
Exploring the Key Signaling Pathways and ncRNAs in Colorectal Cancer
 , ,
, ,
Abstract
:1. Introduction
2. Multiple Major Signaling Pathways in CRC
2.1. Wnt Signaling Pathway
2.2. PI3K/AKT/mTOR Signaling Pathway
2.3. MAPK Signaling Pathway
2.4. TGF-β Signaling Pathway
2.5. p53 Signaling Pathway
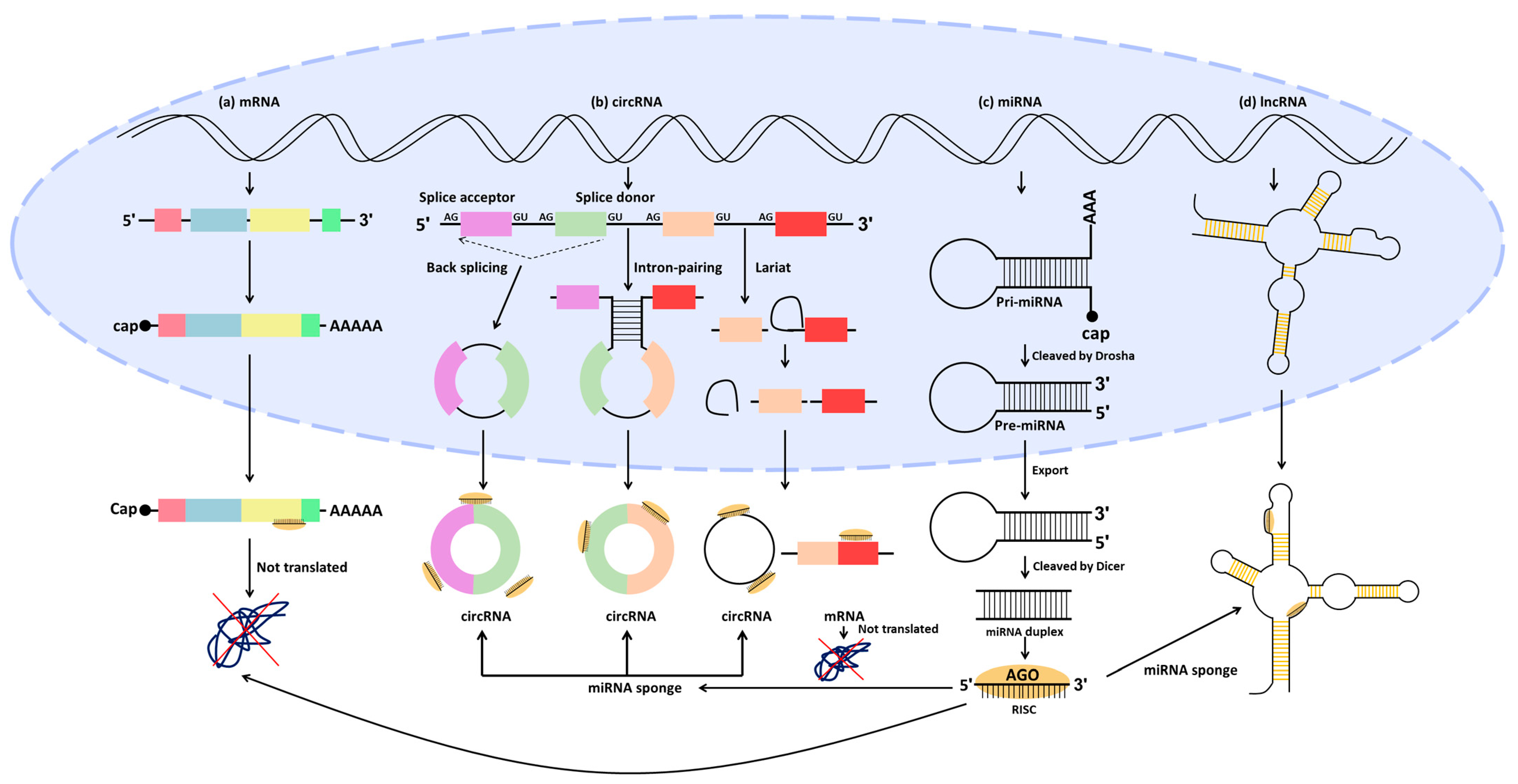
3. ncRNA Regulation in CRC
3.1. miRNA Regulation

{kind=link}
{kind=link}
{kind=link}
| miRNA Expression | miRNAs | Target Genes | Clinical Value | Impact on CRC | Function | Signaling Pathway | Human Samples | Study Model | References |
|---|---|---|---|---|---|---|---|---|---|
| UP | miR-590-5p | PDCD4 | therapeutic target | pathogenesis | induce cell viability, migration, and invasion | TGF-β signaling pathway | 30 tumor and adjacent normal tissues | in vitro | [100] |
| miR-21 | VMP1 | therapeutic target | development | promote migration and invasion, while repressing autophagy and drug sensitivity | PTEN/AKT/TFEB signaling pathway | 4 tumor and adjacent normal tissues | Oncomine dataset, in vitro | [105] | |
| miR-21-5p | TGF-β1 | therapeutic target | NA | induce pyroptosis while inhibiting cell viability | NA | 5 tumor and normal tissues | in vitro | [106] | |
| miR-452-5p | PKN2 | therapeutic target | progression | enhance cell proliferation, cell cycle transition, and chemoresistance, while suppressing apoptosis | MAPK/ERK signaling pathway | 87 tumor and matched para-carcinoma mucosal tissues | TCGA, in vitro, in vivo | [107] | |
| DUSP6 | |||||||||
| miR-106-5p | FAT4 | NA | carcinogenic | promote proliferation, migration, invasion, and angiogenesis | NA | tumor and adjacent normal tissues | TCGA, in vitro | [108] | |
| miR-496 | RASSF6 | therapeutic target | progression | induce cell motility, migration, invasion, and EMT | Wnt signaling pathway | 28 tumor and adjacent normal tissues | TCGA, in vitro | [109] | |
| miR-125b | CFTR | diagnosis biomarker and therapeutic target | metastasis | enhance migration, invasion, metastasis, and EMT | RhoA/ROCK signaling pathway | 58 tumor and adjacent normal tissues | human tissue microarray chips, in vitro, in vivo | [110] | |
| CGN | |||||||||
| miR-298 | PTEN | therapeutic target | development | facilitate cell metabolic activity, cell cycle progression, migration, and invasion, while inhibiting apoptosis | AKT/ERK and AKT/mTOR/P70 S6K signaling pathways | 100 tumors and matched adjacent normal tissues, 100 tumor and normal plasma | in vitro | [111] | |
| miR-429 | LATS2 | diagnosis biomarker and therapeutic target | tumorigenesis | stimulate proliferation and tumor growth | YAP/TAZ signaling pathway | 21 tumor and normal tissues | in vitro, in vivo | [112] | |
| miR-645 | EFNA5 | therapeutic target | metastasis | enhance migration, invasion, metastasis, and tumor growth | NA | 28 tumor and adjacent normal tissues | in vitro, in vivo | [113] | |
| DOWN | miR-150 | CTNNB1 | therapeutic target | progression | inhibit cell proliferation | Wnt signaling pathway | 30 tumor and adjacent normal tissues | in vitro, in vivo | [101] |
| miR-130a-3p | WNT1 | biomarker, therapeutic target | development | repress cell proliferation and tumor growth | Wnt signaling pathway | 30 tumor and adjacent normal tissues | TCGA, in vitro, in vivo | [102] | |
| miR-144-5p | RNF187 | diagnosis biomarker and therapeutic target | progression | suppress migration and invasion | NA | 83 tumor and adjacent normal tissues | in vitro | [103] | |
| miR-1-3p | YWHAZ | therapeutic target | progression | inhibit cell proliferation and EMT | NA | 20 tumor and adjacent normal tissues | GEO database, in vitro | [104] | |
| miR-144-3p | BCL6 | prognosis biomarker and therapeutic target | progression | repress cell proliferation and cell cycle progression | Wnt signaling pathway | 20 tumor and adjacent normal tissues | in vitro | [114] | |
| miR-450a-5p | SOX2 | progression | suppress stemness, vasculature, and tumor growth | NA | 90 glass-slide tumor tissues for array | in vitro, in vivo | [115] | ||
| miR-215-5p | CTNNBIP1 | therapeutic target | progression and metastasis | inhibit clonogenic potential, cell cycle, migration, invasion, metastasis, and tumor growth, while inducing apoptosis | Wnt signaling pathway | primary tumor, paired liver metastatic, and adjacent normal tissues | in vitro, in vivo | [116] | |
| miR-148b | p55PIK | therapeutic target | progression | suppress proliferation, tumor growth | p53 signaling pathway | 10 tumor and normal tissues | in vitro, in vivo | [117] | |
| miR-16 | Survivin | therapeutic target | NA | repress proliferation and induce apoptosis | p53 signaling pathway | 52 tumor and adjacent normal tissues | in vitro | [118] | |
| miR-139-3p | KRT80 | therapeutic target | NA | inhibit proliferation, migration, and invasion | NA | 27 tumor and normal tissues | in vitro | [119] | |
| miR-125a-5p | FNDC3B | prognosis biomarker and therapeutic target | progression | reduce proliferation | PI3K/mTOR signaling pathway | 36 tumor and adjacent normal tissues | in vitro | [120] | |
| miR-217 | |||||||||
| miR-654-3p | SRC | diagnosis and prognosis biomarker, and therapeutic target | development | repress proliferation, migration, and invasion, while promoting apoptosis | NA | 103 tumor and adjacent normal tissues | in vitro, in vivo | [121] | |
| miR-539 | TIPE | therapeutic target | progression | suppress proliferation and tumor growth, while enhancing ferroptosis | SAPK/JNK signaling pathway | 26 tumor and adjacent tissues | in vitro, in vivo | [122] | |
| miR-3622a-3p | SALL4 | therapeutic target | progression and metastasis | reduce proliferation and EMT | Wnt signaling pathway | 80 tumor and adjacent normal tissues | TCGA, in vitro, in vivo | [123] |
3.2. Competing Endogenous RNA (ceRNA) Regulation in CRC
3.2.1. LncRNA/miRNA/mRNA
| LncRNA Expression | LncRNAs | miRNAs | Genes | Clinical Value | Impact on CRC | Function | Signaling Pathway | Human Samples | Study Model | References |
|---|---|---|---|---|---|---|---|---|---|---|
| UP | EGFR-AS1 | miR-133b | STAT3 | early diagnosis biomarker | development and progression | induce cell proliferation, migration, and invasion | NA | 130 tumor and 30 normal tissues | in vitro | [142] |
| CTBP1-AS2 | miR-93-5p | TGF-β1 | NA | progression and metastasis | promote proliferation, invasion, and metastasis, while inhibiting apoptosis | TGF-β signaling pathway | 50 tumor and normal tissues | GEPIA, in vitro, in vivo | [145] | |
| SNHG16 | miR-302a-3p | AKT | therapeutic target | development | enhance proliferation | AKT signaling pathway | NA | in vitro | [146] | |
| COL4A2-AS1 | miR-20b-5p | HIF1A | biomarker and therapeutic target | progression | facilitate proliferation and aerobic glycolysis | NA | 55 tumor and adjacent normal tissues | in vitro, in vivo | [147] | |
| MIR4435-2HG | miR-206 | YAP1 | prognosis biomarker and therapeutic target | metastasis | stimulate invasion, migration, EMT, metastasis, and growth | Hippo signaling pathway | 90 tumor and normal tissues | in vitro, in vivo | [149] | |
| NEAT1 | miR-205-5p | VEGFA | diagnosis biomarker and therapeutic target | development | induce proliferation, migration, and invasion | NA | 30 tumor and adjacent normal tissues | in vitro | [150] | |
| miR-34a | SIRT1 | prognosis biomarker and therapeutic target | metastasis | enhance proliferation, invasion, and tumor growth | Wnt signaling pathway | 100 tumor and normal tissues | GEO database, in vitro, in vivo | [151] | ||
| HOTAIR | miR-206 | CCL2 | therapeutic target | progression | stimulate proliferation and invasion, while inhibiting apoptosis | NA | 32 tumor and normal tissues | TCGA, GEPIA, UALCAN, SurvExpress, in vitro | [152] | |
| HCG18 | miR-1271 | MTDH | therapeutic target | development | promote proliferation and invasion | Wnt signaling pathway | 20 tumor and adjacent normal tissues | StarBase, in vitro | [153] | |
| RoR | miR-6833-3p | SMC4 | NA | tumorigenesis | facilitate proliferation and viability, while inhibiting apoptosis | NA | 24 tumor and normal tissues | in vitro | [154] | |
| SNHG8 | miR-588 | ATG7 | therapeutic target | development | induce proliferation and autophagy | NA | NA | TCGA, in vitro | [155] | |
| MAFG-AS1 | miR-149-3p | HOXB8 | prognosis biomarker and therapeutic target | progression | stimulate proliferation, migration, and invasion | NA | 30 tumor and adjacent normal tissues | in vitro, in vivo | [156] | |
| SNHG6 | miR-181b-5p | JAK2 | biomarker and therapeutic target | progression | enhance proliferation, while repressing apoptosis | NA | 40 tumor and adjacent normal tissues | in vitro | [157] | |
| miR-181c-5p | ||||||||||
| RHPN1-AS1 | miR-7-5p | OGT | therapeutic target | progression | promote proliferation, migration, and invasion, while suppressing apoptosis | NA | NA | in vitro, in vivo | [158] | |
| CASC21 | miR-7-5p | YAP1 | therapeutic target | progression | facilitate migration, invasion, and EMT, while inhibiting apoptosis | NA | NA | GEPIA, in vitro | [159] | |
| HOXD-AS1 | miR-526b-3p | CCND1 | NA | progression | stimulate proliferation, migration, and invasion | NA | 54 tumor and adjacent normal tissues | in vitro | [160] | |
| RP11-757G1.5 | miR-139-5p | YAP1 | prognosis biomarker and therapeutic target | progression and metastasis | enhance proliferation, cell cycle progression, migration, invasion, tumor growth, and metastasis | NA | 43 tumor and 6 adjacent normal tissues | in vitro, in vivo | [161] | |
| MCF2L-AS1 | miR-874-3p | CCNE1 | diagnosis and prognosis biomarkers and therapeutic target | initiation and progression | induce proliferation, migration, invasion, and EMT, while inhibiting apoptosis | NA | 130 tumor and normal tissues | in vitro | [162] | |
| DANCR | miR-185-5p | HMGA2 | NA | progression | promote proliferation, migration, and invasion | NA | 50 tumor and adjacent normal tissues | in vitro | [163] | |
| RNCR3 | miR-1301-3p | AKT1 | therapeutic target | progression | facilitate proliferation and invasion, while suppressing apoptosis | NA | 76 tumor and adjacent normal tissues | in vitro, in vivo | [164] | |
| DOWN | MIR503HG | miR-107 | PAR4 | therapeutic target | suppress migration and invasion | NA | 80 tumor and adjacent normal tissues | in vitro, in vivo | [148] | |
| LINC00485 | miR-581 | EDEM1 | therapeutic target | progression and metastasis | repress proliferation, migration, invasion, tumor growth, and metastasis | NA | 52 tumor and adjacent normal tissues | TCGA, GEO database, in vitro, in vivo | [165] | |
| DPP10-AS1 | miR-127-3p | ADCY1 | therapeutic target | progression | inhibit stemness, sphere formation, proliferation, migration, invasion, and tumor growth, while enhancing apoptosis | NA | 54 rumor and adjacent normal tissues | in vitro, in vivo | [166] | |
| MBNL1-AS1 | miR-412-3p | MYL9 | therapeutic target | progression, occurrence | repress proliferation, invasion, migration, and tumor formation, while increasing apoptosis | NA | NA | TCGA, GEO database, in vitro, in vivo | [167] | |
| FENDRR | miR-18a-5p | ING4 | NA | progression and metastasis | suppress proliferation, migration, invasion, tumor growth, and metastasis | NA | 42 tumor and adjacent normal tissues | GEPIA, in vitro, in vivo | [168] | |
| MCM3AP-AS1 | miR-19a-3p | FOXF2 | biomarker and therapeutic target | progression | inhibit proliferation and migration | NA | 53 tumor and adjacent normal tissues | GEPIA, in vitro | [169] |
3.2.2. CircRNA/miRNA/mRNA
| CircRNA Expression | CircRNAs | miRNAs | Genes | Clinical Value | Impact on CRC | Function | Signaling Pathway | Human Samples | Study Model | References |
|---|---|---|---|---|---|---|---|---|---|---|
| UP | CircVAPA | miR-125a | CREB5 | therapeutic target | progression | induce cycle progression, glycolysis, migration, and invasion | NA | 42 tumor and normal tissues | in vitro | [187] |
| CircCTNNA1 | miR-149-5p | FOXM1 | diagnosis biomarker and therapeutic target | progression | facilitate proliferation, DNA synthesis, migration, invasion, and tumorigenesis | NA | 60 tumor and adjacent normal tissues | TCGA, in vitro, in vivo | [190] | |
| CircGLG1 | miR-622 | KRAS | diagnosis biomarker and therapeutic target | progression | promote proliferation, invasion, and migration | NA | 40 tumor and normal tissues | in vitro | [191] | |
| CircSPARC | miR-485-3p | JAK2 | diagnosis and prognosis biomarkers and therapeutic target | progression and metastasis | enhance proliferation, migration, invasion, tumor growth, and metastasis | JAK/STAT signaling pathway | 84 tumor and adjacent normal tissues, 40 plasmas of patients | GEO database, in vitro, in vivo | [193] | |
| Circ0007142 | miR-122-5p | CDC25A | diagnosis biomarker and therapeutic target | progression | stimulate proliferation, migration, invasion, and tumor growth | NA | 31 tumor and adjacent normal tissues | in vitro, in vivo | [194] | |
| miR-455-5p | SGK1 | therapeutic target | progression | induce proliferation, migration, invasion, and tumor growth while repressing apoptosis | NA | 45 tumor and para-carcinoma tissues | in vitro, in vivo | [195] | ||
| Circ0000467 | miR-382-5p | EN2 | therapeutic target | development | enhance proliferation, migration, invasion, and EMT | NA | 69 tumor and adjacent normal tissues | GEO database, in vitro | [196] | |
| Circ0001178 | miR-382 | ZEB1 | therapeutic target | metastasis | stimulate migration, invasion, EMT, and metastasis | NA | 102 tumor tissues | in vitro, in vivo | [197] | |
| miR-587 | ||||||||||
| miR-616 | ||||||||||
| Circ0053277 | miR-2467-3p | MMP14 | therapeutic target | development and progression | promote proliferation, migration, and EMT | NA | 3 tumor and normal tissues | in vitro | [198] | |
| Circ0060745 | miR-4736 | CSE1L | therapeutic target | NA | accelerate proliferation, migration, and invasion | NA | 28 tumor and para-tumor tissues | in vitro | [199] | |
| Circ102209 | miR-761 | RIN1 | therapeutic target | progression | enhance proliferation, cell cycle progression, migration, invasion, EMT, and tumor growth, while suppressing apoptosis | NA | 56 tumor and para-carcinoma tissues | human circRNA array v2, in vitro, in vivo | [200] | |
| Circ100146 | miR-149 | HMGA2 | early diagnosis biomarker and therapeutic target | progression and metastasis | facilitate proliferation, migration, invasion, tumor growth, and metastasis, while inhibiting apoptosis | NA | 58 tumor and normal tissues | GEO database, in vitro, in vivo | [201] | |
| Circ0004277 | miR-512-5p | PTMA | NA | progression | induce proliferation and tumor growth, while inhibiting apoptosis | NA | 50 tumor and para-carcinoma tissues | GEO database, in vitro, in vivo | [202] | |
| CircFARSA | miR-330-5p | LASP1 | therapeutic target | progression | promote proliferation, migration, invasion, and tumor growth | NA | 40 tumor and adjacent normal tissues | in vitro, in vivo | [203] | |
| Circ000166 | miR-326 | diagnosis biomarker and therapeutic target | progression | stimulate proliferation, while repressing apoptosis | NA | 40 tumor and adjacent normal tissues | GEO database, in vitro | [204] | ||
| CircPRMT5 | miR-377 | E2F3 | therapeutic target | progression | facilitate proliferation and tumor growth | NA | 30 tumor and adjacent normal tissues | in vitro, in vivo | [205] | |
| CircERBIN | miR-125a-5p | 4EBP-1 | therapeutic target | progression | enhance proliferation, migration, invasion, tumor growth, angiogenesis, and metastasis | NA | 59 tumor and adjacent normal tissues | in vitro, in vivo | [206] | |
| miR-138-5p | ||||||||||
| CircMAT2B | miR-610 | E2F1 | therapeutic target | progression | induce proliferation | NA | 70 tumor and adjacent normal tissues | in vitro | [207] | |
| Circ0084615 | miR-599 | DNMT3A | biomarker and therapeutic target | progression and metastasis | promote proliferation, migration, invasion, and metastasis | NA | 50 tumor and adjacent normal tissues | GEO database, in vitro, in vivo | [208] | |
| CircDENND4C | miR-760 | GLUT1 | diagnosis biomarker and therapeutic target | progression | enhance proliferation, migration, glycolysis, and tumor growth | NA | tumor and normal tissues | sequencing, in vitro, in vivo | [209] | |
| CircUBAP2 | miR-582-5p | FOXO1 | biomarker and therapeutic target | progression and metastasis | stimulate migration, invasion, proliferation, autophagy, tumor growth, and metastasis | NA | 3 tumor and normal tissues | in vitro, in vivo | [210] | |
| Circ0089153 | miR-198 | SENP1 | therapeutic target | progression | facilitate proliferation, sphere formation, tube formation, and tumor growth, while suppressing apoptosis | NA | 50 tumor and adjacent noncancerous tissues | in vitro, in vivo | [211] | |
| CircNOX4 | miR-485-5p | CKS1B | NA | progression | induce proliferation, migration, invasion, glycolysis, and tumor growth | NA | 46 tumor and adjacent normal tissues | in vitro, in vivo | [212] | |
| CircHERC4 | miR-556-5p | CTBP2 | prognosis biomarker and therapeutic target | progression and metastasis | promote proliferation, migration, invasion, tumor growth, and metastasis | Notch signaling pathway | 120 tumor and adjacent normal tissues | sequencing, in vitro, in vivo | [213] | |
| Circ0000372 | miR-495 | IL6 | prognosis biomarker and therapeutic target | progression | enhance proliferation, migration, invasion, and tumor growth | JAK/STAT signaling pathway | 60 tumor and adjacent normal tissues | in vitro, in vivo | [214] | |
| CircLDLR | miR-30a-3p | SOAT1 | biomarker and therapeutic target | progression and metastasis | stimulate proliferation, DNA synthesis, cholesterol, migration, invasion, tumor growth, and metastasis | NA | 80 tumor and 15 normal tissues | in vitro, in vivo | [215] | |
| DOWN | Circ0008285 | miR-382-5p | PTEN | therapeutic target | progression | repress proliferation and migration | PI3K/AKT signaling pathway | 56 tumor and noncancerous colorectal mucosa tissues | in vitro | [188] |
| Circ_0009361 | miR-582 | APC2 | therapeutic target | progression and metastasis | inhibit proliferation, EMT, migration, invasion, tumor growth, and metastasis | Wnt signaling pathway | 30 tumor and paracancerous tissues | microarray, in vitro, in vivo | [189] | |
| Circ0065378 | miR-4701-5p | TUSC1 | diagnosis biomarker and therapeutic target | progression | suppress proliferation, migration, invasion, EMT, and tumor growth | NA | 25 tumor and adjacent normal tissues | sequencing, in vitro, in vivo | [192] | |
| CircCUL2 | miR-208a-3p | PPP6C | diagnosis biomarker and therapeutic target | progression | repress proliferation, apoptosis, and tumor growth, while increasing apoptosis | NA | 30 tumor and adjacent normal tissues | microarray, in vitro, in vivo | [216] | |
| CircSMARCA5 | miR-93-5p | ARID4B | biomarker | progression | inhibit proliferation, migration, invasion, and tumor growth | NA | tumor and adjacent tissues | in vitro, in vivo | [217] | |
| Circ0003266 | miR-503-5p | PDCD4 | therapeutic target | progression | suppress proliferation, migration, and invasion, while accelerating apoptosis | NA | 46 tumor and paracancerous tissues | GEO database, in vitro | [218] |
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Giaquinto, A.N.; Jemal, A. Cancer statistics, 2024. CA Cancer J. Clin. 2024, 74, 12–49. [Google Scholar] [CrossRef]
- Hong, Y.; Kim, J.; Choi, Y.J.; Kang, J.G. Clinical study of colorectal cancer operation: Survival analysis. Korean J. Clin. Oncol. 2020, 16, 3. [Google Scholar] [CrossRef]
- Vafapour, Z.; Troy, W.; Rashidi, A. Colon cancer detection by designing and analytical evaluation of a water-based THz metamaterial perfect absorber. IEEE Sens. J. 2021, 21, 19307–19313. [Google Scholar] [CrossRef]
- Sawicki, T.; Ruszkowska, M.; Danielewicz, A.; Niedźwiedzka, E.; Arłukowicz, T.; Przybyłowicz, K.E. A review of colorectal cancer in terms of epidemiology, risk factors, development, symptoms and diagnosis. Cancers 2021, 13, 2025. [Google Scholar] [CrossRef]
- Stefani, C.; Miricescu, D.; Stanescu-Spinu, I.-I.; Nica, R.I.; Greabu, M.; Totan, A.R.; Jinga, M. Growth factors, PI3K/AKT/mTOR and MAPK signaling pathways in colorectal cancer pathogenesis: Where are we now? Int. J. Mol. Sci. 2021, 22, 10260. [Google Scholar] [CrossRef]
- Liebl, M.C.; Hofmann, T.G. The role of p53 signaling in colorectal cancer. Cancers 2021, 13, 2125. [Google Scholar] [CrossRef]
- Tomita, N.; Ishida, H.; Tanakaya, K.; Yamaguchi, T.; Kumamoto, K.; Tanaka, T.; Hinoi, T.; Miyakura, Y.; Hasegawa, H.; Takayama, T. Japanese society for cancer of the colon and rectum (JSCCR) guidelines 2020 for the clinical practice of hereditary colorectal cancer. Int. J. Clin. Oncol. 2021, 26, 1353–1419. [Google Scholar] [CrossRef]
- Ahmad, R.; Singh, J.K.; Wunnava, A.; Al-Obeed, O.; Abdulla, M.; Srivastava, S.K. Emerging trends in colorectal cancer: Dysregulated signaling pathways. Int. J. Mol. Med. 2021, 47, 14. [Google Scholar] [CrossRef]
- Wan, M.-l.; Wang, Y.; Zeng, Z.; Deng, B.; Zhu, B.-s.; Cao, T.; Li, Y.-k.; Xiao, J.; Han, Q.; Wu, Q. Colorectal cancer (CRC) as a multifactorial disease and its causal correlations with multiple signaling pathways. Biosci. Rep. 2020, 40, BSR20200265. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Wang, Z.-Z.; Zhao, M.; Zhang, Y.; Chen, N.-H. Role of non-coding RNA in the pathogenesis of depression. Gene 2020, 735, 144276. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Wang, S.; Wu, W.; Shan, P.; Chen, Y.; Meng, J.; Xing, L.; Yun, J.; Hao, L.; Wang, X. Mechanisms of circRNA/lncRNA-miRNA interactions and applications in disease and drug research. Biomed. Pharmacother. 2023, 162, 114672. [Google Scholar] [CrossRef] [PubMed]
- Nepstad, I.; Hatfield, K.J.; Grønningsæter, I.S.; Reikvam, H. The PI3K-Akt-mTOR signaling pathway in human acute myeloid leukemia (AML) cells. Int. J. Mol. Sci. 2020, 21, 2907. [Google Scholar] [CrossRef] [PubMed]
- Malki, A.; ElRuz, R.A.; Gupta, I.; Allouch, A.; Vranic, S.; Al Moustafa, A.-E. Molecular mechanisms of colon cancer progression and metastasis: Recent insights and advancements. Int. J. Mol. Sci. 2020, 22, 130. [Google Scholar] [CrossRef]
- Zhao, H.; Ming, T.; Tang, S.; Ren, S.; Yang, H.; Liu, M.; Tao, Q.; Xu, H. Wnt signaling in colorectal cancer: Pathogenic role and therapeutic target. Mol. Cancer 2022, 21, 144. [Google Scholar] [CrossRef] [PubMed]
- Steinhart, Z.; Angers, S. Wnt signaling in development and tissue homeostasis. Development 2018, 145, dev146589. [Google Scholar] [CrossRef] [PubMed]
- Hayat, R.; Manzoor, M.; Hussain, A. Wnt signaling pathway: A comprehensive review. Cell Biol. Int. 2022, 46, 863–877. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ortiz, M.A.; Kotula, L. The physiological role of Wnt pathway in normal development and cancer. Medicus. Exp. Biol. Med. 2020, 245, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Najafi, S.M.A. Canonical Wnt Signaling (Wnt/β-Catenin Pathway): A Potential Target for Cancer Prevention and Therapy. Iran. Biomed. J. 2020, 24, 269. [Google Scholar] [CrossRef]
- Bian, J.; Dannappel, M.; Wan, C.; Firestein, R. Transcriptional regulation of Wnt/β-catenin pathway in colorectal cancer. Cells 2020, 9, 2125. [Google Scholar] [CrossRef]
- Schatoff, E.M.; Leach, B.I.; Dow, L.E. Wnt signaling and colorectal cancer. Curr. Color. Cancer Rep. 2017, 13, 101–110. [Google Scholar] [CrossRef]
- Fleming-de-Moraes, C.D.; Rocha, M.R.; Tessmann, J.W.; de Araujo, W.M.; Morgado-Diaz, J.A. Crosstalk between PI3K/Akt and Wnt/β-catenin pathways promote colorectal cancer progression regardless of mutational status. Cancer Biol. Ther. 2022, 23, 1–13. [Google Scholar] [CrossRef]
- Basu, S.; Haase, G.; Ben-Ze’ev, A. Wnt signaling in cancer stem cells and colon cancer metastasis. F1000Research 2016, 5, 699. [Google Scholar] [CrossRef] [PubMed]
- Aghabozorgi, A.S.; Ebrahimi, R.; Bahiraee, A.; Tehrani, S.S.; Nabizadeh, F.; Setayesh, L.; Jafarzadeh-Esfehani, R.; Ferns, G.A.; Avan, A.; Rashidi, Z. The genetic factors associated with Wnt signaling pathway in colorectal cancer. Life Sci. 2020, 256, 118006. [Google Scholar] [CrossRef]
- Network, C.G.A. Comprehensive molecular characterization of human colon and rectal cancer. Nature 2012, 487, 330. [Google Scholar] [CrossRef] [PubMed]
- La Vecchia, S.; Sebastián, C. Metabolic pathways regulating colorectal cancer initiation and progression. Semin. Cell Dev. Biol. 2020, 98, 63–70. [Google Scholar] [CrossRef]
- Yoshida, N.; Kinugasa, T.; Ohshima, K.; Yuge, K.; Ohchi, T.; Fujino, S.; Shiraiwa, S.; Katagiri, M.; Akagi, Y. Analysis of Wnt and β-catenin expression in advanced colorectal cancer. Anticancer Res. 2015, 35, 4403–4410. [Google Scholar] [PubMed]
- Kim, W.K.; Kwon, Y.; Jang, M.; Park, M.; Kim, J.; Cho, S.; Jang, D.G.; Lee, W.-B.; Jung, S.H.; Choi, H.J. β-catenin activation down-regulates cell-cell junction-related genes and induces epithelial-to-mesenchymal transition in colorectal cancers. Sci. Rep. 2019, 9, 18440. [Google Scholar] [CrossRef]
- Zhang, Z.; Chang, Y.; Zhang, J.; Lu, Y.; Zheng, L.; Hu, Y.; Zhang, F.; Li, X.; Zhang, W.; Li, X. HMGB3 promotes growth and migration in colorectal cancer by regulating WNT/β-catenin pathway. PLoS ONE 2017, 12, e0179741. [Google Scholar] [CrossRef]
- Wachsmannova, L.; Mego, M.; Stevurkova, V.; Zajac, V.; Ciernikova, S. Novel strategies for comprehensive mutation screening of the APC gene. Neoplasma 2017, 64, 338–343. [Google Scholar] [CrossRef]
- Meng, F.; Liu, X.; Lin, C.; Xu, L.; Liu, J.; Zhang, P.; Zhang, X.; Song, J.; Yan, Y.; Ren, Z. SMYD2 suppresses APC2 expression to activate the Wnt/β-catenin pathway and promotes epithelial-mesenchymal transition in colorectal cancer. Am. J. Cancer Res. 2020, 10, 997. [Google Scholar]
- Miricescu, D.; Totan, A.; Stanescu-Spinu, I.-I.; Badoiu, S.C.; Stefani, C.; Greabu, M. PI3K/AKT/mTOR signaling pathway in breast cancer: From molecular landscape to clinical aspects. Int. J. Mol. Sci. 2020, 22, 173. [Google Scholar] [CrossRef] [PubMed]
- Ruchi Sharma, V.; Kumar Gupta, G.; Sharma, A.K.; Batra, N.; Sharma, D.K.; Joshi, A.; Sharma, A.K. PI3K/Akt/mTOR intracellular pathway and breast cancer: Factors, mechanism and regulation. Curr. Pharm. Des. 2017, 23, 1633–1638. [Google Scholar] [CrossRef] [PubMed]
- Owusu-Brackett, N.; Shariati, M.; Meric-Bernstam, F. Role of PI3K/AKT/mTOR in Cancer Signaling. In Predictive Biomarkers in Oncology: Applications in Precision Medicine; Springer: Berlin/Heidelberg, Germany, 2019; pp. 263–270. [Google Scholar]
- Wang, Q.; Zhang, P.; Zhang, W.; Zhang, X.; Chen, J.; Ding, P.; Li, L.; Lv, X.; Li, L.; Hu, W. PI3K activation is enhanced by FOXM1D binding to p110 and p85 subunits. Signal Transduct. Target. Ther. 2020, 5, 105. [Google Scholar] [CrossRef] [PubMed]
- Mazloumi Gavgani, F.; Smith Arnesen, V.; Jacobsen, R.G.; Krakstad, C.; Hoivik, E.A.; Lewis, A.E. Class I phosphoinositide 3-Kinase PIK3CA/p110α and PIK3CB/p110β isoforms in endometrial cancer. Int. J. Mol. Sci. 2018, 19, 3931. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Li, G.; Lu, J.; Li, L. Crosstalk between circRNAs and the PI3K/AKT signaling pathway in cancer progression. Signal Transduct. Target. Ther. 2021, 6, 400. [Google Scholar] [CrossRef] [PubMed]
- Papadatos-Pastos, D.; Rabbie, R.; Ross, P.; Sarker, D. The role of the PI3K pathway in colorectal cancer. Crit. Rev. Oncol. Hematol. 2015, 94, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Moafian, Z.; Maghrouni, A.; Soltani, A.; Hashemy, S.I. Cross-talk between non-coding RNAs and PI3K/AKT/mTOR pathway in colorectal cancer. Mol. Biol. Rep. 2021, 48, 4797–4811. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.M.; Gulhati, P.; Rampy, B.A.; Han, Y.; Rychahou, P.G.; Doan, H.Q.; Weiss, H.L.; Evers, B.M. Novel expression patterns of PI3K/Akt/mTOR signaling pathway components in colorectal cancer. J. Am. Coll. Surg. 2010, 210, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.-F.; Yu, B.-H.; Li, D.-L.; Ke, H.-L.; Guo, X.-Z.; Xiao, X.-Y. PI3K expression and PIK3CA mutations are related to colorectal cancer metastases. World J. Gastroenterol. 2012, 18, 3745. [Google Scholar] [CrossRef]
- Suman, S.; Kurisetty, V.; Das, T.P.; Vadodkar, A.; Ramos, G.; Lakshmanaswamy, R.; Damodaran, C. Activation of AKT signaling promotes epithelial–mesenchymal transition and tumor growth in colorectal cancer cells. Mol. Carcinog. 2014, 53, E151–E160. [Google Scholar] [CrossRef]
- Cai, Z.; Ke, J.; He, X.; Yuan, R.; Chen, Y.; Wu, X.; Wang, L.; Wang, J.; Lan, P.; Wu, X. Significance of mTOR signaling and its inhibitor against cancer stem-like cells in colorectal cancer. Ann. Surg. Oncol. 2014, 21, 179–188. [Google Scholar] [CrossRef]
- Xu, W.; Yu, M.; Qin, J.; Luo, Y.; Zhong, M. LACTB regulates PIK3R3 to promote autophagy and inhibit EMT and proliferation through the PI3K/AKT/mTOR signaling pathway in colorectal cancer. Cancer Manag. Res. 2020, 12, 5181–5200. [Google Scholar] [CrossRef]
- Feng, Y.; Ji, D.; Huang, Y.; Ji, B.; Zhang, Y.; Li, J.; Peng, W.; Zhang, C.; Zhang, D.; Sun, Y. TGM3 functions as a tumor suppressor by repressing epithelial-to-mesenchymal transition and the PI3K/AKT signaling pathway in colorectal cancer. Oncol. Rep. 2020, 43, 864–876. [Google Scholar] [CrossRef]
- Su, W.; Feng, B.; Hu, L.; Guo, X.; Yu, M. MUC3A promotes the progression of colorectal cancer through the PI3K/Akt/mTOR pathway. BMC Cancer 2022, 22, 602. [Google Scholar] [CrossRef]
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.-N.; Luo, Y.-H.; Liu, S.-B.; Xu, W.-T.; Zhang, Y.; Zhang, T.; Xue, H.; Zuo, W.-B.; Li, Y.-N.; Wang, C.-Y. Zeaxanthin induces apoptosis via ROS-regulated MAPK and AKT signaling pathway in human gastric cancer cells. Onco Targets Ther. 2020, 13, 10995–11006. [Google Scholar] [CrossRef]
- Asl, E.R.; Amini, M.; Najafi, S.; Mansoori, B.; Mokhtarzadeh, A.; Mohammadi, A.; Lotfinejad, P.; Bagheri, M.; Shirjang, S.; Lotfi, Z. Interplay between MAPK/ERK signaling pathway and MicroRNAs: A crucial mechanism regulating cancer cell metabolism and tumor progression. Life Sci. 2021, 278, 119499. [Google Scholar] [CrossRef]
- Kciuk, M.; Gielecińska, A.; Budzinska, A.; Mojzych, M.; Kontek, R. Metastasis and MAPK pathways. Int. J. Mol. Sci. 2022, 23, 3847. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.Y.; Richardson, B.C. The MAPK signalling pathways and colorectal cancer. Lancet Oncol. 2005, 6, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Yang, J.; Song, P. Correlation of ERK/MAPK signaling pathway with proliferation and apoptosis of colon cancer cells. Oncol. Lett. 2019, 17, 2266–2270. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xie, G.; Li, M.; Du, J.; Wang, M. COPB2 gene silencing inhibits colorectal cancer cell proliferation and induces apoptosis via the JNK/c-Jun signaling pathway. PLoS ONE 2020, 15, e0240106. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liao, Y.; Chen, C.; Sun, W.; Sun, X.; Liu, Y.; Xu, E.; Lai, M.; Zhang, H. p38-regulated FOXC1 stability is required for colorectal cancer metastasis. J. Pathol. Inform. 2020, 250, 217–230. [Google Scholar] [CrossRef]
- Ou, S.; Liao, Y.; Shi, J.; Tang, J.; Ye, Y.; Wu, F.; Wang, W.; Fei, J.; Xie, F.; Bai, L. S100A16 suppresses the proliferation, migration and invasion of colorectal cancer cells in part via the JNK/p38 MAPK pathway. Mol. Med. Rep. 2021, 23, 164. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, X.; Peng, L.; Deng, Q.; Liang, Y.; Qing, H.; Jiang, B. CD24-dependent MAPK pathway activation is required for colorectal cancer cell proliferation. Cancer Sci. 2010, 101, 112–119. [Google Scholar] [CrossRef]
- Itatani, Y.; Kawada, K.; Sakai, Y. Transforming growth factor-β signaling pathway in colorectal cancer and its tumor microenvironment. Int. J. Mol. Sci. 2019, 20, 5822. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Wei, J.; Sun, J. Roles of TGF-β signaling pathway in tumor microenvirionment and cancer therapy. Int. Immunopharmacol. 2020, 89, 107101. [Google Scholar] [CrossRef]
- Pawlak, J.B.; Blobe, G.C. TGF-β superfamily co-receptors in cancer. Dev. Dynam. 2022, 251, 117–143. [Google Scholar] [CrossRef]
- Tzavlaki, K.; Moustakas, A. TGF-β Signaling. Biomolecules 2020, 10, 487. [Google Scholar] [CrossRef]
- Yu, X.-Y.; Sun, Q.; Zhang, Y.-M.; Zou, L.; Zhao, Y.-Y. TGF-β/Smad signaling pathway in tubulointerstitial fibrosis. Front. Pharmacol. 2022, 13, 860588. [Google Scholar] [CrossRef]
- Yang, D.; Hou, T.; Li, L.; Chu, Y.; Zhou, F.; Xu, Y.; Hou, X.; Song, H.; Zhu, K.; Hou, Z. Smad1 promotes colorectal cancer cell migration through Ajuba transactivation. Oncotarget 2017, 8, 110415. [Google Scholar] [CrossRef]
- Shi, W.; He, J.; Huang, Y.; Zeng, Z.; Feng, Z.; Xu, H.; Nie, Y. Integrin β5 enhances the malignancy of human colorectal cancer by increasing the TGF-β signaling. Anti-Cancer Drugs 2021, 32, 717–726. [Google Scholar] [CrossRef]
- Wang, L.; Ai, M.; Nie, M.; Zhao, L.; Deng, G.; Hu, S.; Han, Y.; Zeng, W.; Wang, Y.; Yang, M. EHF promotes colorectal carcinoma progression by activating TGF-β1 transcription and canonical TGF-β signaling. Cancer Sci. 2020, 111, 2310–2324. [Google Scholar] [CrossRef]
- Williams, A.B.; Schumacher, B. p53 in the DNA-damage-repair process. Cold Spring Harb. Perspect. Med. 2016, 6, a026070. [Google Scholar] [CrossRef]
- Borrero, L.J.H.; El-Deiry, W.S. Tumor suppressor p53: Biology, signaling pathways, and therapeutic targeting. Biochim. Biophys. Acta 2021, 1876, 188556. [Google Scholar]
- Fu, X.; Wu, S.; Li, B.; Xu, Y.; Liu, J. Functions of p53 in pluripotent stem cells. Protein Cell 2020, 11, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Wei, W.; Sun, Y. PROTAC degraders with ligands recruiting MDM2 E3 ubiquitin ligase: An updated perspective. Acta Mater. Med. 2022, 1, 244. [Google Scholar] [CrossRef] [PubMed]
- Eskandari, M.; Shi, Y.; Liu, J.; Albanese, J.; Goel, S.; Verma, A.; Wang, Y. The expression of MDM2, MDM4, p53 and p21 in myeloid neoplasms and the effect of MDM2/MDM4 dual inhibitor. Leuk Lymphoma 2021, 62, 167–175. [Google Scholar] [CrossRef]
- Wang, P.; Wang, H.-Y.; Gao, X.-J.; Zhu, H.-X.; Zhang, X.-P.; Liu, F.; Wang, W. Encoding and decoding of p53 dynamics in cellular response to stresses. Cells 2023, 12, 490. [Google Scholar] [CrossRef]
- Solomon, H.; Dinowitz, N.; Pateras, I.S.; Cooks, T.; Shetzer, Y.; Molchadsky, A.; Charni, M.; Rabani, S.; Koifman, G.; Tarcic, O. Mutant p53 gain of function underlies high expression levels of colorectal cancer stem cells markers. Oncogene 2018, 37, 1669–1684. [Google Scholar] [CrossRef]
- Nagao, K.; Koshino, A.; Sugimura-Nagata, A.; Nagano, A.; Komura, M.; Ueki, A.; Ebi, M.; Ogasawara, N.; Tsuzuki, T.; Kasai, K. The complete loss of p53 expression uniquely predicts worse prognosis in colorectal cancer. Int. J. Mol. Sci. 2022, 23, 3252. [Google Scholar] [CrossRef]
- Kruschewski, M.; Mueller, K.; Lipka, S.; Budczies, J.; Noske, A.; Buhr, H.J.; Elezkurtaj, S. The prognostic impact of p53 expression on sporadic colorectal cancer is dependent on p21 status. Cancers 2011, 3, 1274–1284. [Google Scholar] [CrossRef] [PubMed]
- Kondo, I.; Iida, S.; Takagi, Y.; Sugihara, K. MDM2 mRNA expression in the p53 pathway may predict the potential of invasion and liver metastasis in colorectal cancer. Dis. Colon Rectum 2008, 51, 1395–1402. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wei, J.; Zhang, H.; Zheng, X.; Zhou, H.; Luo, Y.; Yang, J.; Deng, Q.; Huang, S.; Fu, Z. PRDX2 promotes the proliferation of colorectal cancer cells by increasing the ubiquitinated degradation of p53. Cell Death Dis. 2021, 12, 605. [Google Scholar] [CrossRef] [PubMed]
- Shen, A.; Liu, L.; Huang, Y.; Shen, Z.; Wu, M.; Chen, X.; Wu, X.; Lin, X.; Chen, Y.; Li, L. Down-regulating HAUS6 suppresses cell proliferation by activating the p53/p21 pathway in colorectal cancer. Front. Cell Dev. Biol. 2022, 9, 772077. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xiao, X.; Li, D.; Chi, Y.; Wei, P.; Wang, Y.; Ni, S.; Tan, C.; Zhou, X.; Du, X. Abnormal expression of GADD45B in human colorectal carcinoma. J. Transl. Med. 2012, 10, 215. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wei, Y.; Feng, Q.; Ren, L.; He, G.; Chang, W.; Zhu, D.; Yi, T.; Lin, Q.; Tang, W. Ribosomal protein S15A promotes malignant transformation and predicts poor outcome in colorectal cancer through misregulation of p53 signaling pathway. Int. J. Oncol. 2016, 48, 1628–1638. [Google Scholar] [CrossRef] [PubMed]
- Science in School. Available online: https://www.scienceinschool.org/article/2023/the-importance-of-non-coding-rnas/ (accessed on 1 February 2024).
- Moraes, F.; Góes, A. A decade of human genome project conclusion: Scientific diffusion about our genome knowledge. Biochem. Mol. Biol. Educ. 2016, 44, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Oyelade, J.; Isewon, I.; Ogunlana, O.; Aworunse, O.; Oyesola, O.; Aromolaran, O.; Dokumu, T.; Ademuwagun, I.; Iheagwam, F.; Babatunde, E. Overview of the human genome. In Genome Plasticity in Health and Disease; Academic Press: Cambridge, MA, USA, 2020; pp. 9–26. [Google Scholar]
- Santosh, B.; Varshney, A.; Yadava, P.K. Non-coding RNAs: Biological functions and applications. Cell Biochem. Funct. 2015, 33, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Micheel, J.; Safrastyan, A.; Wollny, D. Advances in non-coding RNA sequencing. Non-Coding RNA 2021, 7, 70. [Google Scholar] [CrossRef]
- Bhatti, G.K.; Khullar, N.; Sidhu, I.S.; Navik, U.S.; Reddy, A.P.; Reddy, P.H.; Bhatti, J.S. Emerging role of non-coding RNA in health and disease. Metab. Brain Dis. 2021, 36, 1119–1134. [Google Scholar] [CrossRef]
- Luo, R.; Song, J.; Zhang, W.; Ran, L. Identification of MFI2-AS1, a novel pivotal lncRNA for prognosis of stage III/IV colorectal cancer. Dig. Dis. Sci. 2020, 65, 3538–3550. [Google Scholar] [CrossRef] [PubMed]
- Shao, T.; Pan, Y.-h.; Xiong, X.-d. Circular RNA: An important player with multiple facets to regulate its parental gene expression. Mol. Ther. Nucleic Acids 2021, 23, 369–376. [Google Scholar] [CrossRef]
- Gebremedhn, S.; Ali, A.; Hossain, M.; Hoelker, M.; Salilew-Wondim, D.; Anthony, R.V.; Tesfaye, D. MicroRNA-mediated gene regulatory mechanisms in mammalian female reproductive health. Int. J. Mol. Sci. 2021, 22, 938. [Google Scholar] [CrossRef]
- Wang, L.-L.; Chen, W.-Q.; Wang, Y.-R.; Zeng, L.-P.; Chen, T.-T.; Chen, G.-Y.; Chen, J.-H. Numerous long single-stranded DNAs produced by dual amplification reactions for electrochemical detection of exosomal microRNAs. Biosens. Bioelectron. 2020, 169, 112555. [Google Scholar] [CrossRef] [PubMed]
- Wallace, D.R.; Taalab, Y.M.; Heinze, S.; Tariba Lovaković, B.; Pizent, A.; Renieri, E.; Tsatsakis, A.; Farooqi, A.A.; Javorac, D.; Andjelkovic, M. Toxic-metal-induced alteration in miRNA expression profile as a proposed mechanism for disease development. Cells 2020, 9, 901. [Google Scholar] [CrossRef] [PubMed]
- Soni, D.K.; Biswas, R. Role of non-coding RNAs in post-transcriptional regulation of lung diseases. Front. Genet. 2021, 12, 767348. [Google Scholar] [CrossRef]
- Riolo, G.; Cantara, S.; Marzocchi, C.; Ricci, C. miRNA targets: From prediction tools to experimental validation. Methods Protoc. 2020, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Indrieri, A.; Carrella, S.; Carotenuto, P.; Banfi, S.; Franco, B. The pervasive role of the miR-181 family in development, neurodegeneration, and cancer. Int. J. Mol. Sci. 2020, 21, 2092. [Google Scholar] [CrossRef]
- Kalayinia, S.; Arjmand, F.; Maleki, M.; Malakootian, M.; Singh, C.P. MicroRNAs: Roles in cardiovascular development and disease. Cardiovasc. Pathol. 2021, 50, 107296. [Google Scholar] [CrossRef]
- Prasad, A.; Sharma, N.; Prasad, M. Noncoding but coding: Pri-miRNA into the action. Trends Plant Sci. 2021, 26, 204–206. [Google Scholar] [CrossRef]
- Jin, W.; Wang, J.; Liu, C.-P.; Wang, H.-W.; Xu, R.-M. Structural basis for pri-miRNA recognition by Drosha. Mol. Cell 2020, 78, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.; Tran, N. miRNA interplay: Mechanisms and consequences in cancer. Dis. Model. Mech. 2021, 14, dmm047662. [Google Scholar] [CrossRef] [PubMed]
- Klicka, K.; Grzywa, T.M.; Mielniczuk, A.; Klinke, A.; Włodarski, P.K. The role of miR-200 family in the regulation of hallmarks of cancer. Front. Oncol. 2022, 12, 965231. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Bravo Vázquez, L.A.; Pérez Uribe, S.; Roxana Reyes-Pérez, P.; Sharma, A. Current status of microRNA-based therapeutic approaches in neurodegenerative disorders. Cells 2020, 9, 1698. [Google Scholar] [CrossRef] [PubMed]
- Çakmak, H.A.; Demir, M. MicroRNA and cardiovascular diseases. Balkan Med. J. 2020, 37, 60. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.; Wu, W.; Ren, F.; Li, L.; Li, Y.; Li, W.; Wang, A.; Liu, D.; Dong, Y. MiR-346-5p promotes colorectal cancer cell proliferation in vitro and in vivo by targeting FBXL2 and activating the β-catenin signaling pathway. Life Sci. 2020, 244, 117300. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Wang, J.; Cheng, G.; Huang, H. miR-590-5p may regulate colorectal cancer cell viability and migration by targeting PDCD4. Exp. Ther. Med. 2020, 20, 55. [Google Scholar] [CrossRef]
- He, Z.; Dang, J.; Song, A.; Cui, X.; Ma, Z.; Zhang, Y. The involvement of miR-150/β-catenin axis in colorectal cancer progression. Biomed. Pharmacother. 2020, 121, 109495. [Google Scholar] [CrossRef] [PubMed]
- Song, G.-L.; Xiao, M.; Wan, X.-Y.; Deng, J.; Ling, J.-D.; Tian, Y.-G.; Li, M.; Yin, J.; Zheng, R.-Y.; Tang, Y. MiR-130a-3p suppresses colorectal cancer growth by targeting Wnt Family Member 1 (WNT1). Bioengineered 2021, 12, 8407–8418. [Google Scholar] [CrossRef]
- Gao, Z.; Jiang, J.; Hou, L.; Zhang, B. Dysregulation of MiR-144-5p/RNF187 axis contributes to the progression of colorectal cancer. J. Transl. Med. 2022, 10, 65–75. [Google Scholar] [CrossRef]
- Du, G.; Yu, X.; Chen, Y.; Cai, W. MiR-1-3p Suppresses Colorectal Cancer Cell Proliferation and Metastasis by Inhibiting YWHAZ-Mediated Epithelial–Mesenchymal Transition. Front. Oncol. 2021, 11, 634596. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Peng, R.; Zeng, M.; Zhang, Z.; Liu, S.; Jiang, D.; Lu, Y.; Zou, F. An autoregulatory feedback loop of miR-21/VMP1 is responsible for the abnormal expression of miR-21 in colorectal cancer cells. Cell Death Dis. 2020, 11, 1067. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Chen, X.; Ge, S.; Wang, Q.; Liu, Y.; Chen, H.; Xu, J.; Wu, J. MiR-21-5p induces pyroptosis in colorectal cancer via TGFBI. Front. Oncol. 2021, 10, 610545. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Han, L.; Gu, C.; Lai, Y.; Lai, Q.; Li, Q.; He, C.; Meng, Y.; Pan, L.; Liu, S. MiR-452-5p promotes colorectal cancer progression by regulating an ERK/MAPK positive feedback loop. Aging 2021, 13, 7608. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.; Chen, Q.; Lu, Y.; Wei, F.; Chen, C.; Tang, G.; Huang, H. MiR-106b-5p regulates the migration and invasion of colorectal cancer cells by targeting FAT4. Biosci. Rep. 2020, 40, BSR20200098. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yan, B.; Zhang, P.; Liu, S.; Li, Q.; Yang, J.; Yang, F.; Chen, E. MiR-496 promotes migration and epithelial-mesenchymal transition by targeting RASSF6 in colorectal cancer. J. Cell. Physiol. 2020, 235, 1469–1479. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, T.; Han, Y.-N.; Ge, M.; Wang, P.; Sun, L.; Liu, H.; Cao, T.; Nie, Y.; Fan, D. miR-125b promotes colorectal cancer migration and invasion by dual-targeting CFTR and CGN. Cancers 2021, 13, 5710. [Google Scholar] [CrossRef]
- Arabsorkhi, Z.; Gharib, E.; Yaghmoorian Khojini, J.; Farhadieh, M.E.; Nazemalhosseini-Mojarad, E.; Zali, M.R. miR-298 plays a pivotal role in colon cancer invasiveness by targeting PTEN. J. Cell. Physiol. 2020, 235, 4335–4350. [Google Scholar] [CrossRef]
- Chen, X.; Wang, A.-l.; Liu, Y.-y.; Zhao, C.-x.; Zhou, X.; Liu, H.-l.; Lin, M.-b. MiR-429 involves in the pathogenesis of colorectal cancer via directly targeting LATS2. Oxid. Med. Cell. Longev. 2020, 2020, 5316276. [Google Scholar] [CrossRef]
- Li, S.; Hou, X.; Wu, C.; Han, L.; Li, Q.; Wang, J.; Luo, S. MiR-645 promotes invasiveness, metastasis and tumor growth in colorectal cancer by targeting EFNA5. Biomed. Pharmacother. 2020, 125, 109889. [Google Scholar] [CrossRef]
- Sun, N.; Zhang, L.; Zhang, C.; Yuan, Y. miR-144-3p inhibits cell proliferation of colorectal cancer cells by targeting BCL6 via inhibition of Wnt/β-catenin signaling. Cell. Mol. Biol. Lett. 2020, 25, 19. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chen, S.; Zhuo, L.; Zhu, Y.; Zheng, H. Regulation of cancer stem cell properties, angiogenesis, and vasculogenic mimicry by miR-450a-5p/SOX2 axis in colorectal cancer. Cell Death Dis. 2020, 11, 173. [Google Scholar] [CrossRef] [PubMed]
- Machackova, T.; Vychytilova-Faltejskova, P.; Souckova, K.; Trachtova, K.; Brchnelova, D.; Svoboda, M.; Kiss, I.; Prochazka, V.; Kala, Z.; Slaby, O. MiR-215-5p reduces liver metastasis in an experimental model of colorectal cancer through regulation of ECM-receptor interactions and focal adhesion. Cancers 2020, 12, 3518. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Cao, X.; Lai, S.; Luo, X.; Feng, Y.; Wu, J.; Ning, Q.; Xia, X.; Wang, J.; Gong, J. Altered p53 regulation of miR-148b and p55PIK contributes to tumor progression in colorectal cancer. Oncogene 2015, 34, 912–921. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Wang, X.; Li, Z.; Li, B.; Ma, F.; Peng, L.; Zhang, Y.; Xu, A.; Jiang, B. microRNA-16 represses colorectal cancer cell growth in vitro by regulating the p53/survivin signaling pathway. Oncol. Rep. 2013, 29, 1652–1658. [Google Scholar] [CrossRef] [PubMed]
- Yasudome, R.; Seki, N.; Asai, S.; Goto, Y.; Kita, Y.; Hozaka, Y.; Wada, M.; Tanabe, K.; Idichi, T.; Mori, S. Molecular pathogenesis of colorectal cancer: Impact of oncogenic targets regulated by tumor suppressive miR-139-3p. Int. J. Mol. Sci. 2022, 23, 11616. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yang, J.; Wang, H.; Qiao, W.; Guo, Y.; Zhang, S.; Guo, Y. FNDC3B, targeted by miR-125a-5p and miR-217, promotes the proliferation and invasion of colorectal cancer cells via PI3K/mTOR signaling. Onco Targets Ther. 2020, 13, 3501–3510. [Google Scholar] [CrossRef]
- Zhang, H.; Shen, Z.; Zhou, Y.; Zhang, Z.; Wang, Q.; Zhang, M.; Wang, S.; Wang, B. Downregulation of miR-654-3p in colorectal cancer indicates poor prognosis and promotes cell proliferation and invasion by targeting SRC. Front. Genet. 2020, 11, 577948. [Google Scholar] [CrossRef]
- Yang, Y.; Lin, Z.; Han, Z.; Wu, Z.; Hua, J.; Zhong, R.; Zhao, R.; Ran, H.; Qu, K.; Huang, H. miR-539 activates the SAPK/JNK signaling pathway to promote ferropotosis in colorectal cancer by directly targeting TIPE. Cell Death Discov. 2021, 7, 272. [Google Scholar] [CrossRef]
- Chang, S.; Sun, G.; Zhang, D.; Li, Q.; Qian, H. MiR-3622a-3p acts as a tumor suppressor in colorectal cancer by reducing stemness features and EMT through targeting spalt-like transcription factor 4. Cell Death Dis. 2020, 11, 592. [Google Scholar] [CrossRef]
- Liu, H.-T.; Ma, R.-R.; Lv, B.-B.; Zhang, H.; Shi, D.-B.; Guo, X.-Y.; Zhang, G.-H.; Gao, P. LncRNA-HNF1A-AS1 functions as a competing endogenous RNA to activate PI3K/AKT signalling pathway by sponging miR-30b-3p in gastric cancer. Br. J. Cancer 2020, 122, 1825–1836. [Google Scholar] [CrossRef]
- Ala, U. Competing endogenous RNAs, non-coding RNAs and diseases: An intertwined story. Cells 2020, 9, 1574. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Xu, W.-Y.; Xiao, Y.; Jin, B.; Du, S.-D.; Mao, Y.-l.; Zhang, Z.-T. The emerging roles of non-coding competing endogenous RNA in hepatocellular carcinoma. Cancer Cell Int. 2020, 20, 496. [Google Scholar] [CrossRef] [PubMed]
- Mitra, A.; Pfeifer, K.; Park, K.-S. Circular RNAs and competing endogenous RNA (ceRNA) networks. Transl. Cancer Res. 2018, 7 (Suppl. 5), S624. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, J.; Li, J. Role of miRNA sponges in hepatocellular carcinoma. Clin. Chim. Acta. 2020, 500, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Yao, T.; Zhang, Y.; Li, W.; Wang, Z. NEAT1 as a competing endogenous RNA in tumorigenesis of various cancers: Role, mechanism and therapeutic potential. Int. J. Biol. Sci. 2021, 17, 3428. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Lin, Y.; Liu, X.; Chen, J.; Shen, B. Biomarker discovery for the carcinogenic heterogeneity between colon and rectal cancers based on lncRNA-associated ceRNA network analysis. Front. Oncol. 2020, 10, 535985. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Wang, L.; Gong, C.; Wu, W.; Yao, C.; Zhu, S. Long non-coding RNAs: How to regulate the metastasis of non–small-cell lung cancer. J. Cell. Mol. Med. 2020, 24, 3282–3291. [Google Scholar] [CrossRef]
- Yang, X.; Xie, Z.; Lei, X.; Gan, R. Long non-coding RNA GAS5 in human cancer. Oncol. Lett. 2020, 20, 2587–2594. [Google Scholar] [CrossRef]
- Pisignano, G.; Ladomery, M. Post-transcriptional regulation through long non-coding rnas (lncrnas). Non-Coding RNA 2021, 7, 29. [Google Scholar] [CrossRef]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.-L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A. Long non-coding RNAs: Definitions, functions, challenges and recommendations. Nat. Rev. Mol. Cell Biol. 2023, 24, 430–447. [Google Scholar] [CrossRef] [PubMed]
- Neve, B.; Jonckheere, N.; Vincent, A.; Van Seuningen, I. Long non-coding RNAs: The tentacles of chromatin remodeler complexes. Cell. Mol. Life Sci. 2021, 78, 1139–1161. [Google Scholar] [CrossRef] [PubMed]
- Gil, N.; Ulitsky, I. Regulation of gene expression by cis-acting long non-coding RNAs. Nat. Rev. Genet 2020, 21, 102–117. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ding, W.; Yu, W.; Zhang, Y.; Ao, X.; Wang, J. Long non-coding RNAs: Biogenesis, functions, and clinical significance in gastric cancer. Mol. Ther.-Oncolytics 2021, 23, 458–476. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Liu, J.-B.; Deng, J.; Zou, D.-Z.; Wu, J.-J.; Cao, Y.-H.; Yin, J.; Ma, Y.-S.; Da, F.; Li, W. The role of ceRNA-mediated diagnosis and therapy in hepatocellular carcinoma. Hereditas 2021, 158, 44. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet 2016, 17, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Tsagakis, I.; Douka, K.; Birds, I.; Aspden, J.L. Long non-coding RNAs in development and disease: Conservation to mechanisms. J. Pathol. Inform. 2020, 250, 480–495. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Liu, C.; Li, H.; Zhang, L.; Luo, G.; Liang, S.; Lü, M. Research progress on the interactions between long non-coding RNAs and microRNAs in human cancer. Oncol. Lett. 2020, 19, 595–605. [Google Scholar] [CrossRef] [PubMed]
- Atef, M.; Amer, A.; Hafez, Y.; Elsebaey, M.; Saber, S.; Abd El-Khalik, S. Long non-coding RNA EGFR-AS1 in colorectal cancer: Potential role in tumorigenesis and survival via miRNA-133b sponge and EGFR/STAT3 axis regulation. J. Biomed. Sci. 2021, 78, 122–129. [Google Scholar] [CrossRef]
- Shang, A.; Wang, W.; Gu, C.; Chen, W.; Lu, W.; Sun, Z.; Li, D. Long non-coding RNA CCAT1 promotes colorectal cancer progression by regulating miR-181a-5p expression. Aging 2020, 12, 8301. [Google Scholar] [CrossRef]
- Bi, C.; Cui, H.; Fan, H.; Li, L. LncRNA LINC01116 promotes the development of colorectal cancer by targeting miR-9-5p/STMN1. Onco Targets Ther. 2020, 13, 10547–10558. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Yue, W.; Li, M.; Jiang, Z.; Hou, Z.; Liu, W.; Ma, N.; Gan, W.; Li, Y.; Zhou, T. Downregulating long Non-coding RNAs CTBP1-AS2 inhibits colorectal cancer development by modulating the miR-93-5p/TGF-β/SMAD2/3 pathway. Front. Oncol. 2021, 11, 626620. [Google Scholar] [CrossRef] [PubMed]
- Ke, D.; Wang, Q.; Ke, S.; Zou, L.; Wang, Q. Long-non coding RNA SNHG16 supports colon cancer cell growth by modulating miR-302a-3p/AKT axis. Pathol. Oncol. Res. 2020, 26, 1605–1613. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Wang, Y.; Deng, J.; Liu, D.; Zhang, L.; Shao, H.; Wang, Z.; Zhu, W.; Zhao, C.; Ke, Q. Long non-coding RNA COL4A2-AS1 facilitates cell proliferation and glycolysis of colorectal cancer cells via miR-20b-5p/hypoxia inducible factor 1 alpha subunit axis. Bioengineered 2021, 12, 6251–6263. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Li, H.; Zhou, J. Long non-coding RNA MIR503HG inhibits the proliferation, migration and invasion of colon cancer cells via miR-107/Par4 axis. Exp. Cell Res. 2020, 395, 112205. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Yang, Z.; Yang, H.; Li, D.; Qiu, X. Long non-coding RNA MIR4435-2HG promotes colorectal cancer proliferation and metastasis through miR-206/YAP1 axis. Front. Oncol. 2020, 10, 160. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, A.; Sun, Z.; Zhang, J.; Xu, H. Long non-coding RNA NEAT1 promotes colorectal cancer progression by regulating miR-205-5p/VEGFA axis. Hum. Cell 2020, 33, 386–396. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Chen, J.-J.; Lv, Q.; Qin, J.; Huang, Y.-Z.; Yu, M.-H.; Zhong, M. Long non-coding RNA NEAT1 promotes colorectal cancer progression by competitively binding miR-34a with SIRT1 and enhancing the Wnt/β-catenin signaling pathway. Cancer Lett. 2019, 440, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Shengnan, J.; Dafei, X.; Hua, J.; Sunfu, F.; Xiaowei, W.; Liang, X. Long non-coding RNA HOTAIR as a competitive endogenous RNA to sponge miR-206 to promote colorectal cancer progression by activating CCL2. J. Cancer 2020, 11, 4431. [Google Scholar] [CrossRef]
- Li, S.; Wu, T.; Zhang, D.; Sun, X.; Zhang, X. The long non-coding RNA HCG18 promotes the growth and invasion of colorectal cancer cells through sponging miR-1271 and upregulating MTDH/Wnt/β-catenin. Clin. Exp. Pharmacol. Physiol. 2020, 47, 703–712. [Google Scholar] [CrossRef]
- Li, X.; Chen, W.; Jia, J.; You, Z.; Hu, C.; Zhuang, Y.; Lin, Z.; Liu, Y.; Yang, C.; Xu, R. The long non-coding RNA-RoR promotes the tumorigenesis of human colorectal cancer by targeting miR-6833-3p through SMC4. Onco Targets Ther. 2020, 13, 2573–2581. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Fu, Y.; Chen, Y.; Li, X. Long non-coding RNA SNHG8 promotes autophagy as a ceRNA to upregulate ATG7 by sponging microRNA-588 in colorectal cancer. Oncol. Lett. 2021, 22, 577. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Z.; Deng, H.; Liang, M.; Xu, Z.; Lai, M.; Ren, H.; Deng, X.; Su, X. Downregulation of long non-coding RNA MAFG-AS1 represses tumorigenesis of colorectal cancer cells through the microRNA-149-3p-dependent inhibition of HOXB8. Cancer Cell Int. 2020, 20, 511. [Google Scholar] [CrossRef] [PubMed]
- Lai, F.; Deng, W.; Fu, C.; Wu, P.; Cao, M.; Tan, S. Long non-coding RNA SNHG6 increases JAK2 expression by targeting the miR-181 family to promote colorectal cancer cell proliferation. J. Gene Med. 2020, 22, e3262. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Li, H.; Zhang, H.; Zhang, C.; Zhu, Z.; Liang, H.; Zhou, Y. Long noncoding RNA RHPN1-AS1 promotes colorectal cancer progression via targeting miR-7-5p/OGT axis. Cancer Cell Int. 2020, 20, 54. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Nie, P.; Xu, S. Long noncoding RNA CASC21 exerts an oncogenic role in colorectal cancer through regulating miR-7-5p/YAP1 axis. Biomed. Pharmacother. 2020, 121, 109628. [Google Scholar] [CrossRef]
- Yan, F.; Ma, Y.; Liu, L.; Li, L.; Deng, J.; Sun, J. Long noncoding RNA HOXD-AS1 promotes the proliferation, migration, and invasion of colorectal cancer via the miR-526b-3p/CCND1 axis. J. Surg. Res. 2020, 255, 525–535. [Google Scholar] [CrossRef]
- Zhu, X.; Bu, F.; Tan, T.; Luo, Q.; Zhu, J.; Lin, K.; Huang, J.; Luo, C.; Zhu, Z. Long noncoding RNA RP11-757G1. 5 sponges miR-139-5p and upregulates YAP1 thereby promoting the proliferation and liver, spleen metastasis of colorectal cancer. J. Exp. Clin. Cancer Res. 2020, 39, 207. [Google Scholar] [CrossRef]
- Huang, F.K.; Zheng, C.Y.; Huang, L.K.; Lin, C.Q.; Zhou, J.F.; Wang, J.X. Long non-coding RNA MCF2L-AS1 promotes the aggressiveness of colorectal cancer by sponging miR-874-3p and thereby up-regulating CCNE1. J. Gene Med. 2021, 23, e3285. [Google Scholar] [CrossRef]
- Lu, W.; Huang, Z.; Wang, J.; Liu, H. Long non-coding RNA DANCR accelerates colorectal cancer progression via regulating the miR-185-5p/HMGA2 axis. J. Biochem. 2022, 171, 389–398. [Google Scholar] [CrossRef]
- Xu, G.; Wang, H.; Yuan, D.; Yao, J.; Meng, L.; Li, K.; Zhang, Y.; Dang, C.; Zhu, K. RUNX1-activated upregulation of lncRNA RNCR3 promotes cell proliferation, invasion, and suppresses apoptosis in colorectal cancer via miR-1301-3p/AKT1 axis in vitro and in vivo. Clin. Transl. Oncol. 2020, 22, 1762–1777. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Pan, B.; Liu, X.; Qin, J.; Wang, X.; He, B.; Pan, Y.; Sun, H.; Xu, T.; Xu, X. Long intergenic non-coding RNA LINC00485 exerts tumor-suppressive activity by regulating miR-581/EDEM1 axis in colorectal cancer. Aging 2021, 13, 3866. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zhao, H.; Song, Q.; Li, G.; Lin, S.; Xiong, S. Long non-coding RNA DPP10-AS1 exerts anti-tumor effects on colon cancer via the upregulation of ADCY1 by regulating microRNA-127-3p. Aging 2021, 13, 9748. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Wang, Y.; Liu, L.; Li, S.; Yu, W. Long non-coding RNA MBNL1-AS1 regulates proliferation, migration, and invasion of cancer stem cells in colon cancer by interacting with MYL9 via sponging microRNA-412-3p. Clin. Res. Hepatol. Gastroenterol. 2020, 44, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.L.; Xiao, F.; Liu, Y.F.; Chen, H.; Guo, G.C. Long non-coding RNA FENDRR restrains the aggressiveness of CRC via regulating miR-18a-5p/ING4 axis. J. Cell. Biochem. 2020, 121, 3973–3985. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Zeng, W.; Lee, D. lncRNA MCM3AP-AS1 inhibits the progression of colorectal cancer via the miR-19a-3p/FOXF2 axis. J. Gene Med. 2021, 23, e3306. [Google Scholar] [CrossRef] [PubMed]
- Chu, Q.; Zheng, W.; Su, H.; Zhang, L.; Chang, R.; Gao, W.; Xu, T. A highly conserved circular RNA, circRasGEF1B, enhances antiviral immunity by regulating the miR-21-3p/MITA pathway in lower vertebrates. Virol. J. 2021, 95, e02145-20. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Zhang, B.; Sun, C.; Duan, H.Q.; Liu, W.X.; Mu, K.; Zhao, L.; Li, H.R.; Dong, Z.Y.; Cui, Q. Circular RNA derived from TIMP2 functions as a competitive endogenous RNA and regulates intervertebral disc degeneration by targeting miR-185-5p and matrix metalloproteinase 2. Int. J. Mol. Med. 2020, 46, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Ren, H.; Guo, M.; Qian, J.; Yang, Y.; Gu, C. Review on circular RNAs and new insights into their roles in cancer. Comput. Struct. Biotechnol. J. 2021, 19, 910–928. [Google Scholar] [CrossRef]
- Zhou, W.-Y.; Cai, Z.-R.; Liu, J.; Wang, D.-S.; Ju, H.-Q.; Xu, R.-H. Circular RNA: Metabolism, functions and interactions with proteins. Mol. Cancer 2020, 19, 172. [Google Scholar] [CrossRef]
- Huang, Y.; Zhu, Q. Mechanisms regulating abnormal circular RNA biogenesis in cancer. Cancers 2021, 13, 4185. [Google Scholar] [CrossRef] [PubMed]
- Pitolli, C.; Marini, A.; Sette, C.; Pagliarini, V. Non-canonical splicing and its implications in brain physiology and cancer. Int. J. Mol. Sci. 2022, 23, 2811. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Meng, Y.; Jin, Y. RNA structures in alternative splicing and back-splicing. Wiley Interdiscip. Rev. RNA 2021, 12, e1626. [Google Scholar] [CrossRef] [PubMed]
- Van Der Steen, N.; Lyu, Y.; Hitzler, A.K.; Becker, A.C.; Seiler, J.; Diederichs, S. The circular RNA landscape of non-small cell lung cancer cells. Cancers 2020, 12, 1091. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhong, Y.; Wang, X.; Shen, J.; An, W. Advances in circular RNA and its applications. Int. J. Med. Sci. 2022, 19, 975. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, X.; Wang, Y.-Z.; Tian, H.; Xu, S. Research progress of circular RNAs in lung cancer. Cancer Biol. Ther. 2019, 20, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-L. The expanding regulatory mechanisms and cellular functions of circular RNAs. Nat. Rev. Mol. Cell Biol. 2020, 21, 475–490. [Google Scholar] [CrossRef] [PubMed]
- Robic, A.; Kühn, C. Beyond back splicing, a still poorly explored world: Non-canonical circular RNAs. Genes 2020, 11, 1111. [Google Scholar] [CrossRef]
- Huang, A.; Zheng, H.; Wu, Z.; Chen, M.; Huang, Y. Circular RNA-protein interactions: Functions, mechanisms, and identification. Theranostics 2020, 10, 3503. [Google Scholar] [CrossRef]
- Yang, Y.; Yujiao, W.; Fang, W.; Linhui, Y.; Ziqi, G.; Zhichen, W.; Zirui, W.; Shengwang, W. The roles of miRNA, lncRNA and circRNA in the development of osteoporosis. Biol. Res. 2020, 53, 40. [Google Scholar] [CrossRef]
- Li, T.; Wang, W.C.; McAlister, V.; Zhou, Q.; Zheng, X. Circular RNA in colorectal cancer. J. Cell. Mol. Med. 2021, 25, 3667–3679. [Google Scholar] [CrossRef] [PubMed]
- Bhuyan, R.; Bagchi, A. Prediction of the differentially expressed circRNAs to decipher their roles in the onset of human colorectal cancers. Gene 2020, 762, 145035. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Hann, S.S. Biological roles and mechanisms of circular RNA in human cancers. Onco Targets Ther. 2020, 13, 2067–2092. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xu, Y.; Yamaguchi, K.; Hu, J.; Zhang, L.; Wang, J.; Tian, J.; Chen, W. Circular RNA circVAPA knockdown suppresses colorectal cancer cell growth process by regulating miR-125a/CREB5 axis. Cancer Cell Int. 2020, 20, 103. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Luo, J.; Liu, G.; Li, X. Circular RNA hsa_circ_0008285 inhibits colorectal cancer cell proliferation and migration via the miR-382-5p/PTEN axis. Biochem. Biophys. Res. Commun. 2020, 527, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Zheng, X.; Hu, W.; Wang, Q.; Xu, Y.; He, W.; Wu, C.; Zhu, D.; Wu, C.; Jiang, J. Has_circ_0009361 acts as the sponge of miR-582 to suppress colorectal cancer progression by regulating APC2 expression. Clin. Sci. 2019, 133, 1197–1213. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Yao, Y.; Yang, N.; Gong, L.; Kong, Y.; Wu, A. Circular RNA circCTNNA1 promotes colorectal cancer progression by sponging miR-149-5p and regulating FOXM1 expression. Cell Death Dis. 2020, 11, 557. [Google Scholar] [CrossRef] [PubMed]
- Hao, S.; Qu, R.; Hu, C.; Wang, M.; Li, Y. A circular RNA derived from golgi glycoprotein 1 mRNA regulates KRAS expression and promotes colorectal cancer progression by targeting microRNA-622. Onco Targets Ther. 2020, 13, 12637–12648. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Liu, W.; Liu, Y.; Zhu, X. Circular RNA circ_0065378 upregulates tumor suppressor candidate 1 by competitively binding with miR-4701-5p to alleviate colorectal cancer progression. J. Gastroenterol. Hepatol. 2022, 37, 1107–1118. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Y.; Song, H.; Yin, H.; Jiang, T.; Xu, Y.; Liu, L.; Wang, H.; Gao, H.; Wang, R. The circular RNA circSPARC enhances the migration and proliferation of colorectal cancer by regulating the JAK/STAT pathway. Mol. Cancer 2021, 20, 81. [Google Scholar] [CrossRef]
- Yin, W.; Xu, J.; Li, C.; Dai, X.; Wu, T.; Wen, J. Circular RNA circ_0007142 facilitates colorectal cancer progression by modulating CDC25A expression via miR-122-5p. Onco Targets Ther. 2020, 13, 3689–3701. [Google Scholar] [CrossRef] [PubMed]
- Wen, T.; Wu, H.; Zhang, L.; Li, K.; Xiao, X.; Zhang, L.; Zhang, Y. Circular RNA circ_0007142 regulates cell proliferation, apoptosis, migration and invasion via miR-455-5p/SGK1 axis in colorectal cancer. Anti-Cancer Drugs 2021, 32, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Pan, Z. Circular RNA circ_0000467 regulates colorectal cancer development via miR-382-5p/EN2 axis. Bioengineered 2021, 12, 886–897. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Zhang, Z.; Wang, S.; Zhu, W.; Zheng, P.; Wang, W. Circular RNA hsa_circ_0001178 facilitates the invasion and metastasis of colorectal cancer through upregulating ZEB1 via sponging multiple miRNAs. Biol. Chem. 2020, 401, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Liu, M. Circular RNA hsa_circ_0053277 promotes the development of colorectal cancer by upregulating matrix metallopeptidase 14 via miR-2467-3p sequestration. J. Cell. Physiol. 2020, 235, 2881–2890. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ren, Y.; Ma, S.; Wang, S. Circular RNA 0060745, a novel circRNA, promotes colorectal cancer cell proliferation and metastasis through miR-4736 sponging. Onco Targets Ther. 2020, 13, 1941–1951. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhou, H. Circular RNA hsa_circRNA_102209 promotes the growth and metastasis of colorectal cancer through miR-761-mediated Ras and Rab interactor 1 signaling. Cancer Med. 2020, 9, 6710–6725. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Mou, Y.; Shi, X.; Liu, T.; Chen, Z.; Zuo, X. Circular RNA 100146 promotes colorectal cancer progression by the MicroRNA 149/HMGA2 Axis. Mol. Cell. Biol. 2021, 41, e00445-20. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Sun, H.; Liu, X.; Chen, J.; Tian, Z.; Xu, J.; Xiang, B.; Qin, B. Circular RNA hsa_circ_0004277 contributes to malignant phenotype of colorectal cancer by sponging miR-512-5p to upregulate the expression of PTMA. J. Cell. Physiol. 2020; ahead of print. [Google Scholar]
- Lu, C.; Fu, L.; Qian, X.; Dou, L.; Cang, S. Knockdown of circular RNA circ-FARSA restricts colorectal cancer cell growth through regulation of miR-330-5p/LASP1 axis. Arch. Biochem. Biophys. 2020, 689, 108434. [Google Scholar] [CrossRef]
- Hao, Q.; Zhang, Z. hsa_circRNA_000166 facilitated cell growth and limited apoptosis through targeting miR-326/LASP1 axis in colorectal cancer. Gastroent. Res. Pract. 2020, 2020, 8834359. [Google Scholar] [CrossRef]
- Yang, B.; Du, K.; Yang, C.; Xiang, L.; Xu, Y.; Cao, C.; Zhang, J.; Liu, W. CircPRMT5 circular RNA promotes proliferation of colorectal cancer through sponging miR-377 to induce E2F3 expression. J. Cell. Mol. Med. 2020, 24, 3431–3437. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-Y.; Wang, L.; Ren, Y.-X.; Pang, Z.; Liu, Y.; Sun, X.-D.; Tu, J.; Zhi, Z.; Qin, Y.; Sun, L.-N. The circular RNA circ-ERBIN promotes growth and metastasis of colorectal cancer by miR-125a-5p and miR-138-5p/4EBP-1 mediated cap-independent HIF-1α translation. Mol. Cancer 2020, 19, 164. [Google Scholar] [CrossRef]
- Zhao, J.P.; Chen, L.L. Circular RNA MAT2B induces colorectal cancer proliferation via sponging miR-610, resulting in an increased E2F1 expression. Cancer Manag. Res. 2020, 12, 7107–7116. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Yang, S.; Wang, J. Circ_0084615 is an oncogenic circular RNA in colorectal cancer and promotes DNMT3A expression via repressing miR-599. Ppathol. Res. Pract. 2021, 224, 153494. [Google Scholar] [CrossRef]
- Zhang, Z.-J.; Zhang, Y.-H.; Qin, X.-J.; Wang, Y.-X.; Fu, J. Circular RNA circDENND4C facilitates proliferation, migration and glycolysis of colorectal cancer cells through miR-760/GLUT1 axis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 2387–2400. [Google Scholar]
- Chen, F.; Guo, L.; Di, J.; Li, M.; Dong, D.; Pei, D. Circular RNA ubiquitin-associated protein 2 enhances autophagy and promotes colorectal cancer progression and metastasis via miR-582-5p/FOXO1 signaling. J. Genet. Genom. 2021, 48, 1091–1103. [Google Scholar] [CrossRef]
- Fang, G.; Chen, T.; Mao, R.; Huang, X.; Ji, L. Circular RNA circ_0089153 acts as a competing endogenous RNA to regulate colorectal cancer development by the miR-198/SUMO-specific peptidase 1 (SENP1) axis. Bioengineered 2021, 12, 5664–5678. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Tao, G.; Huang, D.; Liang, S.; Zheng, D. Circular RNA NOX4 promotes the development of colorectal cancer via the microRNA-485-5p/CKS1B axis. Oncol. Rep. 2020, 44, 2009–2020. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Chu, Z.; Lai, W.; Lan, Q.; Zeng, Y.; Lu, D.; Jin, S.; Xu, H.; Su, P.; Yin, D. Circular RNA circHERC4 as a novel oncogenic driver to promote tumor metastasis via the miR-556-5p/CTBP2/E-cadherin axis in colorectal cancer. J. Hematol. Oncol. 2021, 14, 194. [Google Scholar] [CrossRef]
- Liu, X.; Qin, Y.; Tang, X.; Wang, Y.; Bian, C.; Zhong, J. Circular RNA circ_0000372 contributes to the proliferation, migration and invasion of colorectal cancer by elevating IL6 expression via sponging miR-495. Anti-Cancer Drugs 2021, 32, 296–305. [Google Scholar] [CrossRef]
- Wang, R.; Wang, J.; Chen, Y.; Chen, Y.; Xi, Q.; Sun, L.; Zhang, X.; Zhang, G.; Ding, X.; Shi, T. Circular RNA circLDLR facilitates cancer progression by altering the miR-30a-3p/SOAT1 axis in colorectal cancer. Cell Death Discov. 2022, 8, 314. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.-L.; Liu, G.-Q.; Li, P.; Li, X.-H. Circular RNA CUL2 regulates the development of colorectal cancer by modulating apoptosis and autophagy via miR-208a-3p/PPP6C. Aging 2022, 14, 497. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.; Xi, Z.; Zhang, Y.; Li, Z.; Huang, L.; Xin, T.; Shen, R.; Wang, T. Circ-SMARCA5 suppresses colorectal cancer progression via downregulating miR-39-3p and upregulating ARID4B. Dig. Liver Dis. 2020, 52, 1494–1502. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.; Feng, X.; Yuan, H.; Gong, Y.; Wang, G. Circ_0003266 sponges miR-503-5p to suppress colorectal cancer progression via regulating PDCD4 expression. BMC Cancer 2021, 21, 284. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.J.; Kim, W.R.; Park, E.G.; Lee, D.H.; Kim, J.-m.; Shin, H.J.; Jeong, H.-s.; Roh, H.-Y.; Kim, H.-S. Exploring the Key Signaling Pathways and ncRNAs in Colorectal Cancer. Int. J. Mol. Sci. 2024, 25, 4548. https://doi.org/10.3390/ijms25084548
Lee YJ, Kim WR, Park EG, Lee DH, Kim J-m, Shin HJ, Jeong H-s, Roh H-Y, Kim H-S. Exploring the Key Signaling Pathways and ncRNAs in Colorectal Cancer. International Journal of Molecular Sciences. 2024; 25(8):4548. https://doi.org/10.3390/ijms25084548
Chicago/Turabian StyleLee, Yun Ju, Woo Ryung Kim, Eun Gyung Park, Du Hyeong Lee, Jung-min Kim, Hae Jin Shin, Hyeon-su Jeong, Hyun-Young Roh, and Heui-Soo Kim. 2024. "Exploring the Key Signaling Pathways and ncRNAs in Colorectal Cancer" International Journal of Molecular Sciences 25, no. 8: 4548. https://doi.org/10.3390/ijms25084548