN28 and reduced expression of SOX2 and NANOG. In P19 malignant germ cells, silencing of LIN28 reduced proliferation and tumor formation [132].
Regarding NANOG, Nettersheim et al. denoted a region upstream the transcription start site the NANOG regulatory region (NRR) [135]. It has binding elements for OCT4 and SOX2 [136,137]. Binding of the transcription factors to NRR increases the expression of NANOG whereas methylation of NRR inhibits the transcription factors which are bound to NRR and inhibits the expression of NANOG. GCNIS, SE, and EC express NANOG whereas TER does not [138]. TGCT cell lines with unmethylated NRR express NANOG whereas cell lines with methylated NRR do not. In EC cells, two transactivating domains in the C-terminal region of NANOG mediate expression of NANOG [139].
3.5. Epidrugs and TGCT
Drugs that inhibit epigenetic regulation of gene expression are denoted epidrugs. They have been studied in TGCT (Table 2). In a cohort study of treatment with the methyltransferase (“writer”) inhibitor 5-azacytidine (5-aza), 2 of 4 TGCT patients had partial remission [140]. Another study reported that one of 17 patients with TGCT treated with 5-aza had no evidence of disease at the end of follow-up [141]. In TCam-2, 5-aza increased expression of POU5F1 and NANOG. Trichostatin works synergistically with retinoic acid [142].
EORTC conducted a trial of NST patients treated with the DNA methylation inhibitor 5-aza-2′-deoxycytidine (5-aza-dC) [145]. The response to 5-aza-dC depended on the cellular expression of POU5F1 and DNMT3B. Treatment with 5-aza-dC increased the expression of DNMT3B. For patients with cisplatin-resistant metastatic TGCT, treatment with cisplatin and an inhibitor of DNA methyltransferase, guadecitabine, did not give long-lasting complete remissions [143].
Epidrugs for TGCT have mainly been evaluated in cell cultures of malignant germ cell lines because most patients with metastatic TGCT are cured with chemotherapy. In the TGCT cell lines NCCIT and 2012p, 5-aza had long-lasting effects [144]. TGCT cell lines have an inverse relation between sensitivity to 5-aza and sensitivity to cisplatin [115]. In EC cells, sensitivity to 5-aza demanded a highly expressed DNMT3B [146,147].
In 1994, Jutterman et al. were the first to report treatment with 5-aza-dC [148]. They found that 5-aza stimulated TCam-2 further and upregulated POU5F1 and NANOG. Three EC cell lines, including Tera-2, were treated with 5-aza [149], and 5-aza-dC demethylated the promoter of POU5F1 and reduced expression of POU5F1. Treatment with low dose 5-aza-dC demethylated promoters for SOX2, NANOG, and MYC [146]. In two NS cell lines, NCCIT and Tera-2, treatment with 5-aza-dC reduced H3K4me2 in the promoter of POU5F1 and inhibited expression of POU5F1.
Regarding inhibitors of histone 3 modifications, treatment of an ovarian germ cell tumor cell line PA-1 with an inhibitor of the histone methylase LSD1, CBB3001, downregulated OCT4 and SOX2 and inhibited tumor growth [121].
Treatment of the EC cell line P19 with a histone lysine demethylase (KDM) inhibitor, tranylcypromine, inhibited a histone demethylase, increased H3K4 methylation, and expression of POU5F1 [150]. In addition, a chimeric inhibitor animacroxam was effective [151].
In cisplatin-sensitive and cisplatin-resistant EC cell lines, two inhibitors of histone lysine deacetylase (HCAD), belinostat and panobiostat, were effective [152]. In NST cell lines, also the HDAC inhibitor depsipeptide was effective [153]. A bromodomain inhibitor, JQ1, binds to the amino-terminal twin of bromodomains of BET proteins that bind to acetylated histone lysines. In an EC cell line, JQ1 downregulated pluripotency factors and caused the cells to differentiate into mesodermal structures [154].
Treatment with the HDAC1 inhibitor, romidepsin, increased H3K acetylations and decreased ARI1A and thereby expression of POU5F1, SOX2, LIN28, and NANOG [155]. Treatment of the EC cell line P19 with the HDAC inhibitor trichostatin (TSA) inhibits cell progression [121]. A study compared treatment of germ cell tumor cell lines with seven epidrugs [156].
The CDK inhibitor YKL-5-124 inhibited both TCam-2 and 2102p whereas SY0351 and NVP2 gave cell line-specific responses [157].
4. iPSC in Oncology
4.1. TGCT
We confirmed and expanded the role iPSC has for the tumorigenesis of TGCT based on epigenetic regulations of driver genes, not least genes in iPSC panels. Epigenetic mechanisms regulate the shift from an OSKM-like panel in SE to an OSLN panel in EC, and the silencing of the OSLN panel in TER. iPSC genes form a biologic background for the genetic and histologic changes between the TGCT subtypes. DNA methylations and histone modifications regulate iPSC genes. Driver genes reprogram malignant germ cells to pluripotency, increase unlimited self-renewal and independency of growth factors, and inhibit the suppression of proliferation, all hallmarks of malignancy [158].
In 1946, Friedman and Moore were first to point out that EC has a unique toti-differentiation potential which is crucial for the TGCT tumorigenesis [159]. In 1980, Pugh and Parkinson commented on the tumorigenesis and classification of TGCT and noted that clinicians generally lump the TGCT subtypes as SE and NS whereas pathologists generally split TGCT in subtypes [160]. Our review categorizes all TGCT subtypes as a continuum of progression. Correspondingly, oncologists generally treat patients with stage 1 SE and NST with active surveillance, and treat patients with metastatic SE and NS with BEP chemotherapy.
The evidence regarding the OSKM and OSLN panels motivates an iPSC hypothesis for the TGCT tumorigenesis. The first premise is that iPSC panels can reprogram cells to iPSC and that only iPSC and EC can differentiate into TER. A second premise is that OSKM and OSLN panels can reprogram both normal somatic cells and malignant germ cells into an iPSC/induced totipotent stem cell pattern. Thus, iPSC panels can reprogram cells with normal and abnormal ploidy into iPSC/induced toti-potential cells.
A third premise is that EC expresses the OSLN panel and is able to differentiate into YST, CC, and TER whereas normal somatic cells reprogrammed with the OSKM panel to iPSC only are able to differentiate into TER. Accordingly, the cellular background before reprogramming for iPSC determines the frames for differentiation of IPSC/toti-potential cells.
GCNIS develops into MGCT within median five years [161]. The median age at diagnosis for SE patients is 35 years and 25 years for NST patients, so it is more likely that thetumorigenesis of TGCT starts during puberty than during fetal life.
The regional site for xenotransplants of TCam-2 to mice is crucial for progression to EC [8]. The fact points out that the microenvironment governs expression of genes in iPSC panels, mediated by epigenetic mechanisms. Thus our review add an epigenetic dimension to the “genvironmental” hypothesis for the TGCT tumorigenesis proposed by Looijenga [162].
Many observations support the progression from SE to EC. An autopsy series of patients with primary SE showed that 63 (44%) of 145 patients had NST metastases at the time of death [163].
Epigenetics have a key role for the TGCT tumorigenesis, and gene mutations have a limited role. The Genomic Cancer Atlas (TGCA) study of patients with TGCT reported that of TGCT subtypes, only SE had mutations in KIT, KRAS, and NRAS [51]. The shifts in gene expression associated with progression between the TGCT subtypes exclude that mutations in general regulate the expression of the driver genes and the progression of the TGCT subtypes.
The iPSC panels also govern clinical characteristics between the TGCT subtypes. The low SOX2 expression in SE and high SOX2 expression in EC explain why SE mostly is the only TGCT subtype in SE tumors, and why NST TGCT most often incorporates more than one NST subtypes. The transition from SE to EC explains why TGCT often combine SE and NST elements.
The pattern of gene expression also explains why SE proliferates more slowly than EC [164], and why patients with SE have a median age at diagnosis that is ten years higher than that for patients with NST. Furthermore, the switch from SOX17 to SOX2 between the TGCT subtypes explains why SE responds better to platin-based chemotherapy than EC, and why SE is sensitive to radiation therapy in contrast to the radiation-insensitivity of NST subtypes [165]. As EC differentiates into TER, the switch in expression of iPSC genes contributes to the cisplatin-resistance in TER.
Our review includes three driver genes with gene locus on chromosome 12. In addition, reprogramming of TCam-2 to EC also upregulated three genes DPPA3, GDF3, and BCAT1 [97], that also have their gene locus on chromosome 12p.
Several gene networks collaborate in the TGCT tumorigenesis. We report that IPSC genes work synergistically in a network. A second network is the CCND2/RB1 signal transduction pathway, and a third network is genes on the short arm of chromosome 12 associated with anaerobic glycolysis [166]. POU5F1 also participates in a fourth network that links macro-environmental factors such as retinoic acid and cisplatin to a downregulated POU5F1, causing downregulated NOXA and PUMA, and reduced apoptosis [86].
The link between i (12p) and extratubular invasion of malignant germ cells implies that CCND2, NANOG, and LDHB (with gene loci on chromosome 12p) are more important for the progression between the TGCT subtypes than KITL and CDK4 (with gene loci on chromosome 12q).
National and international studies support that S-AFP, S-hCG, and S-LDH are prognostic indicators for TGCT. One of the authors (FEvE) was first to report the prognostic value of S-LDH in 1978, and two multivariate analyses later confirmed the prognostic value [167,168,169]. The prognostic value of S-LDH was accepted worldwide after the international large collaborative study in 1997 [26]. Recently, the Global Germ Cell Tumor Collaboration documented the prognostic value of S-LDH for patients with advanced SE (hazard ratio 2.9, p = 0.003) and patients with NS with intermediate prognosis (hazard ratio 2.6, p < 0.001) [170].
As TGCT suppresses LDHC, patients with TGCT and elevated S-LDH do not have detectable S-LDH isoenzyme C [171]. Of present serum tumor markers, only S-LDH-1 is generated from a gene with locus on the short arm of chromosome 12 [172,173,174,175].
DATECA studies showed that patients with SE and NST stage 1 had similar good outcomes whether they were treated with radical orchiectomy followed with active surveillance or with adjuvant radiation therapy to pelvic and retroperitoneal lymph nodes [176,177]. Complementarily, DATECA publication of SE and NST TGCT stage 1 followed with surveillance reported that half of the patients had a raised S-LDH-1 before orchiectomy [178,179]. For the NS TGCT patients, a raised S-LDH-1 was associated with a reduced relapse-free survival (p = 0.003, log-rank test).
Clinically, most patients with metastatic TGCT are cured with a combination of chemotherapy regimen with bleomycin, etoposide, and (cis)platin (denoted BEP) [180]. In addition, some patients are cured with surgery for post-chemotherapy residual TER lesions. Some patients who do not achieve a complete remission with four courses of BEP can be cured with high-dose cisplatin-based chemotherapy [181]. Impressively in recent years, the good five-year overall survival for patient with TGCT improved further [182,183]. The improvement is due to the shift in combination chemotherapy from vinblastine to etoposide in the BEP regimen. For patients with high-risk metastatic TGCT who fail on standard BEP chemotherapy, the decay of two serum tumor markers, S-AFP and S-hCG, pointed to the cisplatin-resistance as early as after the first course of BEP [29,184]. Overall, the prognostic value of the three serum tumor markers is documented in prospective follow-up studies of more than 10,000 patients with TGCT [26,170,182,183].
It is a challenge that patients with metastatic TGCT given a high cumulative dose of cisplatin (>635 mg) have a mortality that increases dramatically after follow-up for ten to twenty years [185]. Accordingly, oncologists might consider to study new systemic treatments as alternative to high-cisplatin-dose chemotherapy.
4.2. Other Malignancies
The driver genes have broad implications in oncology. Small cell lung cancer (SCLC) and small cell esophageal cancer (SCEC) have elevated SOX2 and low RB1 and the two genes reduce inhibition of proliferation [186,187,188]. Synergy between the two genes explains why SCLC, SCEC, and TGCT grow faster than most other malignancies. Other malignancies also highly express the iPSC panels [189]. iPSC panels, especially SOX2 and the OSN panel, are highly expressed in cancer stem cells of other malignancies [189,190,191,192].
POU5F1, SOX2, and NANOG are important for the tumorigenesis of skin, oral squamous, esophageal, lung, and colon cancer [193,194]. SOX2 contributes to the development of brain tumors [195]. NANOG is also an important mediator of cancer induction [196]. For prostate cancer, high expression of SOX2 reduces expression of RB1 and TP53 and changes the cancer from a luminal androgen-receptor-dependent subtype to a basal androgen-receptor-independent subtype [187]. Based on the morphology in prostate cancer, a Gleason score contributes to treatment decisions and predicts outcome of treatment [197]. The increase in Gleason score is denoted as dedifferentiation.
SCLC downregulates both RB1 and TP53 whereas TGCT only downregulates RB1. The difference contributes to different outcomes where most patients with metastatic TGCT are cured with BEP chemotherapy whereas most patients with metastatic SCLC-who despite temporary response to EP-based chemotherapy-progress and die of the cancer.
So far, the United States of America Federal Drug Administration (FDA) has approved seven epidrugs such as 5-aza, 5-aza-cD, belinostat, paninostat, PXD-101, romidepsin, and vorinostat as routine treatment for cutaneous T cell lymphoma, chronic myelomonocytic leukemia (CMML), and multiple myeloma, respectively (Table 2) [198,199,200,201,202,203,204]. A phase III trial of patients with myelodysplastic syndrome showed that 5-aza increased overall survival substantially (hazard ratio 0.58, p = 0.0001, log-rank test) [205]. The FDA also approved a CDK2/CDK6 inhibitor trilaciclib for patients with advanced SCLC treated with EP-containing regimens [206].
In contrast to TGCT, some other malignancies have mutation-based increases in gene expression that are more important than non-mutational increases. Patients with non-small cell lung cancer (NSCLC) with mutated epidermal growth factor receptor (EGFR) respond better to targeted systemic treatment with EGFR tyrosine kinase inhibitors (EGFR-TKI) than patients with a high non-mutational expression of EGFR [207]. Today, the best treatments of patients with NSCLC with activating EGFR mutations are third-generation EGFR TKI rosimertinib and a combination of first-generation EGFR TKI and chemotherapy [208].
5. Perspectives and Conclusions
Continued research may further expand the knowledge of epigenetic regulation of driver genes in oncology and of the TGCT tumorigenesis. In reprogramming cells for iPSC, other genes can substitute for the genes we report in our review [209,210]. These genes may also be relevant in TGCT tumorigenesis. Thus, the number of relevant driver genes in oncology and in the TGCT tumorigenesis may increase in future. New insight in regulation of TGCT driver genes may also be relevant for other malignancies.
Innovative studies of driver genes in many malignancies have general oncologic relevance. For women with hormone-sensitive breast carcinoma, present epidrugs enhance effects of hormone treatment but also increase adverse effects [211]. The findings motivate new epidrugs with fewer adverse effects.
Investigations of epidrugs in TGCT cell lines have been inspired by the clinical achievements with epidrugs in other malignancies. Recent trials support combinations of epidrugs. For elder patients with acute myeloid leukemia, treatment with a combination of 5-aza and the BCL2 inhibitor venetoclax gave a better overall survival than monotherapy with only 5-aza [212]. Another study showed a synergy between a DNMT inhibitor and a HDAC inhibitor [213].
In addition to serum tumor markers, liquid biopsies monitoring hypermethylated driver genes may improve follow-up routines of patients with TGCT and other malignancies [214].
In TGCT, downregulated tumor suppressor gene RASSF1A increases proliferation [110,215]. TGCT also has a downregulated RB1 [216]. It remains to be shown whether the two tumor suppressor genes act in synergy. A recent review pointed to three genes with gene locus on the short arm of chromosome 12, GLUT3, GAPDH, and TPII, that participate in anaerobic glycolysis [166]. The question remains whether LDHB and the three genes collaborate for a Warburg effect in TGCT [217,218].
For patients with TGCT, the target and timing may increase the efficacy of epidrugs. New epidrugs might specifically target the motifs of the combinations of OCT4 and SOX2/SOX17. As for more upfront timing, new trials may recruit patients with high-risk metastatic NS and limited decay of S-AFP and S-hCG after the first course of BEP. Such trials can investigate the efficacy of BEP combined with relevant candidate drugs as alternative to high-dose cisplatin chemotherapy (Figure 4).
In conclusion, genes that reprogram for iPSC have a major role in the TGCT tumorigenesis, and contribute to the aggressiveness in many other malignancies.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms24044148/s1.
Author Contributions
Conceptualization: F.E.v.E. and F.F.N. methodology: K.K. conducted research on gene expression of LDHB in TGCT. R.H. contributed on presentation of MGCT. O.P. conducted immunohistochemistry for EC. writing the initial draft: F.E.v.E. rewriting: F.E.v.E., D.S.K. and F.F.N. funding acquisition: F.E.v.E. supervision: D.S.K., F.F.N. and K.K. All authors have read and agreed to the published version of the manuscript.
Funding
Minister Erna Hamiltons Foundation for art and sciences.
Acknowledgments
The authors thank Rachel Eiges. Shaale Zetek Medical Center, Jerusalem, Israel. [email protected], for a figure of iPSC for the published case of iPSC for a 32 year old man with SE. The authors are very grateful for a grant from minister Erna Hamilton’s foundation for art and science.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AFP | alfa fetoprotein |
| 5-aza | 5-azacytidine |
| 5-aza-cD | 5-aza-2′-deoxycytidine |
| BEP | bleomycin, etoposide, and (cis)platin |
| bromodomaini | bromodomain inhibitor |
| CC | choriocarcinoma |
| CDK | cyclin D kinase |
| CDKi | CDK inhibitor |
| CYT | spermatocyte |
| DNMT | DNA methyltransferase |
| DATECA | the Danish testis cancer study group |
| DNMT | DNA methyl transferase |
| DNMTi | DNA methyltransferase inhibitor |
| EC | embryonal carcinoma |
| EGFR | epidermal growth factor receptor |
| ESC | embryonic stem cells |
| EZH2 | enhancer of Zeste-2 |
| EZH2i | EZH2 inhibitor |
| GCNIS | germ cell neoplasia in situ |
| GON | gonocytes |
| HAT | histone acetylase |
| hCG | human chorionic gonadotropin |
| HDAC | histone deacetylase |
| HDACi | histone deacetylase inhibitor |
| HDMT | histone demethylase |
| HMT | histone methyltransferase |
| iPSC | induced pluri- to totipotent stem cells |
| KDM | lysine demethylase |
| KDMi | lysine demethylase inhibitor |
| Kme | methylated histone 3 lysine |
| LDH | Lactate dehydrogenase |
| LSD1i | LSD1 inhibitor |
| 5mC | 5-methyl cytosine |
| 5mhC | 5-methylhydroxy cytosine |
| MGCT | microinvasive germ cell tumor |
| NGC | normal germ cells |
| NSCLC | non-small cell lung cancer |
| NST | non-seminomatous germ cell tumor |
| OCT4 | the protein of POU5F1 |
| OSKM | the OCT4, SOX2, KLF4, and MYC panel |
| OSLN | the OCT4, SOX2, LIN28, and NANOG panel |
| PGC | primordial germ cells |
| PRC2 | polycomb repressor complex 2 |
| SCLC | small cell lung cancer |
| SCEC | small cell esophageal cancer |
| SE | seminoma |
| SPA | spermatogonia type A |
| SPB | spermatogonia type B |
| TER | teratoma |
| TET | DNA demethylase |
| TGCA | The Genomic Cancer Atlas |
| TKI | tyrosine kinase inhibitor |
| TNM | international tumor, nodes, and metastases classification |
| TSS | transcription start site. |
| WHO | World Health Organization |
| YST | yolk sac tumor |
References
- Waddington, C.H. Evolutionary adaptation. Perspect. Biol. Med. 1959, 2, 379–401. [Google Scholar] [CrossRef]
- Yu, J.; Vodyanik, M.A.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J.L.; Tian, S.; Nie, J.; Jonsdottir, G.A.; Ruotti, V.; Stewart, R.; et al. Induced pluripotent stem cell lines derived from human somatic cells. Science 2007, 318, 1917–1920. [Google Scholar] [CrossRef]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez-Aranda, I.; Ramos-Mejia, V.; Bueno, C.; Munoz-Lopez, M.; Real, P.J.; Macia, A.; Sanchez, L.; Ligero, G.; Garcia-Parez, J.L.; Menendez, P. Human induced pluripotent stem cells develop teratoma more efficiently and faster than human embryonic stem cells regardless the site of injection. Stem Cells 2010, 28, 1568–1570. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.-H.; Hang, W.; Hu, R. Exclusively intertubular seminoma arising in undescendedn testes: Report of two cases. Hum. Pathol. Case Rep. 2018, 11, 15–18. [Google Scholar] [CrossRef]
- Von Eyben, F.E.; Parraga-Alava, J. Meta-analysis of gene expressions in testicular germ cell tumor histologies. Int. J. Mol. Sci. 2020, 21, 4487. [Google Scholar] [CrossRef]
- Jostes, S.V.; Fellermeyer, M.; Arevalo, L.; Merges, G.E.; Kristiansen, G.; Nettersheim, D.; Schorle, H. Unique and redundant roles of SOX2 and SOX17 in regulating the germ cell tumor fate. Int. J. Cancer 2020, 146, 1592–1605. [Google Scholar] [CrossRef] [Green Version]
- Nettersheim, D.; Heimsoeth, A.; Jostes, S.; Schneider, S.; Fellermeyer, M.; Hofmann, A.; Schorle, H. SOX2 is essential for in vivo reprogramming of seminoma-like TCam-2 cells to an embryonal carcinoma-like fate. Oncotarget 2016, 7, 47095–47110. [Google Scholar] [CrossRef] [Green Version]
- Nonaka, D. Differential expression of SOX2 and SOX17 in testicular germ cell tumors. Am. J. Clin. Pathol. 2009, 131, 731–736. [Google Scholar] [CrossRef]
- Eldar-Geva, T.; Gross-Tsur, V.; Hirsch, H.J.; Altarescu, G.; Segal, R.; Zeligson, S.; Golomb, E.; Epsztejn-Litman, S.; Eiges, R. Incomplete methylation of a germ cell tumor (Seminoma) in a Prader-Willi male. Mol. Genet. Genomic Med. 2018, 6, 811–818. [Google Scholar] [CrossRef]
- Schulz, W.A.; Hoffmann, M.J. Transcription factor networks in embryonic stem cells and testicular cancer and the definition of epigenetics. Epigenetics 2007, 2, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Nogales, F.F.; Prat, J.; Schuldt, M.; Cruz-Viruel, N.; Kaur, B.; D’Angelo, E.; Matias-Guiu, X.; Vidal, A.; McCluggage, W.G.; Oosterhuis, J.W. Germ cell tumour growth patterns originating from clear cell carcinomas of the ovary and endometrium: A comparative immunohistochemical study favouring their origin from somatic stem cells. Histopathology 2018, 72, 634–647. [Google Scholar] [CrossRef]
- Oosterhuis, J.W.; Looijenga, L.H.J. Human germ cell tumours from a developmental perspective. Nat. Rev. Cancer 2019, 19, 522–537. [Google Scholar] [CrossRef]
- Wilms, M. Die teratoiden Geschwülste des Hodens, mit einschluss der sogenante Cystoide und Encondrome. Beitr. Pathol. Anat. 1896, 19, 233–366. [Google Scholar]
- Atkin, N.B.; Baker, M.C. Specific chromosome change, i(12p), in testicular tumours? Lancet 1982, 2, 1349. [Google Scholar] [CrossRef]
- Castedo, S.M.; de Jong, B.; Oosterhuis, J.W.; Seruca, R.; te Meerman, G.J.; Dam, A.; Schraffordt Koops, H. Cytogenetic analysis of ten human seminomas. Cancer Res. 1989, 49, 439–443. [Google Scholar]
- Castedo, S.M.; de Jong, B.; Oosterhuis, J.W.; Seruca, R.; Idenburg, V.J.; Dam, A.; te Meerman, G.; Koops, H.S.; Sleijfer, D.T. Chromosomal changes in human primary testicular nonseminomatous germ cell tumors. Cancer Res. 1989, 49, 5696–5701. [Google Scholar]
- Castedo, S.M.; de Jong, B.; Oosterhuis, J.W.; Idenburg, V.J.; Seruca, R.; Buist, J.; te Meerman, G.J.; Schraffordt Koops, H.; Sleijfer, D.T. Chromosomal changes in mature residual teratomas following polychemotherapy. Cancer Res. 1989, 49, 672–676. [Google Scholar]
- Fichtner, A.; Richter, A.; Filmar, S.; Gaisa, N.T.; Schweyer, S.; Reis, H.; Nettersheim, D.; Oing, C.; Gayer, F.A.; Leha, A.; et al. The detection of isochromosome i(12p) in malignant germ cell tumours and tumours with somatic malignant transformation by the use of quantitative real-time polymerase chain reaction. Histopathology 2021, 78, 593–606. [Google Scholar] [CrossRef]
- von Eyben, F.E. Chromosomes, genes, and development of testicular germ cell tumors. Cancer Genet. Cytogenet. 2004, 151, 93–138. [Google Scholar] [CrossRef]
- Zondag, H.A. Enzyme activity in dysgerminoma and seminoma. A study of lactic dehydrogenase isoenzymes in malignant diseases. R. I. Med. J. 1964, 47, 273–281. [Google Scholar]
- von Eyben, F.E.; Blaabjerg, O.; Petersen, P.H.; Horder, M.; Nielsen, H.V.; Andersen, S.K.; Parlev, E. Lactate dehydrogenase isoenzyme 1 in testis cancer. Lancet 1987, 2, 1035–1036. [Google Scholar] [CrossRef]
- von Eyben, F.E. A systematic review of lactate dehydrogenase isoenzyme 1 and germ cell tumors. Clin. Biochem. 2001, 34, 441–454. [Google Scholar] [CrossRef]
- von Eyben, F.E.; Parraga-Alava, J.; Tu, S.M. Testicular germ cell tumors type 2 have high RNA expression of LDHB, the gene for lactate dehydrogenase subunit B. Asian J. Androl. 2021, 23, 357–362. [Google Scholar] [CrossRef]
- Wittekind, C. TNM classification of testicular tumors. Definitions and prerequisites for correct application. Pathologe 2014, 35, 252–255. [Google Scholar] [CrossRef]
- International Germ Cell Consensus Classification Group. International Germ Cell Consensus Classification: A prognostic factor-based staging system for metastatic germ cell cancers. J. Clin. Oncol. 1997, 15, 594–603. [Google Scholar] [CrossRef]
- Einhorn, L.H. Testicular cancer: An oncological success story. Clin. Cancer Res. 1997, 3, 2630–2632. [Google Scholar]
- Gurney, J.K.; Florio, A.A.; Znaor, A.; Ferlay, J.; Laversanne, M.; Sarfati, D.; Bray, F.; McGlynn, K.A. International trends in the incidence of testicular cancer: Lessons from 35 years and 41 countries. Eur. Urol. 2019, 76, 615–623. [Google Scholar] [CrossRef]
- Toner, G.C.; Geller, N.L.; Tan, C.; Nisselbaum, J.; Bosl, G.J. Serum tumor marker half-life during chemotherapy allows early prediction of complete response and survival in nonseminomatous germ cell tumors. Cancer Res. 1990, 50, 5904–5910. [Google Scholar]
- Clemmesen, J. Testis cancer incidence—Suggestion of a world pattern. Int. J. Androl. 1981, 4 (Suppl. S4), 111–120. [Google Scholar] [CrossRef]
- von Eyben, F.E.; Jensen, M.B.; Hoyer, S. Frequency and markers of precursor lesions and implications for the pathogenesis of testicular germ cell tumors. Clin. Genitourin. Cancer 2017, 18, E211–E221. [Google Scholar] [CrossRef] [Green Version]
- Krag Jacobsen, G.; Barlebo, H.; Olsen, J.; Schultz, H.P.; Starklint, H.; Sogaard, H.; Vaeth, M. Testicular germ cell tumours in Denmark 1976–1980. Pathology of 1058 consecutive cases. Acta Radiol. Oncol. 1984, 23, 239–247. [Google Scholar] [CrossRef]
- Norgaard-Pedersen, B.; Schultz, H.; Arends, J.; Brincker, H.; Jacobsen, G.K.; Lindelov, B.; Rorth, M.; Svennekjaer, I.L. Biochemical markers for testicular germ-cell tumors in relation to histology and stage: Some experiences from the Danish Testicular Cancer (DATECA) study from 1976 through 1981. Ann. N. Y. Acad. Sci. 1983, 417, 390–399. [Google Scholar] [CrossRef]
- Trabert, B.; Sigurdson, A.J.; Sweeney, A.M.; Amato, R.J.; Strom, S.S.; McGlynn, K.A. Baldness, acne and testicular germ cell tumours. Int. J. Androl. 2011, 34, e59–e67. [Google Scholar] [CrossRef]
- Dixon, F.J.; Moore, R.A. Testicular tumors. A clinicopathological study. Cancer 1953, 6, 427–454. [Google Scholar] [CrossRef]
- Teilum, G. Endodermal sinus tumors of the ovary and testis. Comparative morphogenesis of the so-called mesoephroma ovarii (Schiller) and extraembryonic (yolk sac-allantoic) structures of the rat’s placenta. Cancer 1959, 12, 1092–1105. [Google Scholar] [CrossRef]
- de Jong, B.; Oosterhuis, J.W.; Castedo, S.M.; Vos, A.; te Meerman, G.J. Pathogenesis of adult testicular germ cell tumors. A cytogenetic model. Cancer Genet. Cytogenet. 1990, 48, 143–167. [Google Scholar] [CrossRef]
- Schulze, C.; Holstein, A.F. On the histology of human seminoma: Development of the solid tumor from intratubular seminoma cells. Cancer 1977, 39, 1090–1100. [Google Scholar] [CrossRef]
- von Eyben, F.E.; Jacobsen, G.K.; Skotheim, R.I. Microinvasive germ cell tumor of the testis. Virchows Arch. 2005, 447, 610–625. [Google Scholar] [CrossRef]
- Holstein, A.F.; Schutte, B.; Becker, H.; Hartman, M. Morphology of normal and malignant germ cells. Int. J. Androl. 1987, 10, 1–18. [Google Scholar] [CrossRef]
- Harrison, N.J.; Baker, D.; Andrews, P.W. Culture adaptation of embryonic stem cells echoes germ cell malignancy. Int. J. Androl. 2007, 30, 275–281, discussion 281. [Google Scholar] [CrossRef]
- Ben-David, U.; Benvenisty, N. The tumorigenicity of human embryonic and induced pluripotent stem cells. Nat. Rev. Cancer 2011, 11, 268–277. [Google Scholar] [CrossRef]
- Blum, B.; Benvenisty, N. The tumorigenicity of human embryonic stem cells. Adv. Cancer Res. 2008, 100, 133–158. [Google Scholar]
- Baker, D.E.; Harrison, N.J.; Maltby, E.; Smith, K.; Moore, H.D.; Shaw, P.J.; Heath, P.R.; Holden, H.; Andrews, P.W. Adaptation to culture of human embryonic stem cells and oncogenesis in vivo. Nat. Biotechnol. 2007, 25, 207–215. [Google Scholar] [CrossRef]
- Kawakami, T.; Okamoto, K.; Sugihara, H.; Hattori, T.; Reeve, A.E.; Ogawa, O.; Okada, Y. The roles of supernumerical X chromosomes and XIST expression in testicular germ cell tumors. J. Urol. 2003, 169, 1546–1552. [Google Scholar] [CrossRef]
- Walt, H.; Emmerich, P.; Jauch, A.; DeLozier-Blanchet, C.D. Characterization of precancerous and neoplastic human testicular germ cells. Recent Results Cancer Res. 1991, 123, 37–44. [Google Scholar]
- Bates, S.; Ryan, K.M.; Phillips, A.C.; Vousden, K.H. Cell cycle arrest and DNA endoreduplication following p21Waf1/Cip1 expression. Oncogene 1998, 17, 1691–1703. [Google Scholar] [CrossRef]
- Baldini, E.; Arlot-Bonnemains, Y.; Mottolese, M.; Sentinelli, S.; Antoniani, B.; Sorrenti, S.; Salducci, M.; Comini, E.; Ulisse, S.; D’Armiento, M. Deregulation of Aurora kinase gene expression in human testicular germ cell tumours. Andrologia 2010, 42, 260–267. [Google Scholar] [CrossRef]
- Yan, X.; Cao, L.; Li, Q.; Wu, Y.; Zhang, H.; Saiyin, H.; Liu, X.; Zhang, X.; Shi, Q.; Yu, L. Aurora C is directly associated with Survivin and required for cytokinesis. Genes Cells 2005, 10, 617–626. [Google Scholar] [CrossRef]
- de Graaff, W.E.; Oosterhuis, J.W.; de Jong, B.; Dam, A.; van Putten, W.L.; Castedo, S.M.; Sleijfer, D.T.; Schraffordt Koops, H. Ploidy of testicular carcinoma in situ. Lab. Investig. 1992, 66, 166–168. [Google Scholar]
- Shen, H.; Shih, J.; Hollern, D.P.; Wang, L.; Bowlby, R.; Tickoo, S.K.; Thorsson, V.; Mungall, A.J.; Newton, Y.; Hegde, A.M.; et al. Integrated molecular characterization of testicular germ cell tumors. Cell. Rep. 2018, 23, 3392–3406. [Google Scholar] [CrossRef]
- Looijenga, L.H.; Rosenberg, C.; van Gurp, R.J.; Geelen, E.; van Echten-Arends, J.; de Jong, B.; Mostert, M.; Wolter Oosterhuis, J. Comparative genomic hybridization of microdissected samples from different stages in the development of a seminoma and a non-seminoma. J. Pathol. 2000, 191, 187–192. [Google Scholar] [CrossRef]
- Cutcutache, I.; Suzuki, Y.; Tan, I.B.; Ramgopal, S.; Zhang, S.; Ramnarayanan, K.; Gan, A.; Lee, H.H.; Tay, S.T.; Ooi, A.; et al. Exome-wide sequencing shows low mutation rates and identifies novel mutated genes in seminomas. Eur. Urol. 2015, 68, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Litchfield, K.; Summersgill, B.; Yost, S.; Sultana, R.; Labreche, K.; Dudakia, D.; Renwick, A.; Seal, S.; Al-Saadi, R.; Broderick, P.; et al. Whole-exome sequencing reveals the mutational spectrum of testicular germ cell tumours. Nat. Commun. 2015, 6, 5973. [Google Scholar] [CrossRef] [Green Version]
- Alagaratnam, S.; Lind, G.E.; Kraggerud, S.M.; Lothe, R.A.; Skotheim, R.I. The testicular germ cell tumour transcriptome. Int. J. Androl. 2011, 34, e133–e151. [Google Scholar] [CrossRef]
- Okamoto, K. Epigenetics: A way to understand the origin and biology of testicular germ cell tumors. Int. J. Urol. 2012, 19, 504–511. [Google Scholar] [CrossRef]
- Ushida, H.; Kawakami, T.; Minami, K.; Chano, T.; Okabe, H.; Okada, Y.; Okamoto, K. Methylation profile of DNA repetitive elements in human testicular germ cell tumor. Mol. Carcinog. 2012, 51, 711–722. [Google Scholar] [CrossRef]
- Smiraglia, D.J.; Szymanska, J.; Kraggerud, S.M.; Lothe, R.A.; Peltomaki, P.; Plass, C. Distinct epigenetic phenotypes in seminomatous and nonseminomatous testicular germ cell tumors. Oncogene 2002, 21, 3909–3916. [Google Scholar] [CrossRef] [Green Version]
- Peltomaki, P. DNA methylation changes in human testicular cancer. Biochim. Biophys. Acta 1991, 1096, 187–196. [Google Scholar] [CrossRef]
- Sicinski, P.; Donaher, J.L.; Geng, Y.; Parker, S.B.; Gardner, H.; Park, M.Y.; Robker, R.L.; Richards, J.S.; McGinnis, L.K.; Biggers, J.D.; et al. Cyclin D2 is an FSH-responsive gene involved in gonadal cell proliferation and oncogenesis. Nature 1996, 384, 470–474. [Google Scholar] [CrossRef]
- Godmann, M.; Gashaw, I.; Eildermann, K.; Schweyer, S.; Bergmann, M.; Skotheim, R.I.; Behr, R. The pluripotency transcription factor Kruppel-like factor 4 is strongly expressed in intratubular germ cell neoplasia unclassified and seminoma. Mol. Hum. Reprod. 2009, 15, 479–488. [Google Scholar] [CrossRef] [Green Version]
- Sikora, K.; Evan, G.; Stewart, J.; Watson, J.V. Detection of the c-myc oncogene product in testicular cancer. Br. J. Cancer 1985, 52, 171–176. [Google Scholar] [CrossRef] [Green Version]
- Strohmeyer, T.; Reissmann, P.; Cordon-Cardo, C.; Hartmann, M.; Ackermann, R.; Slamon, D. Correlation between retinoblastoma gene expression and differentiation in human testicular tumors. Proc. Natl. Acad. Sci. USA 1991, 88, 6662–6666. [Google Scholar] [CrossRef] [Green Version]
- Boublikova, L.; Buchler, T.; Stary, J.; Abrahamova, J.; Trka, J. Molecular biology of testicular germ cell tumors: Unique features awaiting clinical application. Crit. Rev. Oncol. Hematol. 2014, 89, 366–385. [Google Scholar] [CrossRef]
- Goldberg, E.; Eddy, G.; Stewart, J.; Watson, J.V. LDHC: The ultimate testis-specific gene. J. Androl. 2010, 31, 86–94. [Google Scholar] [CrossRef]
- Skude, G.; von Eyben, F.E.; Kristiansen, P. Additional lactate dehydrogenase (LDH) isoenzymes in normal testis and spermatozoa of adult man. Mol. Gen. Genet. 1984, 198, 172–174. [Google Scholar] [CrossRef]
- Saeed, B.A.; Barband, R.S.; Alnasiri, U.S. Lactate dehydrogenase C4 (LDH-C4) is essential for the sperm count and motility: A case-control study. Baghdad J. Biochem. Appl. Biol. Sci. 2021, 2, 146–159. [Google Scholar] [CrossRef]
- Nettersheim, D.; Arndt, I.; Sharma, R.; Riesenberg, S.; Jostes, S.; Schneider, S.; Holzel, M.; Kristiansen, G.; Schorle, H. The cancer/testis-antigen PRAME supports the pluripotency network and represses somatic and germ cell differentiation programs in seminomas. Br. J. Cancer 2016, 115, 454–464. [Google Scholar] [CrossRef] [Green Version]
- Ricci, C.; Franceschini, T.; Giunchi, F.; Grillini, M.; Ambrosi, F.; Massari, F.; Mollica, V.; Colecchia, M.; Fiorentino, M. Immunohistochemical expression of preferentially expressed antigen in melanoma (PRAME) in the uninvolved background testis, germ cell neoplasia in situ, and germ cell tumors of the testis. Am. J. Clin. Pathol. 2022, 157, 644–648. [Google Scholar] [CrossRef]
- Orsatti, A.; Sirolli, M.; Ambrosi, F.; Franceschini, T.; Giunchi, F.; Franchini, E.; Grillini, M.; Massari, F.; Mollica, V.; Bianchi, F.M.; et al. SOX2 and PRAME in the “reprogramming” of seminoma cells. Pathol. Res. Pract. 2022, 237, 154044. [Google Scholar] [CrossRef]
- Cheung, H.H.; Yang, Y.; Lee, T.L.; Rennert, O.; Chan, W.Y. Hypermethylation of genes in testicular embryonal carcinomas. Br. J. Cancer 2016, 114, 230–236. [Google Scholar] [CrossRef] [Green Version]
- Juric, D.; Sale, S.; Hromas, R.A.; Yu, R.; Wang, Y.; Duran, G.E.; Tibshirani, R.; Einhorn, L.H.; Sikic, B.I. Gene expression profiling differentiates germ cell tumors from other cancers and defines subtype-specific signatures. Proc. Natl. Acad. Sci. USA 2005, 102, 17763–17768. [Google Scholar] [CrossRef] [Green Version]
- Sperger, J.M.; Chen, X.; Draper, J.S.; Antosiewicz, J.E.; Chon, C.H.; Jones, S.B.; Brooks, J.D.; Andrews, P.W.; Brown, P.O.; Thomson, J.A. Gene expression patterns in human embryonic stem cells and human pluripotent germ cell tumors. Proc. Natl. Acad. Sci. USA 2003, 100, 13350–13355. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, D.; Rapley, E.; Shipley, J. Testicular germ cell tumours: Predisposition genes and the male germ cell niche. Nat. Rev. Cancer 2011, 11, 278–288. [Google Scholar] [CrossRef]
- Rosenberg, C.; Van Gurp, R.J.; Geelen, E.; Oosterhuis, J.W.; Looijenga, L.H. Overrepresentation of the short arm of chromosome 12 is related to invasive growth of human testicular seminomas and nonseminomas. Oncogene 2000, 19, 5858–5862. [Google Scholar] [CrossRef] [Green Version]
- Ottesen, A.M.; Skakkebaek, N.E.; Lundsteen, C.; Leffers, H.; Larsen, J.; Rajpert-De Meyts, E. High-resolution comparative genomic hybridization detects extra chromosome arm 12p material in most cases of carcinoma in situ adjacent to overt germ cell tumors, but not before the invasive tumor development. Genes Chromosomes Cancer 2003, 38, 117–125. [Google Scholar] [CrossRef]
- Datta, M.W.; Macri, E.; Signoretti, S.; Renshaw, A.A.; Loda, M. Transition from in situ to invasive testicular germ cell neoplasia is associated with the loss of p21 and gain of mdm-2 expression. Mod. Pathol. 2001, 14, 437–442. [Google Scholar] [CrossRef] [Green Version]
- Gashaw, I.; Dushaj, O.; Behr, R.; Biermann, K.; Brehm, R.; Rubben, H.; Grobholz, R.; Schmid, K.W.; Bergmann, M.; Winterhager, E. Novel germ cell markers characterize testicular seminoma and fetal testis. Mol. Hum. Reprod. 2007, 13, 721–727. [Google Scholar] [CrossRef]
- Skotheim, R.I.; Lind, G.E.; Monni, O.; Nesland, J.M.; Abeler, V.M.; Fossa, S.D.; Duale, N.; Brunborg, G.; Kallioniemi, O.; Andrews, P.W.; et al. Differentiation of human embryonal carcinomas in vitro and in vivo reveals expression profiles relevant to normal development. Cancer Res. 2005, 65, 5588–5598. [Google Scholar] [CrossRef] [Green Version]
- Korkola, J.E.; Houldsworth, J.; Chadalavada, R.S.; Olshen, A.B.; Dobrzynski, D.; Reuter, V.E.; Bosl, G.J.; Chaganti, R.S. Down-regulation of stem cell genes, including those in a 200-kb gene cluster at 12p13.31, is associated with in vivo differentiation of human male germ cell tumors. Cancer Res. 2006, 66, 820–827. [Google Scholar] [CrossRef] [Green Version]
- Nettersheim, D.; Gillis, A.J.; Looijenga, L.H.; Schorle, H. TGF-beta1, EGF and FGF4 synergistically induce differentiation of the seminoma cell line TCam-2 into a cell type resembling mixed non-seminoma. Int. J. Androl. 2011, 34, e189–e203. [Google Scholar] [CrossRef]
- Benesova, M.; Trejbalova, K.; Kucerova, D.; Vernerova, Z.; Hron, T.; Szabo, A.; Amouroux, R.; Klezl, P.; Hajkova, P.; Hejnar, J. Overexpression of TET dioxygenases in seminomas associates with low levels of DNA methylation and hydroxymethylation. Mol. Carcinog. 2017, 56, 1837–1850. [Google Scholar] [CrossRef] [Green Version]
- Spiller, C.M.; Gillis, A.J.; Burnet, G.; Stoop, H.; Koopman, P.; Bowles, J.; Looijenga, L.H. Cripto: Expression, epigenetic regulation and potential diagnostic use in testicular germ cell tumors. Mol. Oncol. 2016, 10, 526–537. [Google Scholar] [CrossRef] [Green Version]
- Norgaard-Pedersen, B.; Albrechtsen, R.; Teilum, G. Serum alpha-foetoprotein as a marker for endodermal sinus tumour (yolk sac tumour) or a vitelline component of “teratocarcinoma”. Acta Pathol. Microbiol. Scand. A 1975, 83, 573–589. [Google Scholar] [CrossRef]
- Lempiainen, A.; Sankila, A.; Hotakainen, K.; Haglund, C.; Blomqvist, C.; Stenman, U.H. Expression of human chorionic gonadotropin in testicular germ cell tumors. Urol. Oncol. 2014, 32, 727–734. [Google Scholar] [CrossRef]
- Jacobsen, C.; Honecker, F. Cisplatin resistance in germ cell tumours: Models and mechanisms. Andrology 2015, 3, 111–121. [Google Scholar] [CrossRef]
- Christophersen, N.S.; Helin, K. Epigenetic control of embryonic stem cell fate. J. Exp. Med. 2010, 207, 2287–2295. [Google Scholar] [CrossRef] [Green Version]
- Nettersheim, D.; Heukamp, L.C.; Fronhoffs, F.; Grewe, M.J.; Haas, N.; Waha, A.; Honecker, F.; Waha, A.; Kristiansen, G.; Schorle, H. Analysis of TET expression/activity and 5mC oxidation during normal and malignant germ cell development. PLoS ONE 2013, 8, e82881. [Google Scholar] [CrossRef]
- Liu, J.; Shi, H.; Li, X.; Chen, G.; Larsson, C.; Lui, W.O. miR2233p regulates cell growth and apoptosis via FBXW7 suggesting an oncogenic role in human testicular germ cell tumors. Int. J. Oncol. 2017, 50, 356–364. [Google Scholar] [CrossRef] [Green Version]
- Kristensen, D.G.; Nielsen, J.E.; Jørgensen, A.; Skakkebæk, N.E.; Rajpert-De Meyts, E.; Almstrup, K. Evidence that active demethylation mechanisms maintain the genome of carcinoma in situ cells hypomethylated in the adult testis. Br. J. Cancer 2014, 110, 668–678. [Google Scholar] [CrossRef]
- Kristensen, D.G.; Skakkebaek, N.E.; Rajpert-DeMeyts, E.; Almstrup, K. Epigenetic features of testicular germ cell tumours in relation to epigenetic characteristics of foetal germ cells. Int. J. Dev. Biol. 2013, 57, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Aksoy, I.; Jauch, R.; Chen, J.; Dyla, M.; Divakar, U.; Bogu, G.K.; Teo, R.; Leng Ng, C.K.; Herath, W.; Lili, S.; et al. Oct4 switches partnering from Sox2 to Sox17 to reinterpret the enhancer code and specify endoderm. EMBO J. 2013, 32, 938–953. [Google Scholar] [CrossRef] [Green Version]
- Kushwaha, R.; Jagadish, N.; Kustagi, M.; Mendiratta, G.; Seandel, M.; Soni, R.; Korkola, J.E.; Thodima, V.; Califano, A.; Bosl, G.J.; et al. Mechanism and role of SOX2 repression in seminoma: Relevance to human germline specification. Stem Cell Rep. 2016, 6, 772–783. [Google Scholar] [CrossRef] [Green Version]
- Fritzsche, F.R.; Hasler, A.; Bode, P.K.; Adams, H.; Seifert, H.H.; Sulser, T.; Moch, H.; Barghorn, A.; Kristiansen, G. Expression of histone deacetylases 1, 2 and 3 in histological subtypes of testicular germ cell tumours. Histol. Histopathol. 2011, 26, 1555–1561. [Google Scholar]
- de Jong, J.; Stoop, H.; Gillis, A.J.; Hersmus, R.; van Gurp, R.J.; van de Geijn, G.J.; van Drunen, E.; Beverloo, H.B.; Schneider, D.T.; Sherlock, J.K.; et al. Further characterization of the first seminoma cell line TCam-2. Genes Chromosomes Cancer 2008, 47, 185–196. [Google Scholar] [CrossRef]
- van der Zwan, Y.G.; Rijlaarsdam, M.A.; Rossello, F.J.; Notini, A.J.; de Boer, S.; Watkins, D.N.; Gillis, A.J.; Dorssers, L.C.; White, S.J.; Looijenga, L.H. Seminoma and embryonal carcinoma footprints identified by analysis of integrated genome-wide epigenetic and expression profiles of germ cell cancer cell lines. PLoS ONE 2014, 9, e98330. [Google Scholar] [CrossRef] [Green Version]
- Nettersheim, D.; Jostes, S.; Sharma, R.; Schneider, S.; Hofmann, A.; Ferreira, H.J.; Hoffmann, P.; Kristiansen, G.; Esteller, M.B.; Schorle, H. BMP inhibition in seminomas initiates acquisition of pluripotency via NODAL signaling resulting in reprogramming to an embryonal carcinoma. PLoS Genet. 2015, 11, e1005415. [Google Scholar] [CrossRef] [Green Version]
- Lobo, J.; Guimaraes, R.; Miranda-Goncalves, V.; Monteiro-Reis, S.; Cantante, M.; Antunes, L.; Braga, I.; Mauricio, J.; Looijenga, L.H.; Jeronimo, C.; et al. Differential expression of DNA methyltransferases and demethylases among the various testicular germ cell tumor subtypes. Epigenomics 2020, 12, 1579–1592. [Google Scholar] [CrossRef]
- Villodre, E.S.; Felipe, K.B.; Oyama, M.Z.; Oliveira, F.H.; Lopez, P.; Solari, C.; Sevlever, G.; Guberman, A.; Lenz, G. Silencing of the transcription factors Oct4, Sox2, Klf4, c-Myc or Nanog has different effect on teratoma growth. Biochem. Biophys. Res. Commun. 2019, 517, 324–329. [Google Scholar] [CrossRef]
- Eckert, D.; Biermann, K.; Nettersheim, D.; Gillis, A.J.; Steger, K.; Jack, H.M.; Muller, A.M.; Looijenga, L.H.; Schorle, H. Expression of BLIMP1/PRMT5 and concurrent histone H2A/H4 arginine 3 dimethylation in fetal germ cells, CIS/IGCNU and germ cell tumors. BMC Dev. Biol. 2008, 8, 106. [Google Scholar] [CrossRef] [Green Version]
- Miranda-Goncalves, V.; Lobo, J.; Guimaraes-Teixeira, C.; Barros-Silva, D.; Guimaraes, R.; Cantante, M.; Braga, I.; Mauricio, J.; Oing, C.; Honecker, F.; et al. The component of the m(6)A writer complex VIRMA is implicated in aggressive tumor phenotype, DNA damage response and cisplatin resistance in germ cell tumors. J. Exp. Clin. Cancer Res. 2021, 40, 268. [Google Scholar] [CrossRef]
- Wermann, H.; Stoop, H.; Gillis, A.J.; Honecker, F.; van Gurp, R.J.; Ammerpohl, O.; Richter, J.; Oosterhuis, J.W.; Bokemeyer, C.; Looijenga, L.H. Global DNA methylation in fetal human germ cells and germ cell tumours: Association with differentiation and cisplatin resistance. J. Pathol. 2010, 221, 433–442. [Google Scholar] [CrossRef]
- Fazal, Z.; Singh, R.; Fang, F.; Bikorimana, E.; Baldwin, H.; Corbet, A.; Tomlin, M.; Yerby, C.; Adra, N.; Albany, C.; et al. Hypermethylation and global remodelling of DNA methylation is associated with acquired cisplatin resistance in testicular germ cell tumours. Epigenetics 2021, 16, 1071–1084. [Google Scholar] [CrossRef]
- Buljubasic, R.; Buljubasic, M.; Bojanac, A.K.; Ulamec, M.; Vlahovic, M.; Jezek, D.; Bulic-Jakus, F.; Sincic, N. Epigenetics and testicular germ cell tumors. Gene 2018, 661, 22–33. [Google Scholar] [CrossRef]
- De Jong, J.; Weeda, S.; Gillis, A.J.; Oosterhuis, J.W.; Looijenga, L.H. Differential methylation of the OCT3/4 upstream region in primary human testicular germ cell tumors. Oncol. Rep. 2007, 18, 127–132. [Google Scholar] [CrossRef]
- Mallik, S.; Qin, G.; Jia, P.; Zhao, Z. Molecular signatures identified by integrating gene expression and methylation in non-seminoma and seminoma of testicular germ cell tumours. Epigenetics 2021, 16, 162–176. [Google Scholar] [CrossRef]
- Brait, M.; Maldonado, L.; Begum, S.; Loyo, M.; Wehle, D.; Tavora, F.F.; Looijenga, L.H.; Kowalski, J.; Zhang, Z.; Rosenbaum, E.; et al. DNA methylation profiles delineate epigenetic heterogeneity in seminoma and non-seminoma. Br. J. Cancer 2012, 106, 414–423. [Google Scholar] [CrossRef] [Green Version]
- Chu, W.K.; Hung, L.M.; Hou, C.W.; Chen, J.K. MicroRNA 630 represses NANOG expression through transcriptional and post-transcriptional regulation in human embryonal carcinoma Cells. Int. J. Mol. Sci. 2021, 23, 46. [Google Scholar] [CrossRef]
- Agger, K.; Santoni-Rugiu, E.; Holmberg, C.; Karlstrom, O.; Helin, K. Conditional E2F1 activation in transgenic mice causes testicular atrophy and dysplasia mimicking human CIS. Oncogene 2005, 24, 780–789. [Google Scholar] [CrossRef] [Green Version]
- Lambrot, R.; Kimmins, S. Histone methylation is a critical regulator of the abnormal expression of POU5F1 and RASSF1A in testis cancer cell lines. Int. J. Androl. 2011, 34, 110–123. [Google Scholar] [CrossRef]
- Wang, J.; Lu, F.; Ren, Q.; Sun, H.; Xu, Z.; Lan, R.; Liu, Y.; Ward, D.; Quan, J.; Ye, T.; et al. Novel histone demethylase LSD1 inhibitors selectively target cancer cells with pluripotent stem cell properties. Cancer Res. 2011, 71, 7238–7249. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Yang, Y.; Li, W.; Chen, Q.; Li, J.; Pan, X.; Zhou, L.; Liu, C.; Chen, C.; He, J.; et al. Reciprocal regulation of Akt and Oct4 promotes the self-renewal and survival of embryonal carcinoma cells. Mol. Cell. 2012, 48, 627–640. [Google Scholar] [CrossRef] [Green Version]
- Hinz, S.; Magheli, A.; Weikert, S.; Schulze, W.; Krause, H.; Schrader, M.; Miller, K.; Kempkensteffen, C. Deregulation of EZH2 expression in human spermatogenic disorders and testicular germ cell tumors. World J. Urol. 2010, 28, 631–635. [Google Scholar] [CrossRef]
- Barrand, S.; Andersen, I.S.; Collas, P. Promoter-exon relationship of H3 lysine 9, 27, 36 and 79 methylation on pluripotency-associated genes. Biochem. Biophys. Res. Commun. 2010, 401, 611–617. [Google Scholar] [CrossRef]
- Singh, R.; Fazal, Z.; Bikorimana, E.; Boyd, R.I.; Yerby, C.; Tomlin, M.; Baldwin, H.; Shokry, D.; Corbet, A.K.; Shahid, K.; et al. Reciprocal epigenetic remodeling controls testicular cancer hypersensitivity to hypomethylating agents and chemotherapy. Mol. Oncol. 2022, 16, 683–698. [Google Scholar] [CrossRef]
- Gillis, A.J.; Stoop, H.; Biermann, K.; van Gurp, R.J.; Swartzman, E.; Cribbes, S.; Ferlinz, A.; Shannon, M.; Oosterhuis, J.W.; Looijenga, L.H. Expression and interdependencies of pluripotency factors LIN28, OCT3/4, NANOG and SOX2 in human testicular germ cells and tumours of the testis. Int. J. Androl. 2011, 34, e160–e174. [Google Scholar] [CrossRef]
- de Jong, J.; Looijenga, L.H. Stem cell marker OCT3/4 in tumor biology and germ cell tumor diagnostics: History and future. Crit. Rev. Oncog. 2006, 12, 171–203. [Google Scholar] [CrossRef]
- Gopalan, A.; Dhall, D.; Olgac, S.; Fine, S.W.; Korkola, J.E.; Houldsworth, J.; Chaganti, R.S.; Bosl, G.J.; Reuter, V.E.; Tickoo, S.K. Testicular mixed germ cell tumors: A morphological and immunohistochemical study using stem cell markers, OCT3/4, SOX2 and GDF3, with emphasis on morphologically difficult-to-classify areas. Mod. Pathol. 2009, 22, 1066–1074. [Google Scholar] [CrossRef] [Green Version]
- Wongtrakoongate, P. Epigenetic therapy of cancer stem and progenitor cells by targeting DNA methylation machineries. World J. Stem Cells 2015, 7, 137–148. [Google Scholar] [CrossRef]
- Boer, B.; Kopp, J.; Mallanna, S.; Desler, M.; Chakravarthy, H.; Wilder, P.J.; Bernadt, C.; Rizzino, A. Elevating the levels of Sox2 in embryonal carcinoma cells and embryonic stem cells inhibits the expression of Sox2:Oct-3/4 target genes. Nucleic Acids Res. 2007, 35, 1773–1786. [Google Scholar] [CrossRef]
- Hoang, N.; Zhang, X.; Zhang, C.; Vo, V.; Leng, F.; Saxena, L.; Yin, F.; Lu, F.; Zheng, G.; Bhowmik, P.; et al. New histone demethylase LSD1 inhibitor selectively targets teratocarcinoma and embryonic carcinoma cells. Bioorg. Med. Chem. 2018, 26, 1523–1537. [Google Scholar] [CrossRef]
- Li, W.; Fan, R.; Sun, M.; Jiang, T.; Gong, Y. Identification of Oct4-activating compounds that enhance reprogramming efficiency. Proc. Natl. Acad. Sci. USA 2012, 109, 20853–20858. [Google Scholar] [CrossRef] [Green Version]
- Eini, R.; Stoop, H.; Gillis, A.J.; Biermann, K.; Dorssers, L.C.; Looijenga, L.H. Role of SOX2 in the etiology of embryonal carcinoma, based on analysis of the NCCIT and NT2 cell lines. PLoS ONE 2014, 9, e83585. [Google Scholar] [CrossRef] [Green Version]
- Greber, B.; Lehrech, H.; Adjaye, I. Silencing of core transcription factors in human EC cells highlights the importance of autocrine FGF signaling for self-renewal. BMC Dev. Biol. 2007, 7, 46. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, H.; Theuser, M.; Wruck, W.; Adjaye, J. miR-27 negatively regulates pluripotency-associated genes in human embryonal carcinoma cells. PLoS ONE 2014, 9, e111637. [Google Scholar] [CrossRef]
- Rijlaarsdam, M.A.; Looijenga, L.H. An oncofetal and developmental perspective on testicular germ cell cancer. Semin. Cancer Biol. 2014, 29, 59–74. [Google Scholar] [CrossRef]
- Muller, M.R.; Skowron, M.A.; Albers, P.; Nettersheim, D. Molecular and epigenetic pathogenesis of germ cell tumors. Asian J. Urol. 2021, 8, 144–154. [Google Scholar] [CrossRef]
- Van Der Zwan, Y.G.; Stoop, H.; Rossello, F.; White, S.J.; Looijenga, L.H. Role of epigenetics in the etiology of germ cell cancer. Int. J. Dev. Biol. 2013, 57, 299–308. [Google Scholar] [CrossRef] [Green Version]
- Hopman, A.N.H.; Moshi, J.M.; Hoogduin, K.J.; Ummelen, M.; Henfling, M.E.R.; van Engeland, M.; Wouters, K.A.D.; Stoop, D.S.; Looijenga, L.H.J.; Ramaekers, F.C.S. SOX17 expression and its downregulation by promoter methylation in cervical adenocarcinoma in situ and adenocarcinoma. Histopathology 2020, 76, 383–393. [Google Scholar] [CrossRef] [Green Version]
- Tan, D.S.; Holzner, M.; Weng, M.; Srivastava, Y.; Jauch, R. Sox17 in cellular reprogramming and cancer. Sem. Cancer Biol. 2020, 67, 383–393. [Google Scholar] [CrossRef]
- Jostes, S.; Nettersheim, D.; Schneider, S.; Schorle, H. Cultivation of testicular germ cell cancer cell lines and establishment of gene-edited subclones using CRISPR/Cas9. Methods Mol. Biol. 2021, 2195, 85–97. [Google Scholar]
- West, J.A.; Viswanathan, S.R.; Yabuuchi, A.; Cunniff, K.; Takeuchi, A.; Park, I.H.; Sero, J.E.; Zhu, H.; Perez-Atayde, A.; Frazier, A.L.; et al. A role for Lin28 in primordial germ-cell development and germ-cell malignancy. Nature 2009, 460, 909–913. [Google Scholar] [CrossRef] [Green Version]
- Echigoya, K.; Koyama, M.; Negishi, L.; Takizawa, Y.; Mizukami, Y.; Shimabayashi, H.; Kuroda, A.; Kurumizaka, H. Nucleosome binding by the pioneer transcription factor OCT4. Sci. Rep. 2020, 10, 11832. [Google Scholar] [CrossRef]
- Murray, M.J.; Saini, H.K.; Siegler, C.A.; Hanning, J.E.; Barker, E.M.; van Dongen, S.; Ward, D.M.; Raby, K.L.; Groves, I.J.; Scarpini, C.G.; et al. LIN28 Expression in malignant germ cell tumors downregulates let-7 and increases oncogene levels. Cancer Res. 2013, 73, 4872–4884. [Google Scholar] [CrossRef] [Green Version]
- Nettersheim, D.; Biermann, K.; Gillis, A.J.; Steger, K.; Looijenga, L.H.; Schorle, H. NANOG promoter methylation and expression correlation during normal and malignant human germ cell development. Epigenetics 2011, 6, 114–122. [Google Scholar] [CrossRef] [Green Version]
- Rodda, D.J.; Chew, J.L.; Lim, L.H.; Loh, Y.H.; Wang, B.; Ng, H.H.; Robson, P. Transcriptional regulation of nanog by OCT4 and SOX2. J. Biol. Chem. 2005, 280, 24731–24737. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, T.; Tada, M.; Kubota, H.; Kimura, H.; Hatano, S.Y.; Suemori, H.; Nakatsuji, N.; Tada, T. Octamer and sox elements are required for transcriptional cis regulation of Nanog gene expression. Mol. Cell. Biol. 2005, 25, 2475–2485. [Google Scholar] [CrossRef] [Green Version]
- Hart, A.H.; Hartley, L.; Parker, K.; Ibrahim, M.; Looijenga, L.H.; Pauchnik, M.; Chow, C.W.; Robb, L. The pluripotency homeobox gene NANOG is expressed in human germ cell tumors. Cancer 2005, 104, 2092–2098. [Google Scholar] [CrossRef]
- Do, H.J.; Lee, W.Y.; Lim, H.Y.; Oh, J.H.; Kim, D.K.; Kim, J.H.; Kim, T.; Kim, J.H. Two potent transactivation domains in the C-terminal region of human NANOG mediate transcriptional activation in human embryonic carcinoma cells. J. Cell. Biochem. 2009, 106, 1079–1089. [Google Scholar] [CrossRef]
- Quagliana, J.M.; O’Bryan, R.M.; Baker, L.; Gottlieb, J.; Morrison, F.S.; Eyre, H.J.; Tucker, W.G.; Costanzi, J. Phase II study of 5-azacytidine in solid tumors. Cancer Treat. Rep. 1977, 61, 51–54. [Google Scholar]
- Roth, B.J.; Elson, P.; Sledge, G.W., Jr.; Einhorn, L.H.; Trump, D.L. 5-Azacytidine (NSC 102816) in refractory germ cell tumors. A phase II trial of the Eastern Cooperative Oncology Group. Investig. New Drugs 1993, 11, 201–202. [Google Scholar] [CrossRef]
- Minucci, S.; Horn, V.; Bhattacharyya, N.; Russanova, V.; Ogryzko, V.V.; Gabriele, L.; Howard, B.H.; Ozato, K. A histone deacetylase inhibitor potentiates retinoid receptor action in embryonal carcinoma cells. Proc. Natl. Acad. Sci. USA 1997, 94, 11295–11300. [Google Scholar] [CrossRef] [Green Version]
- Albany, C.; Fazal, Z.; Singh, R.; Bikorimana, E.; Adra, N.; Hanna, N.H.; Einhorn, L.H.; Perkins, S.M.; Sandusky, G.E.; Christensen, B.C.; et al. A phase 1 study of combined guadecitabine and cisplatin in platinum refractory germ cell cancer. Cancer Med. 2021, 10, 156–163. [Google Scholar] [CrossRef]
- Oing, C.; Verem, I.; Mansour, W.Y.; Bokemeyer, C.; Dyshlovoy, S.; Honecker, F. 5-Azacitidine exerts prolonged pro-apoptotic effects and overcomes cisplatin-resistance in non-seminomatous germ cell tumor cells. Int. J. Mol. Sci. 2018, 20, 21. [Google Scholar] [CrossRef] [Green Version]
- Clavel, M.; Monfardini, S.; Fossa, S.; Smyth, J.; Renard, J.; Kaye, S.B. 5-Aza-2’-deoxycytidine (NSC 127716) in non-seminomatous testicular cancer. Phase II from the EORTC Early Clinical Trials Cooperative Group and Genito-Urinary Group. Ann. Oncol. 1992, 3, 399–400. [Google Scholar] [CrossRef]
- Biswal, B.K.; Beyrouthy, M.J.; Hever-Jardine, M.P.; Armstrong, D.; Tomlinson, C.R.; Christensen, B.C.; Marsit, C.J.; Spinella, M.J. Acute hypersensitivity of pluripotent testicular cancer-derived embryonal carcinoma to low-dose 5-aza deoxycytidine is associated with global DNA damage-associated p53 activation, anti-pluripotency and DNA demethylation. PLoS ONE 2012, 7, e53003. [Google Scholar] [CrossRef]
- Beyrouthy, M.J.; Garner, K.M.; Hever, M.P.; Freemantle, S.J.; Eastman, A.; Dmitrovsky, E.; Spinella, M.J. High DNA methyltransferase 3B expression mediates 5-aza-deoxycytidine hypersensitivity in testicular germ cell tumors. Cancer Res. 2009, 69, 9360–9366. [Google Scholar] [CrossRef] [Green Version]
- Juttermann, R.; Li, E.; Jaenisch, R. Toxicity of 5-aza-2’-deoxycytidine to mammalian cells is mediated primarily by covalent trapping of DNA methyltransferase rather than DNA demethylation. Proc. Natl. Acad. Sci. USA 1994, 91, 11797–11801. [Google Scholar] [CrossRef] [Green Version]
- Lind, G.E.; Skotheim, R.I.; Fraga, M.F.; Abeler, V.M.; Esteller, M.; Lothe, R.A. Novel epigenetically deregulated genes in testicular cancer include homeobox genes and SCGB3A1 (HIN-1). J. Pathol. 2006, 210, 441–449. [Google Scholar] [CrossRef]
- Lee, M.G.; Wynder, C.; Schmidt, D.M.; McCafferty, D.G.; Shiekhattar, R. Histone H3 lysine 4 demethylation is a target of nonselective antidepressive medications. Chem. Biol. 2006, 13, 563–567. [Google Scholar] [CrossRef] [Green Version]
- Steinemann, G.; Dittmer, A.; Schmidt, J.; Josuttis, D.; Fähling, M.; Biersack, B.; Beindorff, N.; Jolante Koziolek, E.; Schobert, R.; Brenner, W.; et al. Antitumor and antiangiogenic activity of the novel chimeric inhibitor animacroxam in testicular germ cell cancer. Mol. Oncol. 2019, 13, 2679–2696. [Google Scholar] [CrossRef] [Green Version]
- Lobo, J.; Guimaraes-Teixeira, C.; Barros-Silva, D.; Miranda-Goncalves, V.; Camilo, V.; Guimaraes, R.; Cantante, M.; Braga, I.; Mauricio, J.; Oing, C.; et al. Efficacy of HDAC inhibitors belinostat and panobinostat against cisplatin-sensitive and cisplatin-resistant testicular germ cell tumors. Cancers 2020, 12, 2903. [Google Scholar] [CrossRef]
- Nettersheim, D.; Gillis, A.; Biermann, K.; Looijenga, L.H.; Schorle, H. The seminoma cell line TCam-2 is sensitive to HDAC inhibitor depsipeptide but tolerates various other chemotherapeutic drugs and loss of NANOG expression. Genes Chromosomes Cancer 2011, 50, 1033–1042. [Google Scholar] [CrossRef]
- Jostes, S.; Nettersheim, D.; Fellermeyer, M.; Schneider, S.; Hafezi, F.; Honecker, F.; Schumacher, V.; Geyer, M.; Kristiansen, G.; Schorle, H. The bromodomain inhibitor JQ1 triggers growth arrest and apoptosis in testicular germ cell tumours in vitro and in vivo. J. Cell. Mol. Med. 2017, 21, 1300–1314. [Google Scholar] [CrossRef]
- Nettersheim, D.; Jostes, S.; Fabry, M.; Honecker, F.; Schumacher, V.; Kirfel, J.; Kristiansen, G.; Schorle, H. A signaling cascade including ARID1A, GADD45B and DUSP1 induces apoptosis and affects the cell cycle of germ cell cancers after romidepsin treatment. Oncotarget 2016, 7, 74931–74946. [Google Scholar] [CrossRef]
- Muller, M.R.; Burmeister, A.; Skowron, M.A.; Stephan, A.; Bremmer, F.; Wakileh, G.A.; Petzsch, P.; Kohrer, K.; Albers, P.; Nettersheim, D. Therapeutical interference with the epigenetic landscape of germ cell tumors: A comparative drug study and new mechanistical insights. Clin. Epigenetics 2022, 14, 5. [Google Scholar] [CrossRef]
- Funke, K.; Duster, R.; Wilson, P.D.; Arevalo, L.; Geyer, M.; Schorle, H. Transcriptional CDK inhibitors as potential treatment option for testicular germ cell tumors. Cancers 2022, 14, 1690. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Friedman, N.B.; Moore, R.A. Tumors of the testis; a report on 922 cases. Mil. Surg. 1946, 99, 573–593. [Google Scholar] [CrossRef]
- Pugh, R.C.B.; Parkinson, C. The origin and classification of testicular germ cell tumours. Int. J. Androl. 1981, 4 (Suppl. S4), 15–24. [Google Scholar] [CrossRef]
- Skakkebaek, N.E.; Berthelsen, J.G.; Visfeldt, J. Clinical aspects of testicular carcinoma-in-situ. Int. J. Androl. 1981, 4 (Suppl. S4), 153–160. [Google Scholar] [CrossRef]
- Looijenga, L.H.; Van Agthoven, T.; Biermann, K. Development of malignant germ cells—The genvironmental hypothesis. Int. J. Dev. Biol. 2013, 57, 241–253. [Google Scholar] [CrossRef]
- Bredael, J.J.; Vugrin, D.; Whitmore, W.F., Jr. Autopsy findings in 154 patients with germ cell tumors of the testis. Cancer 1982, 50, 548–551. [Google Scholar] [CrossRef]
- Rabes, H.M. Proliferation of human testicular tumours. Int. J. Androl. 1987, 10, 127–137. [Google Scholar] [CrossRef]
- Shen, L.; Huang, X.; Xie, X.; Su, J.; Yuan, J.; Chen, X. High expression of SOX2 and OCT4 indicates radiation resistance and an independent negative prognosis in cervical squamous cell carcinoma. J. Histochem. Cytochem. 2014, 62, 499–509. [Google Scholar] [CrossRef] [Green Version]
- Oosterhuis, J.W.; Looijenga, L. Germ cell tumors from a developmental perspective: Cells of origin, pathogenesis, and molecular biology (merging patterns). In Pathology and Biology of Human Germ Cell Tumors; Nogales, F.F., Jimenez, R.E., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 23–129. ISBN 978-3-662-53773-2. [Google Scholar]
- von Eyben, F.E. Biochemical markers in advanced testicular tumors: Serum lactate dehydrogenase, urinary chorionic gonadotropin and total urinary estrogens. Cancer 1978, 41, 648–652. [Google Scholar] [CrossRef]
- von Eyben, F.E.; Jacobsen, G.K.; Pedersen, H.; Jacobsen, M.; Clausen, P.P.; Zibrandtsen, P.C.; Gullberg, B. Multivariate analysis of risk factors in patients with metastatic testicular germ cell tumors treated with vinblastine and bleomycin. Invasion Metastasis 1982, 2, 125–135. [Google Scholar]
- Bosl, G.J.; Geller, N.L.; Cirrincione, C.; Vogelzang, N.J.; Kennedy, B.J.; Whitmore, W.F., Jr.; Vugrin, D.; Scher, H.; Nisselbaum, J.; Golbey, R.B. Multivariate analysis of prognostic variables in patients with metastatic testicular cancer. Cancer Res. 1983, 43, 3403–3407. [Google Scholar]
- Seidel, C.; Daugaard, G.; Nestler, T.; Tryakin, A.; Fedyanin, M.; Fankhauser, C.D.; Hermanns, T.; Aparicio, J.; Heinzelbecker, J.; Paffenholz, P.; et al. The prognostic significance of lactate dehydrogenase levels in seminoma patients with advanced disease: An analysis by the Global Germ Cell Tumor Collaborative Group (G3). World J. Urol. 2021, 39, 3407–3414. [Google Scholar] [CrossRef]
- Vogelzang, N.J.; Lange, P.H.; Goldberg, E. Absence of sperm-specific lactate dehydrogenase-x in patients with testis cancer. Oncodev. Biol. Med. 1982, 3, 269–272. [Google Scholar]
- von Eyben, F.E.; de Graaff, W.E.; Marrink, J.; Blaabjerg, O.; Sleijfer, D.T.; Koops, H.S.; Oosterhuis, J.W.; Petersen, P.H.; van Echten-Arends, J.; de Jong, B. Serum lactate dehydrogenase isoenzyme 1 activity in patients with testicular germ cell tumors correlates with the total number of copies of the short arm of chromosome 12 in the tumor. Mol. Gen. Genet. 1992, 235, 140–146. [Google Scholar] [CrossRef]
- von Eyben, F.E.; Blaabjerg, O.; Madsen, E.L.; Petersen, P.H.; Smith-Sivertsen, C.; Gullberg, B. Serum lactate dehydrogenase isoenzyme 1 and tumour volume are indicators of response to treatment and predictors of prognosis in metastatic testicular germ cell tumours. Eur. J. Cancer 1992, 28, 410–415. [Google Scholar] [CrossRef]
- von Eyben, F.E.; Skude, G.; Fossa, S.D.; Klepp, O.; Bormer, O. Serum lactate dehydrogenase (S-LDH) and S-LDH isoenzymes in patients with testicular germ cell tumors. Mol. Gen. Genet. 1983, 189, 326–333. [Google Scholar] [CrossRef]
- von Eyben, F.E.; Blaabjerg, O.; Petersen, P.H.; Mommsen, S.; Madsen, E.L.; Kirpekar, F.; Li, S.S.; Kristiansen, K. Lactate dehydrogenase isoenzyme 1 in testicular germ cell tumors. Recent Results Cancer Res. 1991, 123, 85–92. [Google Scholar]
- Rorth, M.; Jacobsen, G.K.; von der Maase, H.; Madsen, E.L.; Nielsen, O.S.; Pedersen, M.; Schultz, H. Surveillance alone versus radiotherapy after orchiectomy for clinical stage I nonseminomatous testicular cancer. Danish Testicular Cancer Study Group. J. Clin. Oncol. 1991, 9, 1543–1548. [Google Scholar] [CrossRef]
- von der Maase, H.; Specht, L.; Jacobsen, G.K.; Jakobsen, A.; Madsen, E.L.; Pedersen, M.; Rorth, M.; Schultz, H. Surveillance following orchidectomy for stage I seminoma of the testis. Eur. J. Cancer 1993, 29, 1931–1934. [Google Scholar] [CrossRef]
- von Eyben, F.E.; Madsen, E.L.; Blaabjerg, O.; Petersen, P.H.; Jacobsen, G.K.; Specht, L.; Pedersen, B.N.; von der Maase, H. Serum lactate dehydrogenase isoenzyme 1 in patients with seminoma stage I followed with surveillance. Acta Oncol. 2002, 41, 77–83. [Google Scholar] [CrossRef] [Green Version]
- von Eyben, F.E.; Madsen, E.L.; Blaabjerg, O.; Petersen, P.H.; von der Maase, H.; Jacobsen, G.K.; Rorth, M. Serum lactate dehydrogenase isoenzyme 1 and relapse in patients with nonseminomatous testicular germ cell tumors clinical stage I. Acta Oncol. 2001, 40, 536–540. [Google Scholar] [CrossRef]
- Stephenson, A.; Eggener, S.E.; Bass, E.B.; Chelnick, D.M.; Daneshmand, S.; Feldman, D.; Gilligan, T.; Karam, J.A.; Leibovich, B.; Liauw, S.L.; et al. Diagnosis and treatment of early stage testicular cancer: AUA guideline. J. Urol. 2019, 202, 272–281. [Google Scholar] [CrossRef] [Green Version]
- Adra, N.; Einhorn, L.H. Testicular cancer update. Clin. Adv. Hematol. Oncol. 2017, 15, 386–396. [Google Scholar]
- Beyer, J.; Collette, L.; Sauve, N.; Daugaard, G.; Feldman, D.R.; Tandstad, T.; Tryakin, A.; Stahl, O.; Gonzalez-Billalabeitia, E.; De Giorgi, U.; et al. Survival and new prognosticators in metastatic seminoma: Results from the IGCCCG-Update Consortium. J. Clin. Oncol. 2021, 39, 1553–1562. [Google Scholar] [CrossRef]
- Gillessen, S.; Sauve, N.; Collette, L.; Daugaard, G.; de Wit, R.; Albany, C.; Tryakin, A.; Fizazi, K.; Stahl, O.; Gietema, J.A.; et al. Predicting outcomes in men with metastatic nonseminomatous germ cell tumors (NSGCT): Results from the IGCCCG Update Consortium. J. Clin. Oncol. 2021, 39, 1563–1574. [Google Scholar] [CrossRef]
- Fizazi, K.; Delva, R.; Caty, A.; Chevreau, C.; Kerbrat, P.; Rolland, F.; Priou, F.; Geoffrois, L.; Rixe, O.; Beuzeboc, P.; et al. A risk-adapted study of cisplatin and etoposide, with or without ifosfamide, in patients with metastatic seminoma: Results of the GETUG S99 multicenter prospective study. Eur. Urol. 2014, 65, 381–386. [Google Scholar] [CrossRef]
- Fossa, S.D.; Dahl, A.A.; Thorsen, L.; Hellesnes, R.; Kiserud, C.E.; Tandstad, T.; Brydoy, M.; Haugnes, H.S.; Myklebust, T.A. Mortality and second cancer incidence after treatment for testicular cancer: Psychosocial health and lifestyle are modifiable prognostic factors. J. Clin. Oncol. 2022, 40, 12. [Google Scholar] [CrossRef]
- Ishida, H.; Kasajima, A.; Kamei, T.; Miura, T.; Oka, N.; Yazdani, S.; Ozawa, Y.; Fujishima, F.; Sakurada, A.; Nakamura, Y.; et al. SOX2 and Rb1 in esophageal small-cell carcinoma: Their possible involvement in pathogenesis. Mod. Pathol. 2017, 30, 660–671. [Google Scholar] [CrossRef] [Green Version]
- Rudin, C.M.; Durinck, S.; Stawiski, E.W.; Poirier, J.T.; Modrusan, Z.; Shames, D.S.; Bergbower, E.A.; Guan, Y.; Shin, J.; Guillory, J.; et al. Comprehensive genomic analysis identifies SOX2 as a frequently amplified gene in small-cell lung cancer. Nat. Genet. 2012, 44, 1111–1116. [Google Scholar] [CrossRef]
- Kim, J.; Woo, A.J.; Chu, J.; Snow, J.W.; Fujiwara, Y.; Kim, C.G.; Cantor, A.B.; Orkin, S.H. A Myc network accounts for similarities between embryonic stem and cancer cell transcription programs. Cell 2010, 143, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Ben-Porath, I.; Thomson, M.W.; Carey, V.J.; Ge, R.; Bell, G.W.; Regev, A.; Weinberg, R.A. An embryonic stem cell-like gene expression signature in poorly differentiated aggressive human tumors. Nat. Genet. 2008, 40, 499–507. [Google Scholar] [CrossRef]
- Mamun, M.A.; Mannoor, K.; Cao, J.; Qadri, F.; Song, X. SOX2 in cancer stemness: Tumor malignancy and therapeutic potentials. J. Mol. Cell. Biol. 2020, 12, 85–98. [Google Scholar] [CrossRef] [Green Version]
- Hepburn, A.C.; Steele, R.E.; Veeratterapillay, R.; Wilson, L.; Kounatidou, E.E.; Barnard, A.; Berry, P.; Cassidy, J.R.; Moad, M.; El-Sherif, A.; et al. The induction of core pluripotency master regulators in cancers defines poor clinical outcomes and treatment resistance. Oncogene 2019, 38, 4412–4424. [Google Scholar] [CrossRef]
- Hatina, J.; Kripnerova, M.; Houdek, Z.; Pesta, M.; Tichanek, F. Pluripotency stemness and cancer: More questions than answers. Adv. Exp. Med. Biol. 2022, 1376, 77–100. [Google Scholar]
- Feinberg, A.P. The key role of epigenetics in human disease prevention and mitigation. N. Engl. J. Med. 2018, 378, 1323–1334. [Google Scholar] [CrossRef]
- Grubelnik, G.; Bostjancic, E.; Groselj, A.; Zidar, N. Expression of NANOG and its regulation in oral squamous cell carcinoma. Biomed. Res. Int. 2020, 245, 456–464. [Google Scholar] [CrossRef]
- Mansouri, S.; Nejad, R.; Karabork, M.; Ekinci, C.; Solaroglu, I.; Aldape, K.D.; Zadeh, G. Sox2: Regulation of expression and contribution to brain tumors. CNS Oncol. 2016, 5, 159–173. [Google Scholar] [CrossRef]
- Najafzadeh, B.; Asadzadeh, Z.; Motafakker Azad, R.; Mokhtarzadeh, A.; Baghbanzadeh, A.; Alemohammad, H.; Abdoli Shadbad, M.; Vasefifar, P.; Najafi, S.; Baradaran, B. The oncogenic potential of NANOG: An important cancer induction mediator. J. Cell. Physiol. 2021, 236, 2443–2458. [Google Scholar] [CrossRef]
- Egevad, L.; Delahunt, B.; Srigley, J.R.; Samaratunga, H. International Society of Urological Pathology (ISUP) grading of prostate cancer—An ISUP consensus on contemporary grading. APMIS 2016, 124, 433–435. [Google Scholar] [CrossRef]
- Nepali, K.; Liou, J.P. Recent developments in epigenetic cancer therapeutics: Clinical advancement and emerging trends. J. Biomed. Sci. 2021, 28, 27. [Google Scholar] [CrossRef]
- Pojani, E.; Barlocco, D. Romidepsin (FK228), a histone deacetylase inhibitor and its analogues in cancer chemotherapy. Curr. Med. Chem. 2021, 28, 1290–1303. [Google Scholar] [CrossRef]
- De Souza, C.; Chatterji, B.P. HDAC inhibitors as novel anti-cancer therapeutics. Recent Pat. Anticancer Drug Discov. 2015, 10, 145–162. [Google Scholar] [CrossRef]
- Patnaik, M.M.; Lasho, T. Myelodysplastic syndrome/myeloproliferative neoplasm overlap syndromes: A focused review. Hematol. Am. Soc. Hematol. Educ. Program 2020, 2020, 460–464. [Google Scholar] [CrossRef]
- Laubach, J.P.; Moreau, P.; San-Miguel, J.F.; Richarson, P.G. Panobinostat for the treatment of multiple myeloma. Clin. Cancer Res. 2015, 21, 4767–4773. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.Z.; Kwitkowski, V.E.; Del Valle, P.L.; Ricci, M.S.; Saber, H.; Habtemariam, B.A.; Bullock, J.; Bloomquist, E.; Li Shen, Y.; Chen, X.H.; et al. FDA approval: Belinostat for the treatment of patients with relapsed or refractory peripheral T-cell lymphoma. Clin. Cancer Res. 2015, 21, 2666–2670. [Google Scholar] [CrossRef] [Green Version]
- Eckschlager, T.; Plch, J.; Stiborova, M.; Hrabeta, J. Histone deacetylase inhibitors as anticancer drugs. Int. J. Mol. Sci. 2017, 18, 1414. [Google Scholar] [CrossRef] [Green Version]
- Fenaux, P.; Mufti, G.J.; Hellstrom-Lindberg, E.; Santini, V.; Finelli, C.; Giagounidis, A.; Schoch, R.; Gattermann, N.; Sanz, G.; List, A.; et al. Efficacy of azacitidine compared with that of conventional care regimens in the treatment of higher-risk myelodysplastic syndromes: A randomised, open-label, phase III study. Lancet Oncol. 2009, 10, 223–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhillon, S. Trilaciclib: First approval. Drugs 2021, 81, 867–874. [Google Scholar] [CrossRef]
- von Eyben, F.E. Epidermal growth factor receptor inhibition and non-small cell lung cancer. Crit. Rev. Clin. Lab. Sci. 2006, 43, 291–323. [Google Scholar] [CrossRef]
- Qi, Y.; Xia, X.; Shao, L.; Guo, L.; Dong, Y.; Tian, J.; Xu, L.; Niu, R.; Wei, S. An updated network meta-analysis of EGFR-TKIs and combination therapy in the first-line treatment of advanced EGFR mutation positive non-small cell lung cancer. Front. Oncol. 2022, 12, 616546. [Google Scholar] [CrossRef]
- Festuccia, N.; Osorno, R.; Halbritter, F.; Karwacki-Neisius, V.; Navarro, P.; Colby, D.; Wong, F.; Yates, A.; Tomlinson, S.R.; Chambers, I. Esrrb is a direct Nanog target gene that can substitute for Nanog function in pluripotent cells. Cell. Stem Cell 2012, 11, 477–490. [Google Scholar] [CrossRef] [Green Version]
- Rony, I.K.; Baten, A.; Bloomfield, J.A.; Islam, M.E.; Billah, M.M.; Islam, K.D. Inducing pluripotency in vitro: Recent advances and highlights in induced pluripotent stem cells generation and pluripotency reprogramming. Cell. Prolif. 2015, 48, 140–156. [Google Scholar] [CrossRef]
- Wang, C.; Lin, Y.; Zhu, H.; Zhou, Y.; Mao, F.; Huang, X.; Sun, Q.; Li, C. Efficacy and safety profile of histone deacetylase inhibitors for metastatic breast cancer: A meta-analysis. Front. Oncol. 2022, 12, 901152. [Google Scholar] [CrossRef]
- DiNardo, C.D.; Jonas, B.A.; Pullarkat, V.; Thirman, M.J.; Garcia, J.S.; Wei, A.H.; Konopleva, M.; Dohner, H.; Letai, A.; Fenaux, P.; et al. Azacitidine and venetoclax in previously untreated acute myeloid leukemia. N. Engl. J. Med. 2020, 383, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Blagitko-Dorfs, N.; Schlosser, P.; Greve, G.; Pfeifer, D.; Meier, R.; Baude, A.; Brocks, D.; Plass, C.; Lubbert, M. Combination treatment of acute myeloid leukemia cells with DNMT and HDAC inhibitors: Predominant synergistic gene downregulation associated with gene body demethylation. Leukemia 2019, 33, 945–956. [Google Scholar] [CrossRef]
- Lobo, J.; van Zogchel, L.M.J.; Nuru, M.G.; Gillis, A.J.M.; van der Schoot, C.E.; Tytgat, G.A.M.; Looijenga, L.H.J. Combining hypermethylated RASSF1A detection using ddPCR with miR-371a-3p testing: An improved panel of liquid biopsy biomarkers for testicular germ cell tumor patients. Cancers 2021, 13, 5228. [Google Scholar] [CrossRef]
- Markulin, D.; Vojta, A.; Samarzija, I.; Gamulin, M.; Beceheli, I.; Jukic, I.; Maglov, C.; Zoldos, V.; Fucic, A. Association between RASSF1A promoter methylation and testicular germ cell tumor: A meta-analysis and a cohort study. Cancer Genomics Proteomics 2017, 14, 363–372. [Google Scholar]
- Bartkova, J.; Lukas, C.; Sorensen, C.S.; Rajpert-De Meyts, E.; Skakkebaek, N.E.; Lukas, J.; Bartek, J. Deregulation of the RB pathway in human testicular germ cell tumours. J. Pathol. 2003, 200, 149–156. [Google Scholar] [CrossRef]
- Warburg, O. On the origin of cancer cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef]
- Hoff, A.M.; Kraggerud, S.M.; Alagaratnam, S.; Berg, K.C.G.; Johannessen, B.; Holand, M.; Nilsen, G.; Lingjaerde, O.C.; Andrews, P.W.; Lothe, R.A.; et al. Frequent copy number gains of SLC2A3 and ETV1 in testicular embryonal carcinomas. Endocr. Relat. Cancer 2020, 27, 457–468. [Google Scholar] [CrossRef]
Figure 1.
TGCT subtypes in the tumorigenesis of TGCT. (a). The epigenetic landscape for germ cells. The germ cells progress from toti-differentiated cells to unipotent cells, and a gene panel of OCT4, SOX2, LIN28, and NANOG (OSLN) can reprogram germ cells for induced pluri- to totipotent stem cells (iPSC). The red boxes show gene panels for iPSC. The progression of the TGCT subtypes implies a loss of chromosomes from tetraploid malignant germ cells over hypertriploid seminoma to hypotriploid embryonic carcinoma. (b). Microinvasive germ cell tumor (MGCT). MGCT is the first invasive step in the tumorigenesis of TGCT. MGCT is an intermediary precursor of TGCT that has malignant germ cells inside and outside seminiferous tubules in the testis. The figure shows MGCT stained for OCT4, the protein of POU5F1. (c). Embryonal carcinoma (EC). EC has a positive homogeneous immunohistochemical staining for three transcription factors for induced pluripotent stem cells (OCT4, SOX2, and NANOG) and a homogeneous negative staining for the protein of the tumor suppressor gene, RB1. EC stained immunohistochemically positive for OCT4 (1c1), SOX2 (1c2), LIN28 (1c3), and immunohistochemically negative for RB1 (1c4).
Figure 1.
TGCT subtypes in the tumorigenesis of TGCT. (a). The epigenetic landscape for germ cells. The germ cells progress from toti-differentiated cells to unipotent cells, and a gene panel of OCT4, SOX2, LIN28, and NANOG (OSLN) can reprogram germ cells for induced pluri- to totipotent stem cells (iPSC). The red boxes show gene panels for iPSC. The progression of the TGCT subtypes implies a loss of chromosomes from tetraploid malignant germ cells over hypertriploid seminoma to hypotriploid embryonic carcinoma. (b). Microinvasive germ cell tumor (MGCT). MGCT is the first invasive step in the tumorigenesis of TGCT. MGCT is an intermediary precursor of TGCT that has malignant germ cells inside and outside seminiferous tubules in the testis. The figure shows MGCT stained for OCT4, the protein of POU5F1. (c). Embryonal carcinoma (EC). EC has a positive homogeneous immunohistochemical staining for three transcription factors for induced pluripotent stem cells (OCT4, SOX2, and NANOG) and a homogeneous negative staining for the protein of the tumor suppressor gene, RB1. EC stained immunohistochemically positive for OCT4 (1c1), SOX2 (1c2), LIN28 (1c3), and immunohistochemically negative for RB1 (1c4).



Figure 2.
Development of normal and malignant testicular germ cells. (a) Shows the development of normal testicular germ cells from fetal life to spermatozoa in adulthood, and the development of malignant testicular germ cells from tetraploid malignant germ cells to the testicular germ cell tumors type II subtypes. Seminomas are diagnosed at a higher age than non-seminomatous testicular germ cell tumors as reflection of a lower proliferation rate in seminoma. (b). Chromosome 12. The short arm of chromosome 12 (12p) has two candidate regions that are especially important for the tumorigenesis of testicular germ cell tumors type II, shown in red. The candidate regions have gene loci for three driver genes: CCND2, NANOG and LDHB. The candidate regions are shown in red. The figure also shows other genes that literature and OMIM relate to TGCT. (c). IPSC gene panel in TGCT subtypes. The combined OSKM/OSLN panels for seminoma, embryonal carcinoma, and teratoma have dramatic differences with up- and down-regulations of the genes. Down-regulated genes are indicated with red and up-regulated genes are indicated with green.
Figure 2.
Development of normal and malignant testicular germ cells. (a) Shows the development of normal testicular germ cells from fetal life to spermatozoa in adulthood, and the development of malignant testicular germ cells from tetraploid malignant germ cells to the testicular germ cell tumors type II subtypes. Seminomas are diagnosed at a higher age than non-seminomatous testicular germ cell tumors as reflection of a lower proliferation rate in seminoma. (b). Chromosome 12. The short arm of chromosome 12 (12p) has two candidate regions that are especially important for the tumorigenesis of testicular germ cell tumors type II, shown in red. The candidate regions have gene loci for three driver genes: CCND2, NANOG and LDHB. The candidate regions are shown in red. The figure also shows other genes that literature and OMIM relate to TGCT. (c). IPSC gene panel in TGCT subtypes. The combined OSKM/OSLN panels for seminoma, embryonal carcinoma, and teratoma have dramatic differences with up- and down-regulations of the genes. Down-regulated genes are indicated with red and up-regulated genes are indicated with green.
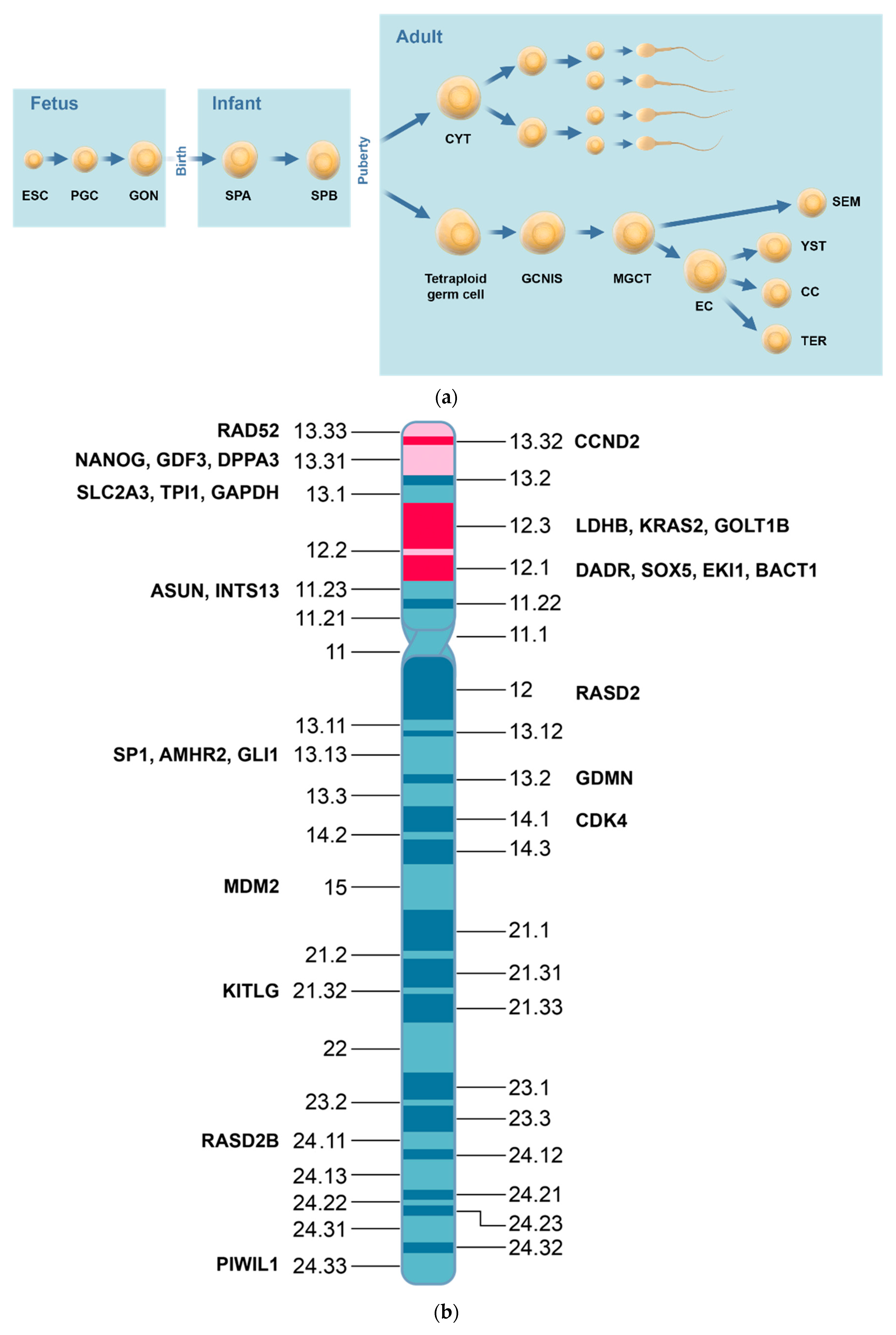
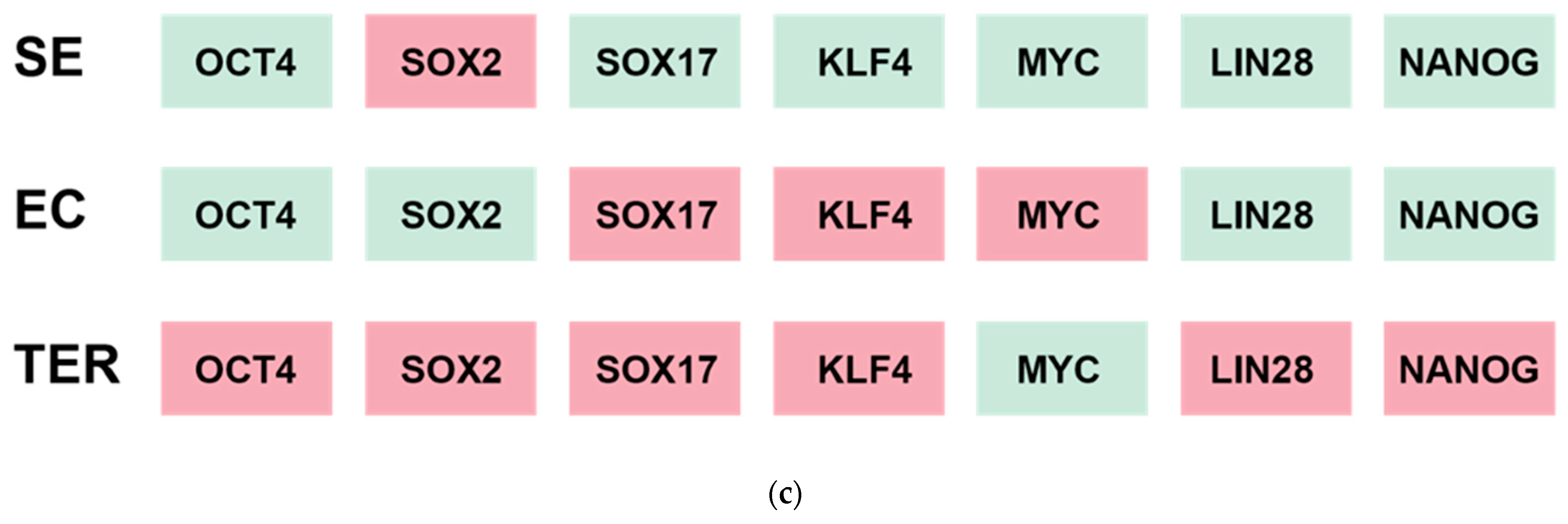
Figure 3.
Epigenetic regulation of gene expression. (a). Histone modifications and gene expression. Histone lysine methylations and acetylations can lead to repressed, poised or activated gene expressions. A combination of histone marks that act both to repress and activate a gene leads to a poised regulation of expression of the gene. (b). Sites for histone marks and gene regulation. Methylated histone 3 lysines differ in action sites on enhancers, promoters, and gene bodies of genes. Red indicates down-regulated gene expression and green indicates upregulated expression of the gene.
Figure 3.
Epigenetic regulation of gene expression. (a). Histone modifications and gene expression. Histone lysine methylations and acetylations can lead to repressed, poised or activated gene expressions. A combination of histone marks that act both to repress and activate a gene leads to a poised regulation of expression of the gene. (b). Sites for histone marks and gene regulation. Methylated histone 3 lysines differ in action sites on enhancers, promoters, and gene bodies of genes. Red indicates down-regulated gene expression and green indicates upregulated expression of the gene.
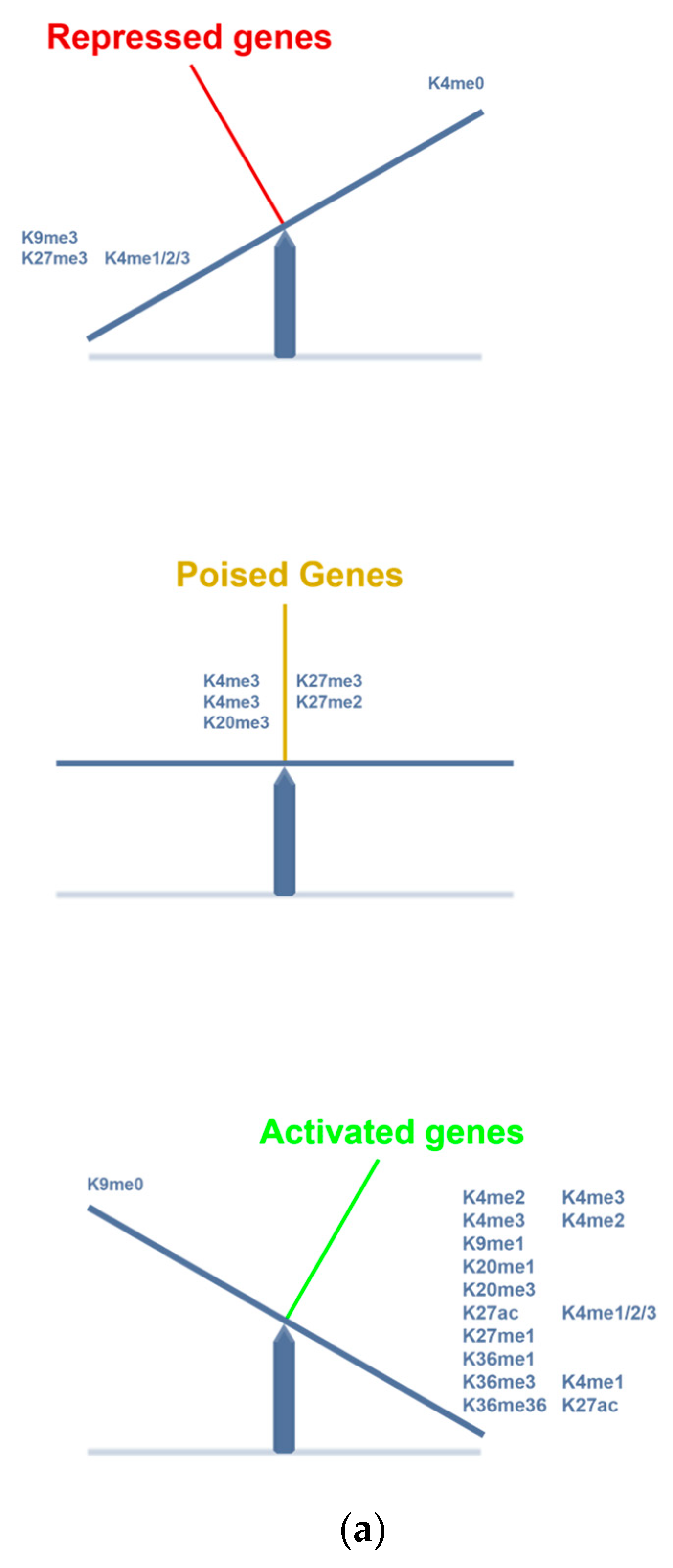

Figure 4.
A new trial of epidrugs in TGCT. Candidate patients are patients with high-risk NS metastatic TGCT with insufficient decay of serum tumor markers after the first course of BEP chemotherapy. The proposed trial investigates whether combination of epidrugs and BEP chemotherapy might give response equal to high-dose salvage chemotherapy. The new elements in the trial are shown in grey boxes. If a patient fails to present with sufficient decay of two serum tumor markers, the patient will be switched to a routine high-dose cisplatin salvage chemotherapy.
Figure 4.
A new trial of epidrugs in TGCT. Candidate patients are patients with high-risk NS metastatic TGCT with insufficient decay of serum tumor markers after the first course of BEP chemotherapy. The proposed trial investigates whether combination of epidrugs and BEP chemotherapy might give response equal to high-dose salvage chemotherapy. The new elements in the trial are shown in grey boxes. If a patient fails to present with sufficient decay of two serum tumor markers, the patient will be switched to a routine high-dose cisplatin salvage chemotherapy.
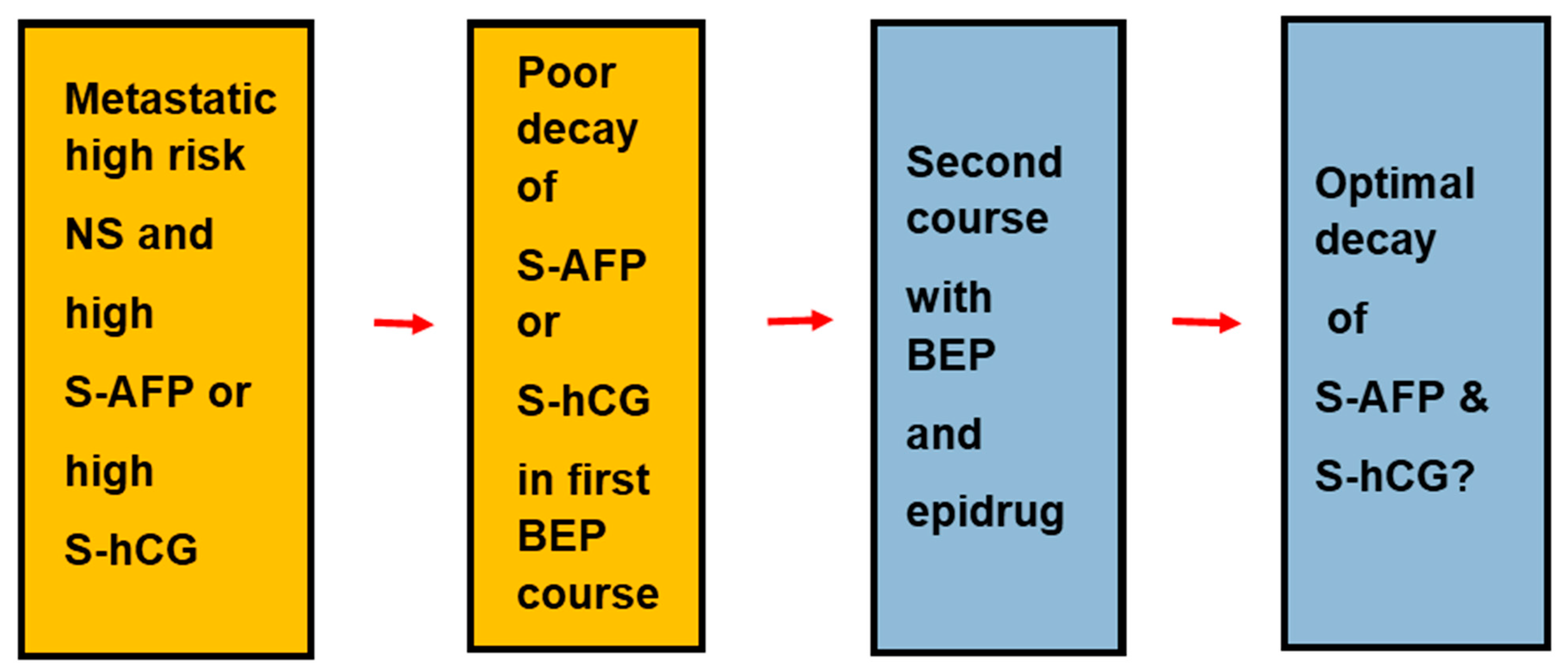

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Histologic types of TGCT and epigenetic regulation.
| Expressed Transcription Factors | Histologic Types | NGC | GCNIS | SE | EC | TER | |
|---|---|---|---|---|---|---|---|
| Epigenetic mediators | High | PRAME LDHC | PRAME, KIT, SOX17, NANOG | PRAME, KIT, SOX17, KLF4 POU5F1, NANOG, MYC | NANOG, POU5F1, DNMT3B | RB1, MYC | |
| Low | RB1 | RB1 | RB1, MYC | POU5F1, SO)X2, LIN28, NANOG | |||
| 5mC | +++ | Neg | Neg/++ | +++ | |||
| 5mhC | Neg | Neg | +++ | ||||
| Epigenetic modifiers | H2A.Z | +/+++ | +++ | Neg/+ | Neg/+ | ||
| H3K4 | Me1 +++ Me2/me3 ++ | Me1/me2/me3 +++ | Me1 ++/+++ | Me1 neg/+ Me2/Me3++/+++ | |||
| H3K9me1/me2 | Neg/+ | ++ | ++ | ||||
| H3K9ac | +++ | Neg/+ | Neg/+ | ||||
| H3K27me2 | |||||||
| H3K27me3 | +++ | Ac/+ | +++ | neg | |||
| Epigenetic modulators | DNMT | Low | High | ||||
| TET | Low | High | low | ||||
| HMT | EZD2 | +++ | Neg/+ | neg | |||
| HDMT | UTX | Neg | Neg | Neg | Neg | ||
| JMJDD3 | Neg/+ | Neg | Neg/+ | Neg/+ | |||
| HAT | High | Low | |||||
| HDAC | Low | High |
The signs +/++/+++ indicats the degree of positive immunohistochemical stainings.
Table 2.
Epidrugs in TGCT and other cancers.
| Pharmacologic Group | Drugs | TGCT | Ref | Other Malignancies | |
|---|---|---|---|---|---|
| Studies | Indications | ||||
| Cell lines | Patients | ||||
| DNMTi | 5-aza | X | X | Myelodysplastic syndromes | |
| 5-aza-cD | X | X | |||
| Guadecitabine | X | ||||
| KDMi | Chaertocin | [141] | |||
| JIB-04 | [141] | ||||
| EZH2i | Tazemetostat | Follicular NH lymphoma | |||
| Bromodomaini | JQ11 | X | [143] | ||
| LSD1i | CBB3001 | X | |||
| HDACi | belinostat | X | [144] | Cutaneous T cell lymphoma | |
| chicamide | X | ||||
| depsipeptide | X | [144] | |||
| panobinostat | X | [144] | Cutaneous T cell lymphoma | ||
| Romidepsin | X | Cutaneous T cell lymphoma | |||
| Quisinostat | X | [141] | |||
| trichostatin | |||||
| vorinostat | X | ||||
| CDKi | dinaciclib | [140] | |||
| Flavo-piridol | [140] | ||||
| NVP-2 | [140] | ||||
| SY0351 | [140] | ||||
| thal-sns-032 | [140] | ||||
| THZ1 | [140] | ||||
| THZ531 | [140] | ||||
| YKL-5-214 | [140] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
von Eyben, F.E.; Kristiansen, K.; Kapp, D.S.; Hu, R.; Preda, O.; Nogales, F.F. Epigenetic Regulation of Driver Genes in Testicular Tumorigenesis. Int. J. Mol. Sci. 2023, 24, 4148. https://doi.org/10.3390/ijms24044148
AMA Style
von Eyben FE, Kristiansen K, Kapp DS, Hu R, Preda O, Nogales FF. Epigenetic Regulation of Driver Genes in Testicular Tumorigenesis. International Journal of Molecular Sciences. 2023; 24(4):4148. https://doi.org/10.3390/ijms24044148
Chicago/Turabian Stylevon Eyben, Finn E., Karsten Kristiansen, Daniel S. Kapp, Rong Hu, Ovidiu Preda, and Francisco F. Nogales. 2023. "Epigenetic Regulation of Driver Genes in Testicular Tumorigenesis" International Journal of Molecular Sciences 24, no. 4: 4148. https://doi.org/10.3390/ijms24044148
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.