
Human T-Cell Responses to Metallic Ion-Doped Bioactive Glasses

 , ,
, ,  , , ,
, , ,  ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Silver, but Not Copper or Tellurium Ion Doping Induces Apoptosis of PBMCs
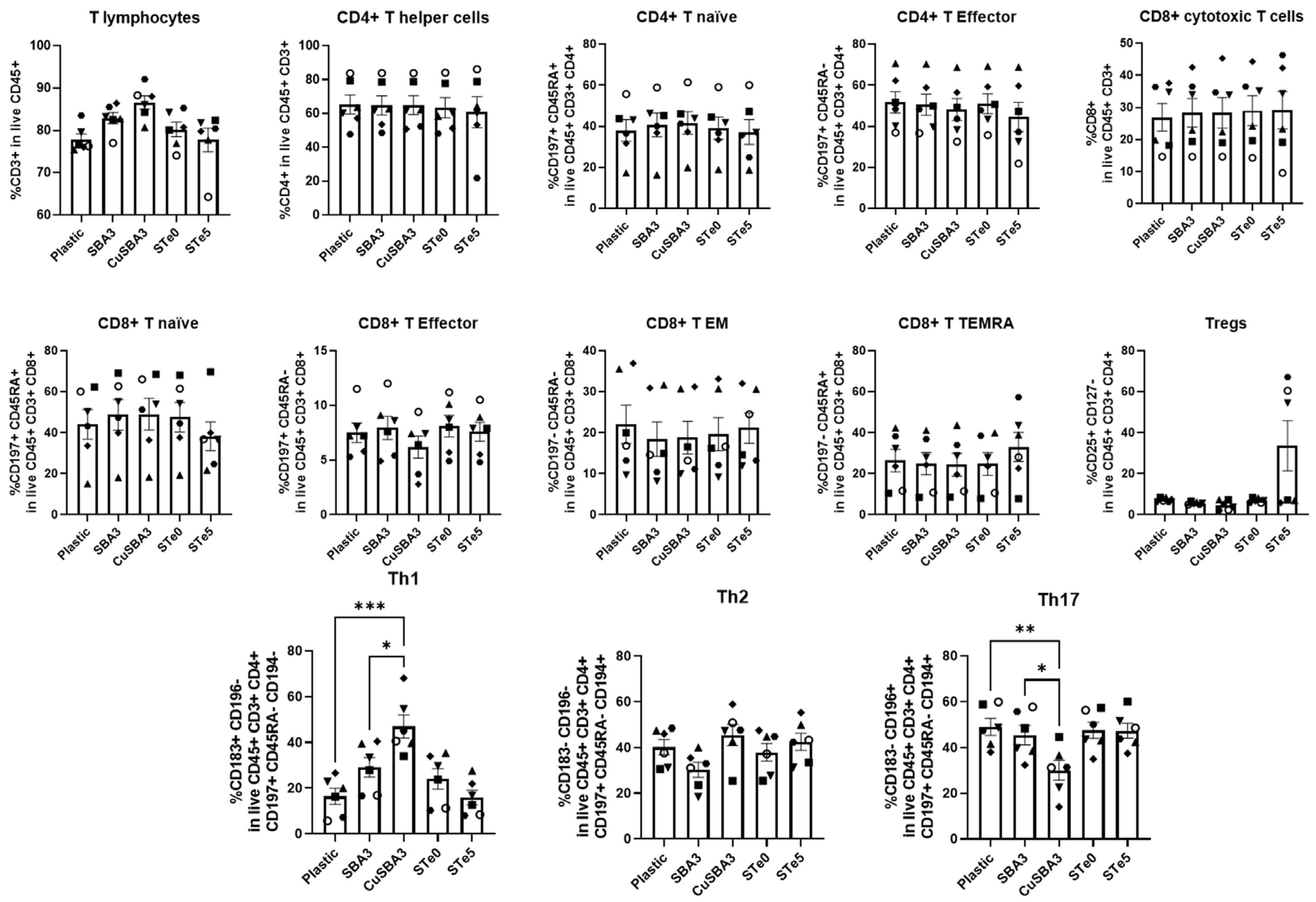
2.2. Immunophenotyping Reveals Th1/Th17 Shift Linked with Copper but Not Tellurium-Doping
2.3. Cytokine Profile Response to Ion-Doped Bioactive Glasses
3. Discussion
4. Materials and Methods
4.1. Bioactive Glasses Preparation
4.2. Ion Release in Simulated Body Fluid (SBF) and Cell Medium
4.3. Blood Specimen Collection
4.4. Peripheral Blood Mononuclear Cells (PBMCs) Isolation
4.5. Assessment of Cell Viability via Flow Cytometry
4.6. Immunobiocompatibility Assay
4.7. Enzyme-Linked Immunosorbent Assay (ELISA)
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abreu, H.; Canciani, E.; Raineri, D.; Cappellano, G.; Rimondini, L.; Chiocchetti, A. Extracellular Vesicles in Musculoskeletal Regeneration: Modulating the Therapy of the Future. Cells 2022, 11, 43. [Google Scholar] [CrossRef] [PubMed]
- Cieza, A.; Causey, K.; Kamenov, K.; Hanson, S.W.; Chatterji, S.; Vos, T. Global Estimates of the Need for Rehabilitation Based on the Global Burden of Disease Study 2019: A Systematic Analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 2006–2017. [Google Scholar] [CrossRef]
- Shen, Y.; Huang, X.; Wu, J.; Lin, X.; Zhou, X.; Zhu, Z.; Pan, X.; Xu, J.; Qiao, J.; Zhang, T.; et al. The Global Burden of Osteoporosis, Low Bone Mass, and Its Related Fracture in 204 Countries and Territories, 1990–2019. Front. Endocrinol. 2022, 13, 882241. [Google Scholar] [CrossRef]
- Agrawal, R.; Kumar, A.; Mohammed, M.K.A.; Singh, S. Biomaterial Types, Properties, Medical Applications, and Other Factors: A Recent Review. J. Zhejiang Univ. Sci. A 2023, 24, 1027–1042. [Google Scholar] [CrossRef]
- Freitas, J.; Santos, S.G.; Gonçalves, R.M.; Teixeira, J.H.; Barbosa, M.A.; Almeida, M.I. Genetically Engineered-MSC Therapies for Non-Unions, Delayed Unions and Critical-Size Bone Defects. Int. J. Mol. Sci. 2019, 20, 3430. [Google Scholar] [CrossRef]
- Salinas, A.J.; Vallet-Regí, M. Bioactive Ceramics: From Bone Grafts to Tissue Engineering. RSC Adv. 2013, 3, 11116–11131. [Google Scholar] [CrossRef]
- Pina, S.; Reis, R.L.; Oliveira, J.M. Ceramic biomaterials for tissue engineering. In Fundamental Biomaterials: Ceramics; Chapter 4; Elsevier: Maryland Heights, MO, USA, 2018. [Google Scholar]
- Negut, I.; Ristoscu, C. Bioactive Glasses for Soft and Hard Tissue Healing Applications—A Short Review. Appl. Sci. 2023, 13, 6151. [Google Scholar] [CrossRef]
- Hench, L.L.; Splinter, R.J.; Allen, W.C.; Greenlee, T.K. Bonding Mechanisms at the Interface of Ceramic Prosthetic Materials. J. Biomed. Mater. Res. 1971, 5, 117–141. [Google Scholar] [CrossRef]
- Hench, L.L. The Story of Bioglass®. J. Mater. Sci. Mater. Med. 2006, 17, 967–980. [Google Scholar] [CrossRef]
- Ali, S.; Farooq, I.; Iqbal, K. A Review of the Effect of Various Ions on the Properties and the Clinical Applications of Novel Bioactive Glasses in Medicine and Dentistry. Saudi Dent. J. 2014, 26, 1–5. [Google Scholar] [CrossRef]
- Miguez-Pacheco, V.; Hench, L.L.; Boccaccini, A.R. Bioactive Glasses beyond Bone and Teeth: Emerging Applications in Contact with Soft Tissues. Acta Biomater. 2015, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Cannio, M.; Bellucci, D.; Roether, J.A.; Boccaccini, D.N.; Cannillo, V. Bioactive Glass Applications: A Literature Review of Human Clinical Trials. Materials 2021, 14, 5440. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Kumar, V.; Baino, F.; Mauro, J.C.; Pickrell, G.; Evans, I.; Bretcanu, O. Mechanical Properties of Bioactive Glasses, Ceramics, Glass-Ceramics and Composites: State-of-the-Art Review and Future Challenges. Mater. Sci. Eng. C 2019, 104, 109895. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Pickrell, G.; Sriranganathan, N.; Kumar, V.; Homa, D. Review and the State of the Art: Sol–Gel and Melt Quenched Bioactive Glasses for Tissue Engineering. J. Biomed. Mater. Res.—Part B Appl. Biomater. 2016, 104, 1248–1275. [Google Scholar] [CrossRef] [PubMed]
- Ciraldo, F.E.; Boccardi, E.; Melli, V.; Westhauser, F.; Boccaccini, A.R. Tackling Bioactive Glass Excessive in Vitro Bioreactivity: Preconditioning Approaches for Cell Culture Tests. Acta Biomater. 2018, 75, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.F. On the Mechanisms of Biocompatibility. Biomaterials 2008, 29, 2941–2953. [Google Scholar] [CrossRef] [PubMed]
- Franz, S.; Rammelt, S.; Scharnweber, D.; Simon, J.C. Immune Responses to Implants—A Review of the Implications for the Design of Immunomodulatory Biomaterials. Biomaterials 2011, 32, 6692–6709. [Google Scholar] [CrossRef] [PubMed]
- Cappellano, G.; Ploner, C.; Lobenwein, S.; Sopper, S.; Hoertnagl, P.; Mayerl, C.; Wick, N.; Pierer, G.; Wick, G.; Wolfram, D. Immunophenotypic Characterization of Human T Cells after in Vitro Exposure to Different Silicone Breast Implant Surfaces. PLoS ONE 2018, 13, e0192108. [Google Scholar] [CrossRef] [PubMed]
- Chraniuk, M.; Panasiuk, M.; Hovhannisyan, L.; Żołędowska, S.; Nidzworski, D.; Ciołek, L.; Woźniak, A.; Jaegermann, Z.; Biernat, M.; Gromadzka, B. The Preliminary Assessment of New Biomaterials Necessitates a Comparison of Direct and Indirect Cytotoxicity Methodological Approaches. Polymers 2022, 14, 4522. [Google Scholar] [CrossRef]
- Zheng, K.; Niu, W.; Lei, B.; Boccaccini, A.R. Immunomodulatory Bioactive Glasses for Tissue Regeneration. Acta Biomater. 2021, 133, 168–186. [Google Scholar] [CrossRef]
- Owen, J.A.; Punt, J.; Stranford, S.A.; Jones, P.P.; Kuby, J. Kuby Immunology, 8th ed.; W.H. Freeman: New York, NY, USA, 2019. [Google Scholar]
- Lozano-Ojalvo, D.; López-Fandiño, R.; López-Expósito, I. PBMC-derived T cells. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Chapter 16; Springer: Cham, Switzerland, 2015. [Google Scholar]
- Zhao, Z.; Zhao, Q.; Gu, B.; Yin, C.; Shen, K.; Tang, H.; Xia, H.; Zhang, X.; Zhao, Y.; Yang, X.; et al. Minimally Invasive Implantation and Decreased Inflammation Reduce Osteoinduction of Biomaterial. Theranostics 2020, 10, 3533. [Google Scholar] [CrossRef]
- Khassawna, T.E.; Serra, A.; Bucher, C.H.; Petersen, A.; Schlundt, C.; Könnecke, I.; Malhan, D.; Wendler, S.; Schell, H.; Volk, H.D.; et al. T Lymphocytes Influence the Mineralization Process of Bone. Front. Immunol. 2017, 8, 562. [Google Scholar] [CrossRef] [PubMed]
- Grassi, F.; Cattini, L.; Gambari, L.; Manferdini, C.; Piacentini, A.; Gabusi, E.; Facchini, A.; Lisignoli, G. T Cell Subsets Differently Regulate Osteogenic Differentiation of Human Mesenchymal Stromal Cells in Vitro. J. Tissue Eng. Regen. Med. 2016, 10, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Cochis, A.; Barberi, J.; Ferraris, S.; Miola, M.; Rimondini, L.; Vernè, E.; Yamaguchi, S.; Spriano, S. Competitive Surface Colonization of Antibacterial and Bioactive Materials Doped with Strontium and/or Silver Ions. Nanomaterials 2020, 10, 120. [Google Scholar] [CrossRef] [PubMed]
- Miola, M.; Verné, E. Bioactive and Antibacterial Glass Powders Doped with Copper by Ion-Exchange in Aqueous Solutions. Materials 2016, 9, 405. [Google Scholar] [CrossRef] [PubMed]
- Tang, A.; Ren, Q.; Wu, Y.; Wu, C.; Cheng, Y. Investigation into the Antibacterial Mechanism of Biogenic Tellurium Nanoparticles and Precursor Tellurite. Int. J. Mol. Sci. 2022, 23, 11697. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.; Lim, D.H.; Choi, I.H.; Kang, T.; Lee, K.; Moon, E.Y.; Yang, Y.; Lee, M.S.; Lim, J.S. Vascular Tube Formation and Angiogenesis Induced by Polyvinylpyrrolidone-Coated Silver Nanoparticles. Toxicol. Lett. 2011, 205, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Kang, Y. Role of Copper in Angiogenesis and Its Medicinal Implications. Curr. Med. Chem. 2009, 16, 1304–1314. [Google Scholar] [CrossRef] [PubMed]
- Miola, M.; Massera, J.; Cochis, A.; Kumar, A.; Rimondini, L.; Vernè, E. Tellurium: A New Active Element for Innovative Multifunctional Bioactive Glasses. Mater. Sci. Eng. C 2021, 123, 111957. [Google Scholar] [CrossRef]
- Lallukka, M.; Houaoui, A.; Miola, M.; Miettinen, S.; Massera, J.; Verné, E. In Vitro Cytocompatibility of Antibacterial Silver and Copper-Doped Bioactive Glasses. Ceram. Int. 2023, 49, 36044–36055. [Google Scholar] [CrossRef]
- Ferraris, S.; Yamaguchi, S.; Barbani, N.; Cazzola, M.; Cristallini, C.; Miola, M.; Vernè, E.; Spriano, S. Bioactive Materials: In Vitro Investigation of Different Mechanisms of Hydroxyapatite Precipitation. Acta Biomater. 2020, 102, 468–480. [Google Scholar] [CrossRef]
- Lallukka, M.; Miola, M.; Najmi, Z.; Cochis, A.; Spriano, S.; Rimondini, L.; Verné, E. Cu-Doped Bioactive Glass with Enhanced in Vitro Bioactivity and Antibacterial Properties. Ceram. Int. 2024, 50, 5091–5103. [Google Scholar] [CrossRef]
- Mehrabi, T.; Mesgar, A.S.; Mohammadi, Z. Bioactive Glasses: A Promising Therapeutic Ion Release Strategy for Enhancing Wound Healing. ACS Biomater. Sci. Eng. 2020, 6, 5399–5430. [Google Scholar] [CrossRef]
- Pajares-Chamorro, N.; Wagley, Y.; Maduka, C.V.; Youngstrom, D.W.; Yeger, A.; Badylak, S.F.; Hammer, N.D.; Hankenson, K.; Chatzistavrou, X. Silver-Doped Bioactive Glass Particles for in Vivo Bone Tissue Regeneration and Enhanced Methicillin-Resistant Staphylococcus Aureus (MRSA) Inhibition. Mater. Sci. Eng. C 2021, 120, 111693. [Google Scholar] [CrossRef]
- Bari, A.; Bloise, N.; Fiorilli, S.; Novajra, G.; Vallet-Regí, M.; Bruni, G.; Torres-Pardo, A.; González-Calbet, J.M.; Visai, L.; Vitale-Brovarone, C. Copper-Containing Mesoporous Bioactive Glass Nanoparticles as Multifunctional Agent for Bone Regeneration. Acta Biomater. 2017, 55, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hu, M.; Zhang, W.; Zhang, X. Construction of Tellurium-Doped Mesoporous Bioactive Glass Nanoparticles for Bone Cancer Therapy by Promoting ROS-Mediated Apoptosis and Antibacterial Activity. J. Colloid Interface Sci. 2022, 610, 719–730. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.F.; Prohaska, J.R.; Knutson, M.D. Metabolic Crossroads of Iron and Copper. Nutr. Rev. 2010, 68, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Haase, H.; Fahmi, A.; Mahltig, B. Impact of Silver Nanoparticles and Silver Ions on Innate Immune Cells. J. Biomed. Nanotechnol. 2014, 10, 1146–1156. [Google Scholar] [CrossRef]
- Sandoval, J.M.; Verrax, J.; Vásquez, C.C.; Calderon, P.B. A Comparative Study of Tellurite Toxicity in Normal and Cancer Cells. Mol. Cell. Toxicol. 2012, 8, 327–334. [Google Scholar] [CrossRef]
- Hardes, J.; Streitburger, A.; Ahrens, H.; Nusselt, T.; Gebert, C.; Winkelmann, W.; Battmann, A.; Gosheger, G. The Influence of Elementary Silver versus Titanium on Osteoblasts Behaviour in Vitro Using Human Osteosarcoma Cell Lines. Sarcoma 2007, 2007, 026539. [Google Scholar] [CrossRef]
- Gosheger, G.; Hardes, J.; Ahrens, H.; Streitburger, A.; Buerger, H.; Erren, M.; Gunsel, A.; Kemper, F.H.; Winkelmann, W.; Von Eiff, C. Silver-Coated Megaendoprostheses in a Rabbit Model—An Analysis of the Infection Rate and Toxicological Side Effects. Biomaterials 2004, 25, 5547–5556. [Google Scholar] [CrossRef]
- Wan, A.T.; Conyers, R.A.J.; Coombs, C.J.; Masterton, J.P. Determination of Silver in Blood, Urine, and Tissues of Volunteers and Burn Patients. Clin. Chem. 1991, 37, 1683–1687. [Google Scholar] [CrossRef]
- Choban, P.S.; Marshall, W.J. Leukopenia Secondary to Silver Sulfadiazine: Frequency, Characteristics and Clinical Consequences. Am. Surg. 1987, 53, 515–517. [Google Scholar]
- Tozzi, P.; Al-Darweesh, A.; Vogt, P.; Stumpe, F. Silver-Coated Prosthetic Heart Valve: A Double-Bladed Weapon. Eur. J. Cardio-Thoracic Surg. 2001, 19, 729–731. [Google Scholar] [CrossRef]
- McCauley, R.L.; Linares, H.A.; Pelligrini, V.; Herndon, D.N.; Robson, M.C.; Heggers, J.P. In Vitro Toxicity of Topical Antimicrobial Agents to Human Fibroblasts. J. Surg. Res. 1989, 46, 267–274. [Google Scholar] [CrossRef]
- Kuroyanagi, Y.; Kim, E.; Shioya, N. Evaluation of a Synthetic Wound Dressing Capable of Releasing Silver Sulfadiazine. J. Burn Care Rehabil. 1991, 12, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Dhinasekaran, D.; Kumar, A. Fabrication of bioactive structures from sol-gel derived bioactive glass. In Bioactive Glasses and Glass-Ceramics: Fundamentals and Applications; Chapter 6; Wiley: Hoboken, NJ, USA, 2022. [Google Scholar]
- Adusei, K.M.; Ngo, T.B.; Sadtler, K. T Lymphocytes as Critical Mediators in Tissue Regeneration, Fibrosis, and the Foreign Body Response. Acta Biomater. 2021, 133, 17–33. [Google Scholar] [CrossRef] [PubMed]
- Wolfram, D.; Rabensteiner, E.; Grundtman, C.; Böck, G.; Mayerl, C.; Parson, W.; Almanzar, G.; Hasenöhrl, C.; Piza-Katzer, H.; Wick, G. T Regulatory Cells and TH17 Cells in Peri-Silicone Implant Capsular Fibrosis. Plast. Reconstr. Surg. 2012, 129, 327e–337e. [Google Scholar] [CrossRef] [PubMed]
- Dey, A.; Manna, S.; Kumar, S.; Chattopadhyay, S.; Saha, B.; Roy, S. Immunostimulatory Effect of Chitosan Conjugated Green Copper Oxide Nanoparticles in Tumor Immunotherapy. Cytokine 2020, 127, 154958. [Google Scholar] [CrossRef]
- Huang, C.; Chen, X.; Xue, Z.; Wang, T. Effect of Structure: A New Insight into Nanoparticle Assemblies from Inanimate to Animate. Sci. Adv. 2020, 6, eaba1321. [Google Scholar] [CrossRef]
- Schuhladen, K.; Stich, L.; Schmidt, J.; Steinkasserer, A.; Boccaccini, A.R.; Zinser, E. Cu, Zn Doped Borate Bioactive Glasses: Antibacterial Efficacy and Dose-Dependent: In Vitro Modulation of Murine Dendritic Cells. Biomater. Sci. 2020, 8, 2143–2155. [Google Scholar] [CrossRef] [PubMed]
- Takatsu, K. Interleukin-5 and IL-5 Receptor in Health and Diseases. Proc. Japan Acad. Ser. B Phys. Biol. Sci. 2011, 87, 463–485. [Google Scholar] [CrossRef] [PubMed]
- Pelaia, C.; Paoletti, G.; Puggioni, F.; Racca, F.; Pelaia, G.; Canonica, G.W.; Heffler, E. Interleukin-5 in the Pathophysiology of Severe Asthma. Front. Physiol. 2019, 10, 1514. [Google Scholar] [CrossRef] [PubMed]
- Marone, G.; Granata, F.; Pucino, V.; Pecoraro, A.; Heffler, E.; Loffredo, S.; Scadding, G.W.; Varricchi, G. The Intriguing Role of Interleukin 13 in the Pathophysiology of Asthma. Front. Pharmacol. 2019, 10, 01387. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.Y.; Sur, S.; Grant, J.A.; Tripple, J.W. Interleukin-4/Interleukin-13 versus Interleukin-5: A Comparison of Molecular Targets in Biologic Therapy for the Treatment of Severe Asthma. Curr. Opin. Allergy Clin. Immunol. 2019, 19, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Asikainen, A.J.; Hagström, J.; Sorsa, T.; Noponen, J.; Kellomäki, M.; Juuti, H.; Lindqvist, C.; Hietanen, J.; Suuronen, R. Soft Tissue Reactions to Bioactive Glass 13-93 Combined with Chitosan. J. Biomed. Mater. Res.—Part A 2007, 83, 530–537. [Google Scholar] [CrossRef] [PubMed]
- Gschwandtner, M.; Derler, R.; Midwood, K.S. More Than Just Attractive: How CCL2 Influences Myeloid Cell Behavior Beyond Chemotaxis. Front. Immunol. 2019, 10, 491100. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Ma, L.; Jiang, C.; Zhang, S. Expression and Potential Role of CCL4 in CD8+T Cells in NSCLC. Clin. Transl. Oncol. 2022, 24, 2420–2431. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.D.; Voyich, J.M.; Burlak, C.; DeLeo, F.R. Neutrophils in the Innate Immune Response. Arch. Immunol. Ther. Exp. 2005, 53, 505. [Google Scholar]
- Jones, J.A.; Chang, D.T.; Meyerson, H.; Colton, E.; Il, K.K.; Matsuda, T.; Anderson, J.M. Proteomic Analysis and Quantification of Cytokines and Chemokines from Biomaterial Surface-Adherent Macrophages and Foreign Body Giant Cells. J. Biomed. Mater. Res.—Part A 2007, 83, 585–596. [Google Scholar] [CrossRef]
- Chang, D.T.; Jones, J.A.; Meyerson, H.; Colton, E.; Il, K.K.; Matsuda, T.; Anderson, J.M. Lymphocyte/Macrophage Interactions: Biomaterial Surface-Dependent Cytokine, Chemokine, and Matrix Protein Production. J. Biomed. Mater. Res.—Part A 2008, 87, 676–687. [Google Scholar] [CrossRef] [PubMed]
- Wick, G.; Grundtman, C.; Mayerl, C.; Wimpissinger, T.F.; Feichtinger, J.; Zelger, B.; Sgonc, R.; Wolfram, D. The Immunology of Fibrosis. Annu. Rev. Immunol. 2013, 31, 107–135. [Google Scholar] [CrossRef] [PubMed]
- Love, R.J.; Jones, K.S. Biomaterials, Fibrosis, and the Use of Drug Delivery Systems in Future Antifibrotic Strategies. Crit. Rev. Biomed. Eng. 2009, 37, 259–281. [Google Scholar] [CrossRef] [PubMed]
- Steen, E.H.; Wang, X.; Balaji, S.; Butte, M.J.; Bollyky, P.L.; Keswani, S.G. The Role of the Anti-Inflammatory Cytokine Interleukin-10 in Tissue Fibrosis. Adv. Wound Care 2020, 9, 184–198. [Google Scholar] [CrossRef] [PubMed]
- Varmette, E.A.; Nowalk, J.R.; Flick, L.M.; Hall, M.M. Abrogation of the Inflammatory Response in LPS-Stimulated RAW 264.7 Murine Macrophages by Zn- and Cu-Doped Bioactive Sol-Gel Glasses. J. Biomed. Mater. Res.—Part A 2009, 90, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Barbeck, M.; Alkildani, S.; Mandlule, A.; Radenković, M.; Najman, S.; Stojanović, S.; Jung, O.; Ren, Y.; Cai, B.; Görke, O.; et al. In Vivo Analysis of the Immune Response to Strontium- and Copper-Doped Bioglass. In Vivo 2022, 36, 2149–2165. [Google Scholar] [CrossRef] [PubMed]
- Weng, L.; Boda, S.K.; Teusink, M.J.; Shuler, F.D.; Li, X.; Xie, J. Binary Doping of Strontium and Copper Enhancing Osteogenesis and Angiogenesis of Bioactive Glass Nanofibers While Suppressing Osteoclast Activity. ACS Appl. Mater. Interfaces 2017, 9, 24484–24496. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Deng, C.; Li, X.; Liu, Y.; Zhang, M.; Qin, C.; Yao, Q.; Wang, L.; Wu, C. Copper-Incorporated Bioactive Glass-Ceramics Inducing Anti-Inflammatory Phenotype and Regeneration of Cartilage/Bone Interface. Theranostics 2019, 9, 6300. [Google Scholar] [CrossRef] [PubMed]
- Rau, J.V.; Curcio, M.; Raucci, M.G.; Barbaro, K.; Fasolino, I.; Teghil, R.; Ambrosio, L.; De Bonis, A.; Boccaccini, A.R. Cu-Releasing Bioactive Glass Coatings and Their in Vitro Properties. ACS Appl. Mater. Interfaces 2019, 11, 5812–5820. [Google Scholar] [CrossRef]
- Chen, Y.; Guan, M.; Ren, R.; Gao, C.; Cheng, H.; Li, Y.; Gao, B.; Wei, Y.; Fu, J.; Sun, J.; et al. Improved Immunoregulation of Ultra-Low-Dose Silver Nanoparticle-Loaded TiO2 Nanotubes via M2 Macrophage Polarization by Regulating GLUT1 and Autophagy. Int. J. Nanomedicine 2020, 15, 2011–2026. [Google Scholar] [CrossRef]
- Kokubo, T.; Takadama, H. How Useful Is SBF in Predicting in Vivo Bone Bioactivity? Biomaterials 2006, 27, 2907–2915. [Google Scholar] [CrossRef] [PubMed]
- Breen, E.J.; Tan, W.; Khan, A. The Statistical Value of Raw Fluorescence Signal in Luminex XMAP Based Multiplex Immunoassays. Sci. Rep. 2016, 6, 26996. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| AgSBA2 | CuSBA3 | STe5 | |
|---|---|---|---|
| Doping method | The surface of the glass (Ion exchange in aqueous solution of AgNO3) | The surface of the glass (Ion exchange in aqueous solution of Cu (CO2CH3)2) | The bulk of the glass (TeO2 in the glass network as an oxide) |
| Doped element content at-% (EDS) | 0.7 ± 0.36 | 8.4 ± 0.18 | 3.4 ± 0.08 |
| Doped ion leaching after 3 days in cell medium | 7.9 ± 1.4 ppm * (ICP-OES) | 11.0 ± 2.4 ppm * (ICP-OES) | 5.6 ± 0.3 ppm ** (ICP-OES) |
| Doped ion leaching after 3 days in Simulated Body Fluid (SBF) | 0.37 ± 0.13 ppm (ICP-MS) | 0.14 ± 0.04 ppm (ICP-MS) | 0.21 ± 0.07 ppm (ICP-MS) |
| Components | SBA2 (% mol) | SBA3 (% mol) |
|---|---|---|
| SiO2 | 48 | 48 |
| Na2O | 18 | 26 |
| CaO | 30 | 22 |
| P2O5 | 3 | 3 |
| B2O3 | 0.43 | 0.43 |
| Al2O3 | 0.57 | 0.57 |
| Components | STe0 (% mol) | STe5 (% mol) |
|---|---|---|
| SiO2 | 48.6 | 43.6 |
| Na2O | 16.7 | 16.7 |
| CaO | 34.2 | 34.2 |
| P2O5 | 0.5 | 0.5 |
| TeO2 | 0.0 | 5.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abreu, H.; Lallukka, M.; Miola, M.; Spriano, S.; Vernè, E.; Raineri, D.; Leigheb, M.; Ronga, M.; Cappellano, G.; Chiocchetti, A. Human T-Cell Responses to Metallic Ion-Doped Bioactive Glasses. Int. J. Mol. Sci. 2024, 25, 4501. https://doi.org/10.3390/ijms25084501
Abreu H, Lallukka M, Miola M, Spriano S, Vernè E, Raineri D, Leigheb M, Ronga M, Cappellano G, Chiocchetti A. Human T-Cell Responses to Metallic Ion-Doped Bioactive Glasses. International Journal of Molecular Sciences. 2024; 25(8):4501. https://doi.org/10.3390/ijms25084501
Chicago/Turabian StyleAbreu, Hugo, Mari Lallukka, Marta Miola, Silvia Spriano, Enrica Vernè, Davide Raineri, Massimiliano Leigheb, Mario Ronga, Giuseppe Cappellano, and Annalisa Chiocchetti. 2024. "Human T-Cell Responses to Metallic Ion-Doped Bioactive Glasses" International Journal of Molecular Sciences 25, no. 8: 4501. https://doi.org/10.3390/ijms25084501