
The Localization of Cell Wall Components in the Quadrifids of Whole-Mount Immunolabeled Utricularia dichotoma Traps


1
Department of Plant Cytology and Embryology, Institute of Botany, Faculty of Biology, Jagiellonian University in Kraków, 9 Gronostajowa St., 30-387 Cracow, Poland
2
Laboratory of Bioimaging, Faculty of Biology, University of Gdańsk, 59 Wita Stwosza St., 80-308 Gdańsk, Poland
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2024, 25(1), 56; https://doi.org/10.3390/ijms25010056
Submission received: 22 November 2023
/
Revised: 13 December 2023
/
Accepted: 19 December 2023
/
Published: 19 December 2023
(This article belongs to the Special Issue From Cell to Organism: Cytological, Experimental Molecular and Medical Aspects)
Abstract
:Utricularia (bladderworts) are carnivorous plants. They produce small hollow vesicles, which function as suction traps that work underwater and capture fine organisms. Inside the traps, there are numerous glandular trichomes (quadrifids), which take part in the secretion of digestive enzymes, the resorption of released nutrients, and likely the pumping out of water. Due to the extreme specialization of quadrifids, they are an interesting model for studying the cell walls. This aim of the study was to fill in the gap in the literature concerning the immunocytochemistry of quadrifids in the major cell wall polysaccharides and glycoproteins. To do this, the localization of the cell wall components in the quadrifids was performed using whole-mount immunolabeled Utricularia traps. It was observed that only parts (arms) of the terminal cells had enough discontinuous cuticle to be permeable to antibodies. There were different patterns of the cell wall components in the arms of the terminal cells of the quadrifids. The cell walls of the arms were especially rich in low-methyl-esterified homogalacturonan. Moreover, various arabinogalactan proteins also occurred. Cell walls in glandular cells of quadrifids were rich in low-methyl-esterified homogalacturonan; in contrast, in the aquatic carnivorous plant Aldrovanda vesiculosa, cell walls in the glandular cells of digestive glands were poor in low-methyl-esterified homogalacturonan. Arabinogalactan proteins were found in the cell walls of trap gland cells in all studied carnivorous plants: Utricularia, and members of Droseraceae and Drosophyllaceae.
1. Introduction
The genus Utricularia is the most diverse genus among carnivorous plants, both in terms of plant size and environmental adaptations, as well as plant body architecture. It is also the richest in number of species, with around 250 species [1,2,3,4]. All of the Utricularia species are rootless herbs. Their vegetative organs go beyond the typical organ classification, i.e., either ‘root’, ‘stem’, or ‘leaf’, and they have intermixed morphological traits and developmental programs [5,6,7,8]. These are related to the heterotopic transfer of the function of the genes to other organs, as was shown in the root genes [9]. Utricularia produce small hollow vesicles (bladders with elastic walls and a mobile trap door), which function as suction traps that work underwater and capture fine organisms [10,11,12,13,14,15,16]. There is agreement that these traps are of a foliar origin. Using Utricularia gibba, Whitewoods et al. [17] showed that simple shifts in gene expression may change leaf morphogenesis, which finally results in trap formation. Utricularia traps attract special interest because they are among the fastest moving plant organs [15,18,19,20,21,22,23]. Poppinga et al. [23] showed that animals were successfully captured by Utricularia australis traps within 9 ms.
There are various types of glandular trichomes on the surface of the Utricularia traps [24,25,26,27,28,29]. The inner part of the trap is densely lined by two types of large trichomes. There are trichomes with four terminal cells called quadrifids, which almost cover the entire inner surface, and trichomes with two terminal cells called bifids, which are located near the trap door. Darwin proposed the names of these trichomes [30]. Both quadrifids and bifids have the same architecture, because are formed by a basal cell, a pedestal cell, and a terminal cell. The terminal cell is regionally differentiated and consists of different parts with distinct structures and functions: the basal part, stalk and arm. Quadrifids participate in the secretion of digestive enzymes and in the resorption of released nutrients, and probably also participate in pumping out water. According to some authors, the main role of bifids is pumping out water [26,27,31,32,33]. Due to the extreme specialization of the quadrifids, they are an interesting model to use to study cell walls. Recently, in a series of works, we showed the major cell wall polysaccharides and glycoproteins in the various glands of carnivorous plants; however, our studies were limited to Aldrovanda vesiculosa [34,35], Dionaea muscipula [36,37], and Drosophyllum lusitanicum [38]. This aim of this study is to fill in the gap in the literature concerning the immunocytochemistry of the quadrifids in the major cell wall polysaccharides and glycoproteins. Plant organs are covered by a cuticle, which is a hydrophobic and protective barrier, but is also impermeable to antibodies. However, structures such as the root hairs and pollen tubes are devoid of a cuticle, which means that the cell walls can be studied using whole-mount immunolabeled organs (without the time-consuming task of embedding them in resin and later cutting the material with a microtome). The use of this technique has produced very good results in root hairs [39,40,41] and pollen tubes [42,43]. Because cuticle discontinuities in the glands of carnivorous plants are known [26,27,44], we wanted to try this method; therefore, we used whole-mount immunolabeled Utricularia traps. However, to what extent a cuticle with discontinuities would be a barrier to the antibodies in these traps was an open question.
2. Results
2.1. Trap Morphology and Structure of a Quadrifid (Figure 1A–F)
A quadrifid has a typical structure and consists of a basal cell, pedestal cell (endodermoid) and four glandular cells. The glandular cells consist of a basal part, a stalk, and an arm (Figure 1C–F). The arm (the middle and distal regions) is highly vacuolated (Figure 1C,D). A large vacuole is surrounded by peripheral cytoplasm with numerous mitochondria. The cytoplasm of the arm is mostly concentrated towards its base, where the nucleus is also located (Figure 1D). Staining with Sudan III shows positive result of cutin occurrence in stalks of trichome terminal cells, the pedestal cell, and trap epidermis (Figure 1E). An autofluorescence of cutin occurs in stalks of trichome terminal cells and the pedestal cell (Figure 1F). Similar results were obtained using staining with Auramine O (Figure S1A).
Figure 1.
Distribution and morphology of the quadrifids of Utricularia dichotoma subsp. novae-zelandiae. (A) Morphology of a trap via SEM, scale bar 500 µm. (B) A sagittally halved trap showing the quadrifids (arrow), SEM, scale bar 500 µm. (C) Morphology of the quadrifids via light microscopy, scale bar 20 µm. (D) Staining with DiOC6 showed numerous mitochondria and nuclei (star) in terminal cells of a quadrifid, arm (A), via fluorescent microscopy, scale bar 10 µm. (E) Lateral view of quadrifid. Staining with Sudan (orange color); note the positive staining of cutin in stalks (white arrow) and cuticle of epidermis (black arrow); basal cell (B), pedestal cell (P), and arm (A) via light microscopy, scale bar 20 µm. (F) Lateral view of quadrifid; note the strong autofluorescence of cutin (blue–white) in stalks (white arrow) and the pedestal cell (P), but no autofluorescence of cutin in arms (A) via fluorescent microscopy, scale bar 20 µm.
Figure 1.
Distribution and morphology of the quadrifids of Utricularia dichotoma subsp. novae-zelandiae. (A) Morphology of a trap via SEM, scale bar 500 µm. (B) A sagittally halved trap showing the quadrifids (arrow), SEM, scale bar 500 µm. (C) Morphology of the quadrifids via light microscopy, scale bar 20 µm. (D) Staining with DiOC6 showed numerous mitochondria and nuclei (star) in terminal cells of a quadrifid, arm (A), via fluorescent microscopy, scale bar 10 µm. (E) Lateral view of quadrifid. Staining with Sudan (orange color); note the positive staining of cutin in stalks (white arrow) and cuticle of epidermis (black arrow); basal cell (B), pedestal cell (P), and arm (A) via light microscopy, scale bar 20 µm. (F) Lateral view of quadrifid; note the strong autofluorescence of cutin (blue–white) in stalks (white arrow) and the pedestal cell (P), but no autofluorescence of cutin in arms (A) via fluorescent microscopy, scale bar 20 µm.
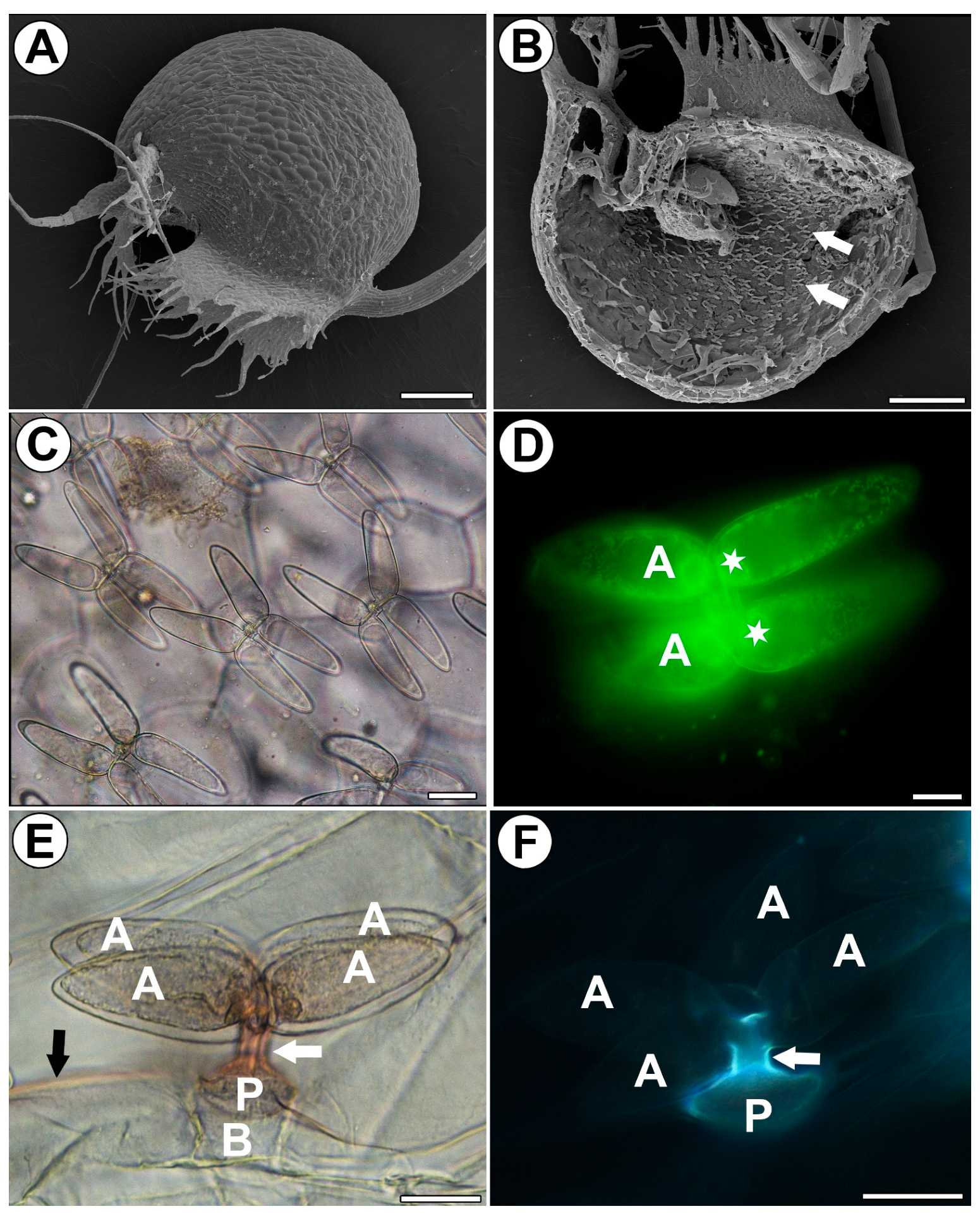
2.2. Distribution of the Arabinogalactan Proteins (AGPs)
We used the JIM8, JIM13 and JIM14 antibodies in order to localize the AGPs (Figure 2A–C). JIM8 reacted with fluorescence in the cell walls of arms of the quadrifids (Figure 2A), but only in the apical and middle parts of the arms. JIM13 gave a fluorescence signal of the AGP epitopes in the cell walls of the arms of the quadrifids (Figure 2B), but not in the basal part. The signal was intense in the middle parts of the arms, and less intense in the apical part of the arms. JIM14 reacted with fluorescence in the cell walls of the arms of the quadrifids (Figure 2C), but only in the apical and middle parts of the arms.
2.3. Distribution of Homogalacturonan
Low-methyl-esterified homogalacturonans (HGs) were detected by the JIM5 and LM19 antibodies (Figure 3A–C). Both JIM5 and LM 19 reacted with a strong fluorescence in the cell walls of the arms of the quadrifids (Figure 3A,C), but only in the apical and middle parts of the arms.
Highly esterified HGs were detected by the JIM7 antibodies. Fluorescence signals were observed in the cell walls in the apical and middle parts of the arms of the quadrifids (Figure 3D). The pectic polysaccharide (1–4)-β-d-galactan was detected by LM5 (Figure 3E,F). The arms of the quadrifids gave only a weak signal (Figure 3F). However, LM5 reacted with a strong fluorescence in the cell walls of the sectioned cells of the trap wall (Figure 3E).
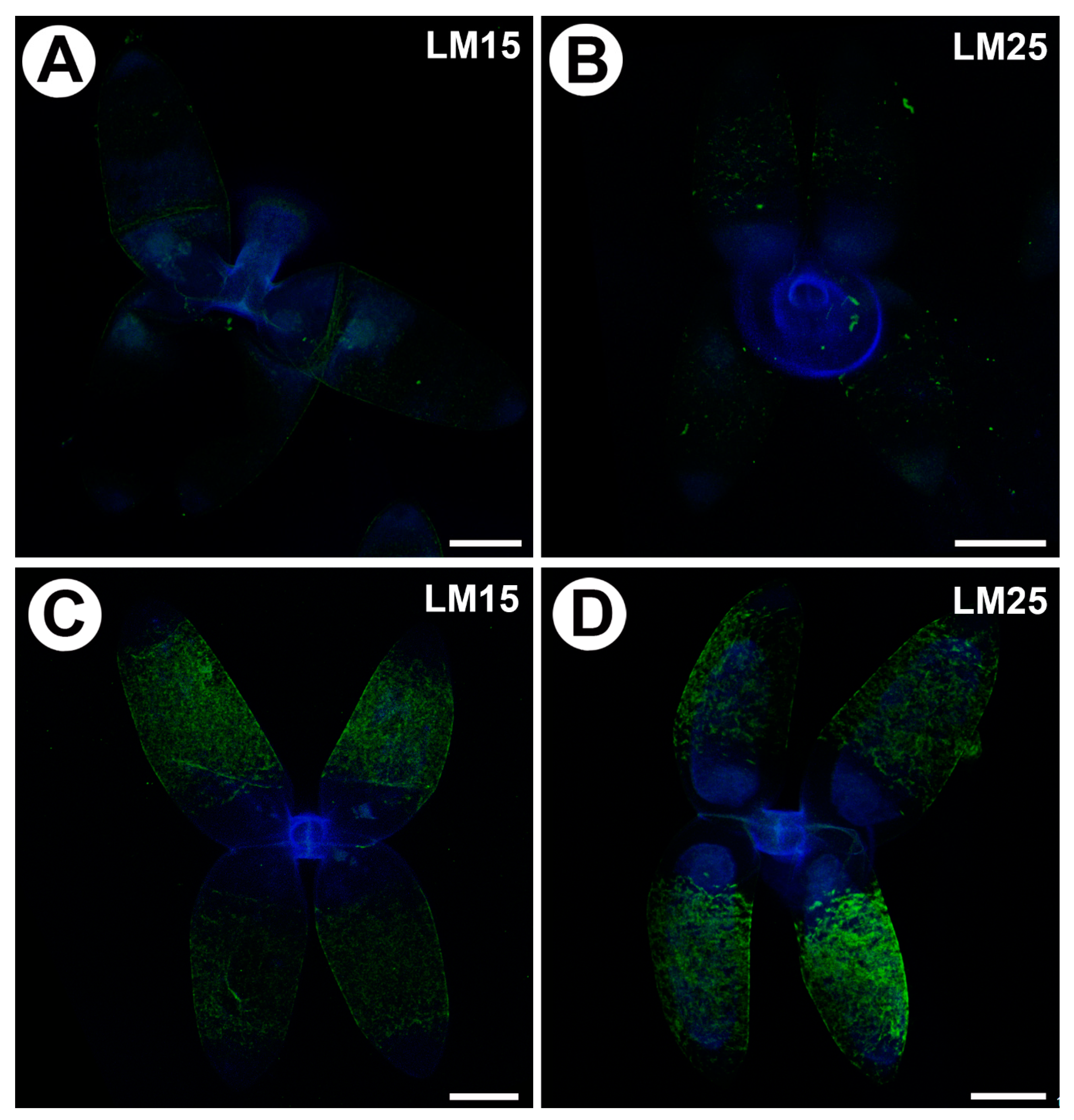
2.4. Distribution of Hemicellulose
Xyloglucan was detected by the LM15 and LM25 antibodies. Both gave a very weak fluorescence signal in the cell walls of the arms of the quadrifids (Figure 4A,B). When the sections were pre-treated with pectate lyase (during which pectins were removed), the LM15 antibody gave a stronger fluorescence signal (Figure 4C); similarly, the antibody signal was stronger in the case of LM25 (Figure 4D).
3. Discussion
3.1. Pros and Cons of the Whole-Mount Immunolabeled Trap Technique
Immunolabeling was not successful in the outer and inner epidermis of the traps. Immunolabeling was successful in the cell walls of the sectioned cells of the trap wall (outer and inner epidermis of the trap), where the cuticle was mechanically damaged and the cell wall was exposed. In the quadrifids, this method gave good results only in the terminal cells. Here, the antibodies penetrated the cell wall only in the middle and apical parts of the arms. Our results can be better understood if the formation and structure of the cuticle in Utricularia traps is considered. Fineran and Lee [25] described the ultrastructure of Utricularia trap cells in detail. They showed that the Utricularia trap epidermal cells have a developed cuticle. Moreover, the pedestal cell of the quadrifids has a cuticle and the lateral wall of the pedestal cell is also impregnated with cutin. According to these authors, the outermost region in the wall of the stalk (part of the terminal cell) is also heavily impregnated with an opaque cuticular material. Cuticular impregnation also occurs at the base of the arms. This would explain why immunolabeling was unsuccessful in these regions. However, in the middle and apical parts of the arm, the cuticle had a more open structure [25]; thus, the cuticle was permeable for antibodies, as we showed. The whole-mount immunolabeled organ technique enabled the dehydration, resin infiltration, and material slicing using a microtome to be avoided, which saved time and reagents. However, in our case, this technique should be regarded as a good preliminary method for quickly assessing the cell wall components in the most exposed cells with a discontinuous cuticle. Carnivorous plants have differently developed cuticles in their glandular structures. These cuticles are permeable because they have discontinuities such as cuticular gaps, cuticular pores, a ruptured cuticle, the open structure of cuticle, and a loose cuticle that is composed of cuticular droplets, e.g., [44,45,46,47,48,49,50,51]. Future research should focus on how these differentially formed cuticular discontinuities permit antibody penetration.
3.2. Cell Wall Polymers and Quadrifid Activity
The cell wall polymers are dynamic components and can be restructured and redistributed during plant development, reproduction, and organ aging [52,53,54,55,56,57]. These components play important roles during stresses and plant interactions with other organisms, e.g., bacteria and fungi [58,59,60,61,62]. Utricularia quadrifids are not only quite specialized trichomes in their structure; they also perform various functions, e.g., secretion of digestive enzymes, resorption of released nutrients, and participation in pumping out water [25,26,44]. Enzymes such as protease, phosphatase, and esterase were localized cytochemically in the quadrifids [31,32,63]. Aminopeptidase, phosphatase, β-hexosaminidase, α- and β-glucosidase were found in Utricularia trap fluid [64,65]. These trichomes are also active in respiration. Adamec [66] demonstrated the great respiratory activity of Utricularia traps and their trichomes immediately after firing, which results in the formation of anoxia, which kills prey inside the traps. Inside aquatic Utricularia traps, a rich community of commensal microorganisms exists, including bacteria, cyanobacteria, microfungi, algae, protozoa, and rotifers [67,68,69]. Thus, the quadrifids are also exposed to stress that is not only associated with contact with captured organisms, but also with a number of organisms that are living as commensals or parasites in the traps. For the above reasons, the presence of AGPs in the cell wall of the secretory cells of the quadrifids is not surprising, especially since these cells are responsible for the secretion of enzymes and subsequent nutrient intake, and these processes involve vesicle trafficking and membrane recycling, in which AGPs participate [70,71,72]. Moreover, the cell walls of the secretory cells in the glands of other carnivorous plants such as Drosophyllum [38], Dionaea [34,35] and Aldrovanda [36,37] are enriched with arabinogalactan proteins. We found that the cell walls of the arms are especially rich in low-methyl-esterified homogalacturonans. It is known that homogalacturonans are involved in plant cell wall porosity, elasticity, and hydration [73,74]. These features are important for transporting substances through the cell wall. Therefore, the presence of these pectins in the cell wall of glandular cells in quadrifids may be crucial for gland function (secretion of digestive enzymes, and later, absorption of nutrients from digested invertebrate bodies). However, to prove this, it would be necessary to study a mutant of Utricularia with a disruption of pectin production. However, in the glands of carnivorous plants from the families Droseraceae and Drosophyllaceae that have been studied to date, the cell walls of secretory cells have been poor in low-methyl-esterified homogalacturonans (as detected by the JIM5 and LM19 antibodies) [34,35,38]. One can only speculate about whether these differences are related to the phylogenetic position of the species or to the functioning of the glands. In the glands of carnivorous plants from the families Droseraceae and Drosophyllaceae that have been studied to date, the cell walls of the secretory cells are enriched with hemicelluloses (detected by the LM15 and LM25 antibodies, which both recognize xyloglucan). We found that in quadrifids, pectins mask the presence of hemicelluloses [75]. Hemicelluloses contribute to strengthening the cell wall by interaction with cellulose [76,77]; their increased presence may be related to the exposure of trichome cells to damage by organisms. We think that to better understand both the cell wall polymers and gland activity in Utricularia, samples cryo-fixed by high-pressure freezing and immunogold techniques are required, particularly since the method we used allows us to examine the components of the wall only to a limited extent. We do not know to what depth of the cell wall the penetration by antibodies reaches.
Future research should examine the composition of Utricularia cell walls in various vegetative organs, and compare this composition not only with trichome cell walls, but also with trap epidermal cells. Since there has been clear progress in genome analysis in Utricularia [9,78,79,80,81], we have hope for a future better understanding of how cell wall composition plays a role in trap function.
4. Materials and Methods
4.1. Plant Material
Utricularia dichotoma subsp. novae-zelandiae (Hook.f) R.W.Jobson [82] plants were grown in the greenhouses of the Botanical Garden of Jagiellonian University. The plants were cultivated in wet peat and exposed to natural sunlight.
4.2. Histological and Immunochemical Analysis
The traps were hand-sliced into two halves using a razor blade, and fixed in 8% (w/v) paraformaldehyde (PFA, Sigma-Aldrich, Sigma-Aldrich Sp. z o.o., Poznań, Poland) mixed with 0.25% (v/v) glutaraldehyde (GA, Sigma-Aldrich, Sigma-Aldrich Sp. z o.o., Poznań, Poland) in a PIPES buffer overnight at 4 °C. The PIPES buffer contained 50 mM PIPES (piperazine-N,N′-bis [2-ethanesulfonic acid], Sigma-Aldrich, Sigma-Aldrich Sp. z o.o., Poznań, Poland), 10 mM EGTA (ethylene gly col-bis[β-aminoethyl ether]N,N,N′,N′-tetraacetic acid, Sigma Aldrich, Poznań, Poland), and 1 mM MgCl2 (Sigma-Aldrich, Sigma-Aldrich Sp. z o.o., Poznań, Poland), pH 6.8. The plant material was washed in a PBS buffer and later blocked with 1% bovine serum albumin (BSA, Sigma-Aldrich) in a PBS buffer and incubated with the following primary antibodies (Table 1) [83]: anti-AGP: JIM8, JIM13, and JIM14 [84,85,86,87]; anti-pectin: JIM5, JIM7, LM19, and LM5 [75,84,88,89]; and anti-hemicelluloses: LM25 and LM15 [75,88,89], overnight at 4 °C. All of the primary antibodies were used in a 1:20 dilution. They were purchased from Plant Probes, Leeds, UK, and the goat anti-rat secondary antibody conjugated with FITC was purchased from Abcam (Abcam plc, Cambridge, UK). The signal of antibodies was visualized using laser 488 emission 498–550 (green fluorescence), and the autofluorescence of cell walls and cutin was visualized with laser 405 emission 415–460 (blue fluorescence). The samples were then cover-slipped using a Mowiol mounting medium (a mixture of Mowiol®4-88 (Sigma-Aldrich, Sigma-Aldrich Sp. z o.o., Poznań, Poland) and glycerol for fluorescence microscopy (Merck, Warsaw, Poland)), with the addition of 2.5% DABCO (The Carl Roth GmbH + Co. KG, Karlsruhe, Germany). The plant material was viewed using a Leica STELLARIS 5 WLL confocal microscope (Wetzlar, Germany) with lightning deconvolution. At least two different replications were performed for each of the analyzed traps, and about five to ten trichomes from each trap were analyzed for each antibody that was used. Negative controls were created by omitting the primary antibody step, which caused no fluorescence signal in any of the control frames for any of the stained slides (Figure S1B). To remove the HG from the cell walls, the traps were pretreated with 0.1 M sodium carbonate pH = 11.4 for 2 h at room temperature. This was followed by digestion with a pectate lyase 10 A (Nzytech) at 10 μg/mL for 2 h at room temperature in 50 mM N-cyclohexyl-3-aminopropane sulfonic acid (CAPS) with the addition of 2 mM of a CaCl2 buffer at pH 10 [38], and then incubation with the JIM5, JIM7, LM15, and LM25 antibodies, as described above.
4.3. Light Microscopy (LM)
The fluorescence of the mitochondria and nuclei in the quadrifids was observed by staining the traps with 20 μM DiOC6 (3,3′-dihexyloxacarbocyanine iodide; Thermo Fisher, Rockland, MD, USA) dissolved in water [90]. The traps were examined using a Nikon Eclipse E400 light microscope. Cutin and cuticle were stained using Sudan III (Sigma-Aldrich Sp. z o.o., Poznań, Poland) and Auramine O (Sigma-Aldrich Sp. z o.o., Poznań, Poland), and later, traps were examined using a Nikon Eclipse E400 light microscope (Tokyo, Japan) with a UV-2A filter (Ex. 330–380 nm, DM. 400 nm, Em. 420-α nm).
4.4. Morphological Observation
For the scanning electron microscopy (SEM), the traps were fixed in methanol and later transferred to ethanol, and then transferred to acetone and dried using supercritical CO2. The material was then sputter-coated with gold and examined at an accelerating voltage of 20 kV using a Hitachi S-4700 scanning electron microscope (Tokyo, Japan), which is housed at the Institute of Geological Sciences, Jagiellonian University, Kraków, Poland [38].
5. Conclusions
The advantage of analyzing whole traps without dehydrating, resin infiltrating, and slicing them using a microtome is that it saves time and reagents. However, our analyses have shown that in intact cells, only those parts of the cells that have a discontinuous cuticle gave positive results.
Here, we show that the cell walls of the secretory cells of the quadrifids were enriched with low-methyl-esterified homogalacturonans is in contrast to the cell walls of gland cells of carnivorous species from the Droseraceae and Drosophyllaceae families. However, a common characteristic of the secretion of these glands from the traps of carnivorous plants is the occurrence of arabinogalactan proteins in the cell walls, which may indirectly indicate the important role of these components in the functioning of glandular structures.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms25010056/s1.
Author Contributions
Conceptualization, B.J.P.; methodology, B.J.P. and M.K.; investigation, B.J.P. and M.K.; resources, B.J.P.; data curation, B.J.P.; writing—preparing the original draft, B.J.P.; writing—review and editing, B.J.P. and M.K.; visualization, B.J.P. and M.K.; supervision, B.J.P.; project administration, B.J.P.; funding acquisition, B.J.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported by the program “Excellence Initiative–Research University” at Jagiellonian University in Cracow, Poland.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank the Botanical Garden of Jagiellonian University for the opportunity to use plants from their garden collection. We sincerely thank the reviewers for their constructive comments, which helped to improve the quality of the article.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
AGPs—arabinogalactan proteins, HG—homogalacturonans, SEM—scanning electron microscope, LM—light microscope.
References
- Taylor, P. The Genus Utricularia: A Taxonomic Monograph; Kew: Kew Bulletin, Additional Series; Her Majesty’s Stationery Office: London, UK, 1989; Volume 4, pp. 1–724. [Google Scholar]
- Guisande, C.; Granado-Lorencio, C.; Andrade-Sossa, C.; Duque, S.R. Bladderworts. Funct. Plant Sci. Biotechnol. 2007, 1, 58–68. [Google Scholar]
- Jobson, R.W.; Baleeiro, P.C.; Guisande, C. Systematics and evolution of Lentibulariaceae: III. Utricularia. In Carnivorous Plants: Physiology, Ecology, and Evolution; Ellison, A.M., Adamec, L., Eds.; Oxford University Press: Oxford, UK, 2018; pp. 89–104. [Google Scholar]
- Miranda, V.F.O.; Silva, S.R.; Reut, M.S.; Dolsan, H.; Stolarczyk, P.; Rutishauser, R.; Płachno, B.J. A Historical Perspective of Bladderworts (Utricularia): Traps, Carnivory and Body Architecture. Plants 2021, 10, 2656. [Google Scholar] [CrossRef] [PubMed]
- Rutishauser, R.; Isler, B. Developmental genetics and morphological evolution of flowering plants, especially bladderworts (Utricularia): Fuzzy Arberian Morphology complements Classical Morphology. Ann. Bot. 2001, 88, 1173–1202. [Google Scholar] [CrossRef]
- Rutishauser, R. Evolution of unusual morphologies in Lentibulariaceae (bladderworts and allies) and Podostemaceae (river-weeds): A pictorial report at the interface of developmental biology and morphological diversification. Ann. Bot. 2016, 117, 811–832. [Google Scholar] [CrossRef]
- Reut, M.S.; Płachno, B.J. Unusual developmental morphology and anatomy of vegetative organs in Utricularia dichotoma—Leaf, shoot and root dynamics. Protoplasma 2020, 257, 371–390. [Google Scholar] [CrossRef] [PubMed]
- Reut, M.S.; Płachno, B.J. Development, Diversity and Dynamics of Plant Architecture in Utricularia subgenus Polypompholyx—Towards Understanding Evolutionary Processes in the Lentibulariaceae. Bot. Rev. 2023, 89, 201–236. [Google Scholar] [CrossRef]
- Carretero-Paulet, L.; Librado, P.; Chang, T.H.; Ibarra-Laclette, E.; Herrera-Estrella, L.; Rozas, J.; Albert, V.A. High gene family turnover rates and gene space adaptation in the compact genome of the carnivorous plant Utricularia gibba. Mol. Biol. Evol. 2015, 32, 1284–1295. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, F.E. The range of structural and functional variation in the traps of Utricularia. Flora 1931, 25, 260–276. [Google Scholar] [CrossRef]
- Lloyd, F.E. The door of Utricularia, an irritable mechanism. Can. J. Bot. 1932, 10, 780–786. [Google Scholar] [CrossRef]
- Lloyd, F.E. The range of structural and functional variety in the traps of Utricularia and Polypompholyx. Flora 1932, 126, 303–328. [Google Scholar] [CrossRef]
- Adamec, L. The smallest but fastest: Ecophysiological characteristics of traps of aquatic carnivorous Utricularia. Plant Signal. Behav. 2011, 6, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Płachno, B.J.; Adamec, L.; Kamińska, I. Relationship between trap anatomy and function in Australian carnivorous bladderworts (Utricularia) of the subgenus Polypompholyx. Aquat. Bot. 2015, 120, 290–296. [Google Scholar] [CrossRef]
- Poppinga, S.; Weisskopf, C.; Westermeier, A.S.; Masselter, T.; Speck, T. Fastest predators in the plant kingdom: Functional morphology and biomechanics of suction traps found in the largest genus of carnivorous plants. AoB Plants 2016, 8, plv140. [Google Scholar] [CrossRef] [PubMed]
- Westermeier, A.S.; Fleischmann, A.; Müller, K.; Schäferhoff, B.; Rubach, C.; Speck, T.; Poppinga, S. Trap diversity and character evolution in carnivorous bladderworts (Utricularia, Lentibulariaceae). Sci. Rep. 2017, 7, 12052. [Google Scholar] [CrossRef] [PubMed]
- Whitewoods, C.D.; Gonçalves, B.; Cheng, J.; Cui, M.; Kennaway, R.; Lee, K.; Bushell, C.; Yu, M.; Piao, C.; Coen, E. Evolution of carnivorous traps from planar leaves through simple shifts in gene expression. Science 2020, 367, 91–96. [Google Scholar] [CrossRef]
- Treat, M. Is the valve of Utricularia sensitive? Harper’s New Mon. Mag. 1876, 52, 382–387. [Google Scholar]
- Vincent, O.; Weißkopf, C.; Poppinga, S.; Masselter, T.; Speck, T.; Joyeux, M.; Quilliet, C.; Marmottant, P. Ultra-fast underwater suction traps. Proc. R. Soc. B Biol. Sci. 2011, 278, 2909–2914. [Google Scholar] [CrossRef]
- Vincent, O.; Roditchev, I.; Marmottant, P. Spontaneous firings of carnivorous aquatic Utricularia traps: Temporal patterns and mechanical oscillations. PLoS ONE 2011, 6, e20205. [Google Scholar] [CrossRef]
- Adamec, L.; Poppinga, S. Measurement of the critical negative pressure inside traps of aquatic carnivorous Utricularia species. Aquat. Bot. 2016, 133, 10–16. [Google Scholar] [CrossRef]
- Müller, U.K.; Berg, O.; Schwaner, J.M.; Brown, M.D.; Li, G.; Voesenek, C.J.; van Leeuwen, J.L. Bladderworts, the smallest known suction feeders, generate inertia-dominated flows to capture prey. New Phytol. 2020, 228, 586–595. [Google Scholar] [CrossRef]
- Poppinga, S.; Daber, L.E.; Westermeier, A.S.; Kruppert, S.; Horstmann, M.; Tollrian, R.; Speck, T. Biomechanical analysis of prey capture in the carnivorous Southern bladderwort (Utricularia australis). Sci. Rep. 2017, 7, 1776. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, F.E. The Carnivorous Plants; Chronica Botanica Company: Waltham, MA, USA, 1942. [Google Scholar]
- Fineran, B.A.; Lee, M.S.L. Organization of quadrifid and bifid hairs in the trap of Utricularia monanthos. Protoplasma 1975, 84, 43–70. [Google Scholar] [CrossRef]
- Fineran, B.A.; Lee, M.S.L. Organization of mature external glands on the trap and other organs of the bladderwort Utricularia monanthos. Protoplasma 1980, 103, 17–34. [Google Scholar] [CrossRef]
- Fineran, B.A. Glandular trichomes in Utricularia: A review of their structure and function. Isr. J. Bot. 1985, 34, 295–330. [Google Scholar]
- Yang, Y.P.; Liu, H.Y.; Chao, Y.S. Trap gland morphology and its systematics implication in Taiwan Utricularia (Lentibuariaceae). Flora 2009, 204, 692–699. [Google Scholar] [CrossRef]
- Płachno, B.J.; Świątek, P.; Adamec, L.; Carvalho, S.; Miranda, V.F.O. The Trap Architecture of Utricularia multifida and Utricularia westonii (subg. Polypompholyx). Front. Plant Sci. 2019, 10, 336. [Google Scholar] [CrossRef] [PubMed]
- Darwin, C.R. Insectivorous Plants; John Murray: London, UK, 1875. [Google Scholar]
- Vintéjoux, C. Ultrastructural and cytochemical observations on the digestive glands of Utricularia neglecta L. (Lentibulariaceae): Distribution of protease and acid phosphatase activities. Port. Acta Biol. 1974, 14, 463–474. [Google Scholar]
- Płachno, B.J.; Adamec, L.; Lichtscheidl, I.K.; Peroutka, M.; Adlassnig, W.; Vrba, J. Fluorescence labelling of phosphatase activity in digestive glands of carnivorous plants. Plant Biol. 2006, 8, 813–820. [Google Scholar] [CrossRef]
- Juang, T.C.C.; Juang, S.D.C.; Liu, Z.H. Direct evidence of the symplastic pathway in the trap of the bladderwort Utricularia gibba L. Bot. Stud. 2011, 52, 47–54. [Google Scholar]
- Płachno, B.J.; Kapusta, M.; Stolarczyk, P.; Świątek, P.; Strzemski, M.; Miranda, V.F.O. Immunocytochemical analysis of the wall ingrowths in the digestive gland transfer cells in Aldrovanda vesiculosa L. (Droseraceae). Cells 2022, 11, 2218. [Google Scholar] [CrossRef]
- Płachno, B.J.; Kapusta, M.; Stolarczyk, P.; Wójciak, M.; Świątek, P. Immunocytochemical analysis of bifid trichomes in Aldrovanda vesiculosa L. Traps. Int. J. Mol. Sci. 2023, 24, 3358. [Google Scholar] [CrossRef] [PubMed]
- Płachno, B.J.; Kapusta, M.; Stolarczyk, P.; Świątek, P. Arabinogalactan proteins in the digestive glands of Dionaea muscipula J. Ellis Traps. Cells 2022, 11, 586. [Google Scholar] [CrossRef] [PubMed]
- Płachno, B.J.; Kapusta, M.; Stolarczyk, P.; Świątek, P. Stellate trichomes in Dionaea muscipula Ellis (Venus Flytrap) Traps, Structure and Functions. Int. J. Mol. Sci. 2023, 24, 553. [Google Scholar] [CrossRef] [PubMed]
- Płachno, B.J.; Kapusta, M.; Stolarczyk, P.; Świątek, P.; Lichtscheidl, I. Differences in the Occurrence of Cell Wall Components between Distinct Cell Types in Glands of Drosophyllum lusitanicum. Int. J. Mol. Sci. 2023, 24, 15045. [Google Scholar] [CrossRef] [PubMed]
- Willats, W.G.; McCartney, L.; Knox, J.P. In-situ analysis of pectic polysaccharides in seed mucilage and at the root surface of Arabidopsis thaliana. Planta 2001, 213, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Larson, E.R.; Tierney, M.L.; Tinaz, B.; Domozych, D.S. Using monoclonal antibodies to label living root hairs: A novel tool for studying cell wall microarchitecture and dynamics in Arabidopsis. Plant Methods 2014, 10, 30. [Google Scholar] [CrossRef] [PubMed]
- Marzec, M.; Szarejko, I.; Melzer, M. Arabinogalactan proteins are involved in root hair development in barley. J. Exp. Bot. 2015, 5, 1245–1257. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Q.; Bruun, L.; Pierson, E.S.; Cresti, M. Periodic deposition of arabinogalactan epitopes in the cell wall of pollen tubes of Nicotiana tabacum L. Planta 1992, 188, 532–538. [Google Scholar] [CrossRef]
- Chebli, Y.; Kaneda, M.; Zerzour, R.; Geitmann, A. The cell wall of the Arabidopsis pollen tube--spatial distribution, recycling, and network formation of polysaccharides. Plant Physiol. 2012, 160, 1940–1955. [Google Scholar] [CrossRef]
- Juniper, B.E.; Robbins, R.J.; Joel, D.M. The Carnivorous Plants; Academic Press: London, UK, 1989. [Google Scholar]
- Williams, S.E.; Pickard, B.G. Secretion, absorption and cuticular structure in Drosera tentacles. Plant Physiol. 1969, 44, 5. [Google Scholar]
- Williams, S.E.; Pickard, B.G. Connections and barriers between cells of Drosera tentacles in relation to their electrophysiology. Planta 1974, 116, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ragetli, H.W.; Weintraub, L.O.E. Characteristics of Drosera tentacles. I. Anatomical and cytological details. Can. J. Bot. 1972, 50, 159–168. [Google Scholar] [CrossRef]
- Joel, D.M.; Rea, P.A.; Juniper, B.E. The cuticle of Dionaea muscipula Ellis (Venus’s Flytrap) in relation to stimulation, secretion and absorption. Protoplasma 1983, 114, 44–51. [Google Scholar] [CrossRef]
- Heide-Jørgensen, H.S. Cuticle development and ultrastructure: Evidence for a procuticle of high osmium affinity. Planta 1991, 183, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Płachno, B.J.; Faber, J.; Jankun, A. Cuticular discontinuities in glandular hairs of Genlisea St.-Hil. in relation to their functions. Acta Bot. Gallica 2005, 152, 125–130. [Google Scholar] [CrossRef]
- Anderson, B. Adaptations to foliar absorption of faeces: A pathway in plant carnivory. Ann. Bot. 2005, 95, 757–761. [Google Scholar] [CrossRef] [PubMed]
- Showalter, A.M.; Basu, D. Glycosylation of arabinogalactan-proteins essential for development in Arabidopsis. Commun. Integr. Biol. 2016, 9, e0125624. [Google Scholar] [CrossRef] [PubMed]
- Rancour, D.M.; Marita, J.M.; Hatfield, R.D. Cell wall composition throughout development for the model grass Brachypodium distachyon. Front. Plant Sci. 2012, 3, 266. [Google Scholar] [CrossRef]
- Leszczuk, A.; Cybulska, J.; Skrzypek, T.; Zdunek, A. Properties of arabinogalactan proteins (AGPs) in apple (Malus × domestica) fruit at different stages of ripening. Biology 2020, 9, 225. [Google Scholar] [CrossRef]
- Leszczuk, A.; Kalaitzis, P.; Blazakis, K.N.; Zdunek, A. The role of arabinogalactan proteins (AGPs) in fruit ripening—A review. Hortic. Res. 2020, 7, 176. [Google Scholar] [CrossRef]
- Hromadová, D.; Soukup, A.; Tylová, E. Arabinogalactan proteins in plant roots—An update on possible functions. Front. Plant Sci. 2021, 12, 674010. [Google Scholar] [CrossRef] [PubMed]
- Jia, K.; Wang, W.; Zhang, Q.; Jia, W. Cell wall integrity signaling in fruit ripening. Int. J. Mol. Sci. 2023, 24, 4054. [Google Scholar] [CrossRef] [PubMed]
- Le Gall, H.; Philippe, F.; Domon, J.M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell wall metabolism in response to abiotic stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.C.; Bulgakov, V.P.; Jinn, T.L. Pectin methylesterases: Cell wall remodeling proteins are required for plant response to heat stress. Front. Plant Sci. 2018, 9, 1612. [Google Scholar] [CrossRef] [PubMed]
- Leszczuk, A.; Szczuka, E.; Lewtak, K.; Chudzik, B.; Zdunek, A. Effect of low temperature on changes in AGP distribution during development of Bellis perennis ovules and anthers. Cells 2021, 10, 1880. [Google Scholar] [CrossRef] [PubMed]
- Pinski, A.; Betekhtin, A.; Skupien-Rabian, B.; Jankowska, U.; Jamet, E.; Hasterok, R. Changes in the cell wall proteome of leaves in response to high temperature stress in Brachypodium distachyon. Int. J. Mol. Sci. 2021, 22, 6750. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S. Cell wall signaling in plant development and defense. Annu. Rev. Plant Biol. 2022, 73, 323–353. [Google Scholar] [CrossRef]
- Heslop-Harrison, Y. Enzyme release in carnivorous plants. In Lysozymes in Biology and Pathology; Dingle, J.T., Dean, R.T., Eds.; North Holland Publishing: Amsterdam, The Netherlands, 1975; pp. 525–578. [Google Scholar]
- Sirová, D.; Adamec, L.; Vrba, J. Enzymatic activities in traps of four aquatic species of the carnivorous genus Utricularia. New Phytol. 2003, 159, 669–675. [Google Scholar] [CrossRef]
- Adamec, L.; Sirová, D.; Vrba, J.; Rejmánková, E. Enzyme production in the traps of aquatic Utricularia species. Biologia 2010, 65, 273–278. [Google Scholar] [CrossRef]
- Adamec, L. Oxygen concentrations inside the traps of the carnivorous plants Utricularia and Genlisea (Lentibulariaceae). Ann. Bot. 2007, 100, 849–856. [Google Scholar] [CrossRef]
- Richards, J.H. Bladder function in Utricularia purpurea (Lentibulariaceae): Is carnivory important? Am. J. Bot. 2001, 88, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Sirová, D.; Borovec, J.; Cerná, B.; Rejmánková, E.; Adamec, L.; Vrba, J. Microbial community development in the traps of aquatic Utricularia species. Aquat. Bot. 2009, 90, 129–136. [Google Scholar] [CrossRef]
- Sirová, D.; Bárta, J.; Šimek, K.; Posch, T.; Pech, J.; Stone, J.; Borovec, J.; Adamec, L.; Vrba, J. Hunters or farmers? Microbiome characteristics help elucidate the diet composition in an aquatic carnivorous plant. Microbiome 2018, 6, 225. [Google Scholar] [CrossRef] [PubMed]
- Samaj, J.; Samajova, O.; Baluska, E.; Lichtscheidl, I.; Knox, J.P.; Volkmann, D. Immunolocalization of LM2 arabinogalactan protein epitope associated with endomembranes of plant cells. Protoplasma 2000, 212, 186–196. [Google Scholar] [CrossRef]
- Olmos, E.; García De La Garma, J.; Gomez-Jimenez, M.C.; Fernandez-Garcia, N. Arabinogalactan proteins are involved in salt-adaptation and vesicle trafficking in Tobacco BY-2 cell cultures. Front. Plant Sci. 2017, 8, 1092. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cheng, M.; Yang, Q.; Li, J.; Wang, X.; Zhou, Q.; Nagawa, S.; Xia, B.; Xu, T.; Huang, R.; et al. Arabinogalactan protein-rare earth element complexes activate plant endocytosis. Proc. Natl. Acad. Sci. USA 2019, 16, 14349–14357. [Google Scholar] [CrossRef] [PubMed]
- Ridley, M.A.; O’Neill, D.; Mohnen, D. Pectins: Structure, biosynthesis, and oligogalacturonide-related signaling. Phytochemistry 2001, 57, 929–967. [Google Scholar] [CrossRef] [PubMed]
- Peaucelle, A.; Braybrook, S.; Höfte, H. Cell wall mechanics and growth control in plants: The role of pectins revisited. Front. Plant Sci. 2012, 3, 121. [Google Scholar] [CrossRef]
- Marcus, S.E.; Verhertbruggen, Y.; Hervé, C.; Ordaz-Ortiz, J.J.; Farkas, V.; Pedersen, H.L.; Willats, W.G.; Knox, J.P. Pectic homogalacturonan masks abundant sets of xyloglucan epitopes in plant cell walls. BMC Plant Biol. 2008, 8, 60. [Google Scholar] [CrossRef]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef]
- Park, Y.B.; Cosgrove, D.J. Xyloglucan and its interactions with other components of the growing cell wall. Plant Cell Physiol. 2015, 56, 180–194. [Google Scholar] [CrossRef] [PubMed]
- Lan, T.; Renner, T.; Ibarra-laclette, E.; Farr, K.M.; Chang, T.H.; Cervantes-Pérez, S.A.; Zheng, C.; Sankoff, D.; Tang, H.; Purbojati, R.W.; et al. Long-read sequencing uncovers the adaptive topography of a carnivorous plant genome. Proc. Natl. Acad. Sci. USA 2017, 114, E4435–E4441. [Google Scholar] [CrossRef] [PubMed]
- Carretero-Paulet, L.; Chang, T.H.; Librado, P.; Ibarra-Laclette, E.; Herrera-Estrella, L.; Rozas, J.; Albert, V.A. Genome-wide analysis of adaptive molecular evolution in the carnivorous plant Utricularia gibba. Genome Biol. Evol. 2015, 7, 444–456. [Google Scholar] [CrossRef]
- Bárta, J.; Stone, J.D.; Pech, J.; Sirová, D.; Adamec, L.; Campbell, M.A.; Štorchová, H. The transcriptome of Utricularia vulgaris, a rootless plant with minimalist genome, reveals extreme alternative splicing and only moderate sequence similarity with Utricularia gibba. BMC Plant Biol. 2015, 15, 78. [Google Scholar] [CrossRef] [PubMed]
- Laspisa, D.; Llla-Berenguer, E.; Bang, S.; Schmitz, R.J.; Parrott, W.; Wallace, J. Mining the Utricularia gibba genome for insulator-like elements for genetic engineering. Front. Plant Sci. 2023, 14, 1279231. [Google Scholar] [CrossRef]
- Jobson, R.W.; Baleeiro, P.C. Radiations of fairy-aprons (Utricularia dichotoma, Lentibulariaceae) in Australia and New Zealand: Molecular evidence and proposal of new subspecies. Aust. Syst. Bot. 2020, 33, 278–310. [Google Scholar] [CrossRef]
- Available online: https://www.kerafast.com/cat/799/paul-knox-phd (accessed on 13 November 2023).
- Knox, J.P.; Day, S.; Roberts, K. A set of cell surface glycoproteins forms an early position, but not cell type, in the root apical meristem of Daucus carota L. Development 1989, 106, 47–56. [Google Scholar] [CrossRef]
- Pennell, R.I.; Knox, J.P.; Scofield, G.N.; Selvendran, R.R.; Roberts, K. A family of abundant plasma membrane-associated glycoproteins related to the arabinogalactan proteins is unique to flowering plants. J. Cell Biol. 1989, 108, 1967–1977. [Google Scholar] [CrossRef]
- Pennell, R.I.; Janniche, L.; Kjellbom, P.; Scofield, G.N.; Peart, J.M.; Roberts, K. Developmental regulation of a plasma membrane arabinogalactan protein epitope in oilseed rape flowers. Plant Cell 1991, 3, 1317–1326. [Google Scholar] [CrossRef]
- Knox, J.P.; Linstead, P.J.; Cooper, J.P.C.; Roberts, K. Developmentally regulated epitopes of cell surface arabinogalactan proteins and their relation to root tissue pattern formation. Plant J. 1991, 1, 317–326. [Google Scholar] [CrossRef]
- Verhertbruggen, Y.; Marcus, S.E.; Haeger, A.; Ordaz-Ortiz, J.J.; Knox, J.P. An extended set of monoclonal antibodies to pectic homogalacturonan. Carbohydr. Res. 2009, 344, 1858–1862. [Google Scholar] [CrossRef] [PubMed]
- McCartney, L.; Marcus, S.E.; Knox, J.P. Monoclonal antibodies to plant cell wall xylans and arabinoxylans. J. Histochem. Cytochem. 2005, 53, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Lichtscheidl, I.; Lancelle, S.; Weidinger, M.; Adlassnig, W.; Koller-Peroutka, M.; Bauer, S.; Krammer, S.; Hepler, P.K. Gland cell responses to feeding in Drosera capensis, a carnivorous plant. Protoplasma 2021, 258, 1291–1306. [Google Scholar] [CrossRef] [PubMed]
Figure 2.
Arabinogalactan proteins detected in the quadrifids of Utricularia dichotoma subsp. novae-zelandiae. Green fluorescence is a signal of antibodies. Blue fluorescence shows the autofluorescence of the cell wall and cuticle. (A) Arabinogalactan proteins (labeled with JIM8) were detected in the quadrifid, scale bar 10 µm. (B) Arabinogalactan proteins (labeled with JIM13) were detected in the quadrifid, scale bar 10 µm. (C) Arabinogalactan proteins (labeled with JIM14) were detected in the quadrifid, scale bar 10 µm.
Figure 2.
Arabinogalactan proteins detected in the quadrifids of Utricularia dichotoma subsp. novae-zelandiae. Green fluorescence is a signal of antibodies. Blue fluorescence shows the autofluorescence of the cell wall and cuticle. (A) Arabinogalactan proteins (labeled with JIM8) were detected in the quadrifid, scale bar 10 µm. (B) Arabinogalactan proteins (labeled with JIM13) were detected in the quadrifid, scale bar 10 µm. (C) Arabinogalactan proteins (labeled with JIM14) were detected in the quadrifid, scale bar 10 µm.
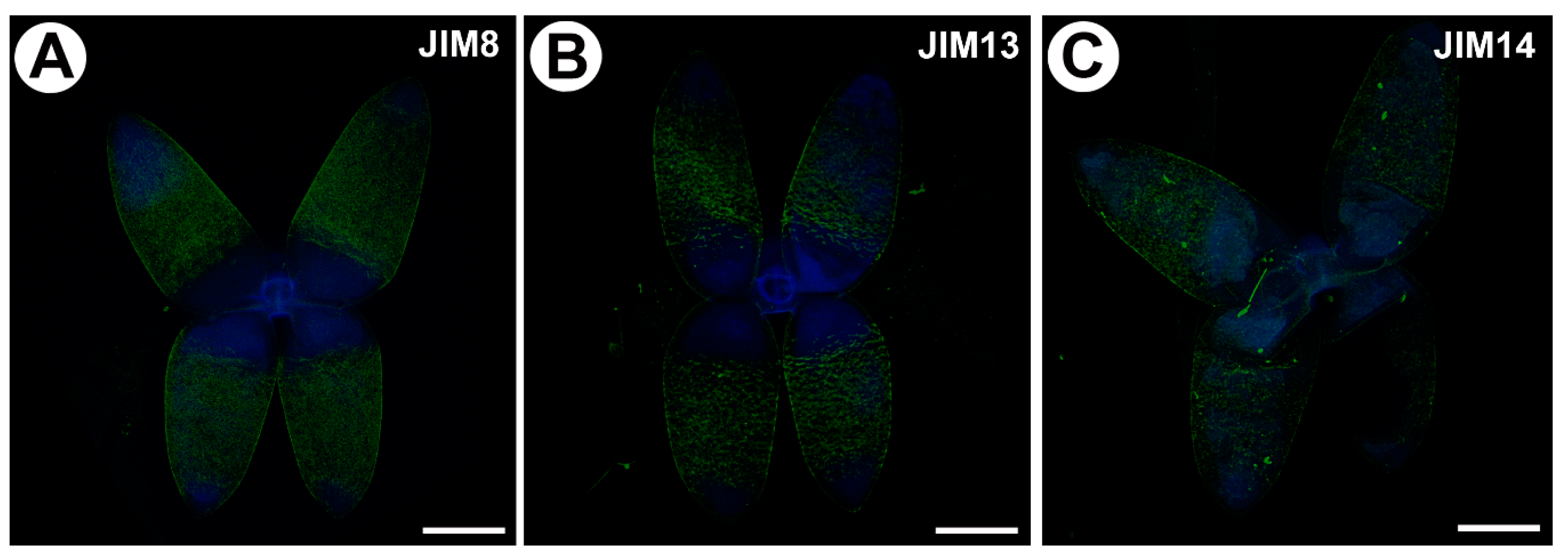
Figure 3.
Homogalacturonan (HG) detected in the in the quadrifids of Utricularia dichotoma subsp. novae-zelandiae. Green fluorescence is a sign of antibodies. Blue fluorescence indicates autofluorescence of the cell wall and cuticle. (A) HG (labeled with JIM5) detected in the quadrifids, scale bar 10 µm. (B) HG (labeled with LM19) detected in the quadrifids, scale bar 500 µm. (C) HG (labeled with LM19) detected in the quadrifids, scale bar 10 µm. (D) HG (labeled with JIM7) detected in the quadrifids, scale bar 10 µm. (E) HG (labeled with LM5) detected in the trap; note the blue autofluorescence of quadrifids inside the trap and the strong green fluorescence signal of antibodies in the cell walls of the trap wall, scale bar 500 µm. (F) HG (labeled with LM5) detected in the quadrifid, scale bar 10 µm.
Figure 3.
Homogalacturonan (HG) detected in the in the quadrifids of Utricularia dichotoma subsp. novae-zelandiae. Green fluorescence is a sign of antibodies. Blue fluorescence indicates autofluorescence of the cell wall and cuticle. (A) HG (labeled with JIM5) detected in the quadrifids, scale bar 10 µm. (B) HG (labeled with LM19) detected in the quadrifids, scale bar 500 µm. (C) HG (labeled with LM19) detected in the quadrifids, scale bar 10 µm. (D) HG (labeled with JIM7) detected in the quadrifids, scale bar 10 µm. (E) HG (labeled with LM5) detected in the trap; note the blue autofluorescence of quadrifids inside the trap and the strong green fluorescence signal of antibodies in the cell walls of the trap wall, scale bar 500 µm. (F) HG (labeled with LM5) detected in the quadrifid, scale bar 10 µm.
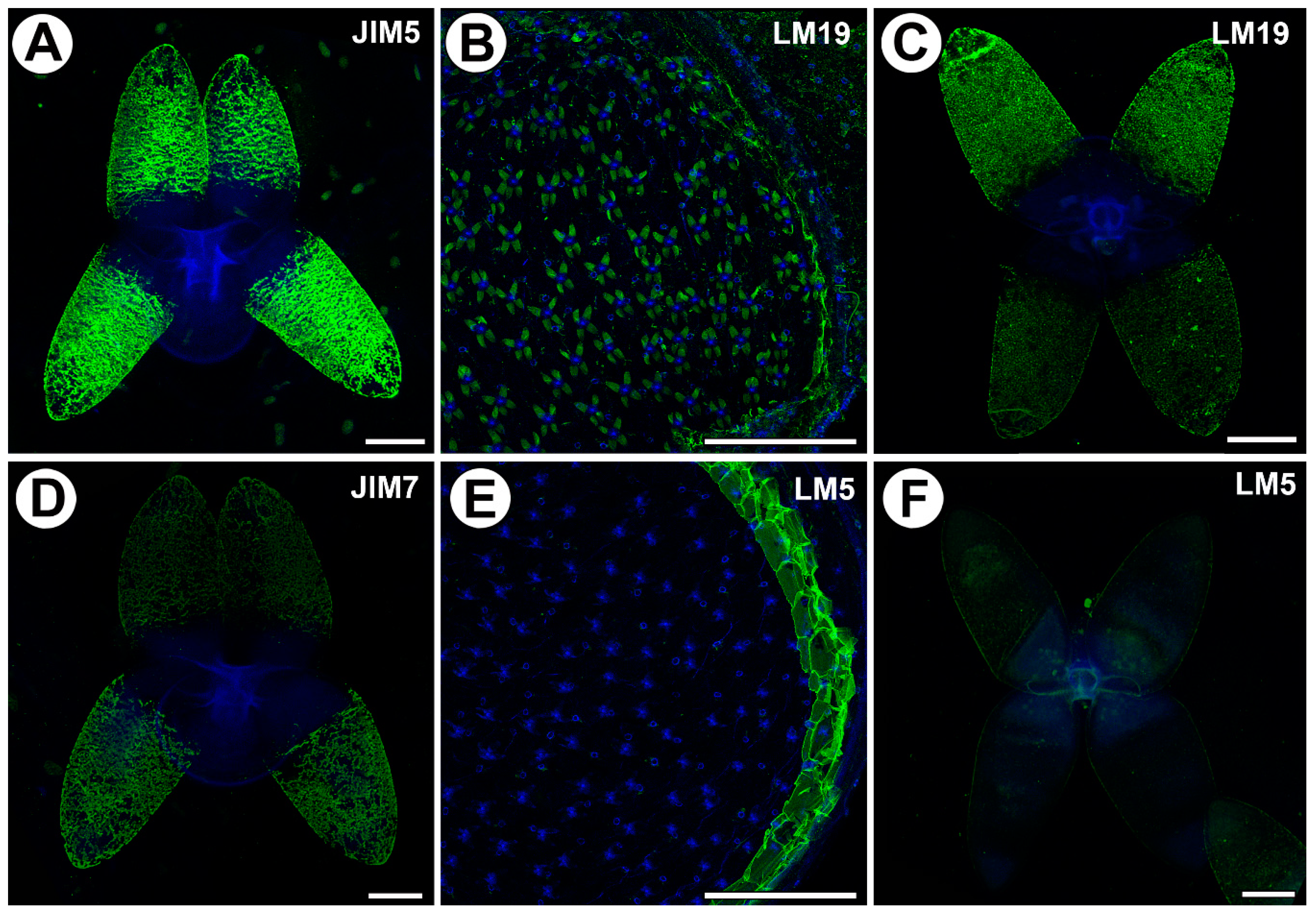
Figure 4.
Xyloglucan detected in the quadrifids of Utricularia dichotoma subsp. novae-zelandiae. Green fluorescence is a sign of antibodies. Blue fluorescence indicates autofluorescence of the cell wall, cuticle and vacuole. (A) Xyloglucan (labeled with LM15) detected in the quadrifids, scale bar 10 µm. (B) Xyloglucan (labeled with LM25) detected in the quadrifids, scale bar 10 µm. (C) Xyloglucan (labeled with LM15) detected in the quadrifids after they had been pre-treated with pectate lyase, scale bar 10 µm. (D) Xyloglucan (labeled with LM25) detected in the quadrifids after they had been pre-treated with pectate lyase, scale bar 10 µm.
Figure 4.
Xyloglucan detected in the quadrifids of Utricularia dichotoma subsp. novae-zelandiae. Green fluorescence is a sign of antibodies. Blue fluorescence indicates autofluorescence of the cell wall, cuticle and vacuole. (A) Xyloglucan (labeled with LM15) detected in the quadrifids, scale bar 10 µm. (B) Xyloglucan (labeled with LM25) detected in the quadrifids, scale bar 10 µm. (C) Xyloglucan (labeled with LM15) detected in the quadrifids after they had been pre-treated with pectate lyase, scale bar 10 µm. (D) Xyloglucan (labeled with LM25) detected in the quadrifids after they had been pre-treated with pectate lyase, scale bar 10 µm.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of the monoclonal antibodies used in the current study, and the epitopes they recognize [83].
Table 1.
List of the monoclonal antibodies used in the current study, and the epitopes they recognize [83].
| Antibody | Epitope |
|---|---|
| AGPs | |
| JIM8 | Arabinogalactan |
| JIM13 | Arabinogalactan/arabinogalactan protein |
| JIM14 | Arabinogalactan/arabinogalactan protein |
| Homogalacturonan | |
| JIM5 | Homogalacturonan (HG) domain of c pectic polysaccharides; recognizes partially methyl-esterified epitopes of HG, and can also bind to unesterified HG |
| JIM7 | HG domain of the pectic polysaccharides; recognizes partially methyl-esterified epitopes of HG, but does not bind to unesterified HG |
| LM5 | Linear tetrasaccharide in (1–4)-β-d-galactans (RGI side chain) |
| LM19 | HG domain in pectic polysaccharides; recognizes a range of HG with a preference to bind strongly to unesterified HG |
| Hemicelluloses | |
| LM15 | XXXG motif of xyloglucan |
| LM25 | XLLG, XXLG and XXXG motifs of xyloglucan |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Płachno, B.J.; Kapusta, M. The Localization of Cell Wall Components in the Quadrifids of Whole-Mount Immunolabeled Utricularia dichotoma Traps. Int. J. Mol. Sci. 2024, 25, 56. https://doi.org/10.3390/ijms25010056
AMA Style
Płachno BJ, Kapusta M. The Localization of Cell Wall Components in the Quadrifids of Whole-Mount Immunolabeled Utricularia dichotoma Traps. International Journal of Molecular Sciences. 2024; 25(1):56. https://doi.org/10.3390/ijms25010056
Chicago/Turabian StylePłachno, Bartosz J., and Małgorzata Kapusta. 2024. "The Localization of Cell Wall Components in the Quadrifids of Whole-Mount Immunolabeled Utricularia dichotoma Traps" International Journal of Molecular Sciences 25, no. 1: 56. https://doi.org/10.3390/ijms25010056
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.