
HCC-Related lncRNAs: Roles and Mechanisms

1
Department of Human and Molecular Genetics, Virginia Commonwealth University, Richmond, VA 23298, USA
2
Department of Human and Molecular Genetics, Massey Comprehensive Cancer Center, and VCU Institute of Molecular Medicine (VIMM), Virginia Commonwealth University, Richmond, VA 23298, USA
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2024, 25(1), 597; https://doi.org/10.3390/ijms25010597
Submission received: 11 November 2023
/
Revised: 21 December 2023
/
Accepted: 22 December 2023
/
Published: 2 January 2024
(This article belongs to the Special Issue Non-coding RNAs' Functionality-Diagnosis and Therapy in Cancer and Other Indications)
Abstract
:Hepatocellular carcinoma (HCC) presents a significant global health threat, particularly in regions endemic to hepatitis B and C viruses, and because of the ongoing pandemic of obesity causing metabolic-dysfunction-related fatty liver disease (MAFLD), a precursor to HCC. The molecular intricacies of HCC, genetic and epigenetic alterations, and dysregulated signaling pathways facilitate personalized treatment strategies based on molecular profiling. Epigenetic regulation, encompassing DNA methyltion, histone modifications, and noncoding RNAs, functions as a critical layer influencing HCC development. Long noncoding RNAs (lncRNAs) are spotlighted for their diverse roles in gene regulation and their potential as diagnostic and therapeutic tools in cancer. In this review, we explore the pivotal role of lncRNAs in HCC, including MAFLD and viral hepatitis, the most prevalent risk factors for hepatocarcinogenesis. The dysregulation of lncRNAs is implicated in HCC progression by modulating chromatin regulation and transcription, sponging miRNAs, and influencing structural functions. The ongoing studies on lncRNAs contribute to a deeper comprehension of HCC pathogenesis and offer promising routes for precision medicine, highlighting the utility of lncRNAs as early biomarkers, prognostic indicators, and therapeutic targets.
1. Introduction
Hepatocellular carcinoma (HCC) is the most common primary liver cancer arising from hepatocytes. It is the sixth most common cancer and the third most common cause of cancer-related deaths globally according to Global Cancer Incidence, Mortality and Prevalence (GLOBOCAN) 2020 [1]. It develops predominantly in the context of chronic liver disease and is characterized by its aggressive nature and limited treatment options. HCC is a significant global health concern with considerable regional variation. The highest incidence rates are observed in regions where HCC-causing chronic hepatitis B virus (HBV) and hepatitis C virus (HCV) infections are endemic, such as Eastern and South-Eastern Asia (Mongolia, Thailand, Cambodia and Vietnam), Northern and Western Africa (e.g., Egypt and Niger), and sub-Saharan Africa [1]. Other significant risk factors include alcoholic liver disease; non-alcoholic fatty liver disease (NAFLD), which has been renamed metabolic dysfunction-associated fatty liver disease (MAFLD); and aflatoxin exposure [1,2]. Recent studies emphasize the increasing prevalence of HCC in Western countries due to the rising rates of MAFLD [3]. Because of the diverse nature of its etiology, understanding the molecular mechanisms underlying HCC has been a major focus of research.
Genetic and epigenetic alterations, including mutations in genes such as TP53, CTNNB1, and TERT, are common in HCC [4]. The dysregulation of key signaling pathways, such as the Wnt/β-catenin, MAPK, and PI3K-AKT pathways, plays a critical role in HCC development and progression [5]. Recent research has elucidated the intricate crosstalk among these pathways and their potential as therapeutic targets [5]. The roles of noncoding RNAs (ncRNAs), epigenetic modifications, and microenvironmental factors in shaping the HCC landscape are being uncovered, offering novel targets for intervention [6,7]. The early diagnosis of HCC is crucial for improving patient outcomes. Surveillance strategies include the use of imaging modalities such as ultrasound, computed tomography (CT), and magnetic resonance imaging (MRI), coupled with alpha-fetoprotein (AFP) measurements. Advances in imaging techniques have improved early detection and staging accuracy [8]. Liquid biopsy approaches, such as circulating tumor DNA (ctDNA) analysis, are also being explored for their potential to detect HCC-related mutations and biomarkers [9].
HCC management depends on the stage at diagnosis and the patient’s overall health [10]. Surgical resection, liver transplantation, and locoregional therapies, like radiofrequency ablation (RFA) and transarterial chemoembolization (TACE), remain crucial for early-stage HCC [11]. Immune checkpoint inhibitors, such as anti-PD-1 antibodies nivolumab and pembrolizumab, have shown promise in advanced HCC treatment [12]. Currently, the most effective FDA-approved treatment for advanced non-resectable HCC is a combination of anti-PD-L1 (atezolizumab) and anti-VEGF (bevacizumab) antibodies, providing an overall response rate of 27% [13,14,15]. Additionally, targeted therapies, including tyrosine kinase inhibitors (TKIs), like sorafenib and lenvatinib, continue to be integral in systemic HCC treatment [16]. Prevention strategies are vital in HCC management. Vaccination against HBV, treatment of HCV infection, and lifestyle modifications to reduce MAFLD risk are essential preventive measures [10,17]. Future research efforts focus on developing innovative therapies, identifying early biomarkers, and refining surveillance strategies to further improve HCC outcomes.
2. Epigenetic Regulation
Epigenetic regulation represents a critical layer of gene expression control that extends beyond the genetic code. It encompasses a range of molecular processes that modify the chromatin landscape and thereby govern gene activity [18]. Epigenetic mechanisms play a fundamental role in cellular differentiation, development, and adaptation to environmental stimuli. Epigenetic regulation in HCC involves a complex interplay of molecular mechanisms that control gene expression and function without altering the underlying DNA sequence. These epigenetic modifications can significantly impact the development and progression of HCC.
2.1. DNA Methylation: Beyond the Methyl Mark
DNA methylation, characterized by the addition of methyl groups to cytosine residues, is a hallmark of epigenetic regulation. Recent research has unveiled the intricate dynamics of DNA methylation patterns in response to developmental cues and environmental factors [19]. Methylation patterns exert direct control over gene expression by influencing transcriptional accessibility and ultimately shaping cellular phenotypes [20]. The hypermethylation of promoter regions in tumor suppressor genes, such as CDKN2A (p16) and APC, can lead to their inactivation [21]. Conversely, global hypomethylation in HCC can contribute to genomic instability and tumorigenesis [22].
2.2. Histone Modifications: The Chromatin Code
Histone modifications, including acetylation, methylation and phosphorylation, serve as the chromatin code that regulates gene accessibility and transcription. Advances in chromatin biology have illuminated the intricate interplay among various histone marks, revealing their roles in orchestrating gene expression programs [23]. Recent studies have emphasized the significance of histone modifications in cellular plasticity and disease processes [24]. Aberrant histone modifications can lead to changes in chromatin structure and gene accessibility, impacting gene expression patterns and resulting in HCC [25].
2.3. Noncoding RNAs: Epigenetic Mediators
In higher eukaryotes, the genome is pervasively transcribed to generate a large number of noncoding RNAs (ncRNAs), particularly long noncoding RNAs (lncRNAs) and microRNAs (miRNAs), which have emerged as crucial epigenetic mediators. They participate in gene silencing, chromatin remodeling, and post-transcriptional regulation [26]. Recent investigations have unraveled the functional diversity of ncRNAs, shedding light on their roles in cellular homeostasis and pathology [27]. The dysregulation of ncRNAs, including miRNAs, such as miR-21 and miR-122, and lncRNAs, is a hallmark of HCC regulating the development and progression of the disease [28]. The field of lncRNA is growing at a very fast speed, unraveling novel mechanisms of action. In this review, we aim to provide a comprehensive and up-to-date description of the role of lncRNAs in hepatocarcinogenesis, highlighting new definitions of lncRNAs, novel mechanisms of action, potential use as diagnostic and prognostic marker, and targets for therapeutic intervention.
3. Introduction to lncRNAs: Mechanisms and Modes of Action
LncRNAs have taken the center stage of molecular biology research, with a surge in investigations uncovering their intricate roles in diverse cellular processes. LncRNAs are typically defined as transcripts exceeding 200 nucleotides (nt) without significant protein-coding potential. However, a recent consensus statement recommends the definition of lncRNAs as ncRNAs greater than 500 nt in length and mainly generated by RNA polymerase II (Pol II), to distinguish them from transcripts generated by RNA polymerase III (Pol III) and some Pol II transcripts, such as snRNAs and intron-derived snoRNAs, which are ~50–500 nt in size [29]. Additionally, lncRNAs coding for biologically active micropeptides are increasingly being identified [30]. Thus, the definition of lncRNAs is undergoing continuous evolution.
There are multiple classification systems for lncRNAs encompassing various categories, such as length of transcript, genomic location and context, sequence and structure conservation, effects on DNA sequences, functional mechanisms and targeting mechanisms, and association with protein coding genes or subcellular structures [31,32]. In relation to the sites of transcription relative to annotated protein-coding genes, lncRNAs are classified as sense, antisense, bidirectional, intronic, and intergenic (Figure 1) [33]. Contemporary studies have illuminated the vast functional repertoire of lncRNAs. These molecules play pivotal roles in gene regulation at both transcriptional and post-transcriptional levels. They participate in chromatin remodeling, epigenetic modifications, and subcellular compartmentalization, exerting influence over fundamental cellular processes and pathways [34]. Understanding the underlying mechanisms and modes of action by which lncRNAs exert their influence is fundamental to unraveling their significance in cellular biology.
3.1. Gene Expression Regulation
One of the primary mechanisms through which lncRNAs operate is the regulation of gene expression. They can modulate transcriptional processes by interacting with chromatin, acting as scaffolds for transcriptional complexes, or influencing the recruitment of transcription factors (Figure 2). lncRNAs also regulate gene expression at post-transcriptional and translational levels.
3.1.1. Regulation of Chromatin
RNAs have inherent chromatin regulatory function because the negatively charged RNAs can neutralize positively charged histone tails, resulting in the opening up of chromatin structure in a rapid manner [35]. LncRNAs are similarly involved in the complex regulation of chromatin architecture either by directly binding to DNA or by interacting with DNA-binding and RNA-binding proteins in either a cis or a trans manner.
LncRNA–Protein Interaction Mediating Chromatin Regulation
Homeodomain transcription factors Hox maintain positional identity, and lncRNA HOTTIP is transcribed from the 5′ end of HOXA locus in an antisense direction [36]. HOTTIP binds to the WD repeat domain 5 (WDR5) protein, a core component of the MLL/SET1 histone methyltransferase complex, and guides this complex to specific genomic loci, including the HOXA gene cluster. This results in the trimethylation of histone H3 at lysine 4 (H3K4me3) and the activation of HOXA genes, providing an example of the cis regulation of gene expression by a lncRNA [36].
Conversely, lncRNAs can interact with chromatin modifiers, sequestering them from promoters of specific genes. Histone deactylase SIRT6 inhibits many pluripotency-associated genes, and lncPRESS1 interacts with and sequesters SIRT6, thereby maintaining the acetylation of histone H3 at lysine 56 and lysine 9 (H3K56ac and H3K9ac) in the promoters of these genes and facilitating the pluripotency of embryonic stem cells (ESCs) [37]. p53 induces the expression of developmental genes and the differentiation of ESCs by inhibiting lncPRESS1 expression, which frees up SIRT6, allowing it to deacetylate and inhibit the expression of pluripotency genes [37].
Polycomb Repressive Complex 2 (PRC2) is an evolutionarily conserved gene-silencing machinery inducing the trimethylation of histone H3 at lysine 27 (H3K27me3), resulting in epigenetic silencing and the transcriptional suppression of target genes. PRC2 complex contains multiple proteins, including EZH2 and SUZ12, and many lncRNAs interact with these proteins recruiting PRC2 to specific genomic sites and causing methylation-induced transcription repression. One such example is HOTAIR, which can recruit PRC2 to numerous sites, and the overexpression of HOTAIR in an epithelial cell shifts the PRC2 occupancy pattern to the embryonic fibroblast pattern, thereby facilitating invasion and metastasis by these cells [38]. The lncRNA ANRIL binds to CBX7 in PRC2 and mediates the CBX7-mediated suppression of cyclin-dependent kinase inhibitors CDKN2A and CDKN2B, thereby preventing cellular senescence [39].
LncRNA-DNA Interaction Mediating Chromatin Regulation
During transcription, lncRNAs can form RNA–DNA hybrids, such as R-loops, which can be recognized by chromatin modifiers or transcription factors to either activate or inhibit transcription. lncRNA TARID, generated from the antisense strand of the TCF21 gene, forms an R-loop at the promoter of the transcription factor TCF21 [40]. GADD45A binds to this R-loop and recruits methylcytosine dioxygenase TET1, thereby facilitating the local demethylation of TCF21 promoter and inducing its expression [40]. TCF21 is a tumor-suppressor gene regulating the cell cycle, and the TARID-regulated mechanism allows TCF21 expression in normal cells while both TARID and TCF21 are hypermethylated, and hence downregulated, in cancer cells [41].
3.1.2. Regulation of Transcription
LncRNAs regulate transcription in multiple ways, including interaction with a variety of transcription factors (TFs) either inhibiting or activating their functions [42]. lncRNA GAS5, expressed in growth-arrested cells, binds to the glucocorticoid receptor (GR), inhibiting the GR-dependent expression of apoptosis-inhibiting genes [43]. Brain-specific lncRNA RMST interacts with transcription factor SOX2, facilitating its DNA-binding and promoting neuronal differentiation [44]. CHASERR is an evolutionarily conserved lncRNA that is transcribed from near the transcription start site of the ATP-dependent chromatin-remodeling enzyme CHD2 [45]. CHASERR expression interferes with transcription factor accessibility to the CHD2 promoter, and CHASERR loss leads to increased CHD2 expression [45]. Homozygous Chaserr knockout mice are embryonically lethal, and heterozygotes show significant growth retardation [45].
3.2. Post-Transcriptional Regulation
One major mechanism by which lncRNAs exert post-transcriptional regulation is by interacting with protein complexes that regulate mRNA splicing and protein turnover. Serine/arginine (SR) splicing factors regulate tissue- and cell-type-specific alternative splicing (AS). MALAT1 is localized in nuclear paraspeckles, where it interacts with SR splicing factors, affects their distribution, and regulates pre-mRNA alternative splicing [46]. The depletion of MALAT1 causes nuclear fragmentation and aberrant mitosis, potentially because of the inclusion of an alternative exon in SAT1 pre-mRNA [46]. Some lncRNAs harbor specific motifs through which they interact with splicing factors; e.g., lncRNA PNCTR has YUCUYY and YYUCUY motifs, through which it interacts with PTBP1, thereby inhibiting the splicing of pre-mRNAs that bear the same motifs [47].
Certain lncRNAs act as competitive endogenous RNAs (ceRNAs) or sponges, sequestering microRNAs (miRNA) and preventing their interaction with target mRNAs. These lncRNAs are abundant and harbor miRNA-complementary sites. ceRNA function has been attributed to numerous lncRNAs, and this regulatory network has been shown to play a role in cancer progression [48]. lncRNA PNUTS have seven binding sites for miR-205, which targets ZEB1 and ZEB2. PNUTS overexpression sponges miR-205, and subsequent increases in ZEB1 and ZEB2 contribute to epithelial–mesenchymal transition (EMT) and invasion by breast cancer cells [49].
3.3. Liquid–Liquid Phase Separation (LLPS)
LLPS inside a cell involves the polymerization of macromolecules, creating unique liquid/fluid phases without any delimiting membrane, similar to an oil droplet in water, which can form or disassemble in seconds, thus quickly and accurately responding to signals and stimuli [50]. Subcellular compartments, such as nucleoli and stress granules, are generated by LLPS, and interaction between RNA-binding proteins (RBPs) containing intrinsically disordered regions (IDRs) and RNAs drives the phase-separation reaction, leading to the formation of ribonucleoprotein (RNP) granules [51].
LncRNAs can function as molecular scaffolds to orchestrate phase separation and facilitate the assembly of membrane-free organelles. lncRNA levels are low and often cancer-specific, with their expression being induced by extracellular cues during cancer development and progression [52]. LncRNAs participate in LLPS via their repetitive sequences, which contribute to molecular crowding and allowing specific cell states and cancer phenotypes [52]. Nuclear paraspeckle, which is rich in RNA and protein, is an example of a membrane-less organelle generated by LLPS, and lncRNA NEAT1 (Nuclear paraspeckle assembly transcript 1) plays an essential role in nuclear paraspeckle formation [53]. NEAT 1 has two transcripts; the longer transcript NEAT1_2 interacts with RBP NONO (non-POU domain-containing octamer binding protein), SFPQ (splicing factor proline and glutamine-rich), and FUS (FUS RNA-binding protein), facilitating paraspeckle formation [54]. Paraspeckles, and hence NEAT1, play a role in the nuclear retention of mRNAs, function as molecular sponges for RBPs, and regulate key physiological and pathological processes, such as corpus luteum formation, and cancer [55,56].
Additional examples of lncRNAs functioning through LLPS are provided in Table 1.
3.4. lncRNAs Coding Micropeptides
Although lncRNAs are by definition ncRNAs, recent studies have shown that some lncRNAs indeed code for functional micropeptides. One of the earliest papers describing this phenomenon was published in 2020, describing the expression of a 60-amino-acid peptide ASRPS from LINC00908, which is downregulated in triple-negative breast cancer (TNBC) [68]. ASRPS binds to STAT3, inhibits STAT3 phosphorylation, and prevents angiogenesis [68]. LINC00665 codes a 52-amino-acid peptide CIP2A-BP, the translation of which is inhibited by TGF-β in TNBC cells [69]. CIP2A-BP inhibits the oncogene CIP2A, thereby inhibiting multiple oncogenic signaling pathways and CIP2A-BP-abrogated lung metastases in the MMTV-PyMT mouse breast cancer model [69]. lncAKR1C2 is an exosomal lncRNA secreted from gastric cancer cells, and it encodes a micropeptide pep-AKR1C2, which is produced in lymphatic endothelial cells and promotes lymph node metastasis by gastric cancer cells by modulating YAP phosphorylation and increased fatty acid β-oxidation [70]. lncRNA AC115619 was shown to code for the micropeptide AC115619-aa, which was downregulated in human HCC and inhibited HCC progression [71]. AC115619-aa interacted with WTAP and inhibited the assembly of N6-methyladenosine (m6A) methyltransferase complex, thereby inhibiting global m6A levels in human HCC cells and affecting the expression of tumor-associated proteins SOCS2 and ATG14 [71]. The number of micropeptides encoded by lncRNAs is growing, especially in the context of cancer. However, for most of these micropeptides, there is a single article describing their detection and function, and in some cases, they are detected in non-human systems [72]. As such, further in-depth validation studies are required to confirm the generalizability of these observations.
4. Significance of Studying lncRNAs in HCC
Studying lncRNAs in HCC holds paramount significance in advancing our understanding of this challenging cancer and improving clinical outcomes. Recent research has underscored the multifaceted roles of lncRNAs in HCC, revealing their potential as critical diagnostic, prognostic, and therapeutic tools (Figure 3).
First and foremost, the identification of lncRNAs as early diagnostic markers for HCC has garnered significant attention. Studies have pinpointed specific lncRNAs, such as HULC (Highly Upregulated in Liver Cancer) and HEIH (hepatocellular carcinoma UpRegulated EZH2-Associated Long Noncoding RNA), with elevated expression in HCC patients. These lncRNAs serve as promising candidates for early biomarkers, allowing for timely intervention and improved patient outcomes [73,74]
LncRNAs have also demonstrated substantial prognostic value in HCC. For instance, HOTAIR, known for its association with poor overall survival and disease-free survival in HCC patients, offers crucial insights for tailoring treatment strategies and predicting patient outcomes [75]. Other lncRNAs, such as H19, have also been linked to HCC progression and poor prognosis, emphasizing the prognostic potential of these molecules [76].
Furthermore, the intricate roles of lncRNAs in HCC pathogenesis have come to light. lncRNAs, including MALAT1, have been implicated in promoting aggressive phenotypes and tumor progression. Additionally, lncRNAs play a pivotal role in epigenetic regulation by modulating DNA methylation and histone modifications, thereby influencing gene expression patterns in HCC cells [77]. Understanding these lncRNA-driven epigenetic mechanisms offers insights into HCC pathogenesis and potential therapeutic interventions.
Importantly, lncRNAs are emerging as potential therapeutic targets in HCC. Recent research has explored innovative strategies to target lncRNAs, including the use of antisense oligonucleotides and small molecules. These approaches hold promise for disrupting critical pathways implicated in HCC development and progression [78]. Additionally, the exploration of lncRNA-directed therapies offers a new dimension to the treatment of this challenging cancer.
5. LncRNAs in NASH and Viral Hepatitis, Risk Factors Contributing to HCC
LncRNAs have emerged as critical players in various liver diseases, including non-alcoholic steatohepatitis (NASH) and viral hepatitis, which are precursors to HCC. A number of lncRNAs have been identified from mouse models of NASH. However, for some of these lncRNAs, the human homologs either do not exist or have not yet been identified. A discussion of the mouse lncRNAs, which lack human relevance, has been omitted from this review.
5.1. NASH
NASH is characterized by hepatic inflammation and fibrosis, often associated with obesity and metabolic syndrome. Recent research has unveiled the regulatory roles of several lncRNAs in NASH pathogenesis. For instance, the lncRNA H19 has garnered attention for its contribution to hepatic fibrosis in NASH. H19 is thought to act as a ceRNA, sponging miRNAs and thereby modulating the expression of genes involved in fibrosis [79]. Through this mechanism, H19 promotes the activation of hepatic stellate cells and the deposition of extracellular matrix components, exacerbating fibrosis progression. MALAT1 was induced in hepatocytes by palmitate treatment as well as in ob/ob mice [80]. It was documented that MALAT1 interacted with SREBP1c mRNA to increase its stability, and in vivo MALAT1 siRNA injection prevented hepatic lipid accumulation and insulin resistance in ob/ob mice [80].
Additionally, lncRNAs like MEG3 have been implicated in NASH-associated oxidative stress and inflammation. MEG3 acts as a molecular sponge for miR-34a, alleviating its inhibitory effect on SIRT1, a key regulator of cellular responses to stress. By enhancing SIRT1 expression, MEG3 attenuates oxidative stress and inflammation in the liver, potentially mitigating NASH development [81].
5.2. Viral Hepatitis
Viral hepatitis, particularly hepatitis B (HBV) and hepatitis C (HCV), is a major risk factor for the development of chronic liver diseases, including cirrhosis and HCC. Emerging evidence suggests that lncRNAs play crucial roles in the host–virus interactions and the progression of viral hepatitis.
The lncRNA HULC has been linked to HBV replication and hepatocarcinogenesis. HULC modulates viral replication by interacting with host factors and promoting the expression of critical viral genes. Furthermore, it contributes to HCC by enhancing the stability of SIRT1 mRNA and inhibiting its degradation by miR-372, leading to increased SIRT1 expression and subsequent tumor growth [81].
In the case of HCV, the lncRNA MALAT1 has been implicated in regulating HCV replication. MALAT1 interacted with HCV-encoded proteins, facilitating viral genome replication [82]. This interaction underscores the intricate interplay between lncRNAs and viral replication processes.
miR-675 is derived from the first exon of H19, and it was shown that H19 and miR-675 were upregulated in HBV-induced chronic hepatitis with the subsequent downregulation of miR-675 target PPARα with associated activation of Akt/mTOR signaling and perturbation of energy metabolism [83].
6. LncRNAs Regulating HCC
A PubMed search performed on 12 December 2023 with the key words lncRNA and HCC revealed 2327 papers, of which 224 were review papers, clearly demonstrating a growing interest in studying lncRNA in hepatocarcinogenesis. However, many of these studies are in vitro using only one cell line; some studies are focused on intrahepatic cholangiocarcinoma, not HCC; and some studies unraveled mouse lncRNAs without identifying the corresponding human homologue. The abundance of lncRNA is an important consideration for determining its mechanism of action and validating whether a particular lncRNA can really modulate a phenotype. Usually, the lncRNA level is low, which might be adequate to modulate the function of a single target-gene locus, especially for cis-acting lncRNAs. However, to sequester a protein or a protein complex or to function as a ceRNA for miRNAs, high levels of lncRNAs are required. It was shown that the lncRNA PNCTR can sequester only ~7.12–27.44% of its target protein PTBP1 in HeLa cells because of the relative abundance of each [47]. However, this level of inhibition was sufficient for PNCTR to regulate cell survival via its regulation of PTBP1 [47]. Given this constraint, stoichiometry analysis, as well as in-depth molecular analysis by multiple techniques, are necessary to determine whether a lncRNA–protein or a lncRNA–miRNA interaction identified by pull-down assays can actually have an effect on biological function and alter a phenotype. The primary mechanism of action for many of the lncRNAs in HCC was shown to be sponging miRNAs, and with the caveat described above, the role of these lncRNAs in HCC requires stringent validation. Here, we highlight those lncRNAs whose function has been interrogated in multiple studies with reproducible results (Table 2), or in the case of a single study, the experiments were performed rigorously using multiple cell lines and in vivo models.
6.1. LncRNAs Functioning as Oncogenes
6.1.1. HOTAIR
HOTAIR is a 2158-nucleotide-long lncRNA that was identified from a custom tilling array of the HOXC locus (12q13.13) [38]. The upregulated expression of HOTAIR in HCC patients correlated with significantly lower cumulative recurrence-free survival [104,105]. As described in Section 3, HOTAIR modulates PRC2, resulting in epigenetic silencing and the transcriptional suppression of target genes [96,106]. Interaction between HOTAIR and EZH2, a component of PRC2, lead to a decrease in miR-218 levels and an increase in its target BMI1, which functions as an oncogene [105]. In HepG2 and Bel7404 cells, HOTAIR knockdown activated p14ARF and p16Ink4a signaling, causing cell cycle arrest and inhibiting the growth in xenograft models [105]. HOTAIR’s competitive binding to RNA helicase DDX5 displaced RNA-binding E3 ligase MEX3B, which stabilized SUZ12, a core subunit of PRC2, and promoted PRC2-mediated gene silencing, especially the PRC2 target genes EpCAM and pleuripotency genes [107]. Liver tumors from HBV X protein (HBx) and c-Myc transgenic mice and chronically HBV-infected patients showed a negative correlation between DDX5 levels, pleuripotency gene expression, and HCC differentiation [107]. According to these findings, a potential role of HOTAIR in the negative regulation of HBV/HCC is suggested, which is contrary to the oncogenic function of HOTAIR and needs further verification.
HOTAIR was shown to regulate exosome production by HepG2 cells by modulating expression and localization of exosomal proteins, such as RAB35, SNAP23, and VAMP3, and through its ability to interact with RAB35 protein [108]. Exosomes are known to promote metastasis. However, it was not studied whether HOTAIR-mediated increase in exosome production accentuates metastasis.
In HCC, HOTAIR disrupted the expression and function of SETD2, influencing histone modifications and DNA repair, which promoted the growth of cancer stem cells (CSCs) [109]. HOTAIR expression is regulated by the transcription factor FOXC1, and it interacts with miR-1, miR-145, miR-122, and RNA-binding motif protein 38 (RBM38) in HepG2, Bel-7402 and Huh7 cells, all contributing to HCC [110,111,112,113].
6.1.2. MALAT1
MALAT1 is a highly conserved nuclear-localized lncRNA transcribed from human chromosome 11q13 as a ~7.5 kb transcript. It is overexpressed in primary tumors and metastases and is prone to copy number changes in several cancer types, including HCC [114]. In HCC patients, it is associated with metastasis and poor prognosis [77].
Nuclear paraspeckles are important structures that retain specific mRNAs in the nucleus [115]. MALAT1 is localized in nuclear paraspeckles, where it interacts with serine/arginine (SR) splicing factors, affects their distribution, and regulates pre-mRNA alternative splicing [46]. In HCC cells, MALAT1 upregulated the splicing factor SRSF1, which caused alternative splicing of RPS6KB1 with subsequent activation of mTORC1, a process that induced the transformation of liver progenitor cells [116]. MALAT1 was shown to activate the Wnt/β-catenin pathway, although the underlying molecular mechanism was not elucidated [116]. MALAT1 was shown to promote glycolysis and suppress gluconeogenesis by increasing the translation of the transcription factor TCF7L2 [117]. However, the mechanism by which MALAT1 increased TCF7L2 translation was not clear because a direct interaction between MALAT1 and TCF7L2 was not examined.
MALAT1 can act as a ceRNA, sequestering miRNAs and preventing them from targeting their downstream mRNA targets. In HCC cells, such as Bel-7402, Hep3B, HepG2, HuH-7, MHCC97, and SMMC-7721, MALAT1 sponges miR-195 causing the activation of EGFR, PI3K/AKT, and JAK/STAT signaling; miR143-3p upregulating ZEB1; miR-146-5p upregulating TNF receptor-associated factor 6 (TRAF6) resulting in activation of AKT; miR-22 increasing snail family transcription factor SNAIL; and miR-30a-5p upregulating vimentin, all of which contribute to augmentation of cell proliferation, invasion, and EMT [118,119,120,121,122].
A recent study unraveled a role of MALAT1 in mitochondria of HCC cells, such as HepG2 and HL7702, where it interacted with multiple loci in mitochondrial DNA (mtDNA), such as D-loop, COX2, ND3, and CYTB genes [123]. Knocking down MALAT1 perturbed mitochondrial transcription, resulting in the inhibition of mitochondrial functions, such as oxidative phosphorylation and ATP production, a decrease in mitophagy, and an increase in apoptosis [123]. It was shown that the translocation of MALAT1 from the nucleus to the mitochondria is mediated by RNA-shuttle protein HuR and mitochondrial membrane protein MTCH2 [123].
6.1.3. HULC
HULC, a 482 bp transcript encoded by a gene in chromosome 6p24.3, is a well-studied lncRNA that has garnered considerable attention in the context of HCC. HULC was first discovered as a highly upregulated transcript in HCC, and its aberrant expression has been associated with HCC development and progression [89]. One of its primary functions is to act as a molecular sponge for miRNAs, such as miR-2001-3p, miR-186, and miR-107, resulting in an increase in the oncogenes ZEB1 (contributing to EMT), HMGA2, and E2F1, respectively [124,125]. Transcription factor E2F1 regulates sphingosine kinase 1 (SPHK1), which is known to promote angiogenesis, and a contribution of HULC/E2F1/SPHK1 axis in augmenting tumor angiogenesis was shown [125]. Protective autophagy contributes to tumorigenesis, and SIRT1 has been identified to positively regulate this process. In HepG2 and Hep3B cells, HULC downregulates miR-6825-5p, miR-6845-5p, and miR-6886-3p, resulting in the upregulation of their target ubiquitin-specific peptidase 22 (USP22) and the inhibition of ubiquitin-mediated degradation of SIRT1 [81]. This mechanism contributed to HULC- and SIRT1-mediated protective autophagy [81]. HULC decreased miR-15a in Hep3B cells, which resulted in the inhibition of PTEN via P62, resulting in the activation of the oncogenic PI3K-AKT-mTOR pathway [126].
HULC is involved in regulating HBV-induced HCC. The HBX protein activated the HULC promoter via CREB, and the induced HULC inhibited the promoter of the tumor suppressor eukaryotic translation elongation factor 1 epsilon 1 (EEF1E1/P18), thus reducing its expression and facilitating HBX-augmented proliferation both in vitro and in vivo [127]. However, the mechanism by which HULC regulates EEF1E1 expression was not explored. Increased lipid production fuels tumor growth by providing a source of energy. In HepG2 and HuH-7 cells, HULC inhibited the miR-9 promoter by inducing CpG island methylation, leading to an increase in miR-9 target peroxisome-proliferator-activated receptor alpha (PPARA) and subsequently PPARA target acyl-CoA synthetase subunit ACSL1, which resulted in increased cholesterol production and stimulation of cell proliferation [128]. Conversely, exogenous cholesterol augmented the HULC promoter activity of retinoid x receptor (RXRA), thus establishing a positive feedback loop. HULC increased the expression of the circadian rhythm regulating gene CLOCK by interacting with 5′-UTR of the CLOCK gene, which was suggested to contribute to the in vivo growth of HepG2 cells [129]. Thus, HULC augments a variety of oncogenic pathways engaging diverse mechanisms to promote HCC.
6.1.4. PVT1
Chromosome 8q amplification is a frequent event in many cancers, including HCC. PVT1 (Plasmacytoma Variant Translocation 1) is located on chromosome 8q24 adjacent to MYC (encoding c-MYC), and MYC and PVT1 genes are co-amplified in multiple cancer patients, including a subset of HCC patients [130]. PVT1 is frequently upregulated in HCC tissues and cell lines, and its overexpression has been associated with aggressive clinicopathological features and poor prognosis in HCC patients [131,132]. It was demonstrated that PVT1 promotes cell proliferation and cancer stem-cell-like properties by binding to and stabilizing the RNA-binding protein NOP2 [132]. PVT1 functions as ceRNA for multiple miRNAs, thereby increasing their targets, such as miR-365 and ATG3, to promote autophagy, and miR-150 and hypoxia-inducible protein 2 (HIG2) to modulate iron metabolism and cell proliferation [133,134]. PVT1 has been shown to inhibit interferon-α-induced apoptosis in SMMC-7721 HCC cells by interacting with the signal transducer and activator of transcription 1 (STAT1) [135].
6.1.5. HOTTIP
HOTTIP (HOXA Transcript at the Distal Tip) is a 7.9 kb lncRNA located in chromosome 7p15 that is frequently upregulated in HCC tissues and cell lines, and its overexpression is correlated with aggressive clinicopathological features, including advanced tumor stages and poorer prognosis for HCC patients [36,95]. As described in Section 3, HOTTIP activates HOXA genes, which promote HCC cell proliferation, migration, and invasion [36,95]. HOTTIP itself is targeted by tumor suppressor miRNAs, miR-192, and miRNA-240, and this regulation was shown to interfere with glutaminolysis by glutaminase GLS1 in HepG2 and SMMC7721 cells [136]. Glutaminolysis is a hallmark of cancer cells, and it was suggested that HOTTIP might promote HCC by regulating glutamine metabolism. LncRNA PAARH functions as an oncogene in HCC and was shown to mediate its effect by sponging multiple miRNAs, such as miR-6760-5p, miR-6512-3p, miR-1298-5p, miR-6720-5p, miR-4516, and miR-6782-5p, resulting in the upregulation of HOTTIP [137]. Thus, there are a number of indirect mechanisms by which HOTTIP might be upregulated, facilitating HCC development and progression.
6.1.6. H19
H19 is located in chromosome 11p15 and is associated with genomic imprinting. The genomic location of H19 is near the insulin-like growth factor 2 (IGF2) gene; while IGF2 is expressed only from the paternally inherited chromosome, H19 is expressed from the maternally inherited chromosome [138]. H19 is a 2.5-kilobase-long RNA molecule that undergoes splicing and polyadenylation, but it does not contain the information to produce a protein. H19 is highly conserved and is expressed in various cell lineages during mammalian development.
Whether H19 functions as an oncogene or tumor suppressor gene is a debatable issue in other cancers, but in HCC, the current literature suggests it as an oncogene [76]. H19 is frequently upregulated in HCC tissues and cell lines, and its elevated expression has been linked to poor clinical outcomes in HCC patients [139]. One of the key mechanisms through which H19 promotes HCC progression is by sponging miR-193b with resultant upregulation of MAPK1, promoting EMT and the transformation of stem cells. In HepG2 cells, H19 expression was induced by tumor-associated macrophages, suggesting that inflammation might regulate H19 expression [139]. H19 was upregulated in HCC tumors generated upon the deletion of Transforming growth factor-β receptor 2 (TGFBR2) via the activation of the transcription factor SOX2, and H19 knockdown inhibited TGFBR2-deletion-induced HCC [140]. However, the H19 targets that mediate this effect were not identified in this study. A potential role of H19 in promoting bone metastasis of HCC has been documented, in which H19 interacted with protein phosphatase 1 catalytic subunit alpha (PPP1CA), which dephosphorylated p38 MAPK with subsequent downregulation of osteoclastogenesis inhibitory factor osteoprotegerin (OPG) [141]. This resulted in the activation of osteoclastogenesis and hence osteolytic bone lesions, a feature of HCC bone metastasis. Additionally, H19 sponged miRNA-200b-3p, resulting in the enhanced expression of ZEB1, which caused increased cell migration and invasion [141].
6.1.7. NEAT1
The nuclear paraspeckle assembly transcript 1 (NEAT1) gene is located in chromosome 11q13.1 and gives rise to two transcripts, a 3.7 kb NEAT1v1 and a 23 kb NEAT1v2, the latter of which is necessary for the formation of nuclear paraspeckles [53,142]. NEAT1, along with MALAT1, was also shown to bind to active chromatin sites, identified by the procedure Capture Hybridization Analysis of RNA Targets (CHART), suggesting a potential role in regulating transcription [143].
NEAT1 functions as an oncogene in many cancers, including HCC, in which it is overexpressed [144,145,146,147]. In Hep3B and HepG2 cells, the inhibition of NEAT1 attenuates proliferation, migration, and invasion, and NEAT1 functions by sponging multiple miRNAs and increasing the levels of the targets of these miRNAs, such as miR-485 and STAT3, miR-204 and ATG3 (which increases autophagy), and miR-139-5p and TGF-β1 [145,146,148].
A recent study, however, unravels an opposite role of NEAT1, mainly its long isoform NEAT1v2/NEAT1_2, which is regulating cancer cell metabolism [149]. Cancer cells use aerobic glycolysis for its survival, a phenomenon known as the “Warburg Effect”. It was shown that in HCC cells, mTOR, which promotes THE Warburg Effect, negatively regulates NEAT1 transcription, thus suppressing paraspeckle biogenesis and liberating the RNA-binding proteins NONO and SFPQ [149]. These RNA-binding proteins bind to U5 in spliceosomes and promote mRNA splicing and the expression of glycolytic enzymes, thus stimulating aerobic glycolysis and HCC growth. The mTOR inhibitor Rapamycin was shown to exert its anti-cancer effect by perturbing this mTOR/NEAT1-mediated mechanism [149]. The authors also showed that NEAT1v2/NEAT1_2 was downregulated in HCC patients, which correlated with poor overall survival [149]. This study underlies the importance of analyzing transcript variants of lncRNAs and checking their functions in the context of cancer.
6.1.8. HEIH
HEIH (hepatocellular carcinoma upregulated EZH2-associated long noncoding RNA), located in chromosome 5q35, is a 1.7 kb transcript that is overexpressed in HCC patients and negatively correlates with cumulative survival [150]. HEIH interacts with EZH2, resulting in increased binding of EZH2 to and H3K27 trimethylation of p16 promoter, downregulating the expression of this tumor suppressor [150]. The overexpression and knockdown of HEIH stimulated and inhibited, respectively, xenograft growths of Hep3B, HepG2, and HuH-7 cells [150]. HEIH was detected in serum and exosome and was suggested as a potential biomarker of HCV-associated HCC [151]. HEIH was shown to contribute to sorafenib resistance by sponging miR-98-5p and activating the PI3K/Akt pathway [152].
6.1.9. SNHG6
SNHG6 (small nucleolar RNA host gene 6) is another lncRNA that is located in chromosome 8q13.1, thus being amplified in many cancers, including HCC [153]. There are five transcripts of SNHG6 (SNHG6-003 to SNHG6-007), among which SNHG6-003 and SNHG6-006 were shown to be highly expressed in 52 HCC patients [153]. Mechanistically, it was shown that SNHG6-003 promoted HCC by sponging miR-26a/b and increasing the expression of its target transforming growth factor-β-activated kinase 1 (TAK1) [153]. In HCC cells, SNHG6 functions as ceRNA for multiple miRNAs inducing their targets, such as miR-101-3p and ZEB1, miR-139-5p and SERPINH1, let-7c-5p and MYC, and miR-6509-5p and HIF1A, all contributing to hepatocarcinogenesis [154,155,156,157].
A recent study linked SNHG6 in the progression of NAFLD to HCC by modulating cholesterol metabolism [158]. SNHG6 expression was induced by cholesterol treatment in HepG2 cells, and SNHG6 facilitated cholesterol-mediated interaction of ER-anchored FAF2 (Fas-associated factor family member 2) with mTORC1 at lysosomes promoting mTORC1-mediated increase in cellular cholesterol biosynthesis, thus establishing a positive feedback loop [158]. Snhg6 expression was induced in a high-fat/high-cholesterol (HF/HC) and diethylnitrosamine (DEN) model of mouse NAFLD/HCC; adeno-associated virus (AAV8)-mediated delivery of SNHG6 accelerated the process, and AAV8 delivering shRNA for SNHG6 reversed the process [158].
6.1.10. Additional lncRNAs Functioning as Oncogenes
The oncogenic function of several lncRNAs in HCC has been described in a single albeit detailed study, which requires additional validation and/or characterization. Here, we highlight some of these lncRNAs affecting specific aspects of HCC development and progression.
LncRNAs Modulating Wnt/β-Catenin Signaling
LncRNA UFC1 was identified to be overexpressed in HCC tissues, and in vitro and in vivo studies unraveled a tumor-promoting role of UFC1 [159]. Mechanistically, UFC1 was shown to interact with and stabilize HuR mRNA with a resultant increase in β-catenin [159]. lncRNA for β-catenin methylation (lnc-β-Catm) was upregulated in liver cancer stem cells (CSCs) and induced the methylation of β-catenin by associating with β-catenin and methyltransferase EZH2 [160]. The methylation of β-catenin protected it from ubiquitination and increased its stability, leading to the activation of Wnt/β-catenin signaling that promoted the self-renewal of liver CSCs [160]. A novel mechanism of lncRNA action was proposed for DANCR (differentiation antagonizing non-protein coding RNA), in which it interacted with β-catenin protein and prevented β-catenin degradation by miR-214, miR-320a, and miR-199a, which resulted in increased stem cell line features in HCC cells [161].
LncRNAs Promoting HCC by Regulating Metabolism
LINC01234, overexpressed in HCC and correlated with poor prognosis, promoted HCC cell proliferation, migration, and drug resistance in vitro and in vivo [162]. It was shown that LINC01234 interacted with the promoter region of arginosuccinate synthase 1 (ASS1), inhibiting its transcription, especially by p53, thus modulating aspartate metabolism [162]. lncRNA RP11-386G11.10, transcriptionally regulated by ZBTB7A, functions as a ceRNA for miR-345-3p, increasing the expression of HNRNPU and its downstream lipogenic enzymes, with resultant accumulation of lipids in HCC cells and the promotion of metastasis and primary tumor growth [163]. A search for RNA-binding proteins (RBPs) regulating HCC identified that RBP CCT3 regulated lipid metabolism in HCC cells by regulating the lncRNA LINC00326, and interference with CCT3/LINC00326 interaction inhibited the growth of HuH-7 tumors implanted in zebrafish [164]. Taurine upregulated gene 1 (TUG1) interacted with PRC2 complex and epigenetically silenced a number of tumor suppressor genes, including p21. In HCC cells, TUG1-mediated downregulation of p21 resulted in upregulation of miR-455-3p with the resultant downregulation of its target AMPKβ2 (adenosine monophosphate-activated protein kinase subunit beta 2) [165]. This resulted in the activation of the mTOR pathway and the induction of hexokinase 2 (HK2), contributing to increased glycolysis, cell growth, and metastasis [165]. TUG1 was also shown to epigenetically silence the transcription factor KLF2, a tumor suppressor, in HCC cells [166]. Similar epigenetic silencing of KLF2 was also shown for lncRNA CDKN2B antisense RNA 1/ANRIL [167].
lncRNAs Regulating EMT and Metastasis
LncRNA-activated by TGF-β (lncRNA-ATB), overexpressed in HCC metastases and associated with poor prognosis, sponged miR-200 family, thereby upregulating ZEB1 and ZEB1 and inducing EMT [168]. Additionally, lncRNA-ATB bound to IL-11 mRNA, increasing its expression and activating STAT3, which facilitated the colonization of metastatic cells to distant organs [168]. lncRNA PRR34-AS1 sponged miRNA-296-5p to increase transcription factors E2F2 and SRY-box transcription factor 12 (SOX12) in HCC cells, activating Wnt/β-catenin pathway and promoting EMT [169]. It was also shown that HNRNPU augmented ZBTB7A expression, thus establishing a positive feedback loop [163]. lncRNA ZFAS1 (ZNFX1 antisense RNA1) levels correlated with intra- and extrahepatic HCC metastasis and poor prognosis, and it promoted HCC progression by squelching miRNA-150 with resultant upregulation of ZEB1 and matrix metalloproteinases MMP14 and MMP16 [170]. Extrahepatic metastatic tissues in HCC patients showed increased alternative splicing, which was associated with increased expression of DEAD-box RNA helicase 17 (DDX17) [171]. It was shown that DDX17 induced the retention of intron 3 of the lncRNA Paxillin antisense RNA 1 (PXN-AS1), creating a new transcript PXN-AS1-IR3 [171]. PXN-AS1-IR3 stimulated HCC metastasis by recruiting TEX10 (testis expressed 10) and p300 to MYC enhancer region and upregulating MYC levels. PXN-AS1 was increased in the serum of HCC patients with extrahepatic metastasis potentially serving as a blood biomarker [171]. In another study, splicing factor MBNL3 was shown to induce exon 4 inclusion of PXN-AS1, and this new transcript interacted with PXN mRNA, protecting it from miR-24-mediated degradation and stimulating HCC progression [172].
lncRNAs Regulating Vascular Invasion
LncRNA MVIH (lncRNA associated with microvascular invasion in HCC) was shown to be associated with increased microvascular invasion and metastasis and decreased recurrence-free survival in 215 HCC patients [173]. MVIH promoted angiogenesis and metastasis by increasing the secretion of phosphoglycerate kinase 1 (PGK1) [173]. lncRNA PAARH exerts its HCC-promoting effect in multiple ways, such as upregulation of HOTTIP by squelching multiple miRNAs and binding to HIF-1α and facilitating its recruitment to VEGF promoter, thereby increasing microvessel density and promoting angiogenesis and metastasis [137]. A major complication of HCC is portal vein tumor thrombus (PVTT), and it was shown that lncRNA ICR (ICAM-1-related) was expressed in Intercellular adhesion molecule 1 (ICAM-1)-positive cancer stem cells (CSCs) regulating CSC function and leading to PVTT development [174]. ICR was regulated by the stem cell transcription factor Nanog, and it positively regulated ICAM-1 levels by binding to ICAM-1 mRNA [174].
lncRNAs Regulating Oncogenic AKT Pathway
LncRNA cancer susceptibility 9 (CASC9) interacted with the RNA-binding protein heterogenous nuclear ribonucleoprotein L (HNRNPL), resulting in the activation of the AKT signaling pathway and contributing to increased tumorigenesis by HCC cells [175]. lncRNA RP11-295G20.2 interacted with the N-terminus of PTEN, facilitating its interaction with p62, translocation to lysosomes, and degradation, which led to the activation of the oncogenic AKT pathway [176].
Miscellaneous Functions of lncRNAs in HCC
The inhibition of the novel lncRNA NIHCOLE (noncoding RNA induced in hepatocellular carcinoma with an oncogenic role in ligation efficiency) induced apoptosis in multiple HCC cells with associated DNA damage because of a decrease in the nonhomologous end-joining (NHEJ) pathway of DNA double-strand breaks [177]. NIHCOLE was shown to associate with several NHEJ factors, including Ku70/Ku80 heterodimer, and promoted ligation efficiency [177]. CDIP transferase opposite strand, pseudogene (CDIPTOSP), also known as lnc-CTHCC, is a cancer testis gene that is highly expressed in the testis and HCC [178]. An lnc-CTHCC knockout mouse was protected from HCC development [178]. Lnc-CTHCC interacted with heterogenous ribonucleoprotein K (hnRNPK), recruiting it to the YAP1 promoter and activating its transcription [178]. Pluripotency and hepatocyte-associated RNA overexpressed in HCC (PHAROH) is a lncRNA that interacted with and sequestered the translation repressor TIAR, leading to increased translation of MYC, thereby promoting HCC [179].
6.2. lncRNAs Functioning as Tumor-Suppressor Genes
6.2.1. MEG3
Maternally Expressed Gene 3 (MEG3) is located on chromosome 14q32. It is a ~1.6 kb transcript and is known for its tumor-suppressive functions [180]. MEG3 is downregulated in HCC tissues and cell lines, especially by the methylation of its promoter by DNA methyltransferases DNMT1 and DNMT2, and its reduced expression is associated with poor prognosis in HCC patients [103,181]. MEG3 upregulated p53 target genes by directly interacting with p53 DNA binding domain, and the overexpression of MEG3 in HepG2 and HuH-7 cells induced apoptosis [103,180]. MEG3 functions as ceRNA for many miRNAs, such as miRNA-664, but the functional implication of these findings needs further validation [103]. In HepG2 cells, MEG3 was shown to inhibit miR-10a-5p, which targets PTEN [182]. MEG3 overexpression induced PTEN and inhibited AKT signaling, and upregulated pro-apoptotic protein Bax and downregulated anti-apoptotic protein Bcl-2 [182]. An MS2 bacteriophage virus-like particles (VLPs) crosslinked with GE11 polypeptide was used to deliver MEG3 systemically, which inhibited xenografts of EGFR-positive HepG2 cells [183].
6.2.2. GAS5
GAS5 (Growth Arrest-Specific 5), located in chromosome 1q25, is known for its role in regulating cell growth and apoptosis and is frequently dysregulated in cancers, including HCC, where its expression levels inversely correlate with patient survival [144,184]. The overexpression of GAS5 in HCC cells inhibited proliferation and invasion and inhibited vimentin via an unknown mechanism [184]. GAS5 levels are higher in sorafenib-resistant HCC cells and RNA-binding protein RBM38 was shown to bind to and stabilize GAS5 in these cells [185]. In HepG2 cells, GAS5 bound to glucose-regulated protein GRP78, and it induced apoptosis by activating the ER stress-signaling pathway [186]. In HepG2 and HuH-7 cells, GAS5 improved cisplatin sensitivity by sponging miR-222 [187].
6.2.3. FENDRR
FOXF1-adjacent noncoding developmental regulatory RNA (FENDRR), located in chromosome 16q24, is downregulated in HCC and inhibits the growth of Hep3B and HepG2 cells in vivo [188]. It interacted with PRC2 and TrxG/MLL complexes, thus mediating the epigenetic silencing of gene expression [189]. FENDRR interacted with the promoter of Glypican-3 (GPC3), an HCC marker, and caused the methylation-induced silencing of GPC3 expression [188]. In MHCC97 cells, FENDRR sponged miR-423-5p, resulting in increased expression of its target growth arrest and DNA damage-inducible beta (GADD45B), leading to the suppression of in vivo tumorigenicity [190]. FENDRR has been suggested to regulate Tregs and immune escape, but these observations need further validation [190].
6.2.4. Additional lncRNAs Functioning as Tumor Suppressors
Downregulated in liver cancer stem cells (DILC) is an ~2.4 kb lncRNA encoded by a gene located in chromosome 13q34 [191]. It is downregulated in liver CSCs, and its knockdown promotes in vivo tumorigenicity by liver CSCs [191]. DILC interacted with the IL-6 promoter, blocking NF-κB-mediated oncogenic IL-6/STAT3 signaling [191]. A recurrent deletion of lncRNA-PRAL (p53 regulation-associated lncRNA), located in chromosome 17p13.1, was identified in HCC patients [192]. The overexpression of lncRNA-PRAL induced apoptosis in HCCLM3 and SMMC-7721 HCC cells but not in Hep3B (p53-deficient) and HuH-7 (p53-mutant) cells, indicating that the tumor suppressor function of lncRNA-PRAL is mediated by p53 [192]. Three stem-loop motifs in the 5′ end of lncRNA-PRAL facilitated the interaction of HSP90 and p53, thereby precluding MDM2-mediated ubiquitination of p53 and increasing p53 stability [192]. The EZH2 transcript variant including exon 14 promotes metastasis. lncRNA LINC01348 was identified to be downregulated in HCC, and overexpression of LINC01348 inhibited in vivo metastasis by SK-Hep1 cells [193]. LINC01348 interacted with splicing factor 3b subunit 3 (SF3B3), interfering with exon 14’s inclusion of EZH2 and downregulating the expression of Snail [193]. lncRNA uc.134, downregulated in HCC, has been shown to interact with CUL4A, thus inhibiting ubiquitination of LATS1 and thereby silencing YAP [194]. lncRNA FTX, transcribed from X chromosome inactivation center, is downregulated in HCC patients and inhibits in vivo metastasis [195]. It sponges miR-374a, which facilitates the upregulation of genes that negatively regulate the Wnt/β-catenin pathway and binds to DNA replication factor MCM2, thereby inhibiting DNA replication [195].
7. lncRNAs Modulating Current HCC Treatment
For advanced HCC, the first line of treatment is a combination of immunotherapy and tyrosine kinase inhibitors (TKIs) such as sorafenib and lenvatinib. A number of in silico studies have identified potential lncRNA prognostic signatures determining responsiveness to TKIs and immunotherapy. Here, we highlight some of the lncRNAs that have been validated using in vivo studies to influence responsiveness and/or resistance to TKIs and modulate immunotherapy response.
Translation regulatory lncRNA 1 (TRERNA1) was upregulated by HBx in HCC cells that sponged miR-22-3p, resulting in the upregulation of NRAS and the activation of downstream RAF/MEK/ERK signaling, thereby contributing to sorafenib resistance [196]. A potential role of MALAT1 in sorafenib resistance, by sponging miR-140-5p and increasing its target Aurora-A kinase, has been described [197]. In HCC cells, the oncogene FOXM1 transcriptionally activated LINC-ROR (long intergenic non-protein coding RNA, regulator of reprogramming), which in turn increased FOXM1 levels by sponging miR-876-5p, thus establishing a positive feedback loop [198]. The upregulation of both FOXM1 and LINC-ROR was shown to confer resistance to sorafenib [198]. Sorafenib-resistant Hep3B and HuH-7 cells showed the enrichment of genes related to stemness and EMT, and single-cell RNA-sequencing in sorafenib-resistant HuH-7 cells identified the lncRNA ZFAS1 (ZNFX1 antisense RNA 1) as the highest upregulated transcript, which positively correlated with multiple stemness and EMT-associated genes in HCC patients [199]. Knocking down ZFAS1 by siRNA in sorafenib-resistant cells restored their sorafenib sensitivity [199]. However, the mechanism by which ZFAS1 regulates stemness genes was not explored in this study. lncRNA linc-VLDLR was shown to be secreted in the extracellular vehicles (EVs) by HepG2 and MzChA1 cells [200]. The treatment of human HCC cells with drugs, such as sorafenib, camptothecin, and doxorubicin, induced linc-VLDLR release in EVs, while incubation with the EVs protected cells from chemotherapy-induced cell death [200]. The knockdown of linc-VLDLR downregulated the drug transporter ABCG2 and ABCG2 overexpression rescued the effects of linc-VLDLR-mediated knockdown on increased sorafenib sensitivity [200]. The mechanism by which linc-VLDLR regulates ABCG2 remains to be explored. PHF8 is a histone lysine demethylase that functions as a transcriptional activator. lncRNA BBOX1-AS1 is upregulated in HCC patients, and it upregulated PHF8 by sponging miR-361-3p, which contributed to HCC progression, autophagy, and sorafenib resistance [201]. The mechanism by which PHF8 regulates these phenotypes remains to be determined. Sorafenib is known to induce ferroptosis in HCC cells. In sorafenib-resistant HCC cells, HIF1α induced the expression of lncRNA URB1-antisense RNA 1 (URB1-AS1), which contributed to ferroptosis via the phase separation of ferritin, and silencing URB1-AS1 restored sensitivity to sorafenib in an in vivo model [202]. A systematic bioinformatics analysis identified lncRNA LINC01132 as a potential oncogene for HCC [203]. It was shown that LINC01132 interacted with the transcription factor NRF1, leading to the induction of T cell activation antigen CD26/DPP4 [203]. Knocking down LINC01132 induced infiltration of CD8+ T cells and augmented the therapeutic efficacy of anti-PDL1 antibody in Hep1-6 cells, thereby suggesting a potential role of LINC01132 in regulating response to immunotherapy [203].
8. Conclusions and Future Perspectives
Through the exploration of various studies and research articles, it is evident that lncRNAs play crucial roles in the initiation, progression, and prognosis of HCC. These molecules have been found to be dysregulated in HCC, acting as both oncogenes and tumor suppressors, and their dysregulation can contribute to the altered expression of essential genes and signaling pathways involved in HCC development and pathogenesis. Furthermore, lncRNAs have exhibited potential diagnostic and prognostic value; e.g., circulating levels of lncRNA SCARNA10 (small cajal-body-specific RNA 10) have been shown to be higher in HBV- or HCV-positive HCC patients with positive correlation with tumor size, tumor stage, vascular invasion, and metastasis, highlighting their significance in clinical practice [204]. By analyzing the plasma or serum of cohorts of patients, several lncRNAs, including MALAT-1, HULC, and HOTAIR, have been implicated as biomarkers for HCC diagnosis but no comprehensive clinical trials are actively exploring these possibilities [205]. Therefore, further investigations are warranted to fully unravel the diverse mechanisms underlying lncRNAs’ involvement in HCC and to explore their therapeutic potential. Many HCC cell lines, such as BEL7402, SMMC7721, MHCC97L, BEL7404, QGY7701, QGY7703, QSG7701, and SKHEP1, used for lncRNA studies, have been found to be either contaminated or not of HCC origin [206]. Therefore, findings obtained from these cell lines must be further validated in additional cell lines. Overall, the body of knowledge garnered so far enhances our understanding of HCC biology and has promising implications for the development of innovative diagnostic methods and targeted therapies in the future. While 83.9% of human mRNAs are orthologous with mouse mRNAs, only 25% of human lncRNAs have mouse orthologs [207]. As such, studying the role of lncRNAs in HCC using mouse models is not always feasible. With ongoing advancements in high-throughput sequencing technologies and bioinformatics, the comprehensive understanding of lncRNAs in HCC is becoming more malleable. Thus, harnessing the potential of lncRNAs as both regulators and therapeutic targets in HCC holds immense promise in improving patient outcomes and revolutionizing the field of HCC and treatment.
Ongoing pre-clinical research explores the possibility of targeting lncRNAs using antisense oligonucleotides (ASOs), CRISPR-based strategies, and RNAi-based therapies. CRISPR and RNAi approaches have been tested mainly at the cell line level, where an oncogenic lncRNA was knocked out or knocked down by siRNA or shRNA, and then the tumorigenic potential of the cell line was checked in an in vivo model [88]. A recent in vivo genome-wide CRISPR activation screening using xenografts of MHCC-97H cells identified many HCC-promoting lncRNAs, and the clinical relevance of these lncRNAs to human HCC was validated [208]. This discovery approach can also be used therapeutically, where multiple HCC-promoting lncRNAs can be simultaneously knocked out with CRISPR/Cas9. Therapeutically, ASOs, which are short single-stranded DNAs, serve as a promising approach because of their ability to specifically hybridize with their target lncRNAs, thus forming DNA–RNA complexes, which can be recognized and degraded by RNase H. The intra-tumoral injection of ASO for MALAT1 significantly inhibited the growth of Hep3B and HuH-7 xenografts in nude mice [209]. HAND2-AS1 is a lncRNA shown to promote liver CSCs, and HAND-AS1 ASO could significantly inhibit the growth of HuH-7 and human patient-derived xenografts (PDX) in NOD-Prkdcscid Il2rgtm1/Bcgen (B-NSG) mice [210]. On the other hand, PRAL is a tumor suppressor lncRNA, and overexpression of PRAL via an adenovirus inhibited xenograft growth of SMMC-7221 cells in nude mice [192]. These therapeutic approaches have been tested using subcutaneous xenografts in immunocompromised mice and intra-tumoral injection. Although these studies demonstrate a preliminary proof of principle, they do not determine whether the intravenous delivery of these approaches works in tumors in the liver and how an intact immune system responds to them. ASOs and similar oligonucleotide-based approaches have the potential to activate an interferon response, and viruses have the potential to develop neutralizing antibodies, thereby neutralizing their efficacy [211]. A potential strategy to overcome these drawbacks is to use targeted nanoparticles to deliver the payload. One example is the delivery of siRNA for the oncogene Astrocyte elevated gene-1 (AEG-1) via hepatocyte-targeted nanoparticles in an orthotopic model of HCC, as well as in a high fat diet-induced model of NASH showing marked therapeutic efficacy [212,213]. Similar strategies might be used for lncRNAs as well. As yet, no lncRNA-targeting therapeutics have entered clinical development, and more in-depth pre-clinical studies are required to bring the lncRNA-targeting approaches to the clinical arena. The field of lncRNA therapeutics is evolving rapidly, offering promising avenues for precision medicine [205]. In conclusion, the significance of lncRNAs as key regulators and potential targets in HCC is undeniably profound.
Author Contributions
Writing—original draft preparation, M.S.; writing—review and editing, D.S.; supervision, D.S.; project administration, D.S.; funding acquisition, D.S. All authors have read and agreed to the published version of the manuscript.
Funding
Present study was supported in part by The National Cancer Institute (NCI) Grants R01CA230561 (DS), R01CA240004 (DS), and R01CA244993 (DS).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| 53BP1 | p53 binding protein 1 |
| AAV | Adeno-associated virus |
| ABCG2 | ATP binding cassette subfamily G member 2 |
| ACSL1 | Acyl-CoA synthetase long-chain family member 1 |
| AEG-1 | Astrocyte elevated gene-1 |
| AFP | Alpha-fetoprotein |
| AKR1C2 | Aldo-keto reductase family 1 member C2 |
| AKT | Akt kinase |
| ALS | Amyotrophic lateral sclerosis |
| AMPKβ2 | Adenosine monophosphate-activated protein kinase subunit beta 2 |
| ANRIL/CDKN2B-AS1 | Antisense noncoding RNA in the INK4 locus/CDKN2B antisense RNA 1 |
| ANXA8 | Annexin A8 |
| APC | APC regulator of WNT signaling pathway |
| ASO | Antisense oligonucleotide |
| ASRPS | A small regulatory peptide of STAT3 |
| ASS1 | Arginosuccinate synthase 1 |
| ATG3 | Autophagy related 3 |
| ATG14 | Autophagy related 14 |
| ATP | Adenosine triphosphate |
| BBOX1-AS1 | BBOX1 antisense RNA 1 |
| BMI1 | BMI1 proto-oncogene, polycomb ring finer |
| B-NSG | NOD-Prkdcscid Il2rgtm1/Bcgen |
| BRD3 | Bromodomain containing 3 |
| CAPRIN1 | Cell cycle associated protein 1 |
| CASC9 | Cancer susceptibility 9 |
| CBX7 | Chromobox 7 |
| CCT3 | Chaperonin containing TCP1 subunit 3 |
| CDIPTOSP | CDIP transferase opposite strand, pseudogene |
| CDKN2A | Cyclin-dependent kinase inhibitor 2A (p16) |
| CDKN2B | Cyclin-dependent kinase inhibitor 2B (p15) |
| CELF1 | CUGBP Elav-like family member 1 |
| ceRNA | Competitive endogenous RNA |
| CHASERR | CHD2 adjacent suppressive regulatory RNA |
| CHD2 | Chromodomain helicase DNA binding protein 2 |
| CIP2A-BP | CIP2A binding peptide |
| CLOCK | Clock circadian regulator |
| COX2 | Cytochrome c oxidase subunit II |
| CREB | cAMP-response element binding protein |
| CSC | Cancer stem cells |
| CT | Computed tomography |
| ctDNA | Circulating tumor deoxyribonucleic acid |
| CRISPR | Clustered regularly interspaced short palindromic repeats |
| CTNNB1 | Catenin beta 1 (β-catenin) |
| CUL4A | Cullin 4A |
| CYTB | Cytochrome b |
| DANCR | Differentiation antagonizing non-protein coding RNA |
| DDR | DNA damage response |
| DDX5 | DEAD-box helicase 5 |
| DDX17 | DEAD-box RNA helicase 17 |
| DDX21 | DExD-box helicase 21 |
| DEN | Diethylnitrosamine |
| DFC | Dense fibrillar component |
| DIGIT | Divergent to GSC, induced by TGF-β family signaling |
| DILC | Downregulated in liver cancer stem cells |
| dilncRNA | Damage-induced lncRNA |
| DNA | Deoxyribonucleic acid |
| DNMT1 | DNA methyltransferase 1 |
| DNMT2 | DNA methyltransferase 2 |
| DPP4 | Dipeptidyl peptidase 4 |
| DSB | Double-strand break repair |
| E2F1 | E2F transcription factor 1 |
| EEF1E1 | Eukaryotic translation elongation factor 1 epsilon 1 |
| EGFR | Epidermal growth factor receptor |
| EMT | Epithelial-mesenchymal transition |
| EPCAM | Epithelial cell adhesion molecule |
| ER | Endoplasmic reticulum |
| ESC | Embryonic stem cells |
| EV | Extracellular vesicle |
| EZH2 | Enhancer of zeste 2 polycomb repressive complex 2 subunit |
| FAF2 | Fas associated factor family member 2 |
| FC | Fibrillar center |
| FDA | Food and drug administration |
| FENDRR | FOXF1 adjacent noncoding developmental regulatory RNA |
| FOXC1 | Forkhead box C1 |
| FOXM1 | Forkhead box M1 |
| FUS | FUS RNA-binding protein |
| GADD45A | Growth arrest and DNA damage-inducible alpha |
| GADD45B | Growth arrest and DNA damage-inducible beta |
| GAS5 | Growth arrest specific 5 |
| GIRGL | glutamine insufficiency regulator of glutaminase lncRNA |
| GLS1 | Glutaminase 1 |
| GPC3 | Glypican-3 |
| GR | Glucocorticoid receptor |
| GRP78 | Glucose regulated protein 78 |
| H19 | H19 imprinted maternally expressed transcript |
| H3K4me3 | Trimethylation of histone H3 at lysine 4 |
| H3K27me3 | Trimethylation of histone H3 at lysine 27 |
| H3K9ac | Acetylation of histone H3 at lysine 9 |
| H3K18ac | Acetylation of histone H3 at lysine 18 |
| H3K56ac | Acetylation of histone H3 at lysine 56 |
| HAND2-AS1 | HAND2 antisense RNA 1 |
| HBV | Hepatitis B virus |
| HCC | Hepatocellular carcinoma |
| HCV | Hepatitis C virus |
| HEIH | Hepatocellular carcinoma upregulated EZH2-associated long noncoding RNA |
| HIF1A | Hypoxia-inducible factor 1 subunit alpha |
| HIG2 | Hypoxia-inducible protein 2 |
| HK2 | Hexokinase 2 |
| HMGA2 | High mobility group AT-hook 2 |
| HNRNPK | Heterogenous nuclear ribonucleoprotein K |
| HNRNPL | Heterogenous nuclear ribonucleoprotein L |
| HNRNPU | Heterogenous nuclear ribonucleoprotein U |
| HOTAIR | HOX transcript antisense RNA |
| HOTTIP | HOXA distal transcript antisense RNA |
| HSP90 | Heat shock protein 90 |
| HULC | Highly upregulated in liver cancer |
| HuR | Human antigen R |
| ICAM1 | Intercellular adhesion molecule 1 |
| ICR | ICAM-1-related |
| IDR | Intrinsically disordered region |
| IGF2 | Insulin-like growth factor 2 |
| IL-6 | Interleukin 6 |
| IL-11 | Interleukin 11 |
| JAK | Janus kinase |
| KLF2 | KLF transcription factor 2 |
| KMT2A/MLL | Lysine methyltransferase 2A |
| LATS1 | Large tumor suppressor kinase 1 |
| LINC-ROR | Long intergenic non-protein coding RNA, regulator of reprogramming |
| LINP1 | LncRNA in non-homologous end joining pathway 1 |
| LLPS | Liquid-liquid phase separation |
| LncRNA | Long noncoding ribonucleic acid |
| lncRNA-ATB | LncRNA-activated by TGF-β |
| LncPRESS1 | LncRNA p53-regulated and ESC-associated |
| LSD1 | Lysine-specific histone demethylase 1 |
| M6A | N6-methyladenosine |
| MAFLD | Metabolic dysfunction-related fatty liver disease |
| MALAT1 | Metastasis associated lung adenocarcinoma transcript 1 |
| MAPK | Mitogen-activated protein kinase |
| MATR3 | Matrin 3 |
| MBNL3 | Muscleblind like splicing regulator 3 |
| MCM2 | Minichromosome maintenance complex component 2 |
| MEG3 | Maternally expressed 3 |
| MEK | MAP kinase-ERK kinase |
| MELTF-AS1 | MELTF antisense RNA 1 |
| MEX3B | Mex-3 RNA-binding family member B |
| miRNA | microRNA |
| MMP | Matrix metallopeptidase |
| MMTV-PyMT | mouse mammary tumor virus (MMTV) long terminal repeat upstream of a cDNA sequence encoding the Polyoma Virus middle T antigen (PyMT) |
| MRI | Magnetic resonance imaging |
| MTCH2 | Mitochondrial carrier 2 |
| mTOR | Mammalian target of rapamycin |
| MVIH | LncRNA associated with microvascular invasion in HCC |
| MYC | MYC proto-oncogene, bHLH transcription factor |
| NAFLD | Non-alcoholic fatty liver disease |
| NASH | Non-alcoholic steatohepatitis |
| NcRNA | Noncoding ribonucleic acid |
| ND3 | NADH dehydrogenase subunit 3 |
| NEAT1 | Nuclear paraspeckle assembly transcript 1 |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| NHEJ | Non-homologous end joining |
| NIHCOLE | Noncoding RNA induced in hepatocellular carcinoma with an oncogenic role in ligation efficiency |
| NONO | Non-POU domain-containing octamer binding protein |
| NOP2 | NOP2 nucleolar protein |
| NORAD | Noncoding RNA activated by DNA damage |
| NRAS | NRAS proto-oncogene, GTPase |
| NSCLC | Non-small cell lung cancer |
| PAARH | Progression and angiogenesis associated RNA in HCC |
| PD-1 | Programmed cell death 1 |
| PD-L1 | Programmed cell death 1 ligand 1 |
| PDX | Patient-derived xenograft |
| PGK1 | Phosphoglycerate kinase 1 |
| PHAROH | Pluripotency and hepatocyte associated RNA overexpressed in HCC |
| PHF8 | PHD finger protein 8 |
| PI3K | Phosphatidylinositol 3-kinase |
| PNCTR | Pyrimidine-rich noncoding transcript |
| PNUTS | Phosphatase 1 nuclear targeting subunit |
| Pol I | RNA polymerase I |
| Pol II | RNA polymerase II |
| Pol III | RNA polymerase III |
| PPARA | Peroxisome proliferator–activated receptor alpha |
| PPP1CA | Protein phosphatase 1 catalytic subunit alpha |
| PRAL | p53 regulation-associated lncRNA |
| PRC2 | Polycomb Repressive Complex 2 |
| PRR34-AS1 | PRR34 antisense RNA 1 |
| PTBP1 | Pyrimidine tract binding protein 1 |
| PTEN | Phosphatase and tensin homolog |
| PUM1 | Pumilio RNA-binding family member 1 |
| PUM2 | Pumilio RNA-binding family member 2 |
| PVT1 | Pvt1 oncogene |
| PVTT | Portal vein tumor thrombus |
| PXN-AS1 | Paxillin antisense RNA 1 |
| RAB35 | RAB35, member RAS oncogene family |
| RBP | RNA-binding protein |
| RFA | Radiofrequency ablation |
| RMST | Rhabdomyosarcoma 2 associated transcript |
| RNA | Ribonucleic acid |
| RNase | Ribonuclease |
| RNAi | RNA interference |
| RNP | Ribonucleoprotein |
| RPS6KB1 | Ribosomal protein S6 kinase B1 |
| RXRA | Retinoid X receptor alpha |
| SAT1 | Spermidine/spermine N1-acetyltransferase 1 |
| SCARNA10 | Small cajal body specific RNA 10 |
| SERPINH1 | Serpin family H member 1 |
| SET1 | Histone methyltransferase SET1 |
| SETD2 | SET domain containing 2, histone lysine methyltransferase |
| SF3B3 | Splicing factor 3b subunit 3 |
| SFPQ | Splicing factor proline and glutamine rich |
| shRNA | Short hairpin RNA |
| siRNA | Small interfering RNA |
| SIRT1 | Sirtuin 1 |
| SIRT6 | Sirtuin 6 |
| SLERT | Box H/ACA small nucleolar RNA (snoRNA)–ended lncRNA |
| SNAIL | Zing-finger transcription factor Snail |
| SNAP23 | Synaptosome associated protein 23 |
| SNHG6 | Small nucleolar RNA host gene 6 |
| SNHG9 | Small nucleolar RNA host gene 9 |
| snRNA | Small nuclear RNA |
| snoRNA | Small nucleolar RNA |
| SOCS2 | Suppressor of cytokine signaling 2 |
| SOX2 | SRY-box transcription factor 2 |
| SOX12 | SRY-box transcription factor 12 |
| SPHK1 | Sphingosine kinase 1 |
| SR | Serine/arginine |
| SREBP1 | Sterol regulatory element binding protein 1 |
| STAT1 | Signal transducer and activator of transcription 1 |
| STAT3 | Signal transducer and activator of transcription 3 |
| SUZ12 | SUZ12 polycomb repressive complex 2 subunit |
| TACE | Transarterial chemoembolization |
| TAK1 | Transforming growth factor-β-activated kinase 1 |
| TARID | TCF21 antisense RNA inducing promoter demethylation |
| TCF7L2 | Transcription factor 7 like 2 |
| TCF21 | Transcription factor 21 |
| TDP-43 | Tar DNA binding protein |
| TERT | Telomerase reverse transcriptase |
| TET1 | Tet methylcytosine dioxygenase 1 |
| TEX10 | Testis expressed 10 |
| TGF-β | Transforming growth factor-β |
| TGFBR2 | Transforming growth factor-β receptor 2 |
| TIAR | TIA1 cytotoxic granule associated RNA-binding protein like 1 |
| TKI | Tyrosine kinase inhibitor |
| TNBC | Triple negative breast cancer |
| TP53 | Tumor protein p53 |
| TRAF6 | TNF receptor associated factor 6 |
| Treg | Regulatory T cells |
| TRERNA1 | Translation regulatory lncRNA 1 |
| TUG1 | Taurine upregulated gene 1 |
| URB1-AS1 | URB1-antisense RNA 1 |
| USP22 | Ubiquitin-specific peptidase 22 |
| VAMP3 | Vesicle associated membrane protein 3 |
| VLP | Virus-like particles |
| VEGF | Vascular endothelial growth factor |
| VLDLR | Very low density lipoprotein receptor |
| WDR5 | WD repeat domain 5 |
| WTAP | WT1 associated protein |
| XIST | X inactive specific transcript |
| YAP1 | Yes1 associated transcriptional regulator |
| YBX1 | Y-box binding protein 1 |
| ZBTB7A | Zinc finger and BTB domain containing 7A |
| ZEB1 | Zinc finger E-box binding homeobox 1 |
| ZEB2 | Zinc finger E-box binding homeobox 2 |
| ZFAS1 | ZNFX1 antisense RNA 1 |
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Eslam, M.; Sanyal, A.J.; George, J.; International Consensus, P. MAFLD: A Consensus-Driven Proposed Nomenclature for Metabolic Associated Fatty Liver Disease. Gastroenterology 2020, 158, 1999–2014.e1991. [Google Scholar] [CrossRef] [PubMed]
- Pennisi, G.; Celsa, C.; Giammanco, A.; Spatola, F.; Petta, S. The Burden of Hepatocellular Carcinoma in Non-Alcoholic Fatty Liver Disease: Screening Issue and Future Perspectives. Int. J. Mol. Sci. 2019, 20, 5613. [Google Scholar] [CrossRef] [PubMed]
- Rebouissou, S.; Nault, J.C. Advances in molecular classification and precision oncology in hepatocellular carcinoma. J. Hepatol. 2020, 72, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, A. Hepatocellular Carcinoma. N. Engl. J. Med. 2019, 380, 1450–1462. [Google Scholar] [CrossRef]
- Mahpour, A.; Mullen, A.C. Our emerging understanding of the roles of long non-coding RNAs in normal liver function, disease, and malignancy. JHEP Rep. 2021, 3, 100177. [Google Scholar] [CrossRef]
- Nagaraju, G.P.; Dariya, B.; Kasa, P.; Peela, S.; El-Rayes, B.F. Epigenetics in hepatocellular carcinoma. Semin. Cancer Biol. 2022, 86, 622–632. [Google Scholar] [CrossRef]
- Zhang, G.; Li, R.; Zhao, X.; Meng, S.; Ye, J.; Zhao, L. Validation of the American Joint Committee on Cancer eighth edition staging system in patients undergoing hepatectomy for hepatocellular carcinoma: A US population-based study. J. Surg. Res. 2018, 222, 55–68. [Google Scholar] [CrossRef]
- Labgaa, I.; Villanueva, A.; Dormond, O.; Demartines, N.; Melloul, E. The Role of Liquid Biopsy in Hepatocellular Carcinoma Prognostication. Cancers 2021, 13, 659. [Google Scholar] [CrossRef]
- Kulik, L.; El-Serag, H.B. Epidemiology and Management of Hepatocellular Carcinoma. Gastroenterology 2019, 156, 477–491.e471. [Google Scholar] [CrossRef]
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikarsky, E.; Roayaie, S.; Lencioni, R.; Koike, K.; Zucman-Rossi, J.; Finn, R.S. Hepatocellular carcinoma. Nat. Rev. Dis. Primers 2021, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- El-Khoueiry, A.B.; Sangro, B.; Yau, T.; Crocenzi, T.S.; Kudo, M.; Hsu, C.; Kim, T.Y.; Choo, S.P.; Trojan, J.; Welling, T.H.R.; et al. Nivolumab in patients with advanced hepatocellular carcinoma (CheckMate 040): An open-label, non-comparative, phase 1/2 dose escalation and expansion trial. Lancet 2017, 389, 2492–2502. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Ryoo, B.Y.; Hsu, C.H.; Numata, K.; Stein, S.; Verret, W.; Hack, S.P.; Spahn, J.; Liu, B.; Abdullah, H.; et al. Atezolizumab with or without bevacizumab in unresectable hepatocellular carcinoma (GO30140): An open-label, multicentre, phase 1b study. Lancet Oncol. 2020, 21, 808–820. [Google Scholar] [CrossRef] [PubMed]
- Casak, S.J.; Donoghue, M.; Fashoyin-Aje, L.; Jiang, X.; Rodriguez, L.; Shen, Y.L.; Xu, Y.; Jiang, X.; Liu, J.; Zhao, H.; et al. FDA Approval Summary: Atezolizumab Plus Bevacizumab for the Treatment of Patients with Advanced Unresectable or Metastatic Hepatocellular Carcinoma. Clin. Cancer Res. 2021, 27, 1836–1841. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.S.; Qin, S.; Ikeda, M.; Galle, P.R.; Ducreux, M.; Kim, T.Y.; Kudo, M.; Breder, V.; Merle, P.; Kaseb, A.O.; et al. Atezolizumab plus Bevacizumab in Unresectable Hepatocellular Carcinoma. N. Engl. J. Med. 2020, 382, 1894–1905. [Google Scholar] [CrossRef]
- Reig, M.; Forner, A.; Rimola, J.; Ferrer-Fabrega, J.; Burrel, M.; Garcia-Criado, A.; Kelley, R.K.; Galle, P.R.; Mazzaferro, V.; Salem, R.; et al. BCLC strategy for prognosis prediction and treatment recommendation: The 2022 update. J. Hepatol. 2022, 76, 681–693. [Google Scholar] [CrossRef] [PubMed]
- Anstee, Q.M.; Reeves, H.L.; Kotsiliti, E.; Govaere, O.; Heikenwalder, M. From NASH to HCC: Current concepts and future challenges. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 411–428. [Google Scholar] [CrossRef]
- Bird, A. Perceptions of epigenetics. Nature 2007, 447, 396–398. [Google Scholar] [CrossRef]
- Hon, G.C.; Rajagopal, N.; Shen, Y.; McCleary, D.F.; Yue, F.; Dang, M.D.; Ren, B. Epigenetic memory at embryonic enhancers identified in DNA methylation maps from adult mouse tissues. Nat. Genet. 2013, 45, 1198–1206. [Google Scholar] [CrossRef]
- Jones, P.A.; Takai, D. The role of DNA methylation in mammalian epigenetics. Science 2001, 293, 1068–1070. [Google Scholar] [CrossRef]
- Esteller, M. Epigenetics in cancer. N. Engl. J. Med. 2008, 358, 1148–1159. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, A.; Portela, A.; Sayols, S.; Battiston, C.; Hoshida, Y.; Mendez-Gonzalez, J.; Imbeaud, S.; Letouze, E.; Hernandez-Gea, V.; Cornella, H.; et al. DNA methylation-based prognosis and epidrivers in hepatocellular carcinoma. Hepatology 2015, 61, 1945–1956. [Google Scholar] [CrossRef] [PubMed]
- Kouzarides, T. Chromatin modifications and their function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef] [PubMed]
- Arrowsmith, C.H.; Bountra, C.; Fish, P.V.; Lee, K.; Schapira, M. Epigenetic protein families: A new frontier for drug discovery. Nat. Rev. Drug Discov. 2012, 11, 384–400. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Mu, H.; Xiao, Y.; Zhao, Z.; Cui, X.; Wu, D. Comprehensive Analysis of Histone Modifications in Hepatocellular Carcinoma Reveals Different Subtypes and Key Prognostic Models. J. Oncol. 2022, 2022, 5961603. [Google Scholar] [CrossRef] [PubMed]
- Rinn, J.L.; Chang, H.Y. Genome regulation by long noncoding RNAs. Annu. Rev. Biochem. 2012, 81, 145–166. [Google Scholar] [CrossRef] [PubMed]
- Gebert, L.F.R.; MacRae, I.J. Regulation of microRNA function in animals. Nat. Rev. Mol. Cell Biol. 2019, 20, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Henson, R.; Wehbe-Janek, H.; Ghoshal, K.; Jacob, S.T.; Patel, T. MicroRNA-21 regulates expression of the PTEN tumor suppressor gene in human hepatocellular cancer. Gastroenterology 2007, 133, 647–658. [Google Scholar] [CrossRef]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; et al. Long non-coding RNAs: Definitions, functions, challenges and recommendations. Nat. Rev. Mol. Cell Biol. 2023, 24, 430–447. [Google Scholar] [CrossRef]
- Hartford, C.C.R.; Lal, A. When Long Noncoding Becomes Protein Coding. Mol. Cell Biol. 2020, 40, e00528-19. [Google Scholar] [CrossRef]
- Ma, L.; Bajic, V.B.; Zhang, Z. On the classification of long non-coding RNAs. RNA Biol. 2013, 10, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Kopp, F.; Mendell, J.T. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef] [PubMed]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef] [PubMed]
- Cao, J. The functional role of long non-coding RNAs and epigenetics. Biol. Proced. Online 2014, 16, 11. [Google Scholar] [CrossRef] [PubMed]
- Dueva, R.; Akopyan, K.; Pederiva, C.; Trevisan, D.; Dhanjal, S.; Lindqvist, A.; Farnebo, M. Neutralization of the Positive Charges on Histone Tails by RNA Promotes an Open Chromatin Structure. Cell Chem. Biol. 2019, 26, 1436–1449.e1435. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.C.; Yang, Y.W.; Liu, B.; Sanyal, A.; Corces-Zimmerman, R.; Chen, Y.; Lajoie, B.R.; Protacio, A.; Flynn, R.A.; Gupta, R.A.; et al. A long noncoding RNA maintains active chromatin to coordinate homeotic gene expression. Nature 2011, 472, 120–124. [Google Scholar] [CrossRef]
- Jain, A.K.; Xi, Y.; McCarthy, R.; Allton, K.; Akdemir, K.C.; Patel, L.R.; Aronow, B.; Lin, C.; Li, W.; Yang, L.; et al. LncPRESS1 Is a p53-Regulated LncRNA that Safeguards Pluripotency by Disrupting SIRT6-Mediated De-acetylation of Histone H3K56. Mol. Cell 2016, 64, 967–981. [Google Scholar] [CrossRef]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef]
- Yap, K.L.; Li, S.; Munoz-Cabello, A.M.; Raguz, S.; Zeng, L.; Mujtaba, S.; Gil, J.; Walsh, M.J.; Zhou, M.M. Molecular interplay of the noncoding RNA ANRIL and methylated histone H3 lysine 27 by polycomb CBX7 in transcriptional silencing of INK4a. Mol. Cell 2010, 38, 662–674. [Google Scholar] [CrossRef]
- Arab, K.; Karaulanov, E.; Musheev, M.; Trnka, P.; Schafer, A.; Grummt, I.; Niehrs, C. GADD45A binds R-loops and recruits TET1 to CpG island promoters. Nat. Genet. 2019, 51, 217–223. [Google Scholar] [CrossRef]
- Arab, K.; Park, Y.J.; Lindroth, A.M.; Schafer, A.; Oakes, C.; Weichenhan, D.; Lukanova, A.; Lundin, E.; Risch, A.; Meister, M.; et al. Long noncoding RNA TARID directs demethylation and activation of the tumor suppressor TCF21 via GADD45A. Mol. Cell 2014, 55, 604–614. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Wang, X.; Youmans, D.T.; Cech, T.R. How do lncRNAs regulate transcription? Sci. Adv. 2017, 3, eaao2110. [Google Scholar] [CrossRef]
- Kino, T.; Hurt, D.E.; Ichijo, T.; Nader, N.; Chrousos, G.P. Noncoding RNA gas5 is a growth arrest- and starvation-associated repressor of the glucocorticoid receptor. Sci. Signal 2010, 3, ra8. [Google Scholar] [CrossRef]
- Ng, S.Y.; Bogu, G.K.; Soh, B.S.; Stanton, L.W. The long noncoding RNA RMST interacts with SOX2 to regulate neurogenesis. Mol. Cell 2013, 51, 349–359. [Google Scholar] [CrossRef]
- Rom, A.; Melamed, L.; Gil, N.; Goldrich, M.J.; Kadir, R.; Golan, M.; Biton, I.; Perry, R.B.; Ulitsky, I. Regulation of CHD2 expression by the Chaserr long noncoding RNA gene is essential for viability. Nat. Commun. 2019, 10, 5092. [Google Scholar] [CrossRef]
- Tripathi, V.; Ellis, J.D.; Shen, Z.; Song, D.Y.; Pan, Q.; Watt, A.T.; Freier, S.M.; Bennett, C.F.; Sharma, A.; Bubulya, P.A.; et al. The nuclear-retained noncoding RNA MALAT1 regulates alternative splicing by modulating SR splicing factor phosphorylation. Mol. Cell 2010, 39, 925–938. [Google Scholar] [CrossRef]
- Yap, K.; Mukhina, S.; Zhang, G.; Tan, J.S.C.; Ong, H.S.; Makeyev, E.V. A Short Tandem Repeat-Enriched RNA Assembles a Nuclear Compartment to Control Alternative Splicing and Promote Cell Survival. Mol. Cell 2018, 72, 525–540 e513. [Google Scholar] [CrossRef]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Grelet, S.; Link, L.A.; Howley, B.; Obellianne, C.; Palanisamy, V.; Gangaraju, V.K.; Diehl, J.A.; Howe, P.H. A regulated PNUTS mRNA to lncRNA splice switch mediates EMT and tumour progression. Nat. Cell Biol. 2017, 19, 1105–1115. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.; Brangwynne, C.P. Liquid phase condensation in cell physiology and disease. Science 2017, 357, eaaf4382. [Google Scholar] [CrossRef]
- Lin, Y.; Protter, D.S.; Rosen, M.K.; Parker, R. Formation and Maturation of Phase-Separated Liquid Droplets by RNA-Binding Proteins. Mol. Cell 2015, 60, 208–219. [Google Scholar] [CrossRef]
- Adnane, S.; Marino, A.; Leucci, E. LncRNAs in human cancers: Signal from noise. Trends Cell Biol. 2022, 32, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Clemson, C.M.; Hutchinson, J.N.; Sara, S.A.; Ensminger, A.W.; Fox, A.H.; Chess, A.; Lawrence, J.B. An architectural role for a nuclear noncoding RNA: NEAT1 RNA is essential for the structure of paraspeckles. Mol. Cell 2009, 33, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Takakuwa, H.; Yamazaki, T.; Souquere, S.; Adachi, S.; Yoshino, H.; Fujiwara, N.; Yamamoto, T.; Natsume, T.; Nakagawa, S.; Pierron, G.; et al. Shell protein composition specified by the lncRNA NEAT1 domains dictates the formation of paraspeckles as distinct membraneless organelles. Nat. Cell Biol. 2023, 25, 1664–1675. [Google Scholar] [CrossRef]
- Nakagawa, S.; Shimada, M.; Yanaka, K.; Mito, M.; Arai, T.; Takahashi, E.; Fujita, Y.; Fujimori, T.; Standaert, L.; Marine, J.C.; et al. The lncRNA Neat1 is required for corpus luteum formation and the establishment of pregnancy in a subpopulation of mice. Development 2014, 141, 4618–4627. [Google Scholar] [CrossRef]
- Adriaens, C.; Standaert, L.; Barra, J.; Latil, M.; Verfaillie, A.; Kalev, P.; Boeckx, B.; Wijnhoven, P.W.; Radaelli, E.; Vermi, W.; et al. p53 induces formation of NEAT1 lncRNA-containing paraspeckles that modulate replication stress response and chemosensitivity. Nat. Med. 2016, 22, 861–868. [Google Scholar] [CrossRef]
- Daneshvar, K.; Ardehali, M.B.; Klein, I.A.; Hsieh, F.K.; Kratkiewicz, A.J.; Mahpour, A.; Cancelliere, S.O.L.; Zhou, C.; Cook, B.M.; Li, W.; et al. lncRNA DIGIT and BRD3 protein form phase-separated condensates to regulate endoderm differentiation. Nat. Cell Biol. 2020, 22, 1211–1222. [Google Scholar] [CrossRef]
- Wu, M.; Xu, G.; Han, C.; Luan, P.F.; Xing, Y.H.; Nan, F.; Yang, L.Z.; Huang, Y.; Yang, Z.H.; Shan, L.; et al. lncRNA SLERT controls phase separation of FC/DFCs to facilitate Pol I transcription. Science 2021, 373, 547–555. [Google Scholar] [CrossRef]
- Xing, Y.H.; Yao, R.W.; Zhang, Y.; Guo, C.J.; Jiang, S.; Xu, G.; Dong, R.; Yang, L.; Chen, L.L. SLERT Regulates DDX21 Rings Associated with Pol I Transcription. Cell 2017, 169, 664–678 e616. [Google Scholar] [CrossRef] [PubMed]
- Elguindy, M.M.; Mendell, J.T. NORAD-induced Pumilio phase separation is required for genome stability. Nature 2021, 595, 303–308. [Google Scholar] [CrossRef]
- Li, R.H.; Tian, T.; Ge, Q.W.; He, X.Y.; Shi, C.Y.; Li, J.H.; Zhang, Z.; Liu, F.Z.; Sang, L.J.; Yang, Z.Z.; et al. A phosphatidic acid-binding lncRNA SNHG9 facilitates LATS1 liquid-liquid phase separation to promote oncogenic YAP signaling. Cell Res. 2021, 31, 1088–1105. [Google Scholar] [CrossRef]
- Lu, X.; Wang, J.; Wang, W.; Lu, C.; Qu, T.; He, X.; Liu, X.; Guo, R.; Zhang, E. Copy number amplification and SP1-activated lncRNA MELTF-AS1 regulates tumorigenesis by driving phase separation of YBX1 to activate ANXA8 in non-small cell lung cancer. Oncogene 2022, 41, 3222–3238. [Google Scholar] [CrossRef]
- Thapar, R.; Wang, J.L.; Hammel, M.; Ye, R.; Liang, K.; Sun, C.; Hnizda, A.; Liang, S.; Maw, S.S.; Lee, L.; et al. Mechanism of efficient double-strand break repair by a long non-coding RNA. Nucleic Acids Res. 2021, 49, 1199–1200. [Google Scholar] [CrossRef]
- Pessina, F.; Giavazzi, F.; Yin, Y.; Gioia, U.; Vitelli, V.; Galbiati, A.; Barozzi, S.; Garre, M.; Oldani, A.; Flaus, A.; et al. Functional transcription promoters at DNA double-strand breaks mediate RNA-driven phase separation of damage-response factors. Nat. Cell Biol. 2019, 21, 1286–1299. [Google Scholar] [CrossRef]
- Pandya-Jones, A.; Markaki, Y.; Serizay, J.; Chitiashvili, T.; Mancia Leon, W.R.; Damianov, A.; Chronis, C.; Papp, B.; Chen, C.K.; McKee, R.; et al. A protein assembly mediates Xist localization and gene silencing. Nature 2020, 587, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Cao, L.; Thorne, R.F.; Zhang, X.D.; Li, J.; Shao, F.; Zhang, L.; Wu, M. LncRNA GIRGL drives CAPRIN1-mediated phase separation to suppress glutaminase-1 translation under glutamine deprivation. Sci. Adv. 2021, 7, eabe5708. [Google Scholar] [CrossRef]
- Wang, C.; Duan, Y.; Duan, G.; Wang, Q.; Zhang, K.; Deng, X.; Qian, B.; Gu, J.; Ma, Z.; Zhang, S.; et al. Stress Induces Dynamic, Cytotoxicity-Antagonizing TDP-43 Nuclear Bodies via Paraspeckle LncRNA NEAT1-Mediated Liquid-Liquid Phase Separation. Mol. Cell 2020, 79, 443–458.e447. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, S.; Zhu, X.; Zhang, L.; Deng, J.; Li, F.; Guo, B.; Zhang, S.; Wu, R.; Zhang, Z.; et al. LncRNA-encoded polypeptide ASRPS inhibits triple-negative breast cancer angiogenesis. J. Exp. Med. 2020, 217, jem.20190950. [Google Scholar] [CrossRef]
- Guo, B.; Wu, S.; Zhu, X.; Zhang, L.; Deng, J.; Li, F.; Wang, Y.; Zhang, S.; Wu, R.; Lu, J.; et al. Micropeptide CIP2A-BP encoded by LINC00665 inhibits triple-negative breast cancer progression. EMBO J. 2020, 39, e102190. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.G.; Yang, J.; Zhu, Y.; Zhu, Q.; Pan, W.; Deng, S.; He, Y.; Zuo, D.; Wang, P.; Han, Y.; et al. The microprotein encoded by exosomal lncAKR1C2 promotes gastric cancer lymph node metastasis by regulating fatty acid metabolism. Cell Death Dis. 2023, 14, 708. [Google Scholar] [CrossRef]
- Zhang, Q.; Wei, T.; Yan, L.; Zhu, S.; Jin, W.; Bai, Y.; Zeng, Y.; Zhang, X.; Yin, Z.; Yang, J.; et al. Hypoxia-Responsive lncRNA AC115619 Encodes a Micropeptide That Suppresses m6A Modifications and Hepatocellular Carcinoma Progression. Cancer Res. 2023, 83, 2496–2512. [Google Scholar] [CrossRef] [PubMed]
- Tornini, V.A.; Miao, L.; Lee, H.J.; Gerson, T.; Dube, S.E.; Schmidt, V.; Kroll, F.; Tang, Y.; Du, K.; Kuchroo, M.; et al. linc-mipep and linc-wrb encode micropeptides that regulate chromatin accessibility in vertebrate-specific neural cells. eLife 2023, 12, e82249. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liao, Z.; Liu, F.; Su, C.; Zhu, H.; Li, Y.; Tao, R.; Liang, H.; Zhang, B.; Zhang, X. Long noncoding RNA HULC promotes hepatocellular carcinoma progression. Aging 2019, 11, 9111–9127. [Google Scholar] [CrossRef] [PubMed]
- Wan, T.; Wang, H.; Gou, M.; Si, H.; Wang, Z.; Yan, H.; Liu, T.; Chen, S.; Fan, R.; Qian, N.; et al. LncRNA HEIH promotes cell proliferation, migration and invasion in cholangiocarcinoma by modulating miR-98-5p/HECTD4. Biomed. Pharmacother. 2020, 125, 109916. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chen, S.; Yang, G.; Gu, F.; Li, M.; Zhong, B.; Hu, J.; Hoffman, A.; Chen, M. Long noncoding RNA HOTAIR as an independent prognostic marker in cancer: A meta-analysis. PLoS ONE 2014, 9, e105538. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zeng, J.; Chen, W.; Fan, J.; Hylemon, P.B.; Zhou, H. Long Noncoding RNA H19: A Novel Oncogene in Liver Cancer. Noncoding RNA 2023, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Guo, J.; Liu, J.; Mao, X.; Xu, K. Long Non-coding RNA MALAT1: A Key Player in Liver Diseases. Front. Med. 2021, 8, 734643. [Google Scholar] [CrossRef]
- Wong, L.S.; Wong, C.M. Decoding the Roles of Long Noncoding RNAs in Hepatocellular Carcinoma. Int. J. Mol. Sci. 2021, 22, 3137. [Google Scholar] [CrossRef]
- He, Z.; Yang, D.; Fan, X.; Zhang, M.; Li, Y.; Gu, X.; Yang, M. The Roles and Mechanisms of lncRNAs in Liver Fibrosis. Int. J. Mol. Sci. 2020, 21, 1482. [Google Scholar] [CrossRef]
- Yan, C.; Chen, J.; Chen, N. Long noncoding RNA MALAT1 promotes hepatic steatosis and insulin resistance by increasing nuclear SREBP-1c protein stability. Sci. Rep. 2016, 6, 22640. [Google Scholar] [CrossRef]
- Xiong, H.; Ni, Z.; He, J.; Jiang, S.; Li, X.; He, J.; Gong, W.; Zheng, L.; Chen, S.; Li, B.; et al. LncRNA HULC triggers autophagy via stabilizing Sirt1 and attenuates the chemosensitivity of HCC cells. Oncogene 2017, 36, 3528–3540. [Google Scholar] [CrossRef] [PubMed]
- Unfried, J.P.; Fortes, P. LncRNAs in HCV Infection and HCV-Related Liver Disease. Int. J. Mol. Sci. 2020, 21, 2255. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, L.; Lu, B.; Zhao, M.; Li, L.; Sun, W.; Qiu, Z.; Zhang, B. LncRNA H19/microRNA-675/PPARalpha axis regulates liver cell injury and energy metabolism remodelling induced by hepatitis B X protein via Akt/mTOR signalling. Mol. Immunol. 2019, 116, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Kogo, R.; Shimamura, T.; Mimori, K.; Kawahara, K.; Imoto, S.; Sudo, T.; Tanaka, F.; Shibata, K.; Suzuki, A.; Komune, S.; et al. Long noncoding RNA HOTAIR regulates polycomb-dependent chromatin modification and is associated with poor prognosis in colorectal cancers. Cancer Res. 2011, 71, 6320–6326. [Google Scholar] [CrossRef]
- Verma, S.; Sahu, B.D.; Mugale, M.N. Role of lncRNAs in hepatocellular carcinoma. Life Sci. 2023, 325, 121751. [Google Scholar] [CrossRef] [PubMed]
- Ji, P.; Diederichs, S.; Wang, W.; Boing, S.; Metzger, R.; Schneider, P.M.; Tidow, N.; Brandt, B.; Buerger, H.; Bulk, E.; et al. MALAT-1, a novel noncoding RNA, and thymosin beta4 predict metastasis and survival in early-stage non-small cell lung cancer. Oncogene 2003, 22, 8031–8041. [Google Scholar] [CrossRef]
- Gutschner, T.; Hammerle, M.; Eissmann, M.; Hsu, J.; Kim, Y.; Hung, G.; Revenko, A.; Arun, G.; Stentrup, M.; Gross, M.; et al. The noncoding RNA MALAT1 is a critical regulator of the metastasis phenotype of lung cancer cells. Cancer Res. 2013, 73, 1180–1189. [Google Scholar] [CrossRef]
- Huang, Z.; Zhou, J.K.; Peng, Y.; He, W.; Huang, C. The role of long noncoding RNAs in hepatocellular carcinoma. Mol. Cancer 2020, 19, 77. [Google Scholar] [CrossRef]
- Panzitt, K.; Tschernatsch, M.M.; Guelly, C.; Moustafa, T.; Stradner, M.; Strohmaier, H.M.; Buck, C.R.; Denk, H.; Schroeder, R.; Trauner, M.; et al. Characterization of HULC, a novel gene with striking up-regulation in hepatocellular carcinoma, as noncoding RNA. Gastroenterology 2007, 132, 330–342. [Google Scholar] [CrossRef]
- Xie, H.; Ma, H.; Zhou, D. Plasma HULC as a promising novel biomarker for the detection of hepatocellular carcinoma. BioMed Res. Int. 2013, 2013, 136106. [Google Scholar] [CrossRef]
- Tu, Z.Q.; Li, R.J.; Mei, J.Z.; Li, X.H. Down-regulation of long non-coding RNA GAS5 is associated with the prognosis of hepatocellular carcinoma. Int. J. Clin. Exp. Pathol. 2014, 7, 4303–4309. [Google Scholar] [PubMed]
- Wang, W.; Zhou, R.; Wu, Y.; Liu, Y.; Su, W.; Xiong, W.; Zeng, Z. PVT1 Promotes Cancer Progression via MicroRNAs. Front. Oncol. 2019, 9, 609. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yu, Y.; Li, H.; Hu, Q.; Chen, X.; He, Y.; Xue, C.; Ren, F.; Ren, Z.; Li, J.; et al. Long non-coding RNA PVT1 promotes tumor progression by regulating the miR-143/HK2 axis in gallbladder cancer. Mol. Cancer 2019, 18, 33. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ren, H. Long noncoding RNA PVT1 promotes tumor cell proliferation, invasion, migration and inhibits apoptosis in oral squamous cell carcinoma by regulating miR-150-5p/GLUT-1. Oncol. Rep. 2020, 44, 1524–1538. [Google Scholar] [CrossRef] [PubMed]
- Quagliata, L.; Matter, M.S.; Piscuoglio, S.; Arabi, L.; Ruiz, C.; Procino, A.; Kovac, M.; Moretti, F.; Makowska, Z.; Boldanova, T.; et al. Long noncoding RNA HOTTIP/HOXA13 expression is associated with disease progression and predicts outcome in hepatocellular carcinoma patients. Hepatology 2014, 59, 911–923. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.C.; Manor, O.; Wan, Y.; Mosammaparast, N.; Wang, J.K.; Lan, F.; Shi, Y.; Segal, E.; Chang, H.Y. Long noncoding RNA as modular scaffold of histone modification complexes. Science 2010, 329, 689–693. [Google Scholar] [CrossRef] [PubMed]
- Unfried, J.P.; Sangro, P.; Prats-Mari, L.; Sangro, B.; Fortes, P. The Landscape of lncRNAs in Hepatocellular Carcinoma: A Translational Perspective. Cancers 2021, 13, 2651. [Google Scholar] [CrossRef]
- Zhang, Y.; Lin, S.; Yang, X.; Zhang, X. Prognostic and Clinicopathological Significance of lncRNA MVIH in Cancer Patients. J. Cancer 2019, 10, 1503–1510. [Google Scholar] [CrossRef]
- Abbastabar, M.; Sarfi, M.; Golestani, A.; Khalili, E. lncRNA involvement in hepatocellular carcinoma metastasis and prognosis. EXCLI J. 2018, 17, 900–913. [Google Scholar] [CrossRef]
- Matouk, I.J.; DeGroot, N.; Mezan, S.; Ayesh, S.; Abu-lail, R.; Hochberg, A.; Galun, E. The H19 non-coding RNA is essential for human tumor growth. PLoS ONE 2007, 2, e845. [Google Scholar] [CrossRef]
- Zhao, H.; Peng, R.; Liu, Q.; Liu, D.; Du, P.; Yuan, J.; Peng, G.; Liao, Y. The lncRNA H19 interacts with miR-140 to modulate glioma growth by targeting iASPP. Arch. Biochem. Biophys. 2016, 610, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Liu, S.; Ye, F.; Shen, Y.; Tie, Y.; Zhu, J.; Wei, L.; Jin, Y.; Fu, H.; Wu, Y.; et al. Long Noncoding RNA MEG3 Interacts with p53 Protein and Regulates Partial p53 Target Genes in Hepatoma Cells. PLoS ONE 2015, 10, e0139790. [Google Scholar] [CrossRef] [PubMed]
- He, J.H.; Han, Z.P.; Liu, J.M.; Zhou, J.B.; Zou, M.X.; Lv, Y.B.; Li, Y.G.; Cao, M.R. Overexpression of Long Non-Coding RNA MEG3 Inhibits Proliferation of Hepatocellular Carcinoma Huh7 Cells via Negative Modulation of miRNA-664. J. Cell Biochem. 2017, 118, 3713–3721. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.J.; Xie, S.L.; Li, Q.; Ma, J.; Wang, G.Y. Large intervening non-coding RNA HOTAIR is associated with hepatocellular carcinoma progression. J. Int. Med. Res. 2011, 39, 2119–2128. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.M.; Zhu, X.; Wang, W.M.; Lu, Y.F.; Hu, B.G.; Wang, H.; Liang, W.C.; Wang, S.S.; Ko, C.H.; Waye, M.M.; et al. Hotair mediates hepatocarcinogenesis through suppressing miRNA-218 expression and activating P14 and P16 signaling. J. Hepatol. 2015, 63, 886–895. [Google Scholar] [CrossRef] [PubMed]
- Rinn, J.L.; Kertesz, M.; Wang, J.K.; Squazzo, S.L.; Xu, X.; Brugmann, S.A.; Goodnough, L.H.; Helms, J.A.; Farnham, P.J.; Segal, E.; et al. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell 2007, 129, 1311–1323. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Xing, Z.; Mani, S.K.; Bancel, B.; Durantel, D.; Zoulim, F.; Tran, E.J.; Merle, P.; Andrisani, O. RNA helicase DEAD box protein 5 regulates Polycomb repressive complex 2/Hox transcript antisense intergenic RNA function in hepatitis B virus infection and hepatocarcinogenesis. Hepatology 2016, 64, 1033–1048. [Google Scholar] [CrossRef]
- Yang, L.; Peng, X.; Li, Y.; Zhang, X.; Ma, Y.; Wu, C.; Fan, Q.; Wei, S.; Li, H.; Liu, J. Long non-coding RNA HOTAIR promotes exosome secretion by regulating RAB35 and SNAP23 in hepatocellular carcinoma. Mol. Cancer 2019, 18, 78. [Google Scholar] [CrossRef]
- Li, H.; An, J.; Wu, M.; Zheng, Q.; Gui, X.; Li, T.; Pu, H.; Lu, D. LncRNA HOTAIR promotes human liver cancer stem cell malignant growth through downregulation of SETD2. Oncotarget 2015, 6, 27847–27864. [Google Scholar] [CrossRef]
- Su, D.N.; Wu, S.P.; Chen, H.T.; He, J.H. HOTAIR, a long non-coding RNA driver of malignancy whose expression is activated by FOXC1, negatively regulates miRNA-1 in hepatocellular carcinoma. Oncol. Lett. 2016, 12, 4061–4067. [Google Scholar] [CrossRef]
- Kong, J.H.; Qiu, Y.J.; Li, Y.; Zhang, H.J.; Wang, W.P. TGF-β1 elevates P-gp and BCRP in hepatocellular carcinoma through HOTAIR/miR-145 axis. Biopharm. Drug Dispos. 2019, 40, 70–80. [Google Scholar] [CrossRef]
- Cheng, D.; Deng, J.G.; Zhang, B.; He, X.Y.; Meng, Z.; Li, G.L.; Ye, H.L.; Zheng, S.Y.; Wei, L.S.; Deng, X.G.; et al. LncRNA HOTAIR epigenetically suppresses miR-122 expression in hepatocellular carcinoma via DNA methylation. EBioMedicine 2018, 36, 159–170. [Google Scholar] [CrossRef]
- Ding, C.; Cheng, S.; Yang, Z.; Lv, Z.; Xiao, H.; Du, C.; Peng, C.; Xie, H.; Zhou, L.; Wu, J.; et al. Long non-coding RNA HOTAIR promotes cell migration and invasion via down-regulation of RNA binding motif protein 38 in hepatocellular carcinoma cells. Int. J. Mol. Sci. 2014, 15, 4060–4076. [Google Scholar] [CrossRef]
- Zhou, D.; Wang, Y.; Hu, H.; Liu, H.; Deng, J.; Li, L.; Zheng, C. lncRNA MALAT1 promotes HCC metastasis through the peripheral vascular infiltration via miRNA-613: A primary study using contrast ultrasound. World J. Surg. Oncol. 2022, 20, 203. [Google Scholar] [CrossRef]
- Sasaki, Y.T.; Ideue, T.; Sano, M.; Mituyama, T.; Hirose, T. MENepsilon/beta noncoding RNAs are essential for structural integrity of nuclear paraspeckles. Proc. Natl. Acad. Sci. USA 2009, 106, 2525–2530. [Google Scholar] [CrossRef]
- Malakar, P.; Shilo, A.; Mogilevsky, A.; Stein, I.; Pikarsky, E.; Nevo, Y.; Benyamini, H.; Elgavish, S.; Zong, X.; Prasanth, K.V.; et al. Long Noncoding RNA MALAT1 Promotes Hepatocellular Carcinoma Development by SRSF1 Upregulation and mTOR Activation. Cancer Res. 2017, 77, 1155–1167. [Google Scholar] [CrossRef]
- Malakar, P.; Stein, I.; Saragovi, A.; Winkler, R.; Stern-Ginossar, N.; Berger, M.; Pikarsky, E.; Karni, R. Long Noncoding RNA MALAT1 Regulates Cancer Glucose Metabolism by Enhancing mTOR-Mediated Translation of TCF7L2. Cancer Res. 2019, 79, 2480–2493. [Google Scholar] [CrossRef]
- Liu, D.; Zhu, Y.; Pang, J.; Weng, X.; Feng, X.; Guo, Y. Knockdown of long non-coding RNA MALAT1 inhibits growth and motility of human hepatoma cells via modulation of miR-195. J. Cell Biochem. 2018, 119, 1368–1380. [Google Scholar] [CrossRef]
- Chen, L.; Yao, H.; Wang, K.; Liu, X. Long Non-Coding RNA MALAT1 Regulates ZEB1 Expression by Sponging miR-143-3p and Promotes Hepatocellular Carcinoma Progression. J. Cell Biochem. 2017, 118, 4836–4843. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Miao, R.; Liu, S.; Wan, Y.; Zhang, S.; Deng, Y.; Bi, J.; Qu, K.; Zhang, J.; Liu, C. Down-regulation of miR-146b-5p by long noncoding RNA MALAT1 in hepatocellular carcinoma promotes cancer growth and metastasis. Oncotarget 2017, 8, 28683–28695. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wang, G.; Tao, K.; Cai, K.; Wu, K.; Ye, L.; Bai, J.; Yin, Y.; Wang, J.; Shuai, X.; et al. Long noncoding RNA metastasis-associated lung adenocarcinoma transcript 1 cooperates with enhancer of zeste homolog 2 to promote hepatocellular carcinoma development by modulating the microRNA-22/Snail family transcriptional repressor 1 axis. Cancer Sci. 2020, 111, 1582–1595. [Google Scholar] [CrossRef]
- Pan, Y.; Tong, S.; Cui, R.; Fan, J.; Liu, C.; Lin, Y.; Tang, J.; Xie, H.; Lin, P.; Zheng, T.; et al. Long Non-Coding MALAT1 Functions as a Competing Endogenous RNA to Regulate Vimentin Expression by Sponging miR-30a-5p in Hepatocellular Carcinoma. Cell Physiol. Biochem. 2018, 50, 108–120. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhou, L.; Li, H.; Sun, T.; Wen, X.; Li, X.; Meng, Y.; Li, Y.; Liu, M.; Liu, S.; et al. Nuclear-Encoded lncRNA MALAT1 Epigenetically Controls Metabolic Reprogramming in HCC Cells through the Mitophagy Pathway. Mol. Ther. Nucleic Acids 2021, 23, 264–276. [Google Scholar] [CrossRef]
- Li, S.P.; Xu, H.X.; Yu, Y.; He, J.D.; Wang, Z.; Xu, Y.J.; Wang, C.Y.; Zhang, H.M.; Zhang, R.X.; Zhang, J.J.; et al. LncRNA HULC enhances epithelial-mesenchymal transition to promote tumorigenesis and metastasis of hepatocellular carcinoma via the miR-200a-3p/ZEB1 signaling pathway. Oncotarget 2016, 7, 42431–42446. [Google Scholar] [CrossRef]
- Lu, Z.; Xiao, Z.; Liu, F.; Cui, M.; Li, W.; Yang, Z.; Li, J.; Ye, L.; Zhang, X. Long non-coding RNA HULC promotes tumor angiogenesis in liver cancer by up-regulating sphingosine kinase 1 (SPHK1). Oncotarget 2016, 7, 241–254. [Google Scholar] [CrossRef]
- Xin, X.; Wu, M.; Meng, Q.; Wang, C.; Lu, Y.; Yang, Y.; Li, X.; Zheng, Q.; Pu, H.; Gui, X.; et al. Long noncoding RNA HULC accelerates liver cancer by inhibiting PTEN via autophagy cooperation to miR15a. Mol. Cancer 2018, 17, 94. [Google Scholar] [CrossRef]
- Du, Y.; Kong, G.; You, X.; Zhang, S.; Zhang, T.; Gao, Y.; Ye, L.; Zhang, X. Elevation of highly up-regulated in liver cancer (HULC) by hepatitis B virus X protein promotes hepatoma cell proliferation via down-regulating p18. J. Biol. Chem. 2012, 287, 26302–26311. [Google Scholar] [CrossRef]
- Cui, M.; Xiao, Z.; Wang, Y.; Zheng, M.; Song, T.; Cai, X.; Sun, B.; Ye, L.; Zhang, X. Long noncoding RNA HULC modulates abnormal lipid metabolism in hepatoma cells through an miR-9-mediated RXRA signaling pathway. Cancer Res. 2015, 75, 846–857. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Zheng, M.; Sun, B.; Wang, Y.; Ye, L.; Zhang, X. A long noncoding RNA perturbs the circadian rhythm of hepatoma cells to facilitate hepatocarcinogenesis. Neoplasia 2015, 17, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.Y.; Moriarity, B.S.; Gong, W.; Akiyama, R.; Tiwari, A.; Kawakami, H.; Ronning, P.; Reuland, B.; Guenther, K.; Beadnell, T.C.; et al. PVT1 dependence in cancer with MYC copy-number increase. Nature 2014, 512, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Yang, Z.; Lv, Z.; Du, C.; Xiao, H.; Peng, C.; Cheng, S.; Xie, H.; Zhou, L.; Wu, J.; et al. Long non-coding RNA PVT1 is associated with tumor progression and predicts recurrence in hepatocellular carcinoma patients. Oncol. Lett. 2015, 9, 955–963. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Yuan, J.H.; Wang, S.B.; Yang, F.; Yuan, S.X.; Ye, C.; Yang, N.; Zhou, W.P.; Li, W.L.; Li, W.; et al. Oncofetal long noncoding RNA PVT1 promotes proliferation and stem cell-like property of hepatocellular carcinoma cells by stabilizing NOP2. Hepatology 2014, 60, 1278–1290. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Peng, X.; Jin, H.; Liu, J. Long non-coding RNA PVT1 promotes autophagy as ceRNA to target ATG3 by sponging microRNA-365 in hepatocellular carcinoma. Gene 2019, 697, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Luo, X.; He, W.; Chen, G.; Li, Y.; Li, W.; Wang, X.; Lai, Y.; Ye, Y. Long Non-Coding RNA PVT1/miR-150/ HIG2 Axis Regulates the Proliferation, Invasion and the Balance of Iron Metabolism of Hepatocellular Carcinoma. Cell Physiol. Biochem. 2018, 49, 1403–1419. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Liu, J.; Liu, B.; Zeng, Y.; Chen, P.; Su, Y. Long noncoding RNA PVT1 inhibits interferon-alpha mediated therapy for hepatocellular carcinoma cells by interacting with signal transducer and activator of transcription 1. Biochem. Biophys. Res. Commun. 2018, 500, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Yan, X.; Jin, Y.; Yang, X.; Yu, X.; Zhou, L.; Han, S.; Yuan, Q.; Yang, M. MiRNA-192 [corrected] and miRNA-204 Directly Suppress lncRNA HOTTIP and Interrupt GLS1-Mediated Glutaminolysis in Hepatocellular Carcinoma. PLoS Genet. 2015, 11, e1005726. [Google Scholar] [CrossRef]
- Wei, H.; Xu, Z.; Chen, L.; Wei, Q.; Huang, Z.; Liu, G.; Li, W.; Wang, J.; Tang, Q.; Pu, J. Long non-coding RNA PAARH promotes hepatocellular carcinoma progression and angiogenesis via upregulating HOTTIP and activating HIF-1alpha/VEGF signaling. Cell Death Dis. 2022, 13, 102. [Google Scholar] [CrossRef]
- Wang, Y.; Hylemon, P.B.; Zhou, H. Long Noncoding RNA H19: A Key Player in Liver Diseases. Hepatology 2021, 74, 1652–1659. [Google Scholar] [CrossRef]
- Ye, Y.; Guo, J.; Xiao, P.; Ning, J.; Zhang, R.; Liu, P.; Yu, W.; Xu, L.; Zhao, Y.; Yu, J. Macrophages-induced long noncoding RNA H19 up-regulation triggers and activates the miR-193b/MAPK1 axis and promotes cell aggressiveness in hepatocellular carcinoma. Cancer Lett. 2020, 469, 310–322. [Google Scholar] [CrossRef]
- Zhang, J.; Han, C.; Ungerleider, N.; Chen, W.; Song, K.; Wang, Y.; Kwon, H.; Ma, W.; Wu, T. A Transforming Growth Factor-beta and H19 Signaling Axis in Tumor-Initiating Hepatocytes That Regulates Hepatic Carcinogenesis. Hepatology 2019, 69, 1549–1563. [Google Scholar] [CrossRef]
- Huang, Z.; Chu, L.; Liang, J.; Tan, X.; Wang, Y.; Wen, J.; Chen, J.; Wu, Y.; Liu, S.; Liao, J.; et al. H19 Promotes HCC Bone Metastasis Through Reducing Osteoprotegerin Expression in a Protein Phosphatase 1 Catalytic Subunit Alpha/p38 Mitogen-Activated Protein Kinase-Dependent Manner and Sponging microRNA 200b-3p. Hepatology 2021, 74, 214–232. [Google Scholar] [CrossRef]
- Souquere, S.; Beauclair, G.; Harper, F.; Fox, A.; Pierron, G. Highly ordered spatial organization of the structural long noncoding NEAT1 RNAs within paraspeckle nuclear bodies. Mol. Biol. Cell 2010, 21, 4020–4027. [Google Scholar] [CrossRef]
- West, J.A.; Davis, C.P.; Sunwoo, H.; Simon, M.D.; Sadreyev, R.I.; Wang, P.I.; Tolstorukov, M.Y.; Kingston, R.E. The long noncoding RNAs NEAT1 and MALAT1 bind active chromatin sites. Mol. Cell 2014, 55, 791–802. [Google Scholar] [CrossRef]
- Slack, F.J.; Chinnaiyan, A.M. The Role of Non-coding RNAs in Oncology. Cell 2019, 179, 1033–1055. [Google Scholar] [CrossRef]
- Zhang, X.N.; Zhou, J.; Lu, X.J. The long noncoding RNA NEAT1 contributes to hepatocellular carcinoma development by sponging miR-485 and enhancing the expression of the STAT3. J. Cell Physiol. 2018, 233, 6733–6741. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhou, Y.; Yang, L.; Ma, Y.; Peng, X.; Yang, S.; Li, H.; Liu, J. LncRNA NEAT1 promotes autophagy via regulating miR-204/ATG3 and enhanced cell resistance to sorafenib in hepatocellular carcinoma. J. Cell Physiol. 2020, 235, 3402–3413. [Google Scholar] [CrossRef]
- Guo, S.; Chen, W.; Luo, Y.; Ren, F.; Zhong, T.; Rong, M.; Dang, Y.; Feng, Z.; Chen, G. Clinical implication of long non-coding RNA NEAT1 expression in hepatocellular carcinoma patients. Int. J. Clin. Exp. Pathol. 2015, 8, 5395–5402. [Google Scholar]
- Tu, J.; Zhao, Z.; Xu, M.; Lu, X.; Chang, L.; Ji, J. NEAT1 upregulates TGF-beta1 to induce hepatocellular carcinoma progression by sponging hsa-mir-139-5p. J. Cell Physiol. 2018, 233, 8578–8587. [Google Scholar] [CrossRef]
- Zhang, H.; Su, X.; Burley, S.K.; Zheng, X.F.S. mTOR regulates aerobic glycolysis through NEAT1 and nuclear paraspeckle-mediated mechanism in hepatocellular carcinoma. Theranostics 2022, 12, 3518–3533. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhang, L.; Huo, X.S.; Yuan, J.H.; Xu, D.; Yuan, S.X.; Zhu, N.; Zhou, W.P.; Yang, G.S.; Wang, Y.Z.; et al. Long noncoding RNA high expression in hepatocellular carcinoma facilitates tumor growth through enhancer of zeste homolog 2 in humans. Hepatology 2011, 54, 1679–1689. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yang, X.; Qi, Q.; Gao, Y.; Wei, Q.; Han, S. lncRNA-HEIH in serum and exosomes as a potential biomarker in the HCV-related hepatocellular carcinoma. Cancer Biomark. 2018, 21, 651–659. [Google Scholar] [CrossRef]
- Shen, Q.; Jiang, S.; Wu, M.; Zhang, L.; Su, X.; Zhao, D. LncRNA HEIH Confers Cell Sorafenib Resistance in Hepatocellular Carcinoma by Regulating miR-98-5p/PI3K/AKT Pathway. Cancer Manag. Res. 2020, 12, 6585–6595. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Zhang, T.; Zhang, D.; Xie, L.; Zou, X.; Lei, L.; Wu, D.; Liu, L. The long non-coding RNA, SNHG6-003, functions as a competing endogenous RNA to promote the progression of hepatocellular carcinoma. Oncogene 2017, 36, 1112–1122. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Yuan, Y.; Li, C.; Guo, T.; Qi, H.; Xiao, Y.; Dong, X.; Liu, Z.; Liu, Q. Upregulation of SNHG6 regulates ZEB1 expression by competitively binding miR-101-3p and interacting with UPF1 in hepatocellular carcinoma. Cancer Lett. 2016, 383, 183–194. [Google Scholar] [CrossRef]
- Wu, G.; Ju, X.; Wang, Y.; Li, Z.; Gan, X. Up-regulation of SNHG6 activates SERPINH1 expression by competitive binding to miR-139-5p to promote hepatocellular carcinoma progression. Cell Cycle 2019, 18, 1849–1867. [Google Scholar] [CrossRef]
- Chen, S.; Xie, C.; Hu, X. lncRNA SNHG6 functions as a ceRNA to up-regulate c-Myc expression via sponging let-7c-5p in hepatocellular carcinoma. Biochem. Biophys. Res. Commun. 2019, 519, 901–908. [Google Scholar] [CrossRef]
- Fan, X.; Zhao, Z.; Song, J.; Zhang, D.; Wu, F.; Tu, J.; Xu, M.; Ji, J. LncRNA-SNHG6 promotes the progression of hepatocellular carcinoma by targeting miR-6509-5p and HIF1A. Cancer Cell Int. 2021, 21, 150. [Google Scholar] [CrossRef]
- Liu, F.; Tian, T.; Zhang, Z.; Xie, S.; Yang, J.; Zhu, L.; Wang, W.; Shi, C.; Sang, L.; Guo, K.; et al. Long non-coding RNA SNHG6 couples cholesterol sensing with mTORC1 activation in hepatocellular carcinoma. Nat. Metab. 2022, 4, 1022–1040. [Google Scholar] [CrossRef]
- Cao, C.; Sun, J.; Zhang, D.; Guo, X.; Xie, L.; Li, X.; Wu, D.; Liu, L. The long intergenic noncoding RNA UFC1, a target of MicroRNA 34a, interacts with the mRNA stabilizing protein HuR to increase levels of beta-catenin in HCC cells. Gastroenterology 2015, 148, 415–426 e418. [Google Scholar] [CrossRef]
- Zhu, P.; Wang, Y.; Huang, G.; Ye, B.; Liu, B.; Wu, J.; Du, Y.; He, L.; Fan, Z. lnc-beta-Catm elicits EZH2-dependent beta-catenin stabilization and sustains liver CSC self-renewal. Nat. Struct. Mol. Biol. 2016, 23, 631–639. [Google Scholar] [CrossRef]
- Yuan, S.X.; Wang, J.; Yang, F.; Tao, Q.F.; Zhang, J.; Wang, L.L.; Yang, Y.; Liu, H.; Wang, Z.G.; Xu, Q.G.; et al. Long noncoding RNA DANCR increases stemness features of hepatocellular carcinoma by derepression of CTNNB1. Hepatology 2016, 63, 499–511. [Google Scholar] [CrossRef]
- Chen, M.; Zhang, C.; Liu, W.; Du, X.; Liu, X.; Xing, B. Long noncoding RNA LINC01234 promotes hepatocellular carcinoma progression through orchestrating aspartate metabolic reprogramming. Mol. Ther. 2022, 30, 2354–2369. [Google Scholar] [CrossRef]
- Xu, K.; Xia, P.; Gongye, X.; Zhang, X.; Ma, S.; Chen, Z.; Zhang, H.; Liu, J.; Liu, Y.; Guo, Y.; et al. A novel lncRNA RP11-386G11.10 reprograms lipid metabolism to promote hepatocellular carcinoma progression. Mol. Metab. 2022, 63, 101540. [Google Scholar] [CrossRef]
- Sondergaard, J.N.; Sommerauer, C.; Atanasoai, I.; Hinte, L.C.; Geng, K.; Guiducci, G.; Brautigam, L.; Aouadi, M.; Stojic, L.; Barragan, I.; et al. CCT3-LINC00326 axis regulates hepatocarcinogenic lipid metabolism. Gut 2022, 71, 2081–2092. [Google Scholar] [CrossRef]
- Lin, Y.H.; Wu, M.H.; Huang, Y.H.; Yeh, C.T.; Cheng, M.L.; Chi, H.C.; Tsai, C.Y.; Chung, I.H.; Chen, C.Y.; Lin, K.H. Taurine up-regulated gene 1 functions as a master regulator to coordinate glycolysis and metastasis in hepatocellular carcinoma. Hepatology 2018, 67, 188–203. [Google Scholar] [CrossRef]
- Huang, M.D.; Chen, W.M.; Qi, F.Z.; Sun, M.; Xu, T.P.; Ma, P.; Shu, Y.Q. Long non-coding RNA TUG1 is up-regulated in hepatocellular carcinoma and promotes cell growth and apoptosis by epigenetically silencing of KLF2. Mol. Cancer 2015, 14, 165. [Google Scholar] [CrossRef]
- Huang, M.D.; Chen, W.M.; Qi, F.Z.; Xia, R.; Sun, M.; Xu, T.P.; Yin, L.; Zhang, E.B.; De, W.; Shu, Y.Q. Long non-coding RNA ANRIL is upregulated in hepatocellular carcinoma and regulates cell apoptosis by epigenetic silencing of KLF2. J. Hematol. Oncol. 2015, 8, 50. [Google Scholar] [CrossRef]
- Yuan, J.H.; Yang, F.; Wang, F.; Ma, J.Z.; Guo, Y.J.; Tao, Q.F.; Liu, F.; Pan, W.; Wang, T.T.; Zhou, C.C.; et al. A long noncoding RNA activated by TGF-beta promotes the invasion-metastasis cascade in hepatocellular carcinoma. Cancer Cell 2014, 25, 666–681. [Google Scholar] [CrossRef]
- Qin, M.; Meng, Y.; Luo, C.; He, S.; Qin, F.; Yin, Y.; Huang, J.; Zhao, H.; Hu, J.; Deng, Z.; et al. lncRNA PRR34-AS1 promotes HCC development via modulating Wnt/beta-catenin pathway by absorbing miR-296-5p and upregulating E2F2 and SOX12. Mol. Ther. Nucleic Acids 2021, 25, 37–52. [Google Scholar] [CrossRef]
- Li, T.; Xie, J.; Shen, C.; Cheng, D.; Shi, Y.; Wu, Z.; Deng, X.; Chen, H.; Shen, B.; Peng, C.; et al. Amplification of Long Noncoding RNA ZFAS1 Promotes Metastasis in Hepatocellular Carcinoma. Cancer Res. 2015, 75, 3181–3191. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.Z.; Li, F.; Cheng, S.T.; Xu, Y.; Deng, H.J.; Gu, D.Y.; Wang, J.; Chen, W.X.; Zhou, Y.J.; Yang, M.L.; et al. DDX17-regulated alternative splicing that produced an oncogenic isoform of PXN-AS1 to promote HCC metastasis. Hepatology 2022, 75, 847–865. [Google Scholar] [CrossRef]
- Yuan, J.H.; Liu, X.N.; Wang, T.T.; Pan, W.; Tao, Q.F.; Zhou, W.P.; Wang, F.; Sun, S.H. The MBNL3 splicing factor promotes hepatocellular carcinoma by increasing PXN expression through the alternative splicing of lncRNA-PXN-AS1. Nat. Cell Biol. 2017, 19, 820–832. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.X.; Yang, F.; Yang, Y.; Tao, Q.F.; Zhang, J.; Huang, G.; Yang, Y.; Wang, R.Y.; Yang, S.; Huo, X.S.; et al. Long noncoding RNA associated with microvascular invasion in hepatocellular carcinoma promotes angiogenesis and serves as a predictor for hepatocellular carcinoma patients’ poor recurrence-free survival after hepatectomy. Hepatology 2012, 56, 2231–2241. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Liu, S.; Cheng, Y.; Lu, L.; Shi, J.; Xu, G.; Li, N.; Cheng, K.; Wu, M.; Cheng, S.; et al. ICAM-1-Related Noncoding RNA in Cancer Stem Cells Maintains ICAM-1 Expression in Hepatocellular Carcinoma. Clin. Cancer Res. 2016, 22, 2041–2050. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, M.; Gross, M.; Goyal, A.; Polycarpou-Schwarz, M.; Miersch, T.; Ernst, A.S.; Leupold, J.; Patil, N.; Warnken, U.; Allgayer, H.; et al. The Long Noncoding RNA Cancer Susceptibility 9 and RNA Binding Protein Heterogeneous Nuclear Ribonucleoprotein L Form a Complex and Coregulate Genes Linked to AKT Signaling. Hepatology 2018, 68, 1817–1832. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Huan, L.; Wang, J.; Wu, Y.; Huang, S.; He, X. LncRNA RP11-295G20.2 regulates hepatocellular carcinoma cell growth and autophagy by targeting PTEN to lysosomal degradation. Cell Discov. 2021, 7, 118. [Google Scholar] [CrossRef]
- Unfried, J.P.; Marin-Baquero, M.; Rivera-Calzada, A.; Razquin, N.; Martin-Cuevas, E.M.; de Braganca, S.; Aicart-Ramos, C.; McCoy, C.; Prats-Mari, L.; Arribas-Bosacoma, R.; et al. Long Noncoding RNA NIHCOLE Promotes Ligation Efficiency of DNA Double-Strand Breaks in Hepatocellular Carcinoma. Cancer Res. 2021, 81, 4910–4925. [Google Scholar] [CrossRef]
- Xia, A.; Yuan, W.; Wang, Q.; Xu, J.; Gu, Y.; Zhang, L.; Chen, C.; Wang, Z.; Wu, D.; He, Q.; et al. The cancer-testis lncRNA lnc-CTHCC promotes hepatocellular carcinogenesis by binding hnRNP K and activating YAP1 transcription. Nat. Cancer 2022, 3, 203–218. [Google Scholar] [CrossRef]
- Yu, A.T.; Berasain, C.; Bhatia, S.; Rivera, K.; Liu, B.; Rigo, F.; Pappin, D.J.; Spector, D.L. PHAROH lncRNA regulates Myc translation in hepatocellular carcinoma via sequestering TIAR. eLife 2021, 10, e68263. [Google Scholar] [CrossRef]
- Tang, X.; Feng, D.; Li, M.; Zhou, J.; Li, X.; Zhao, D.; Hao, B.; Li, D.; Ding, K. Transcriptomic Analysis of mRNA-lncRNA-miRNA Interactions in Hepatocellular Carcinoma. Sci. Rep. 2019, 9, 16096. [Google Scholar] [CrossRef]
- Anwar, S.L.; Krech, T.; Hasemeier, B.; Schipper, E.; Schweitzer, N.; Vogel, A.; Kreipe, H.; Lehmann, U. Loss of imprinting and allelic switching at the DLK1-MEG3 locus in human hepatocellular carcinoma. PLoS ONE 2012, 7, e49462. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, J.; Lv, Y.; Zhang, C.; Guo, S. LncRNA meg3 suppresses hepatocellular carcinoma in vitro and vivo studies. Am. J. Transl. Res. 2019, 11, 4089–4099. [Google Scholar]
- Chang, L.; Wang, G.; Jia, T.; Zhang, L.; Li, Y.; Han, Y.; Zhang, K.; Lin, G.; Zhang, R.; Li, J.; et al. Armored long non-coding RNA MEG3 targeting EGFR based on recombinant MS2 bacteriophage virus-like particles against hepatocellular carcinoma. Oncotarget 2016, 7, 23988–24004. [Google Scholar] [CrossRef]
- Chang, L.; Li, C.; Lan, T.; Wu, L.; Yuan, Y.; Liu, Q.; Liu, Z. Decreased expression of long non-coding RNA GAS5 indicates a poor prognosis and promotes cell proliferation and invasion in hepatocellular carcinoma by regulating vimentin. Mol. Med. Rep. 2016, 13, 1541–1550. [Google Scholar] [CrossRef]
- Gao, X.; Lu, C.; Liu, Z.; Lin, Y.; Huang, J.; Lu, L.; Li, S.; Huang, X.; Tang, M.; Huang, S.; et al. RBM38 Reverses Sorafenib Resistance in Hepatocellular Carcinoma Cells by Combining and Promoting lncRNA-GAS5. Cancers 2023, 15, 2897. [Google Scholar] [CrossRef]
- Zhang, W.Y.; Zhan, H.L.; Li, M.K.; Wu, G.D.; Liu, Z.; Wu, L.F. Long noncoding RNA Gas5 induces cell apoptosis and inhibits tumor growth via activating the CHOP-dependent endoplasmic reticulum stress pathway in human hepatoblastoma HepG2 cells. J. Cell Biochem. 2022, 123, 231–247. [Google Scholar] [CrossRef]
- Zhao, P.; Cui, X.; Zhao, L.; Liu, L.; Wang, D. Overexpression of Growth-Arrest-Specific Transcript 5 Improved Cisplatin Sensitivity in Hepatocellular Carcinoma Through Sponging miR-222. DNA Cell Biol. 2020, 39, 724–732. [Google Scholar] [CrossRef]
- Wang, B.; Xian, J.; Zang, J.; Xiao, L.; Li, Y.; Sha, M.; Shen, M. Long non-coding RNA FENDRR inhibits proliferation and invasion of hepatocellular carcinoma by down-regulating glypican-3 expression. Biochem. Biophys. Res. Commun. 2019, 509, 143–147. [Google Scholar] [CrossRef]
- Grote, P.; Wittler, L.; Hendrix, D.; Koch, F.; Wahrisch, S.; Beisaw, A.; Macura, K.; Blass, G.; Kellis, M.; Werber, M.; et al. The tissue-specific lncRNA Fendrr is an essential regulator of heart and body wall development in the mouse. Dev. Cell 2013, 24, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Zhao, H.; Feng, X.; Li, H.; Qiu, C.; Yi, X.; Tang, H.; Zhang, J. Long Non-coding RNA FENDRR Acts as a miR-423-5p Sponge to Suppress the Treg-Mediated Immune Escape of Hepatocellular Carcinoma Cells. Mol. Ther. Nucleic Acids 2019, 17, 516–529. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sun, W.; Shen, W.; Xia, M.; Chen, C.; Xiang, D.; Ning, B.; Cui, X.; Li, H.; Li, X.; et al. Long non-coding RNA DILC regulates liver cancer stem cells via IL-6/STAT3 axis. J. Hepatol. 2016, 64, 1283–1294. [Google Scholar] [CrossRef]
- Zhou, C.C.; Yang, F.; Yuan, S.X.; Ma, J.Z.; Liu, F.; Yuan, J.H.; Bi, F.R.; Lin, K.Y.; Yin, J.H.; Cao, G.W.; et al. Systemic genome screening identifies the outcome associated focal loss of long noncoding RNA PRAL in hepatocellular carcinoma. Hepatology 2016, 63, 850–863. [Google Scholar] [CrossRef]
- Lin, Y.H.; Wu, M.H.; Liu, Y.C.; Lyu, P.C.; Yeh, C.T.; Lin, K.H. LINC01348 suppresses hepatocellular carcinoma metastasis through inhibition of SF3B3-mediated EZH2 pre-mRNA splicing. Oncogene 2021, 40, 4675–4685. [Google Scholar] [CrossRef]
- Ni, W.; Zhang, Y.; Zhan, Z.; Ye, F.; Liang, Y.; Huang, J.; Chen, K.; Chen, L.; Ding, Y. A novel lncRNA uc.134 represses hepatocellular carcinoma progression by inhibiting CUL4A-mediated ubiquitination of LATS1. J. Hematol. Oncol. 2017, 10, 91. [Google Scholar] [CrossRef]
- Liu, F.; Yuan, J.H.; Huang, J.F.; Yang, F.; Wang, T.T.; Ma, J.Z.; Zhang, L.; Zhou, C.C.; Wang, F.; Yu, J.; et al. Long noncoding RNA FTX inhibits hepatocellular carcinoma proliferation and metastasis by binding MCM2 and miR-374a. Oncogene 2016, 35, 5422–5434. [Google Scholar] [CrossRef]
- Song, W.; Zheng, C.; Liu, M.; Xu, Y.; Qian, Y.; Zhang, Z.; Su, H.; Li, X.; Wu, H.; Gong, P.; et al. TRERNA1 upregulation mediated by HBx promotes sorafenib resistance and cell proliferation in HCC via targeting NRAS by sponging miR-22-3p. Mol. Ther. 2021, 29, 2601–2616. [Google Scholar] [CrossRef]
- Fan, L.; Huang, X.; Chen, J.; Zhang, K.; Gu, Y.H.; Sun, J.; Cui, S.Y. Long Noncoding RNA MALAT1 Contributes to Sorafenib Resistance by Targeting miR-140-5p/Aurora-A Signaling in Hepatocellular Carcinoma. Mol. Cancer Ther. 2020, 19, 1197–1209. [Google Scholar] [CrossRef]
- Zhi, Y.; Abudoureyimu, M.; Zhou, H.; Wang, T.; Feng, B.; Wang, R.; Chu, X. FOXM1-Mediated LINC-ROR Regulates the Proliferation and Sensitivity to Sorafenib in Hepatocellular Carcinoma. Mol. Ther. Nucleic Acids 2019, 16, 576–588. [Google Scholar] [CrossRef]
- Zhou, K.; Nguyen, R.; Qiao, L.; George, J. Single cell RNA-seq analysis identifies a noncoding RNA mediating resistance to sorafenib treatment in HCC. Mol. Cancer 2022, 21, 6. [Google Scholar] [CrossRef]
- Takahashi, K.; Yan, I.K.; Wood, J.; Haga, H.; Patel, T. Involvement of extracellular vesicle long noncoding RNA (linc-VLDLR) in tumor cell responses to chemotherapy. Mol. Cancer Res. 2014, 12, 1377–1387. [Google Scholar] [CrossRef]
- Tao, H.; Zhang, Y.; Li, J.; Liu, J.; Yuan, T.; Wang, W.; Liang, H.; Zhang, E.; Huang, Z. Oncogenic lncRNA BBOX1-AS1 promotes PHF8-mediated autophagy and elicits sorafenib resistance in hepatocellular carcinoma. Mol. Ther. Oncolytics 2023, 28, 88–103. [Google Scholar] [CrossRef]
- Gao, Y.; Tong, M.; Wong, T.L.; Ng, K.Y.; Xie, Y.N.; Wang, Z.; Yu, H.; Loh, J.J.; Li, M.; Ma, S. Long Noncoding RNA URB1-Antisense RNA 1 (AS1) Suppresses Sorafenib-Induced Ferroptosis in Hepatocellular Carcinoma by Driving Ferritin Phase Separation. ACS Nano 2023, 17, 22240–22258. [Google Scholar] [CrossRef]
- Zhang, J.; Pan, T.; Zhou, W.; Zhang, Y.; Xu, G.; Xu, Q.; Li, S.; Gao, Y.; Wang, Z.; Xu, J.; et al. Long noncoding RNA LINC01132 enhances immunosuppression and therapy resistance via NRF1/DPP4 axis in hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 2022, 41, 270. [Google Scholar] [CrossRef]
- Han, Y.; Jiang, W.; Wang, Y.; Zhao, M.; Li, Y.; Ren, L. Serum long non-coding RNA SCARNA10 serves as a potential diagnostic biomarker for hepatocellular carcinoma. BMC Cancer 2022, 22, 431. [Google Scholar] [CrossRef]
- Winkle, M.; El-Daly, S.M.; Fabbri, M.; Calin, G.A. Noncoding RNA therapeutics—Challenges and potential solutions. Nat. Rev. Drug Discov. 2021, 20, 629–651. [Google Scholar] [CrossRef]
- Rebouissou, S.; Zucman-Rossi, J.; Moreau, R.; Qiu, Z.; Hui, L. Note of caution: Contaminations of hepatocellular cell lines. J. Hepatol. 2017, 67, 896–897. [Google Scholar] [CrossRef]
- DiStefano, J.K.; Gerhard, G.S. Long Noncoding RNAs and Human Liver Disease. Annu. Rev. Pathol. 2022, 17, 1–21. [Google Scholar] [CrossRef]
- Wong, L.S.; Wei, L.; Wang, G.; Law, C.T.; Tsang, F.H.; Chin, W.C.; Ng, I.O.; Wong, C.M. In Vivo Genome-Wide CRISPR Activation Screening Identifies Functionally Important Long Noncoding RNAs in Hepatocellular Carcinoma. Cell Mol. Gastroenterol. Hepatol. 2022, 14, 1053–1076. [Google Scholar] [CrossRef]
- Li, G.Z.; Meng, G.X.; Pan, G.Q.; Zhang, X.; Yan, L.J.; Li, R.Z.; Ding, Z.N.; Tan, S.Y.; Wang, D.X.; Tian, B.W.; et al. MALAT1/ mir-1-3p mediated BRF2 expression promotes HCC progression via inhibiting the LKB1/AMPK signaling pathway. Cancer Cell Int. 2023, 23, 188. [Google Scholar] [CrossRef]
- Wang, Y.; Zhu, P.; Luo, J.; Wang, J.; Liu, Z.; Wu, W.; Du, Y.; Ye, B.; Wang, D.; He, L.; et al. LncRNA HAND2-AS1 promotes liver cancer stem cell self-renewal via BMP signaling. EMBO J. 2019, 38, e101110. [Google Scholar] [CrossRef]
- Chakraborty, E.; Sarkar, D. Emerging Therapies for Hepatocellular Carcinoma (HCC). Cancers 2022, 14, 2798. [Google Scholar] [CrossRef]
- Rajasekaran, D.; Srivastava, J.; Ebeid, K.; Gredler, R.; Akiel, M.; Jariwala, N.; Robertson, C.L.; Shen, X.N.; Siddiq, A.; Fisher, P.B.; et al. Combination of Nanoparticle-Delivered siRNA for Astrocyte Elevated Gene-1 (AEG-1) and All-trans Retinoic Acid (ATRA): An Effective Therapeutic Strategy for Hepatocellular Carcinoma (HCC). Bioconjug. Chem. 2015, 26, 1651–1661. [Google Scholar] [CrossRef]
- Srivastava, J.; Robertson, C.L.; Ebeid, K.; Dozmorov, M.; Rajasekaran, D.; Mendoza, R.; Siddiq, A.; Akiel, M.A.; Jariwala, N.; Shen, X.N.; et al. A novel role of astrocyte elevated gene-1 (AEG-1) in regulating nonalcoholic steatohepatitis (NASH). Hepatology 2017, 66, 466–480. [Google Scholar] [CrossRef]
Figure 1.
Different types of lncRNAs based on their genomic location. See text for details. Created in BioRender.com.
Figure 1.
Different types of lncRNAs based on their genomic location. See text for details. Created in BioRender.com.

Figure 2.
Mechanism of gene regulation by lncRNAs. TF: Transcription factor. See text for details. Created in BioRender.com.
Figure 2.
Mechanism of gene regulation by lncRNAs. TF: Transcription factor. See text for details. Created in BioRender.com.
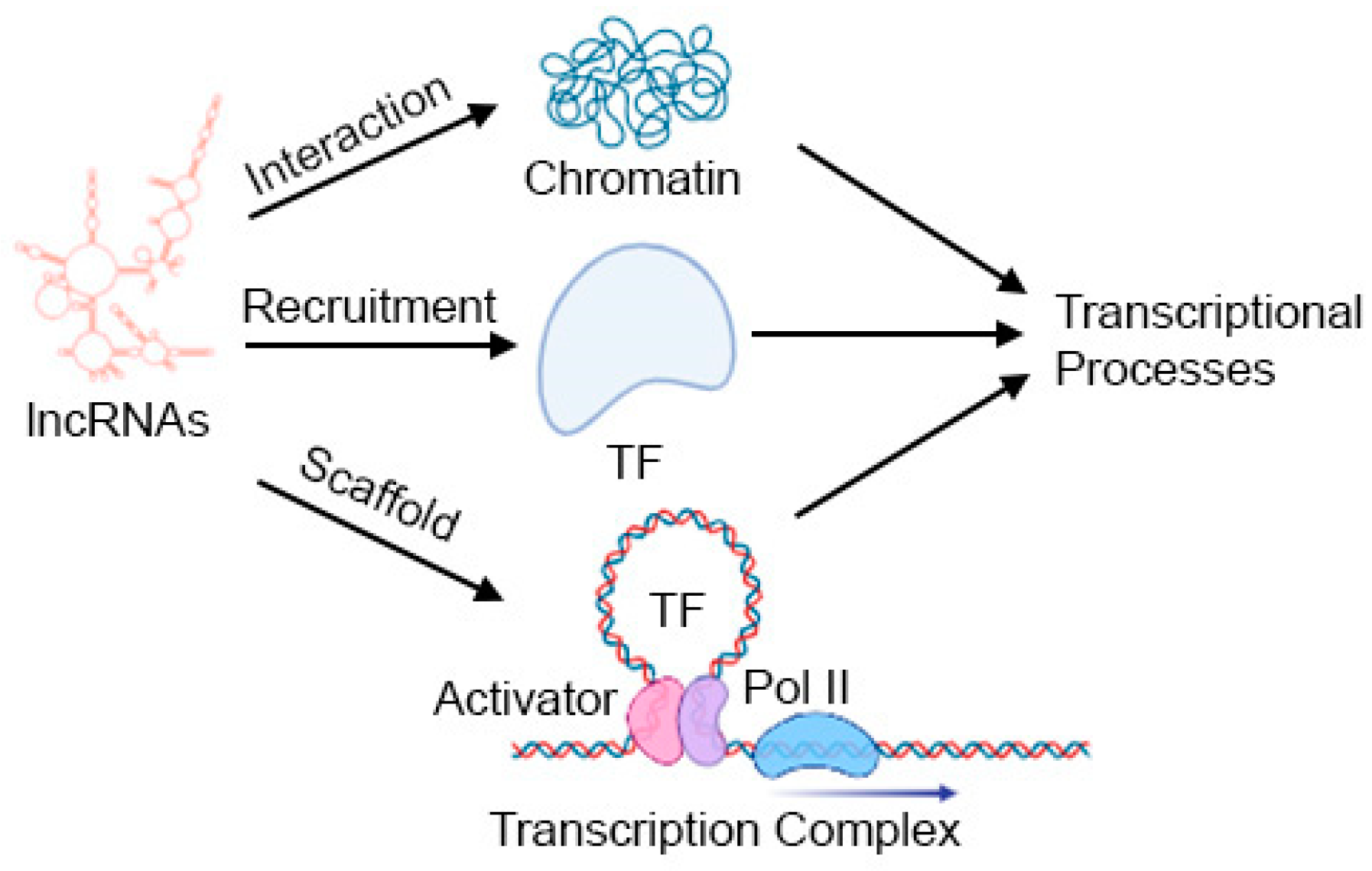
Figure 3.
Significance of studying lncRNAs in HCC. See text for details. Created in BioRender.com.
Figure 3.
Significance of studying lncRNAs in HCC. See text for details. Created in BioRender.com.
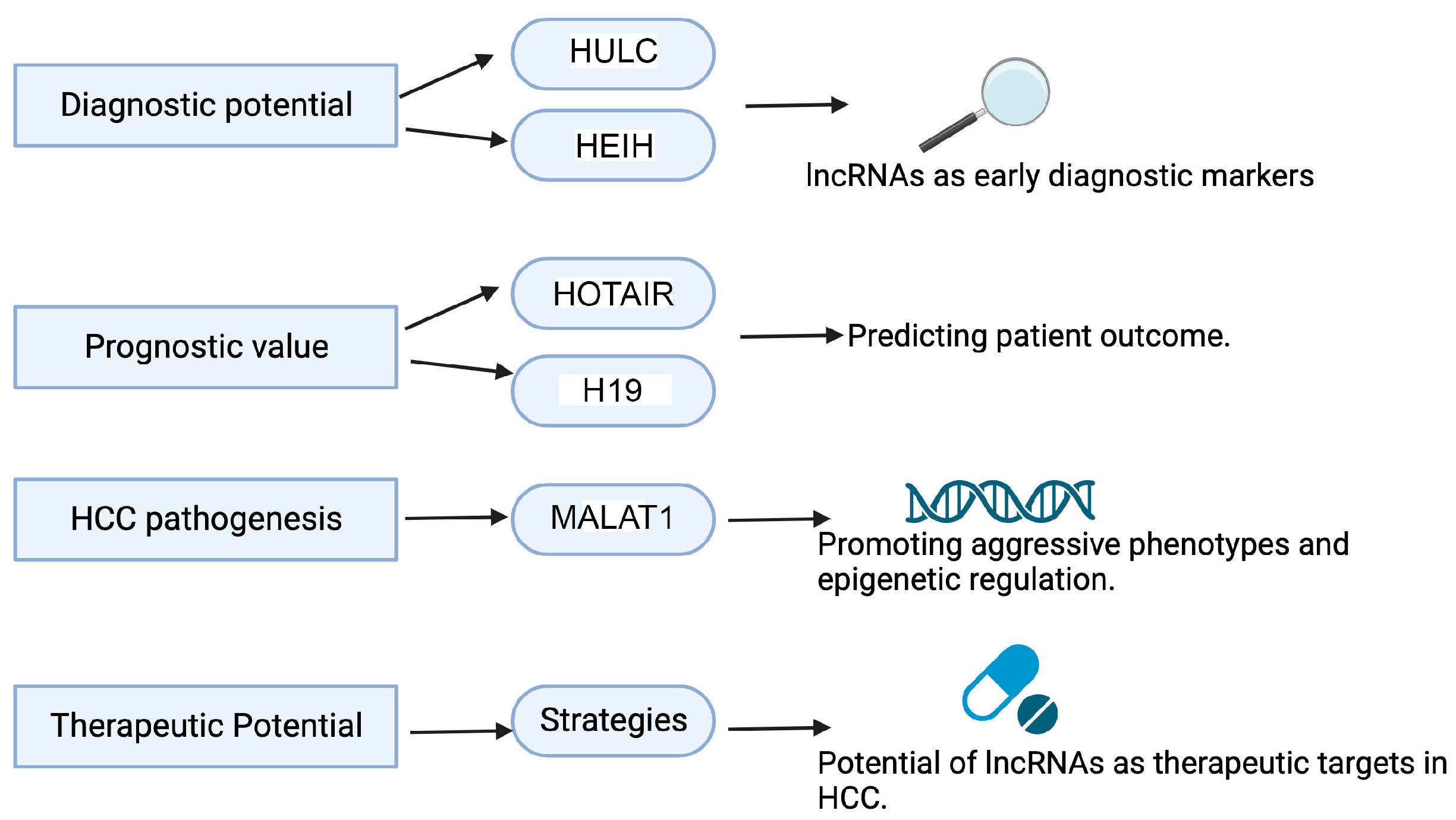

{kind=link}
{kind=link}
{kind=link}
Table 1.
LncRNAs mediating their action by LLPS.
| lncRNAs | Associated Protein | Mechanism | Phenotypes/Outcomes | References |
|---|---|---|---|---|
| DIGIT | BRD3 | Forms phase-separated condensates of BRD3, facilitating the histone H3 acetylation of lysine 18 (H3K18ac) of enhancers of endoderm transcription factors | Endoderm differentiation | [57] |
| SLERT | DDX21 | Formation of fibrillar center (FC) and dense fibrillar component (DFC) in the nucleolus | RNA polymerase I (Pol I) transcription and ribosomal RNA production | [58,59] |
| NORAD | PUM1 and PUM2 | Sequestration of Pumilio proteins in PUM condensates termed NP bodies | Inhibition of Pumilio function, preventing aberrant mitosis and maintaining genomic stability | [60] |
| SNHG9 | LATS1 | LATS1 phase separation inhibits LATS1-mediated YAP phosphorylation | Activates YAP-driven gene transcription, thus promoting breast cancer | [61] |
| MELTF-AS1 | YBX1 | Phase separation of oncogenic RBP YBX1 | Activation of ANXA8 transcription, leading to promotion of non-small-cell lung cancer (NSCLC) | [62] |
| LINP1 | Ku70/Ku80 | Multimerization of Ku to form filamentous Ku-containing aggregates | Facilitates Ku-mediated DNA repair by non-homologous end joining (NHEJ) | [63] |
| dilncRNA | DNA-damage-response (DDR) proteins, such as 53BP1 | Molecular crowding of DDR proteins in LLPS condensates | Facilitates DNA double-strand break (DSB) repair | [64] |
| XiST | PTBP1, MATR3, TDP-43 and CELF1 RBPs | Forms a condensate in the inactive X (Xi)-compartment | X-chromosome inactivation | [65] |
| GIRGL | CAPRIN1 | Sequesters CAPRIN1 and GLS1 mRNA in stress granules | Inhibition of GLS1 mRNA translation, facilitating the survival of cancer cells under glutamine-deprived conditions | [66] |
| NEAT1 | TDP-43 | Formation of nuclear bodies in response to stress | Mitigation of stress, the dysfunction of which might be a cause of ALS | [67] |
Table 2.
Role of dysregulated lncRNAs in HCC, which have been validated by multiple studies.
| lncRNAs | Expression | Role in HCC | Outcome in HCC | References |
|---|---|---|---|---|
| HOTAIR | Upregulated | Oncogene | Promotes metastasis | [38,84,85] |
| MALAT1 | Upregulated | Oncogene | Promotes proliferation, migration, invasion, and metastasis | [86,87,88] |
| HULC | Upregulated | Oncogene | Promotes proliferation and inhibits apoptosis | [85,89,90] |
| GAS5 | Downregulated | Tumor Suppressor | Suppresses proliferation and invasion | [88,91] |
| PVT1 | Upregulated | Oncogene | Promotes proliferation, migration, and invasion | [92,93,94] |
| HOTTIP | Upregulated | Oncogene | Promotes proliferation and angiogenesis | [95,96,97] |
| MVIH | Upregulated | Oncogene | Promotes proliferation, invasion, metastasis and angiogenesis | [98,99] |
| H19 | Upregulated | Oncogene | Promotes proliferation, metastasis and angiogenesis | [99,100,101] |
| MEG3 | Downregulated | Tumor suppressor | Inhibits proliferation, migration and invasion | [102,103] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shah, M.; Sarkar, D. HCC-Related lncRNAs: Roles and Mechanisms. Int. J. Mol. Sci. 2024, 25, 597. https://doi.org/10.3390/ijms25010597
AMA Style
Shah M, Sarkar D. HCC-Related lncRNAs: Roles and Mechanisms. International Journal of Molecular Sciences. 2024; 25(1):597. https://doi.org/10.3390/ijms25010597
Chicago/Turabian StyleShah, Mimansha, and Devanand Sarkar. 2024. "HCC-Related lncRNAs: Roles and Mechanisms" International Journal of Molecular Sciences 25, no. 1: 597. https://doi.org/10.3390/ijms25010597
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.