
A Pilot Immunohistochemical Study Identifies Hedgehog Pathway Expression in Sinonasal Adenocarcinoma


 and
and
Abstract
:1. Introduction
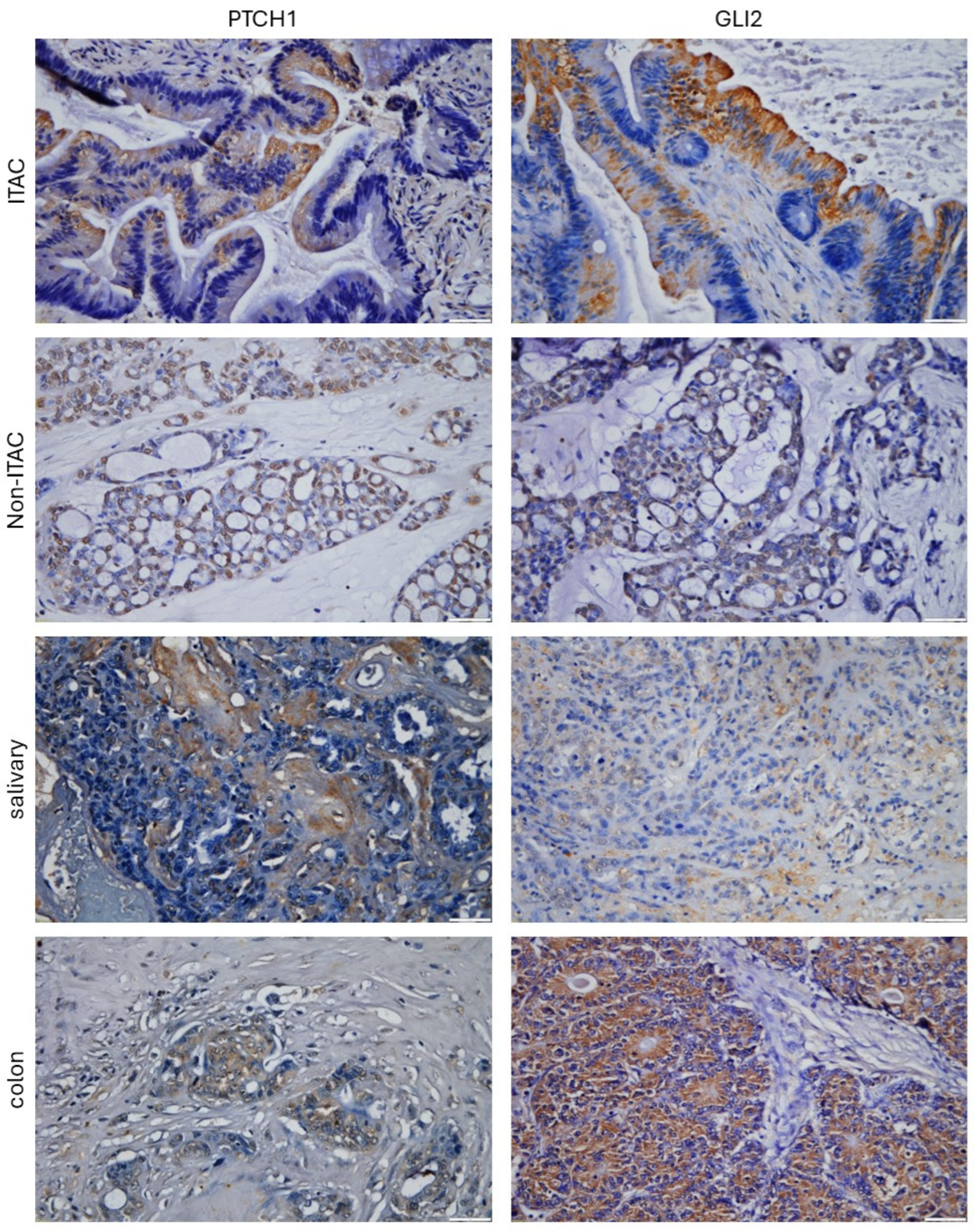
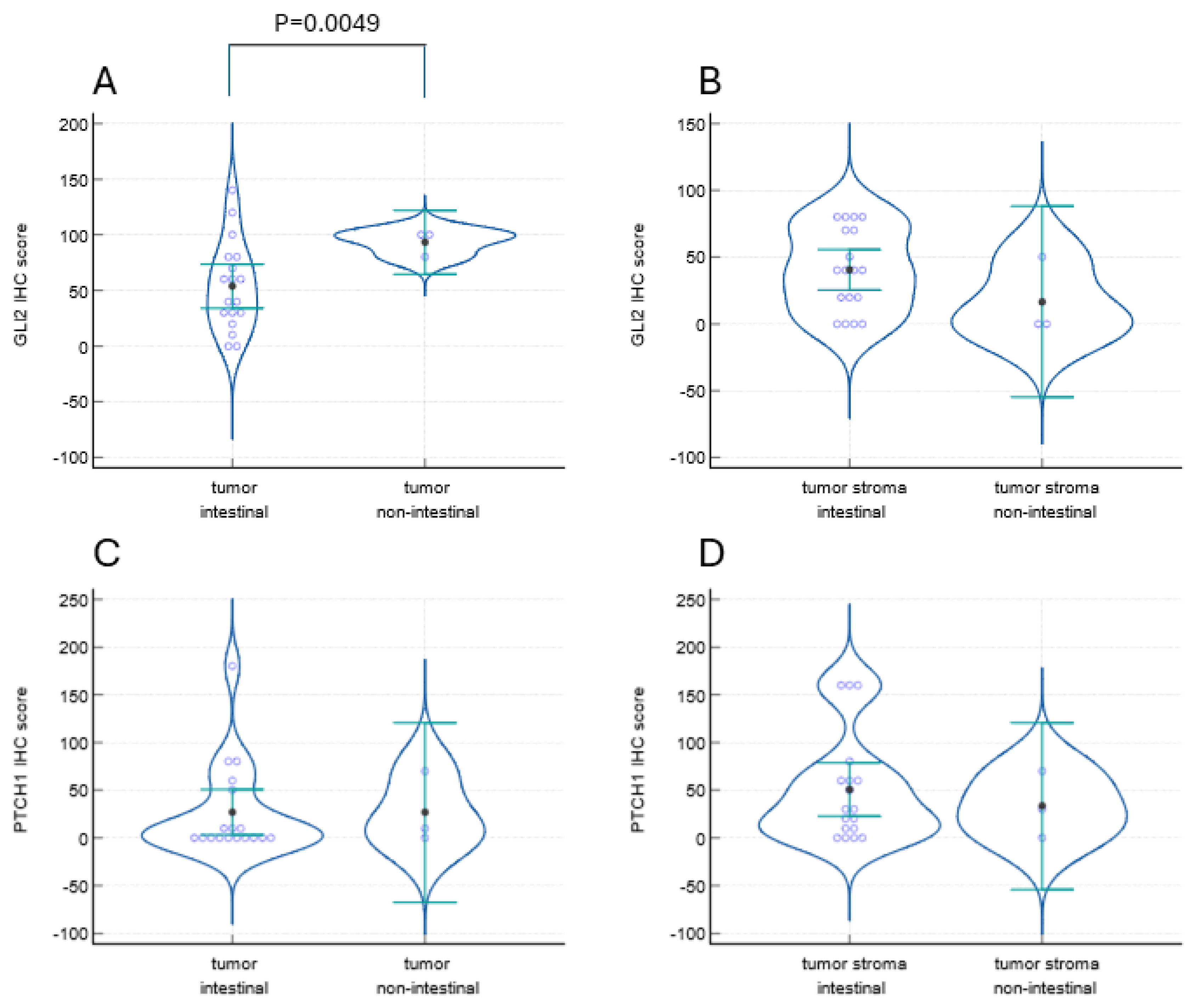
2. Results
3. Discussion
4. Materials and Methods
4.1. Immunohistochemical Staining
4.2. Slide Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gritli-Linde, A.; Lewis, P.; McMahon, A.P.; Linde, A. The Whereabouts of a Morphogen: Direct Evidence for Short- and Graded Long-Range Activity of Hedgehog Signaling Peptides. Dev. Biol. 2001, 236, 364–386. [Google Scholar] [CrossRef] [PubMed]
- Cobourne, M.T.; Green, J.B.A. Hedgehog Signalling in Development of the Secondary Palate. Front. Oral Biol. 2012, 16, 52–59. [Google Scholar] [CrossRef]
- Jaskoll, T.; Leo, T.; Witcher, D.; Ormestad, M.; Astorga, J.; Bringas, P.; Carlsson, P.; Melnick, M. Sonic Hedgehog Signaling Plays an Essential Role during Embryonic Salivary Gland Epithelial Branching Morphogenesis. Dev. Dyn. 2004, 229, 722–732. [Google Scholar] [CrossRef] [PubMed]
- Sagai, T.; Amano, T.; Maeno, A.; Kiyonari, H.; Seo, H.; Cho, S.-W.; Shiroishi, T. SHH Signaling Directed by Two Oral Epithelium-Specific Enhancers Controls Tooth and Oral Development. Sci. Rep. 2017, 7, 13004. [Google Scholar] [CrossRef] [PubMed]
- Abramyan, J. Hedgehog Signaling and Embryonic Craniofacial Disorders. J. Dev. Biol. 2019, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Rui, C.; Zhang, H.; Chen, J.; Jia, X.; Xiao, Y. Sonic Hedgehog Signaling in Epithelial Tissue Development. Regen. Med. Res. 2019, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Petrova, R.; Joyner, A.L. Roles for Hedgehog Signaling in Adult Organ Homeostasis and Repair. Development 2014, 141, 3445–3457. [Google Scholar] [CrossRef] [PubMed]
- Jing, J.; Wu, Z.; Wang, J.; Luo, G.; Lin, H.; Fan, Y.; Zhou, C. Hedgehog Signaling in Tissue Homeostasis, Cancers, and Targeted Therapies. Signal Transduct. Target. Ther. 2023, 8, 315. [Google Scholar] [CrossRef]
- Pettigrew, C.A.; Asp, E.; Emerson, C.P. A New Role for Hedgehogs in Juxtacrine Signaling. Mech. Dev. 2014, 131, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Ramsbottom, S.A.; Pownall, M.E. Regulation of Hedgehog Signalling Inside and Outside the Cell. J. Dev. Biol. 2016, 4, 23. [Google Scholar] [CrossRef]
- Weiss, L.E.; Milenkovic, L.; Yoon, J.; Stearns, T.; Moerner, W.E. Motional Dynamics of Single Patched1 Molecules in Cilia Are Controlled by Hedgehog and Cholesterol. Proc. Natl. Acad. Sci. USA 2019, 116, 5550–5557. [Google Scholar] [CrossRef]
- Leovic, D.; Sabol, M.; Ozretic, P.; Musani, V.; Car, D.; Marjanovic, K.; Zubcic, V.; Sabol, I.; Sikora, M.; Grce, M.; et al. Hh-Gli Signaling Pathway Activity in Oral and Oropharyngeal Squamous Cell Carcinoma. Head Neck 2012, 34, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.; Thurnher, D.; Kloimstein, P.; Leitner, V.; Petzelbauer, P.; Pammer, J.; Brunner, M.; Erovic, B.M. Expression of the Sonic Hedgehog Pathway in Squamous Cell Carcinoma of the Skin and the Mucosa of the Head and Neck. Head Neck 2011, 33, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Honami, T.; Shimo, T.; Okui, T.; Kurio, N.; Hassan, N.M.M.; Iwamoto, M.; Sasaki, A. Sonic Hedgehog Signaling Promotes Growth of Oral Squamous Cell Carcinoma Cells Associated with Bone Destruction. Oral Oncol. 2012, 48, 49–55. [Google Scholar] [CrossRef]
- Fan, H.-X.; Wang, S.; Zhao, H.; Liu, N.; Chen, D.; Sun, M.; Zheng, J.-H. Sonic Hedgehog Signaling May Promote Invasion and Metastasis of Oral Squamous Cell Carcinoma by Activating MMP-9 and E-Cadherin Expression. Med. Oncol. 2014, 31, 41. [Google Scholar] [CrossRef]
- Gan, G.N.; Eagles, J.; Keysar, S.B.; Wang, G.; Glogowska, M.J.; Altunbas, C.; Anderson, R.T.; Le, P.N.; Morton, J.J.; Frederick, B.; et al. Hedgehog Signaling Drives Radioresistance and Stroma-Driven Tumor Repopulation in Head and Neck Squamous Cancers. Cancer Res. 2014, 74, 7024–7036. [Google Scholar] [CrossRef]
- Wang, Y.-F.; Chang, C.-J.; Lin, C.-P.; Chang, S.-Y.; Chu, P.-Y.; Tai, S.-K.; Li, W.-Y.; Chao, K.S.C.; Chen, Y.-J. Expression of Hedgehog Signaling Molecules as a Prognostic Indicator of Oral Squamous Cell Carcinoma. Head Neck 2012, 34, 1556–1561. [Google Scholar] [CrossRef]
- Stelow, E.B.; Mills, S.E.; Jo, V.Y.; Carlson, D.L. Adenocarcinoma of the Upper Aerodigestive Tract. Adv. Anat. Pathol. 2010, 17, 262. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Bhatt, A.A.; Bathla, G.; Kanekar, S.; Soni, N.; Murray, J.; Vijay, K.; Vibhute, P.; Rhyner, P.H. Update from the 5th Edition of the WHO Classification of Nasal, Paranasal, and Skull Base Tumors: Imaging Overview with Histopathologic and Genetic Correlation. Am. J. Neuroradiol. 2023, 44, 1116–1125. [Google Scholar] [CrossRef]
- Rampinelli, V.; Ferrari, M.; Nicolai, P. Intestinal-Type Adenocarcinoma of the Sinonasal Tract: An Update. Curr. Opin. Otolaryngol. Head Neck Surg. 2018, 26, 115–121. [Google Scholar] [CrossRef]
- Leivo, I. Intestinal-Type Adenocarcinoma: Classification, Immunophenotype, Molecular Features and Differential Diagnosis. Head Neck Pathol. 2017, 11, 295–300. [Google Scholar] [CrossRef]
- Vivanco, B.; Llorente, J.L.; Perez-Escuredo, J.; Alvarez Marcos, C.; Fresno, M.F.; Hermsen, M.A. Benign Lesions in Mucosa Adjacent to Intestinal-Type Sinonasal Adenocarcinoma. Pathol. Res. Int. 2011, 2011, 230147. [Google Scholar] [CrossRef] [PubMed]
- Hoeben, A.; van de Winkel, L.; Hoebers, F.; Kross, K.; Driessen, C.; Slootweg, P.; Tjan-Heijnen, V.C.G.; van Herpen, C. Intestinal-Type Sinonasal Adenocarcinomas: The Road to Molecular Diagnosis and Personalized Treatment. Head Neck 2016, 38, 1564–1570. [Google Scholar] [CrossRef] [PubMed]
- Zappia, J.J.; Wolf, G.T.; McClatchey, K.D. Signet-Ring Cell Adenocarcinoma Metastatic to the Maxillary Sinus. Oral Surg. Oral Med. Oral Pathol. 1992, 73, 89–90. [Google Scholar] [CrossRef] [PubMed]
- Cantu, G.; Solero, C.L.; Mariani, L.; Lo Vullo, S.; Riccio, S.; Colombo, S.; Pompilio, M.; Perrone, F.; Formillo, P.; Quattrone, P. Intestinal Type Adenocarcinoma of the Ethmoid Sinus in Wood and Leather Workers: A Retrospective Study of 153 Cases. Head Neck 2011, 33, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.H.; Reh, D.D. Incidence and Survival in Patients with Sinonasal Cancer: A Historical Analysis of Population-Based Data. Head Neck 2012, 34, 877–885. [Google Scholar] [CrossRef]
- Nicolai, P.; Schreiber, A.; Bolzoni Villaret, A.; Lombardi, D.; Morassi, L.; Raffetti, E.; Donato, F.; Battaglia, P.; Turri–Zanoni, M.; Bignami, M.; et al. Intestinal Type Adenocarcinoma of the Ethmoid: Outcomes of a Treatment Regimen Based on Endoscopic Surgery with or without Radiotherapy. Head Neck 2016, 38, E996–E1003. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.N.; Maina, I.W.; Kuan, E.C.; Triantafillou, V.; Trope, M.A.; Carey, R.M.; Workman, A.D.; Tong, C.C.; Kohanski, M.A.; Palmer, J.N.; et al. Adenocarcinoma of the Sinonasal Tract: A Review of the National Cancer Database. J. Neurol. Surg. B Skull Base 2020, 81, 701–708. [Google Scholar] [CrossRef]
- Buller, N.V.J.A.; Rosekrans, S.L.; Westerlund, J.; van den Brink, G.R. Hedgehog Signaling and Maintenance of Homeostasis in the Intestinal Epithelium. Physiology 2012, 27, 148–155. [Google Scholar] [CrossRef]
- Walton, K.D.; Gumucio, D.L. Hedgehog Signaling in Intestinal Development and Homeostasis. Annu. Rev. Physiol. 2021, 83, 359–380. [Google Scholar] [CrossRef]
- van Dop, W.A.; Heijmans, J.; Büller, N.V.J.A.; Snoek, S.A.; Rosekrans, S.L.; Wassenberg, E.A.; van den Bergh Weerman, M.A.; Lanske, B.; Clarke, A.R.; Winton, D.J.; et al. Loss of Indian Hedgehog Activates Multiple Aspects of a Wound Healing Response in the Mouse Intestine. Gastroenterology 2010, 139, 1665–1676.e10. [Google Scholar] [CrossRef]
- Vidal, M.T.A.; Lourenço, S.V.; Soares, F.A.; Gurgel, C.A.; Studart, E.J.B.; Valverde, L.D.F.; Araújo, I.B.D.O.; Ramos, E.A.G.; Xavier, F.C.D.A.; Dos Santos, J.N. The Sonic Hedgehog Signaling Pathway Contributes to the Development of Salivary Gland Neoplasms Regardless of Perineural Infiltration. Tumor Biol. 2016, 37, 9587–9601. [Google Scholar] [CrossRef]
- Cierpikowski, P.; Leszczyszyn, A.; Bar, J. The Role of Hedgehog Signaling Pathway in Head and Neck Squamous Cell Carcinoma. Cells 2023, 12, 2083. [Google Scholar] [CrossRef]
- Mao, L.; Xia, Y.; Zhou, Y.; Dai, R.; Yang, X.; Wang, Y.; Duan, S.; Qiao, X.; Mei, Y.; Hu, B. Activation of Sonic Hedgehog Signaling Pathway in Olfactory Neuroblastoma. Oncology 2009, 77, 231–243. [Google Scholar] [CrossRef]
- Varnat, F.; Duquet, A.; Malerba, M.; Zbinden, M.; Mas, C.; Gervaz, P.; Ruiz i Altaba, A. Human Colon Cancer Epithelial Cells Harbour Active HEDGEHOG-GLI Signalling That Is Essential for Tumour Growth, Recurrence, Metastasis and Stem Cell Survival and Expansion. EMBO Mol. Med. 2009, 1, 338–351. [Google Scholar] [CrossRef]
- Alinger, B.; Kiesslich, T.; Datz, C.; Aberger, F.; Strasser, F.; Berr, F.; Dietze, O.; Kaserer, K.; Hauser-Kronberger, C. Hedgehog Signaling Is Involved in Differentiation of Normal Colonic Tissue Rather than in Tumor Proliferation. Virchows Arch. 2009, 454, 369–379. [Google Scholar] [CrossRef]
- Das, D.; Fletcher, R.B.; Ngai, J. Cellular Mechanisms of Epithelial Stem Cell Self-Renewal and Differentiation during Homeostasis and Repair. WIREs Dev. Biol. 2020, 9, e361. [Google Scholar] [CrossRef]
- Bansal, R.; Nayak, B.B.; Bhardwaj, S.; Vanajakshi, C.N.; Das, P.; Somayaji, N.S.; Sharma, S. Cancer Stem Cells and Field Cancerization of Head and Neck Cancer—An Update. J. Fam. Med. Prim. Care 2020, 9, 3178–3182. [Google Scholar] [CrossRef]
- Mazumdar, T.; Devecchio, J.; Agyeman, A.; Shi, T.; Houghton, J.A. The GLI Genes as the Molecular Switch in Disrupting Hedgehog Signaling in Colon Cancer. Oncotarget 2011, 2, 638–645. [Google Scholar] [CrossRef]
- Gerling, M.; Büller, N.V.J.A.; Kirn, L.M.; Joost, S.; Frings, O.; Englert, B.; Bergström, Å.; Kuiper, R.V.; Blaas, L.; Wielenga, M.C.B.; et al. Stromal Hedgehog Signalling Is Downregulated in Colon Cancer and Its Restoration Restrains Tumour Growth. Nat. Commun. 2016, 7, 12321. [Google Scholar] [CrossRef]
- Guimaraes, V.S.N.; Vidal, M.T.A.; de Faro Valverde, L.; de Oliveira, M.G.; de Oliveira Siquara da Rocha, L.; Coelho, P.L.C.; Soares, F.A.; de Freitas Souza, B.S.; Bezerra, D.P.; Coletta, R.D.; et al. Hedgehog Pathway Activation in Oral Squamous Cell Carcinoma: Cancer-Associated Fibroblasts Exhibit Nuclear GLI-1 Localization. J. Mol. Histol. 2020, 51, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Noman, A.S.M.; Parag, R.R.; Rashid, M.I.; Rahman, M.Z.; Chowdhury, A.A.; Sultana, A.; Jerin, C.; Siddiqua, A.; Rahman, L.; Shirin, A.; et al. Widespread Expression of Sonic Hedgehog (Shh) and Nrf2 in Patients Treated with Cisplatin Predicts Outcome in Resected Tumors and Are Potential Therapeutic Targets for HPV-Negative Head and Neck Cancer. Ther. Adv. Med. Oncol. 2020, 12, 1758835920911229. [Google Scholar] [CrossRef]
- Liu, J.; Xie, Y.; Wu, S.; Lv, D.; Wei, X.; Chen, F.; Wang, Z. Combined Effects of EGFR and Hedgehog Signaling Blockade on Inhibition of Head and Neck Squamous Cell Carcinoma. Int. J. Clin. Exp. Pathol. 2017, 10, 9816–9828. [Google Scholar] [PubMed]
- Zubčić, V.; Rinčić, N.; Kurtović, M.; Trnski, D.; Musani, V.; Ozretić, P.; Levanat, S.; Leović, D.; Sabol, M. GANT61 and Lithium Chloride Inhibit the Growth of Head and Neck Cancer Cell Lines Through the Regulation of GLI3 Processing by GSK3β. Int. J. Mol. Sci. 2020, 21, 6410. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Tian, L.; Cheng, J.; Chen, Z.; Xu, B.; Wang, L.; Li, C.; Huang, Q. Sonic Hedgehog Signaling Pathway Supports Cancer Cell Growth during Cancer Radiotherapy. PLoS ONE 2013, 8, e65032. [Google Scholar] [CrossRef]
- Hehlgans, S.; Booms, P.; Güllülü, Ö.; Sader, R.; Rödel, C.; Balermpas, P.; Rödel, F.; Ghanaati, S. Radiation Sensitization of Basal Cell and Head and Neck Squamous Cell Carcinoma by the Hedgehog Pathway Inhibitor Vismodegib. Int. J. Mol. Sci. 2018, 19, 2485. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhang, L.; Hai, B.; Wang, J.; Baetge, C.L.; Deveau, M.A.; Kapler, G.M.; Feng, J.Q.; Liu, F. Transient Activation of the Hedgehog-Gli Pathway Rescues Radiotherapy-Induced Dry Mouth via Recovering Salivary Gland Resident Macrophages. Cancer Res. 2020, 80, 5531–5542. [Google Scholar] [CrossRef]
- Hu, L.; Du, C.; Yang, Z.; Yang, Y.; Zhu, Z.; Shan, Z.; Zhang, C.; Wang, S.; Liu, F. Transient Activation of Hedgehog Signaling Inhibits Cellular Senescence and Inflammation in Radiated Swine Salivary Glands through Preserving Resident Macrophages. Int. J. Mol. Sci. 2021, 22, 13493. [Google Scholar] [CrossRef] [PubMed]
- Henkin, R.I.; Hosein, S.; Stateman, W.A.; Knoppel, A.B.. Sonic Hedgehog in Nasal Mucus Is a Biomarker for Smell Loss in Patients with Hyposmia. Cell. Mol. Med. Open Access 2016, 2, 9. [Google Scholar] [CrossRef]
- Henkin, R.I.; Knöppel, A.B.; Abdelmeguid, M.; Stateman, W.A.; Hosein, S. Sonic Hedgehog Is Present in Parotid Saliva and Is Decreased in Patients with Taste Dysfunction. J. Oral Pathol. Med. 2017, 46, 829–833. [Google Scholar] [CrossRef]
- Maurya, D.K.; Bohm, S.; Alenius, M. Hedgehog Signaling Regulates Ciliary Localization of Mouse Odorant Receptors. Proc. Natl. Acad. Sci. USA 2017, 114, E9386–E9394. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Tumor Regions | Total n | All | Colon (n = 6) | Intestinal (n = 18) | Non-Intestinal (n = 3) | Salivary (n = 3) |
| GLI1 | Tumor | 30 | 5 (16.7%) | 0 (0%) | 4 (22.2%) | 1 (33.3%) | 0 (0%) |
| Tumor stroma | 30 | 5 (16.7%) | 0 (0%) | 4 (22.2%) | 1 (33.3%) | 0 (0%) | |
| GLI2 | Tumor | 30 | 27 (90%) | 5 (83.3%) | 16 (88.9%) | 3 (100%) | 3 (100%) |
| Tumor stroma | 30 | 20 (66.7%) | 4 (66.7%) | 14 (77.8%) | 1 (33.3%) | 1 (33.3%) | |
| GLI3 | Tumor | 30 | 1 (3.3%) | 0 (0%) | 1 (5.5%) | 0 (0%) | 0 (0%) |
| Tumor stroma | 30 | 1 (3.3%) | 0 (0%) | 0 (0%) | 0 (0%) | 1 (33.3%) | |
| PTCH1 | Tumor | 30 | 16 (53.3%) | 4 (66.7%) | 8 (44.4%) | 2 (66.7%) | 2 (66.7%) |
| Tumor stroma | 30 | 23 (76.7%) | 5 (83.3%) | 14 (77.8%) | 2 (66.7%) | 2 (66.7%) | |
| SHH | Tumor | 30 | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Tumor stroma | 30 | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | |
| IHH | Tumor | 30 | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Tumor stroma | 30 | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | |
| Healthy Regions | Total n | All | Colon (n = 6) | Intestinal (n = 9) | Non-Intestinal (n = 1) | Salivary (n = 2) | |
| GLI1 | Healthy epithelium | 18 | 1 (5.5%) | 0 (0%) | 0 (0%) | 0 (0%) | 1 (50%) |
| Healthy stroma | 18 | 2 (11.4%) | 0 (0%) | 2 (22.2%) | 0 (0%) | 0 (0%) | |
| GLI2 | Healthy epithelium | 18 | 15 (83.3%) | 6 (100%) | 7 (77.8%) | 1 (100%) | 1 (50%) |
| Healthy stroma | 18 | 15 (83.3%) | 6 (100%) | 7 (77.8%) | 1 (100%) | 1 (50%) | |
| GLI3 | Healthy epithelium | 18 | 1 (5.5%) | 0 (0%) | 0 (0%) | 0 (0%) | 1 (50%) |
| Healthy stroma | 18 | 1 (5.5%) | 0 (0%) | 1 (11.1%) | 0 (0%) | 0 (0%) | |
| PTCH1 | Healthy epithelium | 18 | 6 (33.3%) | 2 (33.3%) | 2 (22.2%) | 0 (0%) | 2 (100%) |
| Healthy stroma | 18 | 16 (88.9%) | 5 (83.3%) | 9 (100%) | 1 (100%) | 1 (50%) | |
| SHH | Healthy epithelium | 18 | 3 (16.7%) | 0 (0%) | 2 (22.2%) | 0 (0%) | 1 (50%) |
| Healthy stroma | 18 | 3 (16.7%) | 0 (0%) | 2 (22.2%) | 0 (0%) | 1 (50%) | |
| IHH | Healthy epithelium | 18 | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Healthy stroma | 18 | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leović, M.; Jakovčević, A.; Mumlek, I.; Zagorac, I.; Sabol, M.; Leović, D. A Pilot Immunohistochemical Study Identifies Hedgehog Pathway Expression in Sinonasal Adenocarcinoma. Int. J. Mol. Sci. 2024, 25, 4630. https://doi.org/10.3390/ijms25094630
Leović M, Jakovčević A, Mumlek I, Zagorac I, Sabol M, Leović D. A Pilot Immunohistochemical Study Identifies Hedgehog Pathway Expression in Sinonasal Adenocarcinoma. International Journal of Molecular Sciences. 2024; 25(9):4630. https://doi.org/10.3390/ijms25094630
Chicago/Turabian StyleLeović, Matko, Antonija Jakovčević, Ivan Mumlek, Irena Zagorac, Maja Sabol, and Dinko Leović. 2024. "A Pilot Immunohistochemical Study Identifies Hedgehog Pathway Expression in Sinonasal Adenocarcinoma" International Journal of Molecular Sciences 25, no. 9: 4630. https://doi.org/10.3390/ijms25094630