
Update on the Role and Regulatory Mechanism of Extracellular Matrix in the Pathogenesis of Uterine Fibroids

Department of Obstetrics and Gynecology, University of Chicago, Chicago, IL 60637, USA
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(6), 5778; https://doi.org/10.3390/ijms24065778
Submission received: 10 February 2023
/
Revised: 22 February 2023
/
Accepted: 8 March 2023
/
Published: 17 March 2023
(This article belongs to the Special Issue Latest Review Papers in Molecular Pathology, Diagnostics, and Therapeutics 2023)
Abstract
:Uterine fibroids (UFs), also known as leiomyomas, are benign tumors of the myometrium affecting over 70% of women worldwide, particularly women of color. Although benign, UFs are associated with significant morbidity; they are the primary indication for hysterectomy and a major source of gynecologic and reproductive dysfunction, ranging from menorrhagia and pelvic pain to infertility, recurrent miscarriage, and preterm labor. So far, the molecular mechanisms underlying the pathogenesis of UFs are still quite limited. A knowledge gap needs to be filled to help develop novel strategies that will ultimately facilitate the development of therapies and improve UF patient outcomes. Excessive ECM accumulation and aberrant remodeling are crucial for fibrotic diseases and excessive ECM deposition is the central characteristics of UFs. This review summarizes the recent progress of ascertaining the biological functions and regulatory mechanisms in UFs, from the perspective of factors regulating ECM production, ECM-mediated signaling, and pharmacological drugs targeting ECM accumulation. In addition, we provide the current state of knowledge by discussing the molecular mechanisms underlying the regulation and emerging role of the extracellular matrix in the pathogenesis of UFs and in applications. Comprehensive and deeper insights into ECM-mediated alterations and interactions in cellular events will help develop novel strategies to treat patients with this common tumor.
1. Introduction
Uterine fibroids (UFs), also known as leiomyomas or myomas, are benign tumors of the myometrium affecting over 70% of women worldwide, particularly women of color. Although benign, UFs are associated with significant morbidity; they are the primary indication for hysterectomy and a major source of gynecologic and reproductive dysfunction, ranging from menorrhagia and pelvic pain to infertility, recurrent miscarriage, and preterm labor [1,2,3]. Each year, approximately 300,000 myomectomies and 200,000 hysterectomies are performed in the United States to remove either leiomyoma tumors or the whole uterus [4,5]. Accordingly, the annual USA healthcare costs associated with UFs have been estimated at ~34 billion USD. Therefore, UFs represent significant societal health and financial burdens.
Several risk factors have been shown to impact UF pathogenesis and are associated with a higher probability of UF formation and development. These factors include race, age, parity, family history, food additives, obesity, vitamin D deficiency, and endocrine-disrupting chemical exposure [1,6]. These risk factors affect several key pathways, including inflammation [7,8,9], DNA damage repair pathway, β-catenin pathway, and genetic instability, among others, leading to the pathogenesis of UFs [10,11,12]. Despite the importance to women’s health, there are currently no UF-specific therapeutics because UFs are heterogeneous in composition and size among women, even within the same individual, and vary in number between individuals [13,14,15,16]. These irregularities highlight the challenge of preventing UFs and treating patients with UFs. Moreover, our understanding of the origin and pathogenesis of UFs continues to evolve.
2. Extracellular Matrix and Regulation in Uterine Fibroids
Excessive extracellular matrix (ECM) accumulation and aberrant remodeling are crucial for fibrotic diseases. An increased stiffness characterizes the fibrotic microenvironment, and this rigidity is associated with disease progression. The mechanical network of fibrotic ECM is regulated by ECM-degrading enzymes called matrix metalloproteinases (MMPs). MMPs are commonly classified on the basis of their substrates and the organization of their structural domains into collagenases, gelatinases, stromelysins, matrilysins, membrane-type (MT)-MMPs, and other MMPs. MMPs are often secreted in an inactive pro-MMP form, which is cleaved to the active form by various proteinases, including other MMPs. MMPs can be regulated by endogenous tissue inhibitors of metalloproteinases (TIMPs), and the MMP/TIMP ratio often determines the extent of ECM protein degradation and tissue remodeling [17]. UFs are characterized by the excessive deposition of ECM proteins, such as collagens, fibronectin, and proteoglycans, representing fibrosis [18,19,20]. In addition to MMPs and TIMPs, several factors impact ECM accumulation and deposition in UFs.
2.1. ECM and Hormones
UFs are considered hormone-dependent tumors, based on their association with reproductive age. Estrogen and progesterone are considered the principal promoters of UF growth [1,21,22]. Estrogen-related signaling impacts the biological process via genomic and nongenomic mechanisms. In addition, estrogen is capable of inducing the expression of the progesterone receptor (PR), stimulating progesterone-regulated signaling. Estrogen exerts multiple stimulatory actions on its target cells and accelerates collagen biosynthesis [23,24]. Three-dimensional UF cell cultures exposed to estrogen and medroxyprogesterone acetate (MPA) increased in their expression of collagen I and fibronectin [25].
FK506-binding protein 51 (FKBP51) is known as a chaperone that regulates the responsiveness of steroid hormone receptors. It was linked to several intracellular pathways related to tumorigenesis and chemoresistance [26,27]. FKBP51 was shown to bind PR, glucocorticoid receptor (GR), and androgen receptor (AR) to coregulate their transcriptional activity. It was reported that FKBP51 expression is higher in UFs compared to myometrial tissues. The knockdown of FKBP51 displayed decreased mRNA levels of ECM, TIMP1, and TIMP3, and reduced cell proliferation [28].
Prolactin is a hormone responsible for lactation and has been reported to be present and functional in UFs [29]. Prolactin strongly activates STAT5 and MAPK signaling in rat and human myometrial cell lines. Moreover, Prolactin produced from UFs may stimulate the transdifferentiation of the myometrium (MM) cells to myofibroblasts, contributing to the fibrotic nature of UFs [29].
2.2. ECM and Growth Factors
The ECM acts as a reservoir of profibrotic growth factors and enhances their activity by increasing their stability and prolonging signaling duration. Therefore, a better understanding of ECM composition and metabolism in UFs is critical for developing new therapeutics for UFs. In addition, several growth factors have been shown to trigger the ECM accumulation in UFs, including transforming growth factor-β, activin-A, and platelet-derived growth factor [24].
TGF-β initiates the cascade of signal transduction that elicits biological actions on responding cells via receptors on the plasma membrane. The central mechanism of signal transduction by the TGF-β family receptors follows a well-characterized process of interactions and receptor-mediated phosphorylation. Upon ligand binding and the following cascade steps, the trimeric complex (SMAD2, 3, 4) translocates into the nucleus and associates with high-affinity DNA binding transcription factors and chromatin remodeling proteins, thereby positively or negatively regulating the transcription of the TGF-β-responsive genes [30]. Numerous studies demonstrated that TGF-β signaling plays an important role in the pathogenesis of UFs. Abnormal ECM accumulation and deposition have been shown to be associated with the activation of TGF-β signaling in UFs. Several studies have demonstrated that TGF-β3 stimulates the production and secretion of ECM macromolecules and alters the expression of MMP members [24,31].
Activin-A is a member of the TGF-β superfamily, a large family of over 30 structurally related proteins. Activin A has been recognized as a multifunctional cytokine expressed in a wide range of tissues and cells, and growing evidence implicates activin A in the pathogenesis of UFs. In vitro studies demonstrated that Activin A promoted cell proliferation and increased ECM protein accumulation via p38 MAPK signaling in immortalized UF cells [32,33]. In addition, activin A significantly increased mRNA expression of FN, collagen 1A1, and versican in primary UF cells concomitantly with the activation of Smad-2/3 signaling, but not with changes in ERK and P38 signaling [34].
The platelet-derived growth factor (PDGF) family belongs to the growth factor systems. Its dysregulation is involved in a wide array of pathological conditions, such as fibrosis, neurological disorders, atherosclerosis, and tumorigenesis [35]. The PDGF family consists of four members: PDGF-A, PDGF-B, PDGF-C, and PDGF-D. Members of the PDGF family bind to and signal through the PDGF tyrosine kinase receptors with an extracellular ligand-binding domain and an intracellular tyrosine kinase domain. Upon ligand binding, the receptor dimerization results in receptor autophosphorylation on tyrosine residues. Autophosphorylation further activates the receptor kinase and docking sites for downstream signaling molecules and the modulation of different pathways [36]. PDGF is upregulated in about 80% of UFs compared to adjacent myometrial tissues [37] and can promote the growth of myometrial and UF-derived smooth muscle cells, which is one of the main cell populations contributing to ECM production and secretion. In addition, PDGF-C prolongs the survival of UF-derived SMCs in Matrigel plugs implemented subcutaneously in immunocompromised mice. Furthermore, PDGF can increase the collagen levels in both myometrial and UF cells [38].
2.3. ECM and Cytokines
Cytokines are small proteins with characteristics of intercellular messengers, which have a complex regulatory influence on inflammation and immunity. In addition, cytokines play an important role in many other biological processes, including tumorigenesis [39]. Proinflammatory cytokines have been shown to cause potent and consistent changes in ECM expression in many cell types [40,41]. Several cytokines have been implicated in the development of UFs [42]. Tumor necrosis factor-α (TNF-α) is a cell-signaling protein involved in systemic inflammation and is one of the cytokines responsible for the acute phase reaction. It was reported that TNF-α serum levels in women with clinically symptomatic UFs were significantly higher than in the control group [43]. TNF-a can increase the expression of activin-A, the ECM inducer, in UF and MM cells, suggesting that TNF-a may increase the deposition of ECM, leading to UF pathogenesis [24,44]. Further investigation of TNF-a-induced alterations in ECM production and components will help better understand the cytokine role in the ECM-mediated signal pathway in UFs.
2.4. Cell Types Contributing to ECM Production in Uterine Fibroids
UF heterogeneity exists at many levels, including etiology, clinical symptoms, and pathogenesis, which have significant ramifications for research design and therapeutic decisions [13,45]. The ECM forms a milieu surrounding cells that reciprocally influence cellular function to modulate diverse fundamental aspects of cell biology [46].
Intracellular heterogeneity is present in UFs with multiple cellular compositions [47]. Among them, SMCs and fibroblast populations are dominant in contributing to ECM secretion and participate in the collagen signaling network in the MED12-variant-positive UFs compared to the MM tissues [48]. Accordingly, increased SMC and fibroblast proliferation in UFs is correlated to enhanced ECM accumulation, the characteristic feature of UFs [37,49]. In addition, there was a significant increase in UF cells when cocultured with UF-derived fibroblasts. UF-derived fibroblasts can stimulate the production of collagen type I in the medium cocultured with UF cells [50].
3. ECM and Downstream Signaling
Several studies have demonstrated that ECM contributes to mechanotransduction in UFs. Excessive ECM deposition and abnormal remodeling can regulate downstream signaling in UF cells.
3.1. Stiff ECM and Progesterone Receptor Signaling
The uterine myometrium consists of various cell types and is interspersed with interstitial ECM. Cells grown on mechanically stiff ECM substrate promote progesterone receptor activation via MEK1/2 and AKAP/RhoA/ROCK signaling pathways. Accordingly, UF cells exhibited higher RNA levels of collagen I grown on stiff collagen I-coated- plates compared to soft collagen I-coated plates in response to progesterone treatment [51].
3.2. The Effect of Collagen Cross-Linking on Proliferation and Resistance to MMP Proteolytic Degradation
Collagen, as one of the major ECM components, contains various types of collagens. The amount of collagen can influence the conversion of signals into chemical changes, and intramolecular and intermolecular Lysyl oxidase cross-links can form the mature collagen fibrils that impact these functional signals [20]. The expression levels of collagen cross-linking enzymes lysyl hydroxylases (LH) and lysyl oxidases (LOX) are higher in UFs than MM tissues linked to increased collagen cross-links and resistance to MMP proteolytic degradation, highlighting the role of cross-linking in ECM remodeling. In addition, increased collagen cross-links positively correlated to the UF size and cell-proliferation-related markers [52].
3.3. ECM Stiffness and Other Signaling
It has been reported that ECM stiffness impacts several signaling pathways in UFs. The Wnt signaling pathway is involved in various cellular processes such as embryogenesis, tissue renewal, cell proliferation, differentiation, and tumorigenesis [53,54,55,56]. In canonical Wnt-on signaling, β-catenin avoids destruction in the cytoplasm and translocates into the nucleus. Subsequently, the nuclear β-catenin binds to the TCF/LEF transcription factors and accelerates β-catenin-regulated gene expression. In canonical Wnt-off signaling, a combination of AXIN and APC allows GSK3β to phosphorylate β-catenin and targets it for proteasomal degradation. Wnt/β-catenin signaling is abnormally activated in UFs compared to MM [57], and the mislocalization of β-catenin is correlated to the UF phenotype [58]. Moreover, estrogen triggers β-catenin nuclear translocation and enhanced β-catenin-responsive gene expression in human UF cells. An increase in ECM stiffness occurred when primary UF cells were grown on hydrogels of known stiffness and triggered the upregulation of β-catenin expression in UF cells [57].
The Hippo signaling pathway is highly conserved and plays a critical role in tumorigenesis. It is characterized by the phosphorylation of YAP1 and TAZ. Several factors can regulate the localization of Yes-associated protein (YAP), therefore modulating the downstream signaling. Hippo pathways can integrate mechanical signaling with cell growth. YAP is a key transcriptional coactivator of the Hippo pathway and acts as a sensor and regulator of a wide range of physical and mechanical stresses, including the stiffness of the ECM [59]. The increased ECM stiffness activates YAP, promoting cell growth and fibrotic gene transcription. On the other hand, in the soft matrix, YAP becomes phosphorylated or degraded in the cytoplasm. When YAP, a suppressor of the YAP-TEAD complex, is phosphorylated, it becomes transcriptionally inactive. A study by Purdy et al. showed that decreased substrate stiffness reduced YAP/TAZ nuclear localization in both the MM and UFs [60]. The pharmacological inhibition of YAP with verteporfin reduced key targets of fibrosis and genes involved in mechanotransduction in UF cells. In addition, the antifibrotic drug nintedanib inhibited YAP and showed antifibrotic effects on UF cells with decreased fibronectin levels [61].
4. Targeting ECM
4.1. Targeting Hormone-Dependent Growth with ECM Changes
The significant variations in the quantity and distribution of the ECM also create variation in the degree of UF tissue stiffness from soft to firm to solid to hard, with a major impact on the gene expression and biology of these tumor lesions. UF growth is regulated by hormone levels in the body (estrogen and progesterone are both promoters of UF growth), providing the basis for new therapy developments, with others in the pipeline. Hormonal contraception containing estrogen and progestin effectively prevents unintended pregnancy and lowers the risk of some female cancers. However, it may increase other risks and cause some side effects in women. Current pharmacological treatment options for UFs aim to halt their hormonal-dependent growth using gonadotropin-releasing hormone (GnRH) analogs (agonists and antagonists). For example, Bozzini et al. [62] identified significant positive correlations between UF volume reduction and collagen content, progesterone receptors, and blood supply, and negative correlations with estrogen receptors after treatment with GnRH agonists (goserelin). The following efforts aimed to target the GnRH to improve outcomes and reduce hypoestrogenic side effects, such as bone loss, in clinical trials. Such concerns led to the hypothesis that one might maximize the benefits of blocking UF-associated menstrual bleeding and minimize the hypoestrogenic side effect. This idea led to the design of a series of studies that demonstrate that new-generation oral GnRH antagonists, including elagolix [63], relugolix [64], and linzagolix [65], are promising, well tolerated, and noninvasive options, and offer potential prospects for the future therapy of UFs. These clinical trial studies support the hormone threshold approach to increase the quality of life of premenopausal women with clinically significant UFs. Notably, a recent study demonstrated that the new-generation GnRH antagonists relugolix and elagolix decreased the production of the ECM component fibronectin in UF cells, accompanied by MAPK inhibition [66].
4.2. Collagenase
Since UFs are collagen-rich, fibrotic tumors, the digestion of collagen would degrade the ECM, altering the stiffness. The injection of a highly purified form of collagenase Clostridium histolyticum into UFs resulted in a remarkable reduction in fibrosis and stiffness, concomitantly with a decrease in cell proliferation markers [61,67].
4.3. Vitamin D and ECM
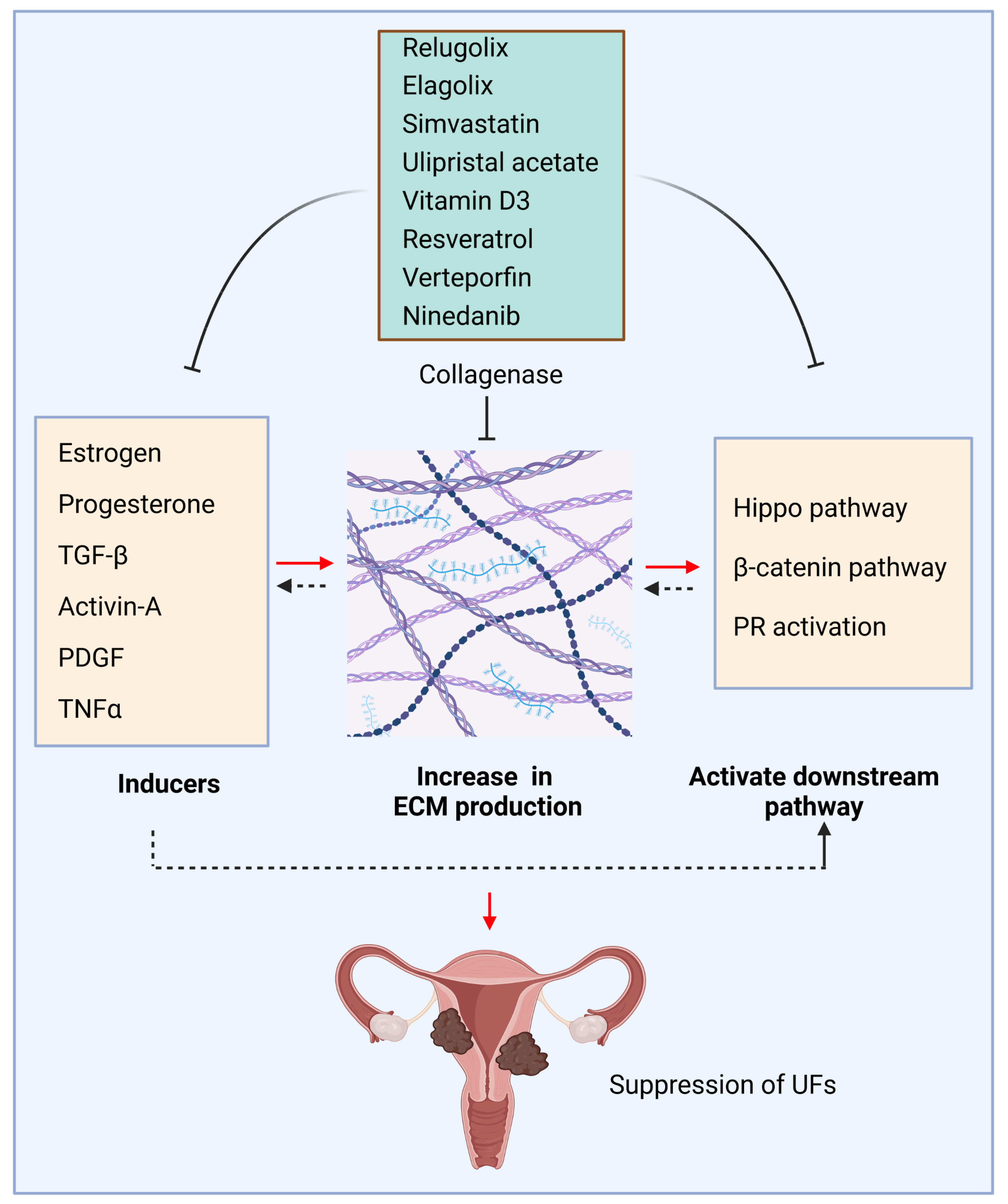
Vitamin D forms a complex with a specific receptor named the vitamin D receptor (VDR) to mediate its pleiotropic functions via steroid transcriptional mechanisms [1]. Vitamin D deficiency is one of the critical risk factors for UF pathogenesis [68,69]. Our previous studies demonstrated that vitamin D3 reduced UF cell proliferation in vitro and UF growth in vivo animal models. Vitamin D3 decreased key ECM components, including collagen type I, fibronectin, and proteoglycans [70]. In addition, Vitamin D3 altered the expression and activities of matrix metalloproteinase 2 and 9 in human UF cells, indicating its role in ECM remodeling. These results underscore the potential role of vitamin D3 in an effective, safe, nonsurgical medical treatment option for UFs [71]. The key ECM inducers and relevant pathways and inhibitors are summarized in Table 1 and Figure 1.
4.4. Epigenetic-Mediated ECM Changes
HDACs are a class of enzymes that remove acetyl groups from an N-acetyl lysine amino acid on a histone, allowing the histones to wrap the DNA more tightly, thereby regulating gene expression. In addition, nonhistone proteins are also targeted by HATs and HDACs, modulating various biological events. Studies have demonstrated that abnormal HDAC signaling contributes to many diseases, including tumorigenesis [72,73,74]. In UFs, HDAC expression and activity are abnormally upregulated compared to MM tissues [58,75]. In one study, the HDAC inhibitor suberoylanilide hydroxamic acid (SAHA) selectively inhibited UF growth but not MM cells [75]. In addition, SAHA significantly inhibited ECM production, including FN and collagen I, accompanied by a decrease in TGF-β3, MMP9, and cell-cycle-related markers (C-MYC and CCND1) [75]. Resveratrol is a polyphenolic phytoalexin found in peanuts, grapes, and other plants with characteristics of antioxidant and anti-HDAC activity [76]. Resveratrol treatment suppressed UF growth in a UF xenograft model and decreased the expression levels of collagen 1, a-SMA, and FN, demonstrating the potential role of the antifibrotic effect of HDAC inhibition [77].
Bromodomains (BRDs) are evolutionarily conserved protein–protein interaction modules that selectively recognize and bind to acetylated lysine residues—particularly in histones—and thereby have important roles in regulating gene expression. Bromodomains (BRD)-containing proteins are responsible for transducing regulatory signals from acetylated lysine residues into various biological phenotypes. BRD proteins can have various functions via multiple gene regulatory mechanisms, including chromatin remodeling, histone modification, histone recognition, and scaffolding, and various activities in transcriptional coregulation; therefore, they can lead to the alteration of gene transcription [78,79]. The BET (bromodomain and extraterminal domain) family of proteins, consisting of BRD2, BRD3, BRD4, and testis-specific BRDT, are widely acknowledged as major transcriptional regulators in biology. They are characterized by two tandem bromodomains (BRDs) that bind to lysine-acetylated histones and transcription factors, recruit transcription factors and coactivators to target gene sites, and activate RNA polymerase II machinery for transcriptional elongation. The pharmacological inhibition of BET proteins with BRD inhibitors has been shown as a promising therapeutic strategy for treating many human diseases, including cancer [80]. BRD4 expression is abnormally upregulated in breast cancer tissues and cells, and the targeted inhibition of BRD4 by pharmacological inhibitors and the genetic loss of function analysis have been shown to significantly inhibit the malignancy of breast cancer cells and concomitantly decrease the fibronectin protein levels [81]. Attention has now expanded to the biological role of non-BET BRD proteins. Among them, non-BET BRD-containing protein 9 (BRD9) is associated with various diseases, including tumorigenesis. Our recent studies demonstrated that the pharmacological inhibition of non-BET BRD9 suppressed the UF phenotype with decreased ECM proteins, including fibronectin in UF cells [82], reinforcing the view that BRD proteins may be involved in the pathogenesis of UFs.
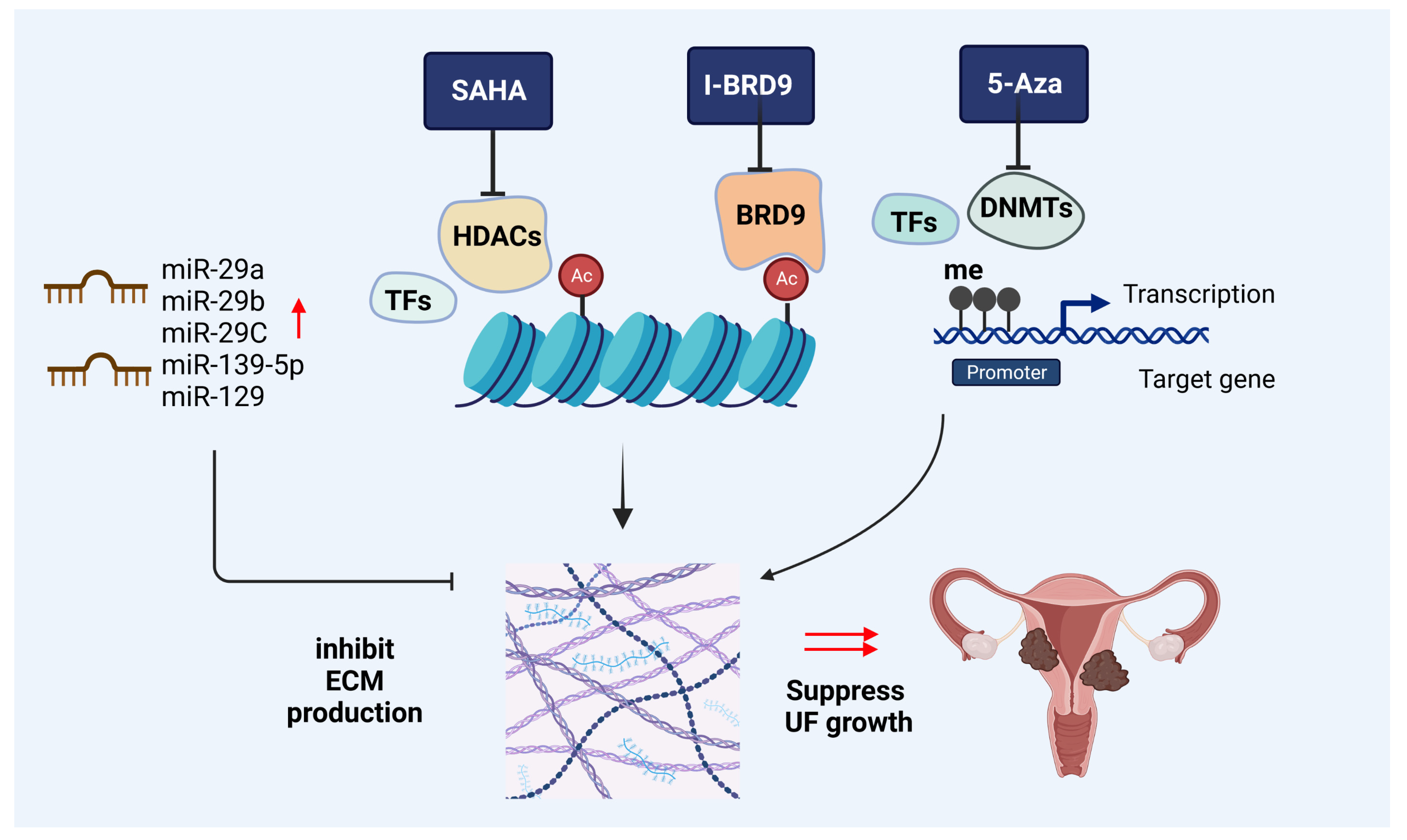
DNA methylation is one of the common epigenetic regulation mechanisms in eukaryotes. The hypermethylation of the CpG island in the promoter region generally results in the repression of gene expression, while hypomethylation leads to active transcription. Cytosine methylation is catalyzed by specific DNA methyltransferases (DNMTs) that transfer a methyl group from the donor S-adenosyl methionine to the 5′-position of the pyrimidinic ring. The cytosine methylation can be oxidized by the TET dioxygenases to ultimately cause DNA demethylation [83]. Aberrant DNA methylation dynamics occur in many diseases, including tumorigenesis. In UFs, abnormal DNA methylation is involved in the pathogenesis of UFs [83,84,85]. DNMT activity was shown to be upregulated, indicating an important role of DNA methylation in UFs [86]. Targeting DNMT with 5-aza-2′-deoxycytidine, the DNMT inhibitor, reduced the collagen I levels, concomitantly with a decrease in the wnt/βcatenin pathway, MMP7, and c-MYC in UF cells [86] (Figure 2).
MicroRNAs (or miRNAs) are small, noncoding RNAs (∼22 nt long) that regulate post-transcriptional gene expression. By binding to the target mRNAs’ 3′-UTR (untranslated region), miRNAs prevent protein production by inducing mRNA degradation and/or directly repressing translation. A number of studies demonstrated that miRNAs play an important role in the pathogenesis of UFs [9,87,88,89,90]. The expression levels of the mir-29 family, including mir-29a, mir-29b, and mir-29c, were downregulated in UFs compared to the MM tissues. The overexpression of each mir-29 family member reduced the ECM production in UF cells [91,92,93,94]. Another microRNA, miR-139-5p, regulated fibrotic potentials via the modulation of collagen type I and phosphorylated p38 MAPK in UFs [95]. The expression of miR-129 was also lower in UFs. The targeted inhibition of mir-129 modulated the accumulation of ECM, accompanied by a decrease in cell proliferation and TET1 expression [96].
These studies suggested that the epigenetic targeting of UF could reverse the UF phenotype via decreasing ECM composition and accumulation, one of the key hallmarks of UFs (Figure 2).
4.5. ECM and Other Inhibitors
Several other factors have been shown to regulate ECM production and stiffness (Table 1). Simvastatin is used to help lower “bad” cholesterol and fats and raise “good” cholesterol, and exhibits anti-UF effects both in vitro and in vivo [97,98]. The treatment of UF cells with simvastatin decreased the production of collagen, fibronectin, versican, and brevican and reduced the levels of mechanical signaling proteins involved in β1 integrin downstream signaling [99,100].
Ulipristal acetate as a selective progesterone receptor modulator showed a decrease in the total volume of UFs and an improvement in quality-of-life measures [101,102,103]. In addition, in a randomized control study, the administration of ulipristal acetate decreased fibronectin, and versican, concomitantly altering the expression levels of MMP2 and MMP9 [104].

{kind=link}
{kind=link}
Table 1.
Summarized ECM research in uterine fibroids since 2018.
| Inducers/Inhibitors | Biological Samples | Changes in ECM | Approach | Co-Changes | Publication Time | References |
|---|---|---|---|---|---|---|
| Activin A | UF cell line | Increase in ECM accumulation | WB | p38 MAPK | July 2021 | [33] |
| Activin A | UF cell line | Excessive ECM | WB, qPCR, and IC | p38 MAPK | October 2018 | [32] |
| Butylated hydroxytoluene | ELT-3 UF cell line | Increase in CO1A1 | WB, IF | PI3K/AKT and MAPK | August 2021 | [105] |
| Leptin | UF cell line | Increase in ECM formation | JAK2/STAT3 and MAPK/ERK | May 2022 | [106] | |
| TBBPA | 3D human UF spheroids | Increase in collagen and fibrosis | Masson’s trichrome stain, PCR, and light microscopy | TGF-β signaling | February 2022 | [107] |
| Cadmium prolonged exposure | UF cells | Decrease in collagens, FNs, laminins; increase in MMPs | IF and MMP antibody array | TGF-β signaling, cell proliferation, and migration | August 2021 | [108] |
| Collagenase Clostridium histolyticum | UF tissues | Decrease in ECM stiffness | Rheometry | Cell proliferation and Hippo signaling | July 2021 | [61] |
| CRSR | UF rat model | Decrease in ECM deposition | Microarray | MAPK, PPAR, Notch, and TGF-β | May 2019 | [109] |
| Decrease in FKBP51 | UF cells | Decrease in ECM formation | qPCR | Cell survival and proliferation | June 2022 | [28] |
| Fucoidan | ELT-3, human UF cells, xenograft model | Decrease in FN and COL1A1 | WB | TGF-β3 signaling | September 2018 | [110] |
| Isoliquiritigenin | ELT3, UtSMC, uterine myometrium hyperplasia mouse model | Decrease in ECM accumulation | WB and IHC | MMP, ERK1/2, p38, and JNK | August 2019 | [111] |
| miR-139-5p | UF cell line | Decrease in contractility of the ECM | Migration, collagen gel contraction, and wound healing | p38 MAPK | August 2021 | [95] |
| miR-21a-5p | UF and MM cell lines | FN, Collagen 1A1, CTGF, versican, and DPT | qPCR and collagen assay | TGF-β3 and MMPs | May 2018 | [112] |
| Relugolix and elagolix | UF cells | Decrease in collagen 1A1, FN, and versican | pERK/ERK | August 2022 | [66] | |
| Resveratrol | Xenograft and UF primary cells | Decrease in collagen I, FN, and a-SMA | WB, qPCR | Proliferation and apoptosis | April 2019 | [77] |
| Simvastatin | UF SCs | Decrease in collagen I and fibronectin | WB | TGF-β and β-catenin signaling | February 2022 | [97] |
| Simvastatin | UF cell lines and primary cells, 2D and 3D | Decrease in collagen I, collagen III, FN, versican, and brevican | WB and IHC | Apoptosis | December 2018 | [99] |
| S1P | UF and myometrium cells | Decrease in FN and collagen 1A1 | qPCR | Decrease in activin A | June 2021 | [113] |
| Butylated hydroxytoluene | UF primary cells | Decrease in collagen I and fibronectin | WB | Cell cycle and TGF-β3 signaling | February 2022 | [75] |
| Ultrasound-guided collagenase injection | UF tissues | Decrease in content, density, and fibers of ECM | Masson’s trichrome stain, second harmonic generation, and Picrosirius stain | Decrease in UF-related pain | September 2021 | [114] |
| Ulipristal acetate | UF patients | Decrease in versican and FN1 | Masson trichrome staining and IHC | Decrease in UF size | February 2018 | [104] |
| Vitamin D | Xenograft | Decrease in collagen I and plasminogen activator inhibitor 1 | WB | TGF-β3 signaling | January 2020 | [115] |
5. Conclusions and Future Perspectives
Although the heterogeneity of UFs is present and underscores the challenge for the consideration of basic, translational, or clinical studies or standard clinical practice, considerable progress has been made in recent years to study the role and mechanisms underlying the ECM-mediated cellular and molecular events which contribute to the pathogenesis of UFs. ECM deposition and remodeling can be regulated via hormones, growth factors, cytokines, and MMPs. Excessive ECM can induce mechanotransduction through integrin activation and increase tissue stiffness by altering bidirectional signaling, which leads to UF progression. Pharmacological drugs can suppress the UF phenotype by inhibiting ECM production via multiple mechanisms, including epigenetics (Figure 2). However, several aspects need to be further elucidated, including (1) characterizing the functional interaction of ECM-secreted cells with other types of cells, (2) elucidating the impact of ECM on the microenvironment, (3) characterizing the ECM-mediated signaling via its interactions with other biological pathways via autocrine and or paracrine mechanisms, and (4) investigating the molecular and functional structure of ECM that regulates mechanotransduction and stiffness in both UFs and the at-risk MM. The deep mechanistic and functional investigation will help us better understand the regulatory mechanism of ECM-secreted cell types, cell–cell interaction, and environmental impacts on the risk and development of UFs. The clinical application of ECM/stiffness measurement and molecular inhibitors targeting the ECM might provide a promising option for the personalized treatment of patients with UFs.
Author Contributions
Q.Y. conceived the manuscript, Q.Y. wrote the original manuscript, Q.Y. and A.A.-H. edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported in part by the National Institutes of Health (grant numbers: RO1 ES028615, RO1 HD094378, U54 MD007602, RO1 HD094380, and HD106285).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yang, Q.; Ciebiera, M.; Bariani, M.V.; Ali, M.; Elkafas, H.; Boyer, T.G.; Al-Hendy, A. Comprehensive Review of Uterine Fibroids: Developmental Origin, Pathogenesis, and Treatment. Endocr. Rev. 2022, 43, 678–719. [Google Scholar] [CrossRef]
- Segars, J.H.; Al-Hendy, A. Uterine Leiomyoma: New Perspectives on an Old Disease. Semin. Reprod. Med. 2017, 35, 471–472. [Google Scholar] [CrossRef]
- Bulun, S.E.; Moravek, M.B.; Yin, P.; Ono, M.; Coon, J.S.t.; Dyson, M.T.; Navarro, A.; Marsh, E.E.; Zhao, H.; Maruyama, T.; et al. Uterine Leiomyoma Stem Cells: Linking Progesterone to Growth. Semin. Reprod. Med. 2015, 33, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, C.M.; Steiner, C.A. Hysterectomy rates in the United States 1990–1997. Obstet. Gynecol. 2002, 99, 229–234. [Google Scholar] [CrossRef]
- Cardozo, E.R.; Clark, A.D.; Banks, N.K.; Henne, M.B.; Stegmann, B.J.; Segars, J.H. The estimated annual cost of uterine leiomyomata in the United States. Am. J. Obstet. Gynecol. 2012, 206, 211.e1–e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mas, A.; Stone, L.; O’Connor, P.M.; Yang, Q.; Kleven, D.; Simon, C.; Walker, C.L.; Al-Hendy, A. Developmental Exposure to Endocrine Disruptors Expands Murine Myometrial Stem Cell Compartment as a Prerequisite to Leiomyoma Tumorigenesis. Stem Cells 2017, 35, 666–678. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Ali, M.; El Andaloussi, A.; Al-Hendy, A. The emerging spectrum of early life exposure-related inflammation and epigenetic therapy. Cancer Stud. Mol. Med. 2018, 4, 13–23. [Google Scholar] [CrossRef]
- Orciani, M.; Caffarini, M.; Biagini, A.; Lucarini, G.; Delli Carpini, G.; Berretta, A.; Di Primio, R.; Ciavattini, A. Chronic Inflammation May Enhance Leiomyoma Development by the Involvement of Progenitor Cells. Stem Cells Int. 2018, 2018, 1716246. [Google Scholar] [CrossRef]
- Chuang, T.D.; Khorram, O. miR-200c regulates IL8 expression by targeting IKBKB: A potential mediator of inflammation in leiomyoma pathogenesis. PLoS ONE 2014, 9, e95370. [Google Scholar] [CrossRef] [PubMed]
- Muralimanoharan, S.; Shamby, R.; Stansbury, N.; Schenken, R.; de la Pena Avalos, B.; Javanmardi, S.; Dray, E.; Sung, P.; Boyer, T.G. Aberrant R-loop-induced replication stress in MED12-mutant uterine fibroids. Sci. Rep. 2022, 12, 6169. [Google Scholar] [CrossRef] [PubMed]
- Prusinski Fernung, L.E.; Yang, Q.; Sakamuro, D.; Kumari, A.; Mas, A.; Al-Hendy, A. Endocrine disruptor exposure during development increases incidence of uterine fibroids by altering DNA repair in myometrial stem cells. Biol. Reprod. 2018, 99, 735–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Nair, S.; Laknaur, A.; Ismail, N.; Diamond, M.P.; Al-Hendy, A. The Polycomb Group Protein EZH2 Impairs DNA Damage Repair Gene Expression in Human Uterine Fibroids. Biol. Reprod. 2016, 94, 69. [Google Scholar] [CrossRef] [PubMed]
- Stewart, E.A.; Laughlin-Tommaso, S.K.; Catherino, W.H.; Lalitkumar, S.; Gupta, D.; Vollenhoven, B. Uterine fibroids. Nat. Rev. Dis. Primers. 2016, 2, 16043. [Google Scholar] [CrossRef] [PubMed]
- Bulun, S.E. Uterine fibroids. N. Engl. J. Med. 2013, 369, 1344–1355. [Google Scholar] [CrossRef] [Green Version]
- Jayes, F.L.; Liu, B.; Feng, L.; Aviles-Espinoza, N.; Leikin, S.; Leppert, P.C. Evidence of biomechanical and collagen heterogeneity in uterine fibroids. PLoS ONE 2019, 14, e0215646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yatsenko, S.A.; Mittal, P.; Wood-Trageser, M.A.; Jones, M.W.; Surti, U.; Edwards, R.P.; Sood, A.K.; Rajkovic, A. Highly heterogeneous genomic landscape of uterine leiomyomas by whole exome sequencing and genome-wide arrays. Fertil. Steril. 2017, 107, 457–466.e459. [Google Scholar] [CrossRef] [Green Version]
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and Biological Attributes of Matrix Metalloproteinases. Prog. Mol. Biol. Transl. Sci. 2017, 147, 1–73. [Google Scholar] [CrossRef] [Green Version]
- Rogers, R.; Norian, J.; Malik, M.; Christman, G.; Abu-Asab, M.; Chen, F.; Korecki, C.; Iatridis, J.; Catherino, W.H.; Tuan, R.S.; et al. Mechanical homeostasis is altered in uterine leiomyoma. Am. J. Obstet. Gynecol. 2008, 198, 474.e1–474.e11. [Google Scholar] [CrossRef] [Green Version]
- Rafique, S.; Segars, J.H.; Leppert, P.C. Mechanical Signaling and Extracellular Matrix in Uterine Fibroids. Semin. Reprod. Med. 2017, 35, 487–493. [Google Scholar] [CrossRef]
- Leppert, P.C.; Jayes, F.L.; Segars, J.H. The extracellular matrix contributes to mechanotransduction in uterine fibroids. Obstet. Gynecol. Int. 2014, 2014, 783289. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, H.; Ishi, K.; Serna, V.A.; Kakazu, R.; Bulun, S.E.; Kurita, T. Progesterone is essential for maintenance and growth of uterine leiomyoma. Endocrinology 2010, 151, 2433–2442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borahay, M.A.; Asoglu, M.R.; Mas, A.; Adam, S.; Kilic, G.S.; Al-Hendy, A. Estrogen Receptors and Signaling in Fibroids: Role in Pathobiology and Therapeutic Implications. Reprod. Sci. 2017, 24, 1235–1244. [Google Scholar] [CrossRef]
- Zbucka, M.; Miltyk, W.; Bielawski, T.; Surazynski, A.; Palka, J.; Wolczynski, S. Mechanism of collagen biosynthesis up-regulation in cultured leiomyoma cells. Folia. Histochem. Cytobiol. 2007, 45 (Suppl. S1), S181–S185. [Google Scholar] [PubMed]
- Islam, M.S.; Ciavattini, A.; Petraglia, F.; Castellucci, M.; Ciarmela, P. Extracellular matrix in uterine leiomyoma pathogenesis: A potential target for future therapeutics. Hum. Reprod. Update 2018, 24, 59–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, M.; Britten, J.; Cox, J.; Patel, A.; Catherino, W.H. Gonadotropin-releasing hormone analogues inhibit leiomyoma extracellular matrix despite presence of gonadal hormones. Fertil. Steril. 2016, 105, 214–224. [Google Scholar] [CrossRef] [Green Version]
- Rein, T. FK506 binding protein 51 integrates pathways of adaptation: FKBP51 shapes the reactivity to environmental change. Bioessays 2016, 38, 894–902. [Google Scholar] [CrossRef]
- Li, L.; Lou, Z.; Wang, L. The role of FKBP5 in cancer aetiology and chemoresistance. Br. J. Cancer 2011, 104, 19–23. [Google Scholar] [CrossRef] [Green Version]
- New, E.P.; Semerci, N.; Ozmen, A.; Guo, X.; Jonnalagadda, V.A.; Kim, J.W.; Anderson, M.L.; Guzeloglu-Kayisli, O.; Imudia, A.N.; Lockwood, C.J.; et al. FKBP51 Contributes to Uterine Leiomyoma Pathogenesis by Inducing Cell Proliferation and Extracellular Matrix Deposition. Reprod. Sci. 2022, 29, 1939–1949. [Google Scholar] [CrossRef]
- DiMauro, A.; Seger, C.; Minor, B.; Amitrano, A.M.; Okeke, I.; Taya, M.; Rackow, A.R.; Kumar, D.; Kottman, R.M.; Bhagavath, B.; et al. Prolactin is Expressed in Uterine Leiomyomas and Promotes Signaling and Fibrosis in Myometrial Cells. Reprod. Sci. 2022, 29, 2525–2535. [Google Scholar] [CrossRef]
- Yang, Q.; Al-Hendy, A. The Regulatory Functions and the Mechanisms of Long Non-Coding RNAs in Cervical Cancer. Cells 2022, 11, 1149. [Google Scholar] [CrossRef]
- Joseph, D.S.; Malik, M.; Nurudeen, S.; Catherino, W.H. Myometrial cells undergo fibrotic transformation under the influence of transforming growth factor beta-3. Fertil. Steril. 2010, 93, 1500–1508. [Google Scholar] [CrossRef]
- Bao, H.; Sin, T.K.; Zhang, G. Activin A induces leiomyoma cell proliferation, extracellular matrix (ECM) accumulation and myofibroblastic transformation of myometrial cells via p38 MAPK. Biochem. Biophys. Res. Commun. 2018, 504, 447–453. [Google Scholar] [CrossRef]
- Bao, H.; Sin, T.K.; Zhang, G. Activin A induces tumorigenesis of leiomyoma via regulation of p38beta MAPK-mediated signal cascade. Biochem. Biophys. Res. Commun. 2020, 529, 379–385. [Google Scholar] [CrossRef]
- Islam, M.S.; Catherino, W.H.; Protic, O.; Janjusevic, M.; Gray, P.C.; Giannubilo, S.R.; Ciavattini, A.; Lamanna, P.; Tranquilli, A.L.; Petraglia, F.; et al. Role of activin-A and myostatin and their signaling pathway in human myometrial and leiomyoma cell function. J. Clin. Endocrinol. Metab. 2014, 99, E775–E785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadopoulos, N.; Lennartsson, J. The PDGF/PDGFR pathway as a drug target. Mol. Aspects Med. 2018, 62, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Folestad, E.; Kunath, A.; Wagsater, D. PDGF-C and PDGF-D signaling in vascular diseases and animal models. Mol. Aspects Med. 2018, 62, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Suo, G.; Jiang, Y.; Cowan, B.; Wang, J.Y. Platelet-derived growth factor C is upregulated in human uterine fibroids and regulates uterine smooth muscle cell growth. Biol. Reprod. 2009, 81, 749–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, M.; Wang, H.; Zhang, Y.; Lu, S.; Wang, Z. Expression and functional analysis of platelet-derived growth factor in uterine leiomyomata. Cancer Biol. Ther. 2006, 5, 28–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, I.L. Cytokine-mediated inflammation, tumorigenesis, and disease-associated JAK/STAT/SOCS signaling circuits in the CNS. Brain Res. Brain Res. Rev. 2005, 48, 166–177. [Google Scholar] [CrossRef]
- Giblin, M.J.; Ontko, C.D.; Penn, J.S. Effect of cytokine-induced alterations in extracellular matrix composition on diabetic retinopathy-relevant endothelial cell behaviors. Sci. Rep. 2022, 12, 12955. [Google Scholar] [CrossRef]
- Uitto, J.; Kouba, D. Cytokine modulation of extracellular matrix gene expression: Relevance to fibrotic skin diseases. J. Dermatol. Sci. 2000, 24 (Suppl. S1), S60–S69. [Google Scholar] [CrossRef]
- Ciavattini, A.; Di Giuseppe, J.; Stortoni, P.; Montik, N.; Giannubilo, S.R.; Litta, P.; Islam, M.S.; Tranquilli, A.L.; Reis, F.M.; Ciarmela, P. Uterine fibroids: Pathogenesis and interactions with endometrium and endomyometrial junction. Obstet. Gynecol. Int. 2013, 2013, 173184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciebiera, M.; Wlodarczyk, M.; Wrzosek, M.; Wojtyla, C.; Blazej, M.; Nowicka, G.; Lukaszuk, K.; Jakiel, G. TNF-alpha serum levels are elevated in women with clinically symptomatic uterine fibroids. Int. J. Immunopathol. Pharmacol. 2018, 32, 2058738418779461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, M.S.; Protic, O.; Giannubilo, S.R.; Toti, P.; Tranquilli, A.L.; Petraglia, F.; Castellucci, M.; Ciarmela, P. Uterine leiomyoma: Available medical treatments and new possible therapeutic options. J. Clin. Endocrinol. Metab. 2013, 98, 921–934. [Google Scholar] [CrossRef] [Green Version]
- Peddada, S.D.; Laughlin, S.K.; Miner, K.; Guyon, J.P.; Haneke, K.; Vahdat, H.L.; Semelka, R.C.; Kowalik, A.; Armao, D.; Davis, B.; et al. Growth of uterine leiomyomata among premenopausal black and white women. Proc. Natl. Acad. Sci. USA 2008, 105, 19887–19892. [Google Scholar] [CrossRef] [Green Version]
- Lu, P.; Takai, K.; Weaver, V.M.; Werb, Z. Extracellular matrix degradation and remodeling in development and disease. Cold Spring Harb. Perspect Biol. 2011, 3. [Google Scholar] [CrossRef] [PubMed]
- Holdsworth-Carson, S.J.; Zaitseva, M.; Vollenhoven, B.J.; Rogers, P.A. Clonality of smooth muscle and fibroblast cell populations isolated from human fibroid and myometrial tissues. Mol. Hum. Reprod. 2014, 20, 250–259. [Google Scholar] [CrossRef] [Green Version]
- Goad, J.; Rudolph, J.; Zandigohar, M.; Tae, M.; Dai, Y.; Wei, J.J.; Bulun, S.E.; Chakravarti, D.; Rajkovic, A. Single-cell sequencing reveals novel cellular heterogeneity in uterine leiomyomas. Hum. Reprod. 2022, 37, 2334–2349. [Google Scholar] [CrossRef] [PubMed]
- Luo, N.; Guan, Q.; Zheng, L.; Qu, X.; Dai, H.; Cheng, Z. Estrogen-mediated activation of fibroblasts and its effects on the fibroid cell proliferation. Transl. Res. 2014, 163, 232–241. [Google Scholar] [CrossRef]
- Moore, A.B.; Yu, L.; Swartz, C.D.; Zheng, X.; Wang, L.; Castro, L.; Kissling, G.E.; Walmer, D.K.; Robboy, S.J.; Dixon, D. Human uterine leiomyoma-derived fibroblasts stimulate uterine leiomyoma cell proliferation and collagen type I production, and activate RTKs and TGF beta receptor signaling in coculture. Cell Commun. Signal. 2010, 8, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordeiro Mitchell, C.N.; Islam, M.S.; Afrin, S.; Brennan, J.; Psoter, K.J.; Segars, J.H. Mechanical stiffness augments ligand-dependent progesterone receptor B activation via MEK 1/2 and Rho/ROCK-dependent signaling pathways in uterine fibroid cells. Fertil. Steril. 2021, 116, 255–265. [Google Scholar] [CrossRef]
- Kamel, M.; Wagih, M.; Kilic, G.S.; Diaz-Arrastia, C.R.; Baraka, M.A.; Salama, S.A. Overhydroxylation of Lysine of Collagen Increases Uterine Fibroids Proliferation: Roles of Lysyl Hydroxylases, Lysyl Oxidases, and Matrix Metalloproteinases. Biomed. Res. Int. 2017, 2017, 5316845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, W.; Khan, S.K.; Gvozdenovic-Jeremic, J.; Kim, Y.; Dahlman, J.; Kim, H.; Park, O.; Ishitani, T.; Jho, E.H.; Gao, B.; et al. Hippo signaling interactions with Wnt/beta-catenin and Notch signaling repress liver tumorigenesis. J. Clin. Investig. 2017, 127, 137–152. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, M.K.; Shao, C.; Wang, J.; Wei, Q.; Wang, X.; Collier, Z.; Tang, S.; Liu, H.; Zhang, F.; Huang, J.; et al. Wnt/beta-catenin signaling plays an ever-expanding role in stem cell self-renewal, tumorigenesis and cancer chemoresistance. Genes. Dis. 2016, 3, 11–40. [Google Scholar] [CrossRef] [Green Version]
- Deng, F.; Peng, L.; Li, Z.; Tan, G.; Liang, E.; Chen, S.; Zhao, X.; Zhi, F. YAP triggers the Wnt/beta-catenin signalling pathway and promotes enterocyte self-renewal, regeneration and tumorigenesis after DSS-induced injury. Cell Death Dis. 2018, 9, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moumen, M.; Chiche, A.; Decraene, C.; Petit, V.; Gandarillas, A.; Deugnier, M.A.; Glukhova, M.A.; Faraldo, M.M. Myc is required for beta-catenin-mediated mammary stem cell amplification and tumorigenesis. Mol. Cancer 2013, 12, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, Y.A.; Jamaluddin, M.F.B.; Adebayo, M.; Bajwa, P.; Scott, R.J.; Dharmarajan, A.M.; Nahar, P.; Tanwar, P.S. Extracellular matrix (ECM) activates beta-catenin signaling in uterine fibroids. Reproduction 2018, 155, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Shahin, S.M.; Sabri, N.A.; Al-Hendy, A.; Yang, Q. Activation of beta-Catenin Signaling and its Crosstalk With Estrogen and Histone Deacetylases in Human Uterine Fibroids. J. Clin. Endocrinol. Metab. 2020, 105, e1517–e1535. [Google Scholar] [CrossRef]
- Feng, J.; Gou, J.; Jia, J.; Yi, T.; Cui, T.; Li, Z. Verteporfin, a suppressor of YAP-TEAD complex, presents promising antitumor properties on ovarian cancer. Onco. Targets Ther. 2016, 9, 5371–5381. [Google Scholar] [CrossRef] [Green Version]
- Purdy, M.P.; Ducharme, M.; Haak, A.J.; Ravix, J.; Tan, Q.; Sicard, D.; Prakash, Y.S.; Tschumperlin, D.J.; Stewart, E.A. YAP/TAZ are Activated by Mechanical and Hormonal Stimuli in Myometrium and Exhibit Increased Baseline Activation in Uterine Fibroids. Reprod. Sci. 2020, 27, 1074–1085. [Google Scholar] [CrossRef]
- Islam, M.S.; Afrin, S.; Singh, B.; Jayes, F.L.; Brennan, J.T.; Borahay, M.A.; Leppert, P.C.; Segars, J.H. Extracellular matrix and Hippo signaling as therapeutic targets of antifibrotic compounds for uterine fibroids. Clin. Transl. Med. 2021, 11, e475. [Google Scholar] [CrossRef]
- Bozzini, N.; Rodrigues, C.J.; Petti, D.A.; Bevilacqua, R.G.; Goncalves, S.P.; Pinotti, J.A. Effects of treatment with gonadotropin releasing hormone agonist on the uterine leiomyomata structure. Acta. Obstet. Gynecol. Scand. 2003, 82, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Schlaff, W.D.; Ackerman, R.T.; Al-Hendy, A.; Archer, D.F.; Barnhart, K.T.; Bradley, L.D.; Carr, B.R.; Feinberg, E.C.; Hurtado, S.M.; Kim, J.; et al. Elagolix for Heavy Menstrual Bleeding in Women with Uterine Fibroids. N. Engl. J. Med. 2020, 382, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Al-Hendy, A.; Lukes, A.S.; Poindexter, A.N., 3rd; Venturella, R.; Villarroel, C.; Critchley, H.O.D.; Li, Y.; McKain, L.; Arjona Ferreira, J.C.; Langenberg, A.G.M.; et al. Treatment of Uterine Fibroid Symptoms with Relugolix Combination Therapy. N. Engl. J. Med. 2021, 384, 630–642. [Google Scholar] [CrossRef]
- Donnez, J.; Taylor, H.S.; Stewart, E.A.; Bradley, L.; Marsh, E.; Archer, D.; Al-Hendy, A.; Petraglia, F.; Watts, N.; Gotteland, J.P.; et al. Linzagolix with and without hormonal add-back therapy for the treatment of symptomatic uterine fibroids: Two randomised, placebo-controlled, phase 3 trials. Lancet 2022, 400, 896–907. [Google Scholar] [CrossRef] [PubMed]
- Wright, D.; Britten, J.; Malik, M.; Catherino, W.H. Relugolix and elagolix directly inhibit leiomyoma extracellular matrix production in 2-dimesnional and 3-dimensional cell cultures. FS Sci. 2022, 3, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Jayes, F.L.; Liu, B.; Moutos, F.T.; Kuchibhatla, M.; Guilak, F.; Leppert, P.C. Loss of stiffness in collagen-rich uterine fibroids after digestion with purified collagenase Clostridium histolyticum. Am. J. Obstet. Gynecol. 2016, 215, 596.e1–596.e8. [Google Scholar] [CrossRef]
- Ciebiera, M.; Wlodarczyk, M.; Ciebiera, M.; Zareba, K.; Lukaszuk, K.; Jakiel, G. Vitamin D and Uterine Fibroids-Review of the Literature and Novel Concepts. Int. J. Mol. Sci. 2018, 19, 2051. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, R.; Tabrizi, R.; Hessami, K.; Ashari, H.; Nowrouzi-Sohrabi, P.; Hosseini-Bensenjan, M.; Asadi, N. Correlation of low serum vitamin-D with uterine leiomyoma: A systematic review and meta-analysis. Reprod. Biol. Endocrinol. 2020, 18, 85. [Google Scholar] [CrossRef]
- Halder, S.K.; Osteen, K.G.; Al-Hendy, A. 1,25-dihydroxyvitamin d3 reduces extracellular matrix-associated protein expression in human uterine fibroid cells. Biol. Reprod. 2013, 89, 150. [Google Scholar] [CrossRef]
- Brakta, S.; Diamond, J.S.; Al-Hendy, A.; Diamond, M.P.; Halder, S.K. Role of vitamin D in uterine fibroid biology. Fertil. Steril. 2015, 104, 698–706. [Google Scholar] [CrossRef] [Green Version]
- Wawruszak, A.; Gumbarewicz, E.; Okon, E.; Jeleniewicz, W.; Czapinski, J.; Halasa, M.; Okla, K.; Smok-Kalwat, J.; Bocian, A.; Rivero-Muller, A.; et al. Histone deacetylase inhibitors reinforce the phenotypical markers of breast epithelial or mesenchymal cancer cells but inhibit their migratory properties. Cancer. Manag. Res. 2019, 11, 8345–8358. [Google Scholar] [CrossRef] [Green Version]
- Eckschlager, T.; Plch, J.; Stiborova, M.; Hrabeta, J. Histone Deacetylase Inhibitors as Anticancer Drugs. Int. J. Mol. Sci. 2017, 18, 1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Falahati, A.; Khosh, A.; Mohammed, H.; Kang, W.; Corachan, A.; Bariani, M.V.; Boyer, T.G.; Al-Hendy, A. Targeting Class I Histone Deacetylases in Human Uterine Leiomyosarcoma. Cells 2022, 11, 3801. [Google Scholar] [CrossRef]
- Carbajo-Garcia, M.C.; Garcia-Alcazar, Z.; Corachan, A.; Monleon, J.; Trelis, A.; Faus, A.; Pellicer, A.; Ferrero, H. Histone deacetylase inhibition by suberoylanilide hydroxamic acid: A therapeutic approach to treat human uterine leiomyoma. Fertil. Steril. 2022, 117, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Venturelli, S.; Berger, A.; Bocker, A.; Busch, C.; Weiland, T.; Noor, S.; Leischner, C.; Schleicher, S.; Mayer, M.; Weiss, T.S.; et al. Resveratrol as a pan-HDAC inhibitor alters the acetylation status of histone [corrected] proteins in human-derived hepatoblastoma cells. PLoS ONE 2013, 8, e73097. [Google Scholar] [CrossRef]
- Chen, H.Y.; Lin, P.H.; Shih, Y.H.; Wang, K.L.; Hong, Y.H.; Shieh, T.M.; Huang, T.C.; Hsia, S.M. Natural Antioxidant Resveratrol Suppresses Uterine Fibroid Cell Growth and Extracellular Matrix Formation In Vitro and In Vivo. Antioxidants 2019, 8, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujisawa, T.; Filippakopoulos, P. Functions of bromodomain-containing proteins and their roles in homeostasis and cancer. Nat. Rev. Mol. Cell Biol. 2017, 18, 246–262. [Google Scholar] [CrossRef]
- Kulikowski, E.; Rakai, B.D.; Wong, N.C.W. Inhibitors of bromodomain and extra-terminal proteins for treating multiple human diseases. Med. Res. Rev. 2021, 41, 223–245. [Google Scholar] [CrossRef]
- Cheung, K.L.; Kim, C.; Zhou, M.M. The Functions of BET Proteins in Gene Transcription of Biology and Diseases. Front. Mol. Biosci. 2021, 8, 728777. [Google Scholar] [CrossRef]
- Lu, L.; Chen, Z.; Lin, X.; Tian, L.; Su, Q.; An, P.; Li, W.; Wu, Y.; Du, J.; Shan, H.; et al. Inhibition of BRD4 suppresses the malignancy of breast cancer cells via regulation of Snail. Cell Death Differ. 2020, 27, 255–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Ali, M.; He, C.; Boyer, T.G.; Al-Hendy, A. Inhibition of BRD proteins suppresses the phenotype of uterine fibroids via regulation of N6-methyladenosine regulators. Fertil. Steril. 2021, 116, e11. [Google Scholar] [CrossRef]
- Yang, Q.; Mas, A.; Diamond, M.P.; Al-Hendy, A. The Mechanism and Function of Epigenetics in Uterine Leiomyoma Development. Reprod. Sci. 2016, 23, 163–175. [Google Scholar] [CrossRef] [Green Version]
- Navarro, A.; Yin, P.; Ono, M.; Monsivais, D.; Moravek, M.B.; Coon, J.S.t.; Dyson, M.T.; Wei, J.J.; Bulun, S.E. 5-Hydroxymethylcytosine promotes proliferation of human uterine leiomyoma: A biological link to a new epigenetic modification in benign tumors. J. Clin. Endocrinol. Metab. 2014, 99, E2437–E2445. [Google Scholar] [CrossRef]
- Navarro, A.; Yin, P.; Monsivais, D.; Lin, S.M.; Du, P.; Wei, J.J.; Bulun, S.E. Genome-wide DNA methylation indicates silencing of tumor suppressor genes in uterine leiomyoma. PLoS ONE 2012, 7, e33284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbajo-Garcia, M.C.; Corachan, A.; Segura-Benitez, M.; Monleon, J.; Escrig, J.; Faus, A.; Pellicer, A.; Cervello, I.; Ferrero, H. 5-aza-2′-deoxycitidine inhibits cell proliferation, extracellular matrix formation and Wnt/beta-catenin pathway in human uterine leiomyomas. Reprod. Biol. Endocrinol. 2021, 19, 106. [Google Scholar] [CrossRef]
- Chuang, T.D.; Panda, H.; Luo, X.; Chegini, N. miR-200c is aberrantly expressed in leiomyomas in an ethnic-dependent manner and targets ZEBs, VEGFA, TIMP2, and FBLN5. Endocr. Relat. Cancer 2012, 19, 541–556. [Google Scholar] [CrossRef] [Green Version]
- Chuang, T.D.; Luo, X.; Panda, H.; Chegini, N. miR-93/106b and their host gene, MCM7, are differentially expressed in leiomyomas and functionally target F3 and IL-8. Mol. Endocrinol. 2012, 26, 1028–1042. [Google Scholar] [CrossRef]
- Chuang, T.D.; Rehan, A.; Khorram, O. Tranilast induces MiR-200c expression through blockade of RelA/p65 activity in leiomyoma smooth muscle cells. Fertil. Steril. 2020, 113, 1308–1318. [Google Scholar] [CrossRef]
- Ciebiera, M.; Wlodarczyk, M.; Zgliczynski, S.; Lozinski, T.; Walczak, K.; Czekierdowski, A. The Role of miRNA and Related Pathways in Pathophysiology of Uterine Fibroids-From Bench to Bedside. Int. J. Mol. Sci. 2020, 21, 3016. [Google Scholar] [CrossRef] [Green Version]
- Marsh, E.E.; Steinberg, M.L.; Parker, J.B.; Wu, J.; Chakravarti, D.; Bulun, S.E. Decreased expression of microRNA-29 family in leiomyoma contributes to increased major fibrillar collagen production. Fertil. Steril. 2016, 106, 766–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiang, W.; Liu, Z.; Serna, V.A.; Druschitz, S.A.; Liu, Y.; Espona-Fiedler, M.; Wei, J.J.; Kurita, T. Down-regulation of miR-29b is essential for pathogenesis of uterine leiomyoma. Endocrinology 2014, 155, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Chuang, T.D.; Khorram, O. Mechanisms underlying aberrant expression of miR-29c in uterine leiomyoma. Fertil. Steril. 2016, 105, 236–245.e231. [Google Scholar] [CrossRef] [Green Version]
- Zota, A.R.; Geller, R.J.; VanNoy, B.N.; Marfori, C.Q.; Tabbara, S.; Hu, L.Y.; Baccarelli, A.A.; Moawad, G.N. Phthalate Exposures and MicroRNA Expression in Uterine Fibroids: The FORGE Study. Epigenet. Insights 2020, 13, 2516865720904057. [Google Scholar] [CrossRef] [Green Version]
- Ahn, S.H.; Kim, H.; Lee, I.; Lee, J.H.; Cho, S.; Choi, Y.S. MicroRNA-139-5p Regulates Fibrotic Potentials via Modulation of Collagen Type 1 and Phosphorylated p38 MAPK in Uterine Leiomyoma. Yonsei. Med. J. 2021, 62, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.L.; Zhao, L.; Han, S.C.; Bi, J.L.; Liu, H.X.; Yue, C.; Lin, L. MiR-129 is involved in the occurrence of uterine fibroid through inhibiting TET1. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 4419–4426. [Google Scholar] [CrossRef]
- Afrin, S.; Ali, M.; El Sabeh, M.; Yang, Q.; Al-Hendy, A.; Borahay, M.A. Simvastatin inhibits stem cell proliferation in human leiomyoma via TGF-beta3 and Wnt/beta-Catenin pathways. J. Cell Mol. Med. 2022, 26, 1684–1698. [Google Scholar] [CrossRef]
- Borahay, M.A.; Vincent, K.; Motamedi, M.; Sbrana, E.; Kilic, G.S.; Al-Hendy, A.; Boehning, D. Novel effects of simvastatin on uterine fibroid tumors: In vitro and patient-derived xenograft mouse model study. Am. J. Obstet. Gynecol. 2015, 213, 196.e1–196.e8. [Google Scholar] [CrossRef] [Green Version]
- Malik, M.; Britten, J.; Borahay, M.; Segars, J.; Catherino, W.H. Simvastatin, at clinically relevant concentrations, affects human uterine leiomyoma growth and extracellular matrix production. Fertil. Steril. 2018, 110, 1398–1407.e1. [Google Scholar] [CrossRef]
- Afrin, S.; Islam, M.S.; Patzkowsky, K.; Malik, M.; Catherino, W.H.; Segars, J.H.; Borahay, M.A. Simvastatin ameliorates altered mechanotransduction in uterine leiomyoma cells. Am. J. Obstet. Gynecol. 2020, 223, 733.e1–733.e14. [Google Scholar] [CrossRef]
- Galliano, D. Ulipristal acetate in uterine fibroids. Fertil. Steril. 2015, 103, 359–360. [Google Scholar] [CrossRef]
- Talaulikar, V.S.; Manyonda, I.T. Ulipristal acetate: A novel option for the medical management of symptomatic uterine fibroids. Adv. Ther. 2012, 29, 655–663. [Google Scholar] [CrossRef]
- Talaulikar, V.S.; Manyonda, I. Ulipristal acetate for use in moderate to severe symptoms of uterine fibroids. Womens Health 2014, 10, 565–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, J.; Malik, M.; Britten, J.; Lewis, T.; Catherino, W.H. Ulipristal Acetate and Extracellular Matrix Production in Human Leiomyomas In Vivo: A Laboratory Analysis of a Randomized Placebo Controlled Trial. Reprod. Sci. 2018, 25, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Chiang, Y.F.; Chen, H.Y.; Ali, M.; Shieh, T.M.; Huang, Y.J.; Wang, K.L.; Chang, H.Y.; Huang, T.C.; Hong, Y.H.; Hsia, S.M. The Role of Cell Proliferation and Extracellular Matrix Accumulation Induced by Food Additive Butylated Hydroxytoluene in Uterine Leiomyoma. Nutrients 2021, 13, 3074. [Google Scholar] [CrossRef] [PubMed]
- Reschke, L.; Afrin, S.; El Sabah, M.; Charewycz, N.; Miyashita-Ishiwata, M.; Borahay, M.A. Leptin induces leiomyoma cell proliferation and extracellular matrix deposition via JAK2/STAT3 and MAPK/ERK pathways. FS Sci. 2022, 3, 383–391. [Google Scholar] [CrossRef]
- Liu, J.; Yu, L.; Castro, L.; Yan, Y.; Clayton, N.P.; Bushel, P.; Flagler, N.D.; Scappini, E.; Dixon, D. Short-term tetrabromobisphenol A exposure promotes fibrosis of human uterine fibroid cells in a 3D culture system through TGF-beta signaling. FASEB J. 2022, 36, e22101. [Google Scholar] [CrossRef]
- Yan, Y.; Liu, J.; Lawrence, A.; Dykstra, M.J.; Fannin, R.; Gerrish, K.; Tucker, C.J.; Scappini, E.; Dixon, D. Prolonged cadmium exposure alters benign uterine fibroid cell behavior, extracellular matrix components, and TGFB signaling. FASEB J. 2021, 35, e21738. [Google Scholar] [CrossRef]
- Yu, C.H.; Zhao, J.S.; Zhao, H.; Peng, T.; Shen, D.C.; Xu, Q.X.; Li, Y.; Webb, R.C.; Wang, M.H.; Shi, X.M.; et al. Transcriptional profiling of uterine leiomyoma rats treated by a traditional herb pair, Curcumae rhizoma and Sparganii rhizoma. Braz. J. Med. Biol. Res. 2019, 52, e8132. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.Y.; Huang, T.C.; Lin, L.C.; Shieh, T.M.; Wu, C.H.; Wang, K.L.; Hong, Y.H.; Hsia, S.M. Fucoidan Inhibits the Proliferation of Leiomyoma Cells and Decreases Extracellular Matrix-Associated Protein Expression. Cell Physiol. Biochem. 2018, 49, 1970–1986. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.H.; Kung, H.L.; Chen, H.Y.; Huang, K.C.; Hsia, S.M. Isoliquiritigenin Suppresses E2-Induced Uterine Leiomyoma Growth through the Modulation of Cell Death Program and the Repression of ECM Accumulation. Cancers 2019, 11, 1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardozo, E.R.; Foster, R.; Karmon, A.E.; Lee, A.E.; Gatune, L.W.; Rueda, B.R.; Styer, A.K. MicroRNA 21a-5p overexpression impacts mediators of extracellular matrix formation in uterine leiomyoma. Reprod. Biol. Endocrinol. 2018, 16, 46. [Google Scholar] [CrossRef] [PubMed]
- Bernacchioni, C.; Ciarmela, P.; Vannuzzi, V.; Greco, S.; Vannuccini, S.; Malentacchi, F.; Pellegrino, P.; Capezzuoli, T.; Sorbi, F.; Cencetti, F.; et al. Sphingosine 1-phosphate signaling in uterine fibroids: Implication in activin A pro-fibrotic effect. Fertil. Steril. 2021, 115, 1576–1585. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Sims, H.; Trueheart, I.; Simpson, K.; Wang, K.C.; Patzkowsky, K.; Wegman, T.; Soma, J.M.; Dixon, R.; Jayes, F.; et al. A Phase I Clinical Trial to Assess Safety and Tolerability of Injectable Collagenase in Women with Symptomatic Uterine Fibroids. Reprod. Sci. 2021, 28, 2699–2709. [Google Scholar] [CrossRef] [PubMed]
- Corachan, A.; Ferrero, H.; Escrig, J.; Monleon, J.; Faus, A.; Cervello, I.; Pellicer, A. Long-term vitamin D treatment decreases human uterine leiomyoma size in a xenograft animal model. Fertil. Steril. 2020, 113, 205–216.e4. [Google Scholar] [CrossRef]
Figure 1.
ECM production and inhibition in uterine fibroids. ECM inducers, including hormones, cytokines, and growth factors, trigger the deposition and accumulation of ECM, activating downstream pathways and creating a pathological loop to promote the pathogenesis of UFs. Conversely, targeted inhibition of ECM inducers and relevant pathways decreases the production of ECM, concomitantly with the changes in mechanotransduction and relevant signaling, which contribute to the development and progression of UFs. Black dotted arrow: need further investigation; This figure was created using BioRender software. PR: progesterone receptor.
Figure 1.
ECM production and inhibition in uterine fibroids. ECM inducers, including hormones, cytokines, and growth factors, trigger the deposition and accumulation of ECM, activating downstream pathways and creating a pathological loop to promote the pathogenesis of UFs. Conversely, targeted inhibition of ECM inducers and relevant pathways decreases the production of ECM, concomitantly with the changes in mechanotransduction and relevant signaling, which contribute to the development and progression of UFs. Black dotted arrow: need further investigation; This figure was created using BioRender software. PR: progesterone receptor.

Figure 2.
Epigenetic-mediated targeting on ECM production. Targeted inhibition of HDACs, BRDs, and DNMTs and overexpression of targeted microRNAs can decrease the ECM deposition and accumulation in UF cells, leading to inhibited UF growth. Red up-arrow: overexpression of the microRNAs, which lead to the inhibition of ECM production; black ┴: inhibition; HDACs: histone deacetylases; BRD9: bromodomain-containing protein 9; DNMTs: DNA methyltransferases; ECM: extracellular matrix; TFs: transcription factors; SAHA: suberoylanilide hydroxamic acid (HDAC inhibitor); I-BRD9: BRD9 inhibitor; 5-Aza: 5-Aza-2-deoxycytidine (DNMT inhibitor). This figure was created using BioRender software.
Figure 2.
Epigenetic-mediated targeting on ECM production. Targeted inhibition of HDACs, BRDs, and DNMTs and overexpression of targeted microRNAs can decrease the ECM deposition and accumulation in UF cells, leading to inhibited UF growth. Red up-arrow: overexpression of the microRNAs, which lead to the inhibition of ECM production; black ┴: inhibition; HDACs: histone deacetylases; BRD9: bromodomain-containing protein 9; DNMTs: DNA methyltransferases; ECM: extracellular matrix; TFs: transcription factors; SAHA: suberoylanilide hydroxamic acid (HDAC inhibitor); I-BRD9: BRD9 inhibitor; 5-Aza: 5-Aza-2-deoxycytidine (DNMT inhibitor). This figure was created using BioRender software.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yang, Q.; Al-Hendy, A. Update on the Role and Regulatory Mechanism of Extracellular Matrix in the Pathogenesis of Uterine Fibroids. Int. J. Mol. Sci. 2023, 24, 5778. https://doi.org/10.3390/ijms24065778
AMA Style
Yang Q, Al-Hendy A. Update on the Role and Regulatory Mechanism of Extracellular Matrix in the Pathogenesis of Uterine Fibroids. International Journal of Molecular Sciences. 2023; 24(6):5778. https://doi.org/10.3390/ijms24065778
Chicago/Turabian StyleYang, Qiwei, and Ayman Al-Hendy. 2023. "Update on the Role and Regulatory Mechanism of Extracellular Matrix in the Pathogenesis of Uterine Fibroids" International Journal of Molecular Sciences 24, no. 6: 5778. https://doi.org/10.3390/ijms24065778
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.