

VCAN Hypomethylation and Expression as Predictive Biomarkers of Drug Sensitivity in Upper Urinary Tract Urothelial Carcinoma
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
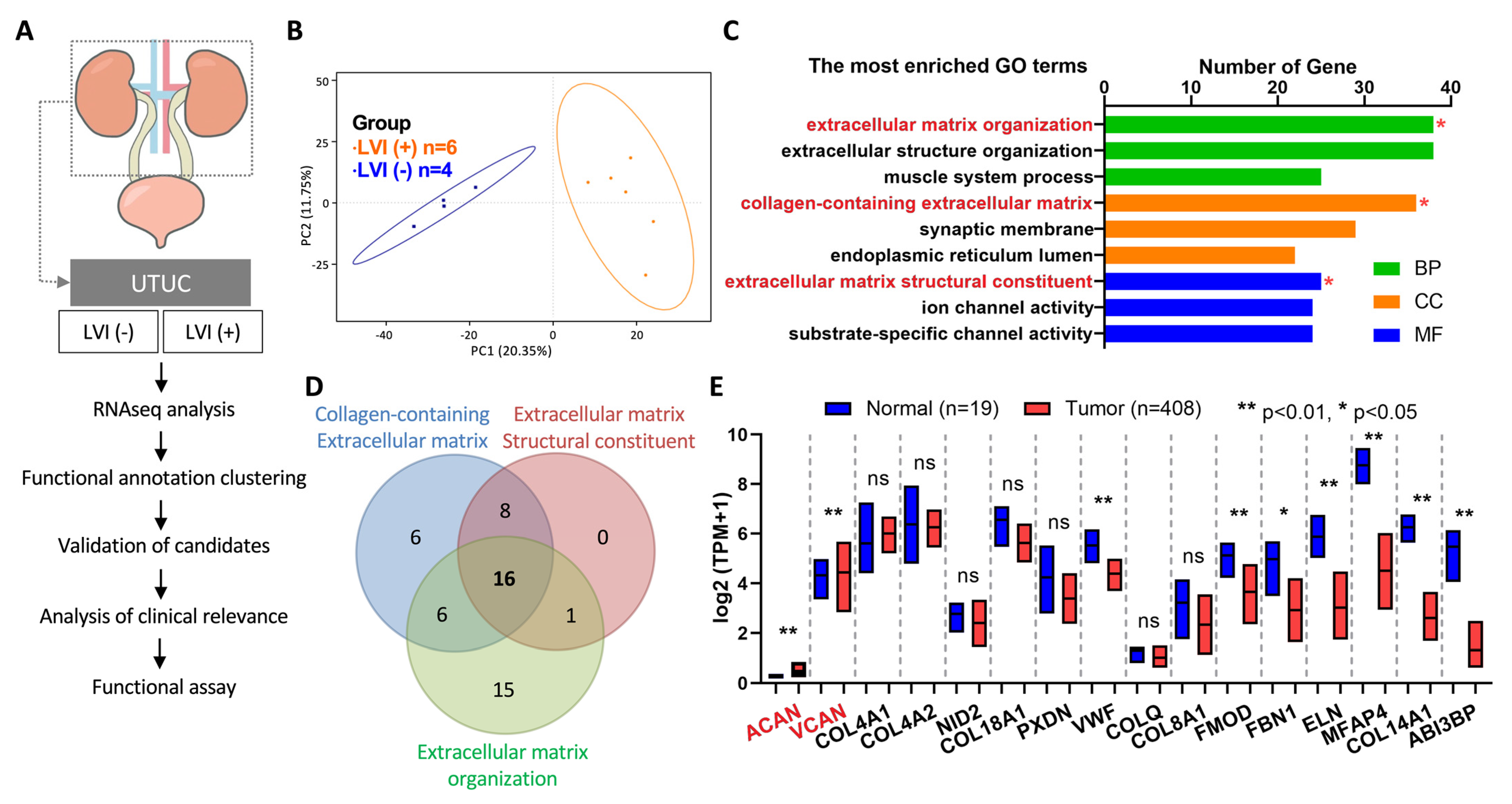
2.1. RNAseq Analysis Identified VCAN as an Oncogene in LVI-Positive UTUC
2.2. Association of VCAN Expression with Progression of BLCA in TCGA Database
2.3. VCAN Gene Locus in Chromosome 5: 83.471,618-83,582,303 Was Hypermethylated in UTUC Patients
2.4. High Expression of VCAN in LVI-Positive Tumors from UTUC Patients
2.5. VCAN Promoted the Migration of UTUC Cancer Cells
2.6. Correlation Analysis of VCAN-Associated Gene Expression with Molecular Subtypes in UTUC Tumors
2.7. VCAN Enhanced Chemoresistance of Cancer Cells
3. Discussion
4. Materials and Methods
4.1. Tissue Samples
4.2. Immunohistochemistry
4.3. Cell Lines, shRNA, RNA Isolation and Quantitative Real-Time PCR
4.4. Western Blotting Analysis
4.5. Whole Transcriptome RNAseq
4.6. Methylation-Specific PCR
4.7. RTCA Cell Proliferation Assay
4.8. RTCA Cell Migration Assay
4.9. Drug Sensitivity Assay
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Margulis, V.; Shariat, S.F.; Matin, S.F.; Kamat, A.M.; Zigeuner, R.; Kikuchi, E.; Lotan, Y.; Weizer, A.; Raman, J.D.; Wood, C.G. Outcomes of radical nephroureterectomy: A series from the Upper Tract Urothelial Carcinoma Collaboration. Cancer 2009, 115, 1224–1233. [Google Scholar] [CrossRef]
- Kang, M.; Jeong, C.W.; Kwak, C.; Kim, H.H.; Ku, J.H. The Characteristics of Recurrent Upper Tract Urothelial Carcinoma after Radical Nephroureterectomy without Bladder Cuff Excision. Yonsei Med. J. 2015, 56, 375. [Google Scholar] [CrossRef]
- Margulis, V.; Puligandla, M.; Trabulsi, E.J.; Plimack, E.R.; Kessler, E.R.; Matin, S.F.; Godoy, G.; Alva, A.; Hahn, N.M.; Carducci, M.A.; et al. Phase II Trial of Neoadjuvant Systemic Chemotherapy Followed by Extirpative Surgery in Patients with High Grade Upper Tract Urothelial Carcinoma. J. Urol. 2020, 203, 690–698. [Google Scholar] [CrossRef] [PubMed]
- Birtle, A.; Johnson, M.H.; Chester, J.D.; Jones, R.; Dolling, D.S.; Bryan, R.T.; Harris, C.; Winterbottom, A.; Blacker, A.; Catto, J.W.; et al. Adjuvant chemotherapy in upper tract urothelial carcinoma (the POUT trial): A phase 3, open-label, randomised controlled trial. Lancet 2020, 395, 1268–1277. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, P.H.; Stadler, W.M. The Role of Chemotherapy in Upper Tract Urothelial Carcinoma. Adv. Urol. 2009, 2009, 419028. [Google Scholar] [CrossRef] [PubMed]
- Rouprêt, M.; Babjuk, M.; Burger, M.; Čapoun, O.; Cohen, D.; Comperat, E.; Cowan, N.C.; Domínguez-Escrig, J.; Gontero, P.; Mostafid, H.; et al. European Association of Urology Guidelines on Upper Urinary Tract Urothelial Carcinoma: 2020 Update. Eur. Urol. 2021, 79, 62–79. [Google Scholar] [CrossRef] [PubMed]
- Tabayoyong, W.; Li, R.; Gao, J.; Kamat, A. Optimal Timing of Chemotherapy and Surgery in Patients with Muscle-Invasive Bladder Cancer and Upper Urinary Tract Urothelial Carcinoma. Urol. Clin. N. Am. 2018, 45, 155–167. [Google Scholar] [CrossRef]
- Xylinas, E.; Rink, M.; Margulis, V.; Clozel, T.; Lee, R.K.; Comploj, E.; Novara, G.; Raman, J.D.; Lotan, Y.; Weizer, A.; et al. Impact of renal function on eligibility for chemotherapy and survival in patients who have undergone radical nephro-ureterectomy. BJU Int. 2013, 112, 453–461. [Google Scholar] [CrossRef]
- Bajorin, D.F.; Witjes, J.A.; Gschwend, J.E.; Schenker, M.; Valderrama, B.P.; Tomita, Y.; Bamias, A.; Lebret, T.; Shariat, S.F.; Park, S.H.; et al. Adjuvant Nivolumab versus Placebo in Muscle-Invasive Urothelial Carcinoma. N. Engl. J. Med. 2021, 384, 2102–2114. [Google Scholar] [CrossRef]
- Luo, H.L.; Chen, Y.T.; Chuang, Y.C.; Cheng, Y.T.; Lee, W.C.; Kang, C.H.; Chiang, P.H. Subclassification of upper urinary tract urothelial carcinoma by the neutrophil-to-lymphocyte ratio (NLR) improves prediction of oncological outcome. BJU Int. 2014, 113, E144–E149. [Google Scholar] [CrossRef]
- Xiong, G.; Chen, X.; Li, X.; Fang, D.; Zhang, L.; Yang, L.; Zhang, L.; Yao, L.; He, Z.; Zhou, L. Prevalence and factors associated with baseline chronic kidney disease in China: A 10-year study of 785 upper urinary tract urothelial carcinoma patients. J. Formos. Med. Assoc. 2014, 113, 521–526. [Google Scholar] [CrossRef]
- Luo, H.L.; Chiang, P.H.; Chen, Y.T.; Cheng, Y.T. Lymphovascular invasion is a pathological feature related to aggressive cancer behavior and predicts early recurrence in prostate cancer. Kaohsiung J. Med. Sci. 2012, 28, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Luo, K.; Chen, C.; Wang, G.; Jin, J.; Kong, M.; Li, B.; Liu, Q.; Li, J.; Rong, T.; et al. Identification and Validation of Lymphovascular Invasion as a Prognostic and Staging Factor in Node-Negative Esophageal Squamous Cell Carcinoma. J. Thorac. Oncol. 2016, 11, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.J.; Shin, S.H.; Cho, J.S.; Park, M.H.; Yoon, J.H.; Jegal, Y.J. The Role of Lymphovascular Invasion as a Prognostic Factor in Patients with Lymph Node-Positive Operable Invasive Breast Cancer. J. Breast Cancer 2011, 14, 198. [Google Scholar] [CrossRef]
- Skancke, M.; Arnott, S.M.; Amdur, R.L.; Siegel, R.S.; Obias, V.J.; Umapathi, B.A. Lymphovascular Invasion and Perineural Invasion Negatively Impact Overall Survival for Stage II Adenocarcinoma of the Colon. Dis. Colon Rectum 2019, 62, 181–188. [Google Scholar] [CrossRef]
- Hurel, S.; Rouprêt, M.; Ouzzane, A.; Rozet, F.; Xylinas, E.; Zerbib, M.; Berod, A.A.; Ruffion, A.; Adam, E.; Cussenot, O.; et al. Impact of lymphovascular invasion on oncological outcomes in patients with upper tract urothelial carcinoma after radical nephroureterectomy. BJU Int. 2013, 111, 1199–1207. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.K.; Lee, J.Y.; Kim, J.W.; Hah, Y.S.; Cho, K.S. Effect of neoadjuvant chemotherapy on locally advanced upper tract urothelial carcinoma: A systematic review and meta-analysis. Crit. Rev. Oncol./Hematol. 2019, 135, 59–65. [Google Scholar] [CrossRef]
- Song, S.H.; Ye, C.H.; Lee, S.; Hong, S.K.; Byun, S.S.; Lee, S.E.; Oh, J.J. Association between lymphovascular invasion and oncologic outcomes among upper urinary tract urothelial carcinoma patients who underwent radical nephroureterectomy. J. Cancer Res. Clin. Oncol. 2019, 145, 2863–2870. [Google Scholar] [CrossRef] [PubMed]
- Danzig, M.R.; Mallin, K.; McKiernan, J.M.; Stadler, W.M.; Sridhar, S.S.; Morgan, T.M.; Bochner, B.H.; Lee, C.T. Prognostic importance of lymphovascular invasion in urothelial carcinoma of the renal pelvis. Cancer 2018, 124, 2507–2514. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wu, B.; Zha, Z.; Zhao, H.; Yuan, J.; Feng, Y. The Prognostic Value of Lymphovascular Invasion in Patients with Upper Tract Urinary Carcinoma After Surgery: An Updated Systematic Review and Meta-Analysis. Front. Oncol. 2020, 10, 487. [Google Scholar] [CrossRef]
- Lee, H.Y.; Li, C.C.; Huang, C.N.; Ke, H.L.; Li, W.M.; Liang, P.I.; Yang, S.F.; Tu, H.P.; Wu, W.J.; Yeh, H.C. Prognostic Significance of Lymphovascular Invasion in Upper Urinary Tract Urothelial Carcinoma is Influenced by Tumor Location. Ann. Surg. Oncol. 2014, 22, 1392–1400. [Google Scholar] [CrossRef]
- Papadas, A.; Arauz, G.; Cicala, A.; Wiesner, J.; Asimakopoulos, F. Versican and Versican-matrikines in Cancer Progression, Inflammation, and Immunity. J. Histochem. Cytochem. 2020, 68, 871–885. [Google Scholar] [CrossRef] [PubMed]
- Said, N.; Sanchez-Carbayo, M.; Smith, S.C.; Theodorescu, D. RhoGDI2 suppresses lung metastasis in mice by reducing tumor versican expression and macrophage infiltration. J. Clin. Investig. 2012, 122, 1503–1518. [Google Scholar] [CrossRef] [PubMed]
- Touab, M.; Villena, J.; Barranco, C.; Arumí-Uría, M.; Bassols, A. Versican Is Differentially Expressed in Human Melanoma and May Play a Role in Tumor Development. Am. J. Pathol. 2002, 160, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Yeung, T.L.; Leung, C.S.; Wong, K.K.; Samimi, G.; Thompson, M.S.; Liu, J.; Zaid, T.M.; Ghosh, S.; Birrer, M.J.; Mok, S.C. TGF-β Modulates Ovarian Cancer Invasion by Upregulating CAF-Derived Versican in the Tumor Microenvironment. Cancer Res. 2013, 73, 5016–5028. [Google Scholar] [CrossRef]
- Kim, S.; Takahashi, H.; Lin, W.W.; Descargues, P.; Grivennikov, S.; Kim, Y.; Luo, J.L.; Karin, M. Carcinoma-produced factors activate myeloid cells through TLR2 to stimulate metastasis. Nature 2009, 457, 102–106. [Google Scholar] [CrossRef]
- Suhovskih, A.V.; Aidagulova, S.V.; Kashuba, V.I.; Grigorieva, E.V. Proteoglycans as potential microenvironmental biomarkers for colon cancer. Cell Tissue Res. 2015, 361, 833–844. [Google Scholar] [CrossRef]
- Keire, P.A.; Kang, I.; Wight, T.N. Versican: Role in cancer tumorigenesis. In Extracellular Matrix in Tumor Biology; Springer: Cham, Switzerland, 2017; pp. 51–74. [Google Scholar] [CrossRef]
- Ball, S.J.; Mann, D.A.; Wilson, J.A.; Fisher, A.J. The Role of the Fibroblast in Inflammatory Upper Airway Conditions. Am. J. Pathol. 2016, 186, 225–233. [Google Scholar] [CrossRef]
- Kotton, D.N.; Morrisey, E.E. Lung regeneration: Mechanisms, applications and emerging stem cell populations. Nat. Med. 2014, 20, 822–832. [Google Scholar] [CrossRef]
- Chang, M.Y.; Tanino, Y.; Vidova, V.; Kinsella, M.G.; Chan, C.K.; Johnson, P.Y.; Wight, T.N.; Frevert, C.W. A rapid increase in macrophage-derived versican and hyaluronan in infectious lung disease. Matrix Biol. 2014, 34, 1–12. [Google Scholar] [CrossRef]
- Petrey, A.C.; De La Motte, C.A. Hyaluronan, a Crucial Regulator of Inflammation. Front. Immunol. 2014, 5, 101. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.; Watanabe, H. Versican: A Dynamic Regulator of the Extracellular Matrix. J. Histochem. Cytochem. 2020, 68, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Rainiero, H.R.; Emmerich, P.B.; Sievers, C.K.; Maloney, C.J.; Pitera, R.T.; Payne, S.; Depke, M.; Pasch, C.A.; Clipson, L.; Johnson, J.A.; et al. Abstract 1904: Versican production is driven by both epithelial and stromal cells in pancreatic cancer. Tumor Biol. 2019, 79, 1904. [Google Scholar] [CrossRef]
- Gupta, N.; Kumar, R.; Seth, T.; Garg, B.; Sharma, A. Targeting of stromal versican by miR-144/199 inhibits multiple myeloma by downregulating FAK/STAT3 signalling. RNA Biol. 2020, 17, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wang, X.; Wu, J.L.; Quan, W.Q.; Ma, L.; Yang, F.; Wu, K.Y.; Wan, H.Y. Tumor-Produced Versican V1 Enhances hCAP18/LL-37 Expression in Macrophages through Activation of TLR2 and Vitamin D3 Signaling to Promote Ovarian Cancer Progression In Vitro. PLoS ONE 2013, 8, e56616. [Google Scholar] [CrossRef]
- Xia, L.; Huang, W.; Tian, D.; Zhang, L.; Qi, X.; Chen, Z.; Shang, X.; Nie, Y.; Wu, K. Forkhead box Q1 promotes hepatocellular carcinoma metastasis by transactivating ZEB2 and VersicanV1 expression. Hepatology 2014, 59, 958–973. [Google Scholar] [CrossRef] [PubMed]
- Bögels, M.; Braster, R.; Nijland, P.G.; Gül, N.; Van De Luijtgaarden, W.; Fijneman, R.J.; Meijer, G.A.; Jimenez, C.R.; Beelen, R.H.; Van Egmond, M. Carcinoma origin dictates differential skewing of monocyte function. OncoImmunology 2012, 1, 798–809. [Google Scholar] [CrossRef]
- Hu, F.; Dzaye, O.; Hahn, A.; Yu, Y.; Scavetta, R.J.; Dittmar, G.; Kaczmarek, A.K.; Dunning, K.R.; Ricciardelli, C.; Rinnenthal, J.L.; et al. Glioma-derived versican promotes tumor expansion via glioma-associated microglial/macrophages Toll-like receptor 2 signaling. Neuro-Oncology 2015, 17, 200–210. [Google Scholar] [CrossRef]
- Said, N.; Theodorescu, D. RhoGDI2 suppresses bladder cancer metastasis via reduction of inflammation in the tumor microenvironment. OncoImmunology 2012, 1, 1175–1177. [Google Scholar] [CrossRef]
- Evanko, S.P.; Angello, J.C.; Wight, T.N. Formation of Hyaluronan- and Versican-Rich Pericellular Matrix Is Required for Proliferation and Migration of Vascular Smooth Muscle Cells. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 1004–1013. [Google Scholar] [CrossRef]
- Evanko, S.P.; Johnson, P.Y.; Braun, K.R.; Underhill, C.B.; Dudhia, J.; Wight, T.N. Platelet-Derived Growth Factor Stimulates the Formation of Versican–Hyaluronan Aggregates and Pericellular Matrix Expansion in Arterial Smooth Muscle Cells. Arch. Biochem. Biophys. 2001, 394, 29–38. [Google Scholar] [CrossRef]
- Schönherr, E.; Kinsella, M.G.; Wight, T.N. Genistein Selectively Inhibits Platelet-Derived Growth Factor-Stimulated Versican Biosynthesis in Monkey Arterial Smooth Muscle Cells. Arch. Biochem. Biophys. 1997, 339, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wu, J.; Lee, D.Y.; Yee, A.; Cao, L.; Zhang, Y.; Kiani, C.; Yang, B.B. Versican protects cells from oxidative stress-induced apoptosis. Matrix Biol. 2005, 24, 3–13. [Google Scholar] [CrossRef]
- LaPierre, D.P.; Lee, D.Y.; Li, S.Z.; Xie, Y.Z.; Zhong, L.; Sheng, W.; Deng, Z.; Yang, B.B. The Ability of Versican to Simultaneously Cause Apoptotic Resistance and Sensitivity. Cancer Res. 2007, 67, 4742–4750. [Google Scholar] [CrossRef]
- Zimmermann, D.R.; Ruoslahti, E. Multiple domains of the large fibroblast proteoglycan, versican. EMBO J. 1989, 8, 2975–2981. [Google Scholar] [CrossRef]
- Locke, W.J.; Guanzon, D.; Ma, C.; Liew, Y.J.; Duesing, K.; Fung, K.Y.C.; Ross, J.W. DNA Methylation Cancer Biomarkers: Translation to the Clinic. Front. Genet. 2019, 10, 1150. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, A.; Nakanishi, M. Navigating the DNA methylation landscape of cancer. Trends Genet. 2021, 37, 1012–1027. [Google Scholar] [CrossRef]
- Gorter, A.; Zijlmans, H.; Van Gent, H.; Trimbos, J.B.; Fleuren, G.J.; Jordanova, E.S. Versican expression is associated with tumor-infiltrating CD8-positive T cells and infiltration depth in cervical cancer. Mod. Pathol. 2010, 23, 1605–1615. [Google Scholar] [CrossRef] [PubMed]
- Dos Reis, D.C.; Damasceno, K.A.; De Campos, C.B.; Veloso, E.S.; Pêgas, G.R.A.; Kraemer, L.R.; Rodrigues, M.A.; Mattos, M.S.; Gomes, D.A.; Campos, P.P.; et al. Versican and Tumor-Associated Macrophages Promotes Tumor Progression and Metastasis in Canine and Murine Models of Breast Carcinoma. Front. Oncol. 2019, 9, 577. [Google Scholar] [CrossRef]
- Gao, D.; Joshi, N.; Choi, H.; Ryu, S.; Hahn, M.; Catena, R.; Sadik, H.; Argani, P.; Wagner, P.; Vahdat, L.T.; et al. Myeloid Progenitor Cells in the Premetastatic Lung Promote Metastases by Inducing Mesenchymal to Epithelial Transition. Cancer Res. 2012, 72, 1384–1394. [Google Scholar] [CrossRef]
- Zheng, P.; Wen, J.; Ang, L.C.; Sheng, W.; Viloria-Petit, A.; Wang, Y.; Wu, Y.; Kerbel, R.S.; Yang, B.B. Versican/PG-M G3 domain promotes tumor growth and angiogenesis. FASEB J. 2004, 18, 754–756. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Yee, A.J. Versican V2 isoform enhances angiogenesis by regulating endothelial cell activities and fibronectin expression. FEBS Lett. 2012, 587, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Yee, A.J.; Akens, M.; Yang, B.L.; Finkelstein, J.; Zheng, P.S.; Deng, Z.; Yang, B. The effect of versican G3 domain on local breast cancer invasiveness and bony metastasis. Breast Cancer Res. 2007, 9, R47. [Google Scholar] [CrossRef] [PubMed]
- Tomiyama, E.; Fujita, K.; Hashimoto, M.; Adomi, S.; Kawashima, A.; Minami, T.; Yoshimura, K.; Uemura, H.; Nonomura, N. Comparison of molecular profiles of upper tract urothelial carcinoma vs. urinary bladder cancer in the era of targeted therapy: A narrative review. Transl. Androl. Urol. 2022, 11, 1747–1761. [Google Scholar] [CrossRef]
- Chung, Y.; Cheng, Y.; Kao, Y.; Tsai, W.; Huang, G.; Chen, Y.; Shen, Y.; Tai, M.; Chiang, P. MiR-26a-5p as a useful therapeutic target for upper tract urothelial carcinoma by regulating WNT5A/β-catenin signaling. Sci. Rep. 2022, 12, 6955. [Google Scholar] [CrossRef]
- Roshan Moniri, M.; Young, A.; Reinheimer, K.; Rayat, J.; Dai, L.-J.; Warnock, G.L. Dynamic Assessment of Cell Viability, Proliferation and Migration Using Real Time Cell Analyzer System (RTCA). Cytotechnology 2015, 67, 379–386. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, H.-L.; Chang, Y.-L.; Liu, H.-Y.; Wu, Y.-T.; Sung, M.-T.; Su, Y.-L.; Huang, C.-C.; Wang, P.-C.; Peng, J.-M. VCAN Hypomethylation and Expression as Predictive Biomarkers of Drug Sensitivity in Upper Urinary Tract Urothelial Carcinoma. Int. J. Mol. Sci. 2023, 24, 7486. https://doi.org/10.3390/ijms24087486
Luo H-L, Chang Y-L, Liu H-Y, Wu Y-T, Sung M-T, Su Y-L, Huang C-C, Wang P-C, Peng J-M. VCAN Hypomethylation and Expression as Predictive Biomarkers of Drug Sensitivity in Upper Urinary Tract Urothelial Carcinoma. International Journal of Molecular Sciences. 2023; 24(8):7486. https://doi.org/10.3390/ijms24087486
Chicago/Turabian StyleLuo, Hao-Lun, Yin-Lun Chang, Hui-Ying Liu, Yen-Ting Wu, Ming-Tse Sung, Yu-Li Su, Chun-Chieh Huang, Pei-Chia Wang, and Jei-Ming Peng. 2023. "VCAN Hypomethylation and Expression as Predictive Biomarkers of Drug Sensitivity in Upper Urinary Tract Urothelial Carcinoma" International Journal of Molecular Sciences 24, no. 8: 7486. https://doi.org/10.3390/ijms24087486