
An In Vitro Study on the Cytotoxic, Antioxidant, and Antimicrobial Properties of Yamogenin—A Plant Steroidal Saponin and Evaluation of Its Mechanism of Action in Gastric Cancer Cells
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Cytotoxic Effect of Yamogenin and in Combination with Oxaliplatin or Capecitabine
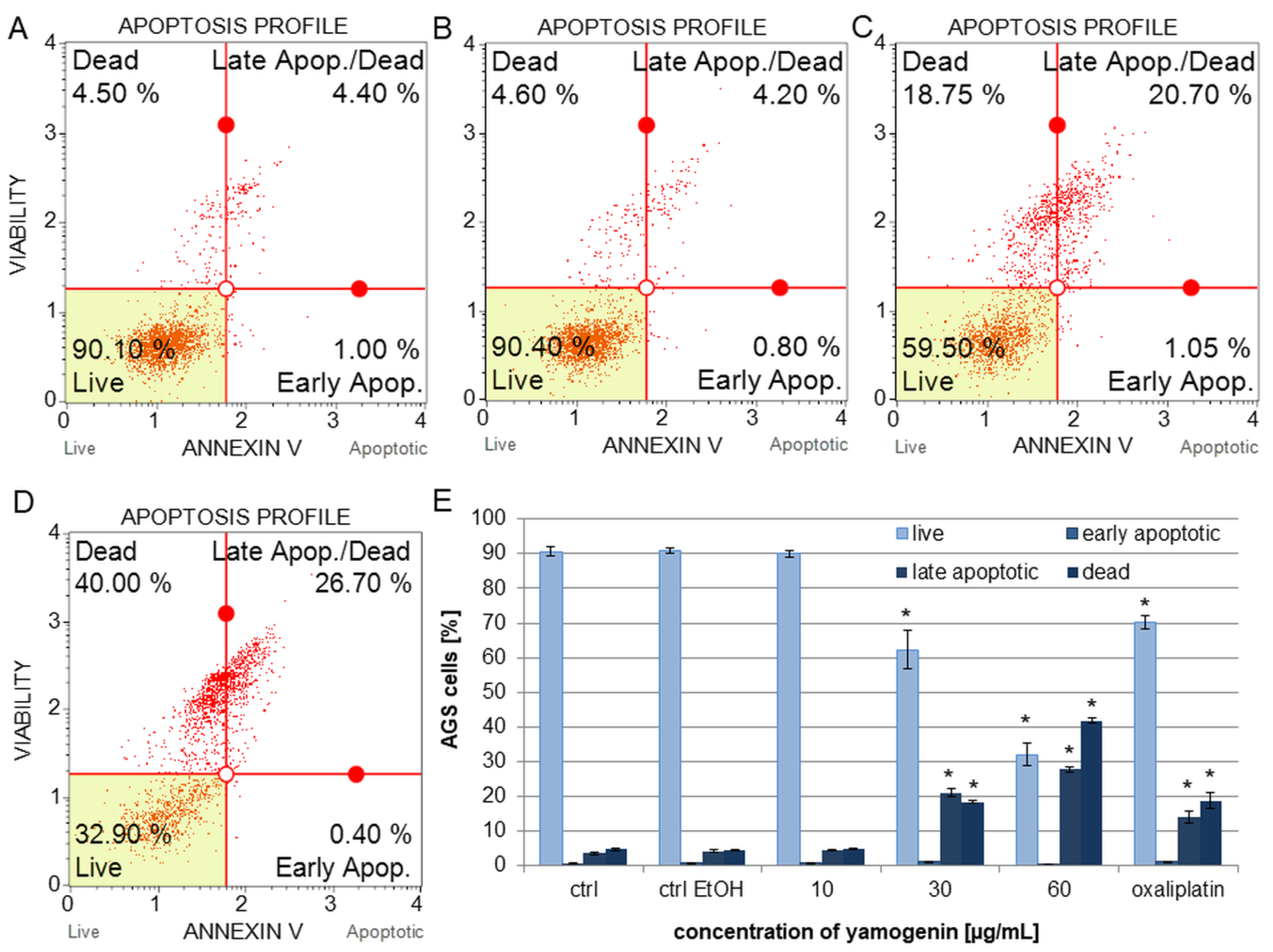
2.2. Yamogenin Increased the Amount of Apoptotic and Dead AGS Cells
2.3. Yamogenin Decreased Mitochondrial Potential in AGS Cells
2.4. Yamogenin Increased the Level of Reactive Oxygen Species (ROS) in AGS Cells
2.5. Yamogenin Induced Cell Cycle Arrest in subG1 Phase in AGS Cells
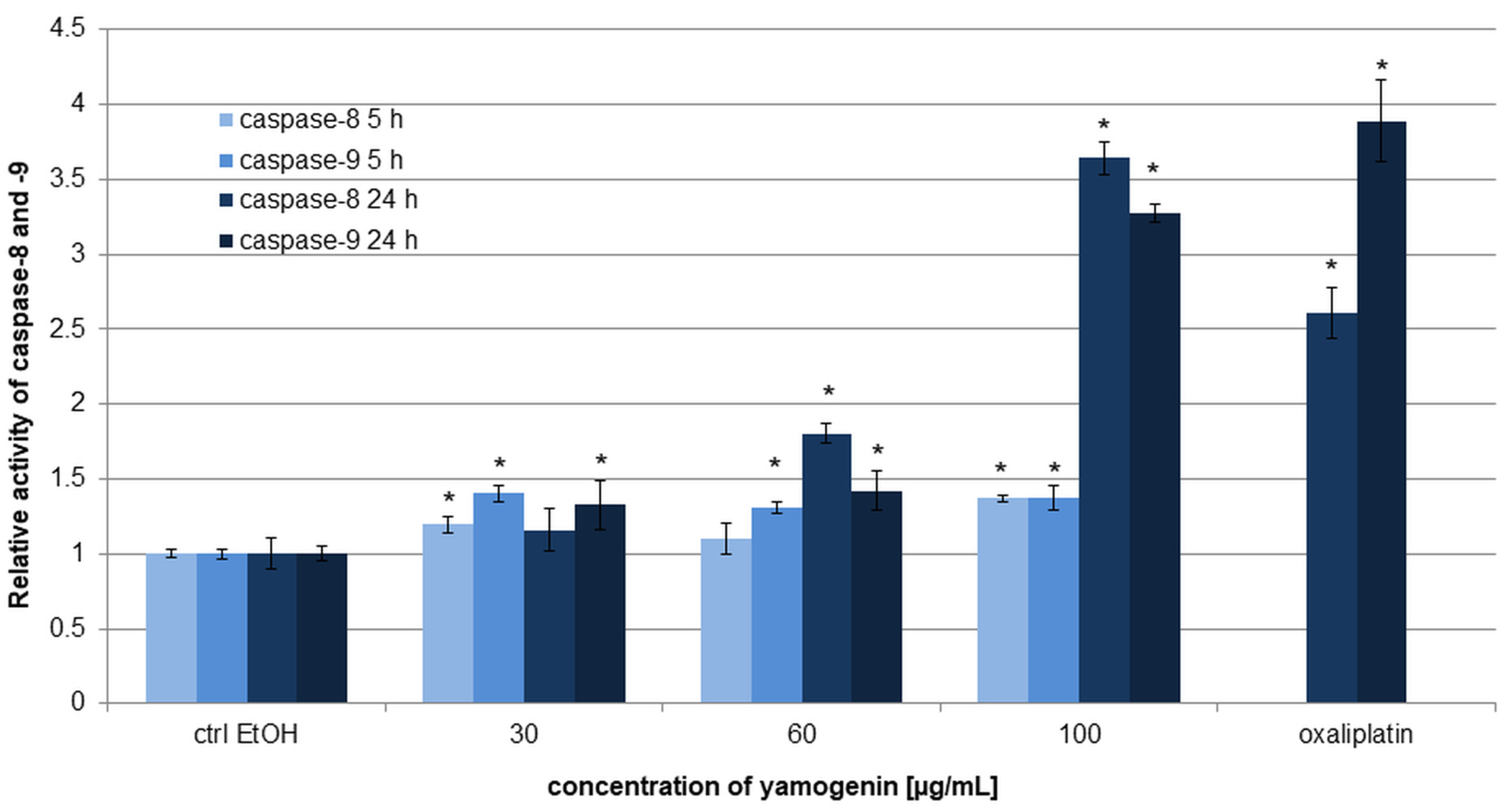
2.6. Yamogenin Triggered Activation of Caspase-8 and -9 in AGS Cells
2.7. The Effect of Yamogenin on Expression of Genes at mRNA Level in AGS Cells
2.8. Antioxidant Activity of Yamogenin and Inhibition of Protein Denaturation
2.8.1. Antiradical Potential of Yamogenin
2.8.2. Inhibition of Protein Denaturation by Yamogenin
2.9. Antimicrobial Activity of Yamogenin
3. Discussion
4. Materials and Methods
4.1. Preparation of Yamogenin Solution
4.2. Cell Culture
4.3. MTT Assay
4.4. Annexin and Dead Cell Assay
4.5. Cell Cycle Analysis of AGS Cells Treated with Yamogenin
4.6. Estimation of Mitochondria Depolarization in AGS Cells Treated with Yamogenin
4.7. Reactive Oxygen Species (ROS) Production in AGS Cells Treated with Yamogenin
4.8. Caspases-8/9 Activity in AGS Cells Treated with Yamogenin
4.9. RT-PCR Analysis of Gene Expression at mRNA Level in AGS Cells Treated with Yamogenin
4.10. Antioxidant and Inhibition of Protein Denaturation Assays
4.10.1. Materials
4.10.2. DPPH Assay
4.10.3. ABTS Assay
4.10.4. Inhibition of Protein Denaturation
4.11. Antimicrobial Activity of Yamogenin
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xie, Y.; Shi, L.; He, X.; Luo, Y. Gastrointestinal Cancers in China, the USA, and Europe. Gastroenterol. Rep. 2021, 9, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Keum, N.; Giovannucci, E. Global Burden of Colorectal Cancer: Emerging Trends, Risk Factors and Prevention Strategies. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 713–732. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Forman, D.; Waskito, L.; Yamaoka, Y.; Crabtree, J. Epidemiology of Helicobacter pylori and CagA-Positive Infections and Global Variations in Gastric Cancer. Toxins 2018, 10, 163. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Xu, J.; Wang, Y.; Xiang, L.; He, X. Total Saponins from Tupistra Chinensis Baker Inhibits Growth of Human Gastric Cancer Cells in Vitro and in Vivo. J. Ethnopharmacol. 2021, 278, 114323. [Google Scholar] [CrossRef]
- Desai, A.; Qazi, G.; Ganju, R.; El-Tamer, M.; Singh, J.; Saxena, A.; Bedi, Y.; Taneja, S.; Bhat, H. Medicinal Plants and Cancer Chemoprevention. Curr. Drug Metab. 2008, 9, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Esmeeta, A.; Adhikary, S.; Dharshnaa, V.; Swarnamughi, P.; Ummul Maqsummiya, Z.; Banerjee, A.; Pathak, S.; Duttaroy, A.K. Plant-Derived Bioactive Compounds in Colon Cancer Treatment: An Updated Review. Biomed. Pharmacother. 2022, 153, 113384. [Google Scholar] [CrossRef] [PubMed]
- Podolak, I.; Galanty, A.; Sobolewska, D. Saponins as Cytotoxic Agents: A Review. Phytochem. Rev. 2010, 9, 425–474. [Google Scholar] [CrossRef] [PubMed]
- Rehan, M.; Shafiullah; Mir, S.A. Structural Diversity, Natural Sources, and Pharmacological Potential of Plant-Based Saponins with Special Focus on Anticancer Activity: A Review. Med. Chem. Res. 2020, 29, 1707–1722. [Google Scholar] [CrossRef]
- Stefanowicz-Hajduk, J.; Hering, A.; Gucwa, M.; Czerwińska, M.; Ochocka, J.R. Yamogenin-Induced Cell Cycle Arrest, Oxidative Stress, and Apoptosis in Human Ovarian Cancer Cell Line. Molecules 2022, 27, 8181. [Google Scholar] [CrossRef]
- Lepage, C.; Léger, D.Y.; Bertrand, J.; Martin, F.; Beneytout, J.L.; Liagre, B. Diosgenin Induces Death Receptor-5 through Activation of P38 Pathway and Promotes TRAIL-Induced Apoptosis in Colon Cancer Cells. Cancer Lett. 2011, 301, 193–202. [Google Scholar] [CrossRef]
- Raju, J.; Patlolla, J.M.R.; Swamy, M.V.; Rao, C.V. Diosgenin, a Steroid Saponin of Trigonella Foenum Graecum (Fenugreek), Inhibits Azoxymethane-Induced Aberrant Crypt Foci Formation in F344 Rats and Induces Apoptosis in HT-29 Human Colon Cancer Cells. Cancer Epidemiol. Biomark. Prev. 2004, 13, 1392–1398. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Xu, Y.; Hua, C.; Li, C.; Zhang, Y. Molecular Cloning and Functional Characterization of a Sterol 3-O-Glucosyltransferase Involved in Biosynthesis of Steroidal Saponins in Trigonella foenum-graecum. Front. Plant Sci. 2021, 12, 809579. [Google Scholar] [CrossRef] [PubMed]
- Stefanowicz-Hajduk, J.; Król-Kogus, B.; Sparzak-Stefanowska, B.; Kimel, K.; Ochocka, J.R.; Krauze-Baranowska, M. Cytotoxic Activity of Standardized Extracts, a Fraction, and Individual Secondary Metabolites from Fenugreek Seeds against SKOV-3, HeLa and MOLT-4 Cell Lines. Pharm. Biol. 2021, 59, 422–435. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Fan, L. Improving the Bioactive Ingredients and Functions of Asparagus from Efficient to Emerging Processing Technologies: A Review. Food Chem. 2021, 358, 129903. [Google Scholar] [CrossRef]
- Cheng, Q.; Zeng, L.; Wen, H.; Brown, S.E.; Wu, H.; Li, X.; Lin, C.; Liu, Z.; Mao, Z. Steroidal Saponin Profiles and Their Key Genes for Synthesis and Regulation in Asparagus officinalis L. by Joint Analysis of Metabolomics and Transcriptomics. BMC Plant Biol. 2023, 23, 207. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Wang, N.; Liu, H.; Li, Z.; Lu, L.; Wang, C. The Bioactive Compounds and Biological Functions of Asparagus officinalis L.—A Review. J. Funct. Foods 2020, 65, 103727. [Google Scholar] [CrossRef]
- Wei, R.; Qin, X.; Li, Z. Comparison of the Inedible Parts of White and Green Asparagus Based on Metabolomics and Network Pharmacology. Food Funct. 2023, 14, 7478–7488. [Google Scholar] [CrossRef]
- Caraux, G.; Pinloche, S. PermutMatrix: A Graphical Environment to Arrange Gene Expression Profiles in Optimal Linear Order. Bioinformatics 2005, 21, 1280–1281. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Cavalcante, G.C.; Schaan, A.P.; Cabral, G.F.; Santana-da-Silva, M.N.; Pinto, P.; Vidal, A.F.; Ribeiro-dos-Santos, Â. A Cell’s Fate: An Overview of the Molecular Biology and Genetics of Apoptosis. Int. J. Mol. Sci. 2019, 20, 4133. [Google Scholar] [CrossRef]
- Peter, M.E.; Krammer, P.H. Mechanisms of CD95 (APO-1/Fas)-Mediated Apoptosis. Curr. Opin. Immunol. 1998, 10, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Suliman, A.; Lam, A.; Datta, R.; Srivastava, R.K. Intracellular Mechanisms of TRAIL: Apoptosis through Mitochondrial-Dependent and -Independent Pathways. Oncogene 2001, 20, 2122–2133. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.; Xiong, J.; Goeddel, D.V. The TNF Receptor 1-Associated Protein TRADD Signals Cell Death and NF-κB Activation. Cell 1995, 81, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Wajant, H. The Fas Signaling Pathway: More Than a Paradigm. Science 2002, 296, 1635–1636. [Google Scholar] [CrossRef] [PubMed]
- Kischkel, F.C.; Hellbardt, S.; Behrmann, I.; Germer, M.; Pawlita, M.; Krammer, P.H.; Peter, M.E. Cytotoxicity-Dependent APO-1 (Fas/CD95)-Associated Proteins Form a Death-Inducing Signaling Complex (DISC) with the Receptor. EMBO J. 1995, 14, 5579–5588. [Google Scholar] [CrossRef] [PubMed]
- Tait, S.W.G.; Green, D.R. Mitochondria and Cell Death: Outer Membrane Permeabilization and Beyond. Nat. Rev. Mol. Cell Biol. 2010, 11, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Kepp, O.; Trojel-Hansen, C.; Kroemer, G. Mitochondrial Control of Cellular Life, Stress, and Death. Circ. Res. 2012, 111, 1198–1207. [Google Scholar] [CrossRef] [PubMed]
- Shimura, T. Mitochondrial Signaling Pathways Associated with DNA Damage Responses. Int. J. Mol. Sci. 2023, 24, 6128. [Google Scholar] [CrossRef] [PubMed]
- Yao, M.; Li, R.; Yang, Z.; Ding, Y.; Zhang, W.; Li, W.; Liu, M.; Zhao, C.; Wang, Y.; Tang, H.; et al. PP9, a Steroidal Saponin, Induces G2/M Arrest and Apoptosis in Human Colorectal Cancer Cells by Inhibiting the PI3K/Akt/GSK3β Pathway. Chem. Biol. Interact. 2020, 331, 109246. [Google Scholar] [CrossRef]
- Wang, N.; Feng, Y.; Zhu, M.; Siu, F.-M.; Ng, K.-M.; Che, C.-M. A Novel Mechanism of XIAP Degradation Induced by Timosaponin AIII in Hepatocellular Carcinoma. Biochim. Biophys. Acta BBA—Mol. Cell Res. 2013, 1833, 2890–2899. [Google Scholar] [CrossRef]
- Corbiere, C.; Liagre, B.; Terro, F.; Beneytout, J.-L. Induction of Antiproliferative Effect by Diosgenin through Activation of P53, Release of Apoptosis-Inducing Factor (AIF) and Modulation of Caspase-3 Activity in Different Human Cancer Cells. Cell Res. 2004, 14, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Raju, J.; Bird, R.P. Diosgenin, a Naturally Occurring Furostanol Saponin Suppresses 3-Hydroxy-3-Methylglutaryl CoA Reductase Expression and Induces Apoptosis in HCT-116 Human Colon Carcinoma Cells. Cancer Lett. 2007, 255, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Coussens, L.M.; Werb, Z. Inflammation and Cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Greten, F.R.; Grivennikov, S.I. Inflammation and Cancer: Triggers, Mechanisms, and Consequences. Immunity 2019, 51, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Marino, P.; Pepe, G.; Basilicata, M.G.; Vestuto, V.; Marzocco, S.; Autore, G.; Procino, A.; Gomez-Monterrey, I.M.; Manfra, M.; Campiglia, P. Potential Role of Natural Antioxidant Products in Oncological Diseases. Antioxidants 2023, 12, 704. [Google Scholar] [CrossRef] [PubMed]
- Karami-Mohajeri, S.; Mohammadinejad, R.; Ashrafizadeh, M.; Mohamadi, N.; Mohajeri, M.; Sharififar, F. Diosgenin: Mechanistic Insights on Its Anti-Inflammatory Effects. Anti-Inflamm. Anti-Allergy Agents Med. Chem. 2021, 21, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Sarveswaran, R.; Jayasuriya, W.J.A.B.N.; Suresh, T.S. In Vitro Assays to Investigate the Anti-Inflammatory Activity of Herbal Extracts: A Review. World J. Pharm. Res. 2017, 6, 131–141. [Google Scholar] [CrossRef]
- Manivannan, J.; Shanthakumar, J.; Arunagiri, P.; Raja, B.; Balamurugan, E. Diosgenin Interferes Coronary Vasoconstriction and Inhibits Osteochondrogenic Transdifferentiation of Aortic VSMC in CRF Rats. Biochimie 2014, 102, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Michalak, O.; Krzeczyński, P.; Jaromin, A.; Cmoch, P.; Cybulski, M.; Trzcińska, K.; Miszta, P.; Mehta, P.; Gubernator, J.; Filipek, S. Antioxidant Activity of Novel Diosgenin Derivatives: Synthesis, Biological Evaluation, and in Silico ADME Prediction. Steroids 2022, 188, 109115. [Google Scholar] [CrossRef]
- Spiegel, M.; Krzyżek, P.; Dworniczek, E.; Adamski, R.; Sroka, Z. In Silico Screening and In Vitro Assessment of Natural Products with Anti-Virulence Activity against Helicobacter pylori. Molecules 2021, 27, 20. [Google Scholar] [CrossRef]
- Akash, S.; Bayıl, I.; Mahmood, S.; Mukerjee, N.; Mili, T.A.; Dhama, K.; Rahman, M.A.; Maitra, S.; Mohany, M.; Al-Rejaie, S.S.; et al. Mechanistic Inhibition of Gastric Cancer-Associated Bacteria Helicobacter pylori by Selected Phytocompounds: A New Cutting-Edge Computational Approach. Heliyon 2023, 9, e20670. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.d.S.; da Silva, A.R.P.; Araújo, N.J.S.; Filho, J.M.B.; Tavares, J.F.; de Freitas, T.S.; Junior, F.N.P.; de Sousa, E.O.; Maia, F.P.A.; de Vasconcelos, J.E.L.; et al. Evaluation of the Antibacterial and Inhibitory Activity of NorA and MepA Efflux Pumps from Staphylococcus aureus by Diosgenin. Life Sci. 2022, 308, 120978. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.-S.; Cai, L.; Li, Y.; Wang, J.-P.; Xiao, H.; Ding, Z.-T. Spirostanol Steroids from the Roots of Allium tuberosum. Steroids 2015, 100, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Fouedjou, R.T.; Teponno, R.B.; Quassinti, L.; Bramucci, M.; Petrelli, D.; Vitali, L.A.; Fiorini, D.; Tapondjou, L.A.; Barboni, L. Steroidal Saponins from the Leaves of Cordyline fruticosa (L.) A. Chev. and Their Cytotoxic and Antimicrobial Activity. Phytochem. Lett. 2014, 7, 62–68. [Google Scholar] [CrossRef]
- Qin, X.-J.; Sun, D.-J.; Ni, W.; Chen, C.-X.; Hua, Y.; He, L.; Liu, H.-Y. Steroidal Saponins with Antimicrobial Activity from Stems and Leaves of Paris polyphylla var. yunnanensis. Steroids 2012, 77, 1242–1248. [Google Scholar] [CrossRef] [PubMed]
- Bułakowska, A.; Sławiński, J.; Hering, A.; Gucwa, M.; Ochocka, J.R.; Hałasa, R.; Balewski, Ł.; Stefanowicz-Hajduk, J. New Chalcone Derivatives Containing 2,4-Dichlorobenzenesulfonamide Moiety with Anticancer and Antioxidant Properties. Int. J. Mol. Sci. 2023, 25, 274. [Google Scholar] [CrossRef] [PubMed]
- Stefanowicz-Hajduk, J.; Hering, A.; Gucwa, M.; Sztormowska-Achranowicz, K.; Kowalczyk, M.; Soluch, A.; Ochocka, J.R. An In Vitro Anticancer, Antioxidant, and Phytochemical Study on Water Extract of Kalanchoe daigremontiana Raym.-Hamet and H. Perrier. Molecules 2022, 27, 2280. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.H.; Wang, Z.; Hwang, S.H.; Kang, Y.-H.; Lee, J.-Y.; Lim, S.S. Comprehensive Evaluation of the Antioxidant Capacity of Perilla frutescens Leaves Extract and Isolation of Free Radical Scavengers Using Step-Wise HSCCC Guided by DPPH-HPLC. Int. J. Food Prop. 2017, 20, 921–934. [Google Scholar] [CrossRef]
- Chomnawang, M.T.; Surassmo, S.; Nukoolkarn, V.S.; Gritsanapan, W. Antimicrobial Effects of Thai Medicinal Plants against Acne-Inducing Bacteria. J. Ethnopharmacol. 2005, 101, 330–333. [Google Scholar] [CrossRef]
- Lekogo, B.M.; Coroller, L.; Mathot, A.G.; Mafart, P.; Leguerinel, I. Modelling the Influence of Palmitic, Palmitoleic, Stearic and Oleic Acids on Apparent Heat Resistance of Spores of Bacillus cereus NTCC 11145 and Clostridium sporogenes Pasteur 79.3. Int. J. Food Microbiol. 2010, 141, 242–247. [Google Scholar] [CrossRef]
- Hałasa, R.; Turecka, K.; Orlewska, C.; Werel, W. Comparison of Fluorescence Optical Respirometry and Microbroth Dilution Methods for Testing Antimicrobial Compounds. J. Microbiol. Methods 2014, 107, 98–105. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 µg/mL | ||
|---|---|---|
| Yamogenin | Ascorbic Acid | |
| DPPH | 704.7 ± 5.9 | 14.15 ± 0.13 |
| ABTS | 631.09 ± 3.51 | 7.33 ± 0.17 |
| Microorganism | [mg/mL] | Ampicillin [mg/mL] | |
|---|---|---|---|
| MIC | MBC | MIC | |
| Staphylococcus aureus ATCC6538 | 0.35 | >3.5 | 0.00008 |
| Klebsiella pneumoniae ATCC13883 | 3.5 | >3.5 | 0.001 |
| Escherichia coli ATCC8739 | 3.5 | >3.5 | 0.0039 |
| Salmonella enterica ATCC13076 | 3.5 | >3.5 | 0.0005 |
| Helicobacter pylori ATCC43504 | 3.5 | >3.5 | 0.0032 |
| Campylobacter jejuni ZMF | 3.5 | >3.5 | 0.032 |
| Campylobacter coli ZMF | 3.5 | >3.5 | 0.016 |
| Bacillus cereus PCM 1948, 2019 (ATCC11778) | 3.5 | >3.5 | 0.125 |
| Listeria monocytogenes PCM2191 | 3.5 | >3.5 | 0.016 |
| Lactobacillus paracasei PCM2639 | >3.5 | >3.5 | >0.125 |
| Lactobacillus acidophilus PCM2499 | >3.5 | >3.5 | >0.125 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stefanowicz-Hajduk, J.; Graczyk, P.; Hering, A.; Gucwa, M.; Nowak, A.; Hałasa, R. An In Vitro Study on the Cytotoxic, Antioxidant, and Antimicrobial Properties of Yamogenin—A Plant Steroidal Saponin and Evaluation of Its Mechanism of Action in Gastric Cancer Cells. Int. J. Mol. Sci. 2024, 25, 4627. https://doi.org/10.3390/ijms25094627
Stefanowicz-Hajduk J, Graczyk P, Hering A, Gucwa M, Nowak A, Hałasa R. An In Vitro Study on the Cytotoxic, Antioxidant, and Antimicrobial Properties of Yamogenin—A Plant Steroidal Saponin and Evaluation of Its Mechanism of Action in Gastric Cancer Cells. International Journal of Molecular Sciences. 2024; 25(9):4627. https://doi.org/10.3390/ijms25094627
Chicago/Turabian StyleStefanowicz-Hajduk, Justyna, Piotr Graczyk, Anna Hering, Magdalena Gucwa, Anna Nowak, and Rafał Hałasa. 2024. "An In Vitro Study on the Cytotoxic, Antioxidant, and Antimicrobial Properties of Yamogenin—A Plant Steroidal Saponin and Evaluation of Its Mechanism of Action in Gastric Cancer Cells" International Journal of Molecular Sciences 25, no. 9: 4627. https://doi.org/10.3390/ijms25094627