
Estrogen-Related Receptor α: A Key Transcription Factor in the Regulation of Energy Metabolism at an Organismic Level and a Target of the ABA/LANCL Hormone Receptor System
 , , , and
, , , and 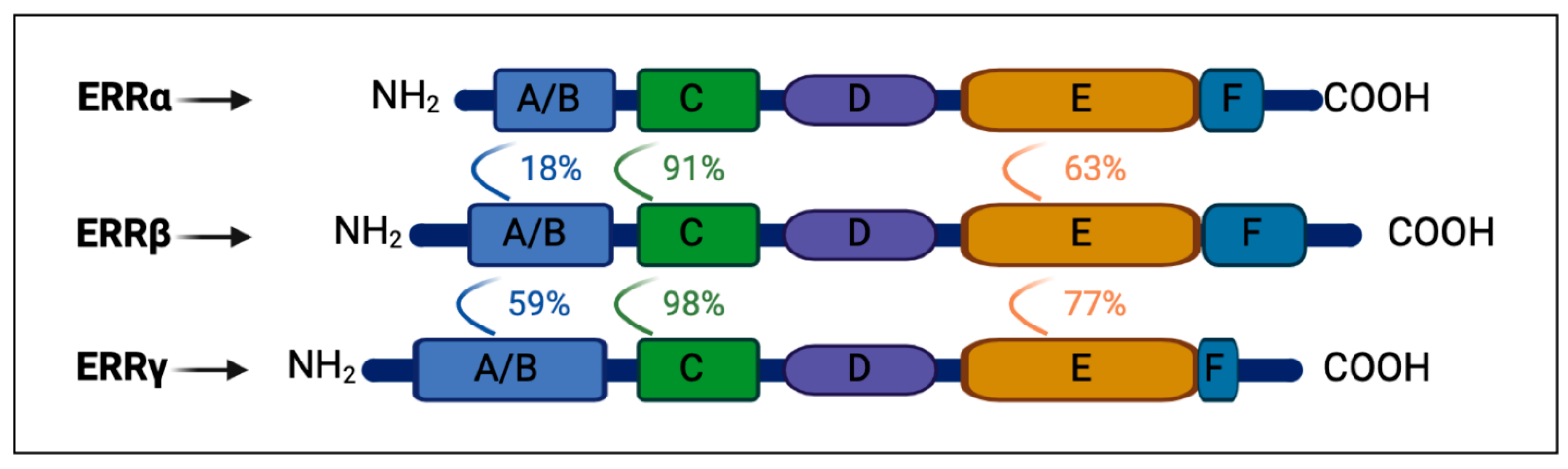{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. ERRs: Protein Family and Tissue Expression
1.2. ERRα: Master Regulator of Mitochondrial Biogenesis, Oxidative Phosphorylation, and Energy Production
1.3. Functional Collaboration between PGC-1α and ERRα: The PGC-1α/ERRα Axis
1.4. AMPK and SIRT1 Control the ERRα/PGC-1α Axis
1.5. ERRs and Protection from Oxidative Stress, the Yang of Oxphos
1.6. The ABA-Sensing System Is an Ancient Stress-Sensing and -Responding Hormone/Receptors System
2. Cell- and Organ-Specific Functions of ERRα
2.1. Heart
2.2. Skeletal Muscle
2.3. Kidney
2.4. Brain
2.5. Brown Adipose Tissue (BAT)
2.6. ERRα in Tumors
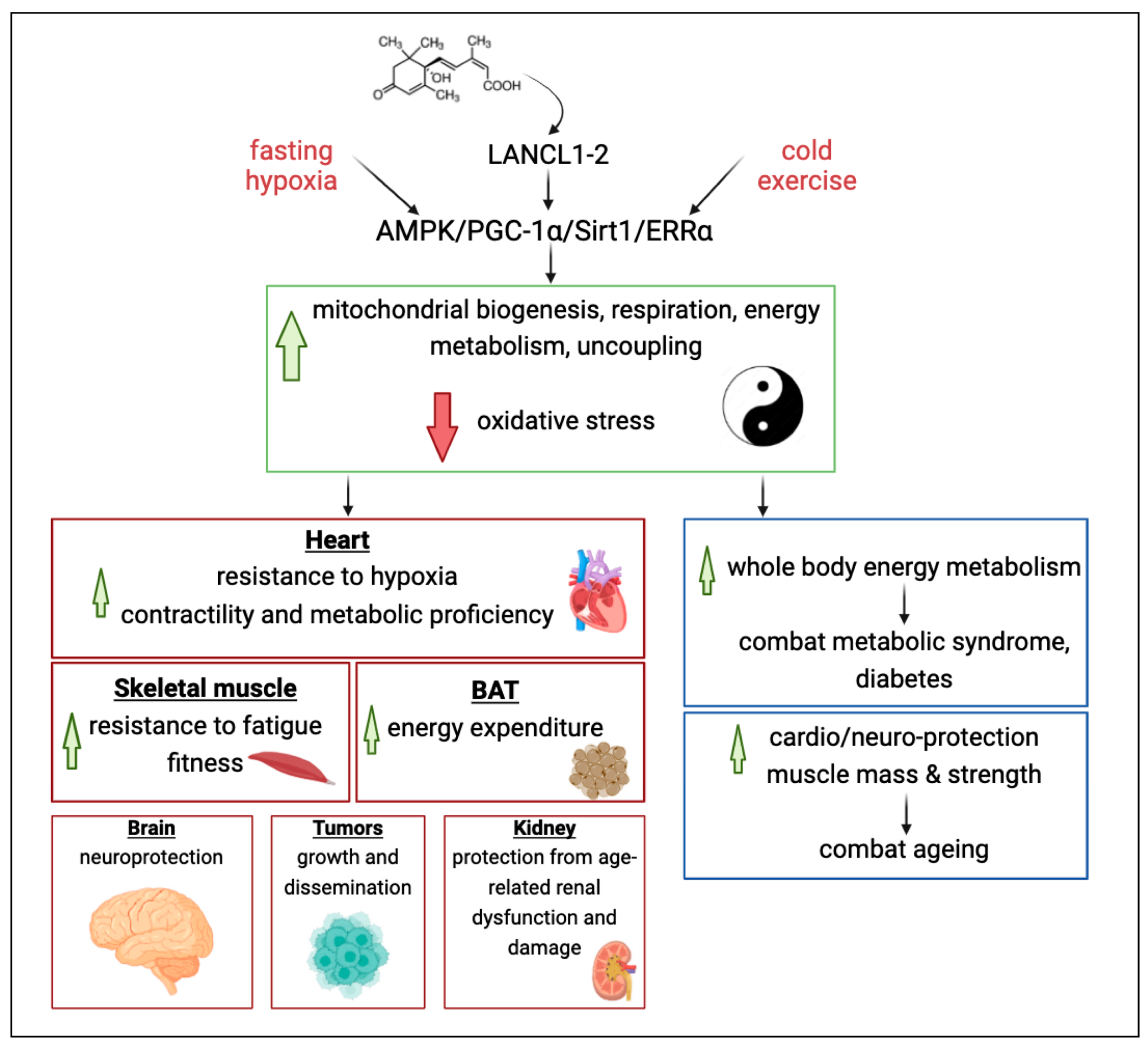
3. The ABA/LANCL1-2 Hormone/Receptor System Controls the PGC-1α/ERRα Axis
3.1. Cardiomyocytes
3.2. Adipocytes
4. Conclusions and Future Perspectives
4.1. Conclusions
- Enhancing ERRα activity promotes mitochondrial health and protects against oxidative stress in several aerobic organs and tissues, particularly in the heart, skeletal muscle, brain, and kidney.
- Repressing ERRα activity is a means to undermine mitochondrial energy metabolism (supporting cell duplication) and cytoskeletal dynamics (allowing cell migration) in cancer cells.
- The ABA/LANCL1-2 hormone/receptor system emerges as a new controller of ERRα expression levels and transcriptional activity via the AMPK/SIRT1/PGC-1α axis.
- A reciprocal feed-forward transcriptional relationship exists between the LANCL proteins and transcriptional coactivators ERRα/PGC-1α, which could be exploited with natural or synthetic LANCL agonists [118] to improve mitochondrial function in multiple clinical settings.
4.2. Future Perspectives
4.2.1. Increase Muscle and Adipocyte Energy Expenditure
4.2.2. Reduce Oxidative Stress
4.2.3. Improve Cardiomyocyte Function
4.2.4. Improve Skeletal Muscle Fitness
4.2.5. Improve Whole-Body Glucose Disposal to Combat Diabetes
4.2.6. Explore the Neuroprotective Role of the LANCL/ERRα System in the Brain
4.2.7. Explore the Nature of the Molecular Interaction between the ABA/LANCL System and ERRα
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Giguère, V.; Yang, N.; Segui, P.; Evans, R.M. Identification of a new class of steroid hormone receptors. Nature 1988, 331, 91–94. [Google Scholar] [CrossRef]
- Eudy, J.D.; Yao, S.; Weston, M.D.; Ma-Edmonds, M.; Talmadge, C.B.; Cheng, J.J.; Kimberling, W.J.; Sumegi, J. Isolation of a gene encoding a novel member of the nuclear receptor superfamily from the critical region of Usher syndrome type IIa at 1q41. Genomics 1998, 50, 382–384. [Google Scholar] [CrossRef]
- Divekar, S.D.; Tiek, D.M.; Fernandez, A.; Riggins, R.B. Estrogen-related receptor beta (ERRbeta)—Renaissance receptor or receptor renaissance? Nucl. Recept. Signal 2016, 14, e002. [Google Scholar] [CrossRef]
- Huss, J.M.; Garbacz, W.G.; Xie, W. Constitutive activities of estrogen-related receptors: Transcriptional regulation of metabolism by the ERR pathways in health and disease. Biochim. Biophys. Acta 2015, 1852, 1912–1927. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, M.; Yen, P.M.; Singh, B.K. Estrogen-related receptor alpha: An under-appreciated potential target for the treatment of metabolic diseases. Int. J. Mol. Sci. 2020, 21, 1645. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Liu, J.; Gu, L.; Ma, X.; Huang, B.; Pan, X. Research progress on the reproductive and non-reproductive endocrine tumors by estrogen-related receptors. J. Steroid Biochem. Mol. Biol. 2016, 158, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Heckler, M.M.; Riggins, R.B. ERRbeta splice variants differentially regulate cell cycle progression. Cell Cycle 2015, 14, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Bombail, V.; Collins, F.; Brown, P.; Saunders, P.T. Modulation of ER alpha transcriptional activity by the orphan nuclear receptor ERR beta and evidence for differential effects of long- and short-form splice variants. Mol. Cell Endocrinol. 2010, 314, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Bielli, P.; Pagliarini, V.; Pieraccioli, M.; Caggiano, C.; Sette, C. Splicing dysregulation as oncogenic driver and passenger factor in brain tumors. Cells 2019, 9, 10. [Google Scholar] [CrossRef]
- Lü, Z.; Zhu, K.; Pang, Z.; Liu, L.; Jiang, L.; Liu, B.; Shi, H.; Ping, H.; Chi, C.; Gong, L. Identification, characterization and mRNA transcript abundance profiles of estrogen related receptor (ERR) in Sepiella japonica imply its possible involvement in female reproduction. Anim. Reprod. Sci. 2019, 211, 106231. [Google Scholar] [CrossRef]
- Vu, E.H.; Kraus, R.J.; Mertz, J.E. Phosphorylation-dependent sumoylation of estrogen-related receptor alpha1. Biochemistry 2007, 46, 9795–9804. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, A.M.; Wilson, B.J.; Yang, X.J.; Giguère, V. Phosphorylation-dependent sumoylation regulates estrogen-related receptor-alpha and -gamma transcriptional activity through a synergy control motif. Mol. Endocrinol. 2008, 22, 570–584. [Google Scholar] [CrossRef]
- Xia, H.; Dufour, C.R.; Giguère, V. ERRα as a bridge between transcription and function: Role in liver metabolism and disease. Front. Endocrinol. 2019, 10, 206. [Google Scholar] [CrossRef]
- Mohideen-Abdul, K.; Tazibt, K.; Bourguet, M.; Hazemann, I.; Lebars, I.; Takacs, M.; Cianférani, S.; Klaholz, B.P.; Moras, D.; Billas, I.M.L. Importance of the sequence-directed DNA shape for specific binding site recognition by the estrogen-related receptor. Front. Endocrinol. 2017, 8, 140. [Google Scholar] [CrossRef] [PubMed]
- Horard, B.; Castet, A.; Bardet, P.L.; Laudet, V.; Cavailles, V.; Vanacker, J.M. Dimerization is required for transactivation by estrogen-receptor-related (ERR) orphan receptors: Evidence from amphioxus ERR. J. Mol. Endocrinol. 2004, 33, 493–509. [Google Scholar] [CrossRef] [PubMed]
- Casaburi, I.; Chimento, A.; De Luca, A.; Nocito, M.; Sculco, S.; Avena, P.; Trotta, F.; Rago, V.; Sirianni, R.; Pezzi, V. Cholesterol as an endogenous ERRalpha agonist: A new perspective to cancer treatment. Front. Endocrinol. 2018, 9, 525. [Google Scholar] [CrossRef] [PubMed]
- Misra, J.; Kim, D.K.; Choi, H.S. ERRγ: A junior orphan with a senior role in metabolism. Trends Endocrinol. Metab. 2017, 28, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Helsen, C.; Claessens, F. Looking at nuclear receptors from a new angle. Mol. Cell. Endocrinol. 2014, 382, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Gibson, D.A.; Saunders, P.T. Estrogen dependent signaling in reproductive tissues—A role for estrogen receptors and estrogen related receptors. Mol. Cell. Endocrinol. 2012, 348, 361–372. [Google Scholar] [CrossRef]
- Misawa, A.; Inoue, S. Estrogen-related receptors in breast cancer and prostate cancer. Front. Endocrinol. 2015, 6, 83. [Google Scholar] [CrossRef]
- Liu, X.; Nishimura, H.; Fujiyama, A.; Matsushima, A.; Shimohigashi, M.; Shimohigashi, Y. α-Helix-peptides comprising the human nuclear receptor ERRγ competitively provoke inhibition of functional homomeric dimerization. Biopolymers 2016, 106, 547–554. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, D.; Yang, C.; Sherman, M. Molecular basis for the constitutive activity of estrogen-related receptor alpha-1. J. Biol. Chem. 2001, 276, 28465–28470. [Google Scholar] [CrossRef]
- Li, D.; Cai, Y.; Teng, D.; Li, W.; Tang, Y.; Liu, G. Computational insights into the interaction mechanisms of estrogen-related receptor alpha with endogenous ligand cholesterol. Chem. Biol. Drug Des. 2019, 94, 1316–1329. [Google Scholar] [CrossRef] [PubMed]
- Ghanbari, F.; Mader, S.; Philip, A. Cholesterol as an Endogenous Ligand of ERRalpha Promotes ERRalpha-Mediated Cellular Proliferation and Metabolic Target Gene Expression in Breast Cancer Cells. Cells 2020, 9, 1765. [Google Scholar] [CrossRef] [PubMed]
- Villena, J.A.; Kralli, A. ERRalpha: A metabolic function for the oldest orphan. Trends Endocrinol. Metab. 2008, 19, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Thouennon, E.; Delfosse, V.; Bailly, R.; Blanc, P.; Boulahtouf, A.; Grimaldi, M.; Barducci, A.; Bourguet, W.; Balaguer, P. Insights into the activation mechanism of human estrogen-related receptor gamma by environmental endocrine disruptors. Cell. Mol. Life Sci. 2019, 76, 4769–4781. [Google Scholar] [CrossRef] [PubMed]
- Misra, J.; Kim, D.K.; Jung, Y.S.; Kim, H.B.; Kim, Y.H.; Yoo, E.K.; Kim, B.G.; Kim, S.; Lee, I.K.; Harris, R.A.; et al. O-GlcNAcylation of orphan nuclear receptor estrogen-related receptor γ promotes hepatic gluconeogenesis. Diabetes 2016, 65, 2835–2848. [Google Scholar] [CrossRef]
- Zhang, L.D.; Chen, L.; Zhang, M.; Qi, H.J.; Chen, L.; Chen, H.F.; Zhong, M.K.; Shi, X.J.; Li, Q.Y. Downregulation of ERRalpha inhibits angiogenesis in human umbilical vein endothelial cells through regulating VEGF production and PI3K/Akt/STAT3 signaling pathway. Eur. J. Pharmacol. 2015, 769, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Festuccia, N.; Owens, N.; Navarro, P. Esrrb, an estrogen-related receptor involved in early development, pluripotency, and reprogramming. FEBS Lett. 2018, 592, 852–877. [Google Scholar] [CrossRef]
- Likhite, N.; Yadav, V.; Milliman, E.J.; Sopariwala, D.H.; Lorca, S.; Narayana, N.P.; Sheth, M.; Reineke, E.L.; Giguère, V.; Narkar, V. Loss of estrogen-related receptor alpha facilitates angiogenesis in endothelial cells. Mol. Cell. Biol. 2019, 39, e00411-18. [Google Scholar] [CrossRef]
- Schreiber, S.N.; Knutti, D.; Brogli, K.; Uhlmann, T.; Kralli, A. The transcriptional coactivator PGC-1 regulates the expression and activity of the orphan nuclear receptor estrogen-related receptor alpha (ERRalpha). J. Biol. Chem. 2003, 278, 9013–9018. [Google Scholar] [CrossRef] [PubMed]
- Cerutti, C.; Shi, J.R.; Vanacker, J.M. Multifaceted Transcriptional Network of Estrogen-Related Receptor Alpha in Health and Disease. Int. J. Mol. Sci. 2023, 24, 4265. [Google Scholar] [CrossRef]
- Huss, J.M.; Torra, I.P.; Staels, B.; Giguère, V.; Kelly, D.P. Estrogen-related receptor alpha directs peroxisome proliferator-activated receptor alpha signaling in the transcriptional control of energy metabolism in cardiac and skeletal muscle. Mol. Cell. Biol. 2004, 24, 9079–9091. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Evans, R. PPARs and ERRs: Molecular mediators of mitochondrial metabolism. Curr. Opin. Cell Biol. 2015, 33, 49–54. [Google Scholar] [CrossRef]
- Kida, Y.S.; Kawamura, T.; Wei, Z.; Sogo, T.; Jacinto, S.; Shigeno, A.; Kushige, H.; Yoshihara, E.; Liddle, C.; Ecker, J.R.; et al. ERRs mediate a metabolic switch required for somatic cell reprogramming to pluripotency. Cell Stem Cell 2015, 16, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.L.; Ueno, M.; de Souza, C.T.; Pereira-da-Silva, M.; Gasparetti, A.L.; Bezzera, R.M.N.; Alberici, L.C.; Vercesi, A.E.; Saad, M.J.A.; Velloso, L.A. Cold-induced PGC-1alpha expression modulates muscle glucose uptake through an insulin receptor/Akt-independent, AMPK-dependent pathway. Am. J. Physiol. Endocrinol. Metab. 2004, 287, E686–E695. [Google Scholar] [CrossRef] [PubMed]
- Puigserver, P.; Wu, Z.; Park, C.W.; Graves, R.; Wright, M.; Spiegelman, B.M. A cold-inducible coactivator of nuclear receptors linked to adaptive thermogenesis. Cell 1998, 92, 829–839. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.C.; Puigserver, P.; Chen, G.; Donovan, J.; Wu, Z.; Rhee, J.; Adelmant, G.; Stafford, J.; Kahn, C.R.; Granner, D.K.; et al. Control of hepatic gluconeogenesis through the transcriptional coactivator PGC-1. Nature 2001, 413, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Lehman, J.J.; Barger, P.M.; Kovacs, A.; Saffitz, J.E.; Medeiros, D.M.; Kelly, D.P. Peroxisome proliferator–activated receptor γ coactivator-1 promotes cardiac mitochondrial biogenesis. J. Clin. Investg. 2000, 106, 847–856. [Google Scholar] [CrossRef]
- Goto, M.; Terada, S.; Kato, M.; Katoh, M.; Yokozeki, T.; Tabata, I.; Shimokawa, T. cDNA cloning and mRNA analysis of PGC-1 in epitrochlearis muscle in swimming-exercised rats. Biochem. Biophys. Res. Commun. 2000, 274, 350–354. [Google Scholar] [CrossRef]
- Herzig, S.; Long, F.; Jhala, U.S.; Hedrick, S.; Quinn, R.; Bauer, A.; Rudolph, D.; Schutz, G.; Yoon, C.; Puigserver, P.; et al. CREB regulates hepatic gluconeogenesis through the coactivator PGC-1. Nature 2001, 413, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Puigserver, P.; Andersson, U.; Zhang, C.; Adelmant, G.; Mootha, V.; Troy, A.; Cinti, S.; Lowell, B.; Scarpulla, R.C.; et al. Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell 1999, 98, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Vega, R.B.; Huss, J.M.; Kelly, D.P. The coactivator PGC-1 cooperates with peroxisome proliferator-activated receptor α in transcriptional control of nuclear genes encoding mitochondrial fatty acid oxidation enzymes. Mol. Cell. Biol. 2000, 20, 1868–1876. [Google Scholar] [CrossRef] [PubMed]
- Ichida, M.; Nemoto, S.; Finkel, T. Identification of a specific molecular repressor of the peroxisome proliferator-activated receptor gamma Coactivator-1 alpha (PGC-1alpha). J. Biol. Chem. 2002, 277, 50991–50995. [Google Scholar] [CrossRef] [PubMed]
- Laganière, J.; Tremblay, G.B.; Dufour, C.R.; Giroux, S.; Rousseau, F.; Giguère, V. A polymorphic autoregulatory hormone response element in the human estrogen-related receptor alpha (ERRalpha) promoter dictates peroxisome proliferator-activated receptor gamma coactivator-1alpha control of ERRalpha expression. J. Biol. Chem. 2004, 279, 18504–18510. [Google Scholar] [CrossRef]
- Ramjiawan, A.; Bagchi, R.A.; Albak, L.; Czubryt, M.P. Mechanism of cardiomyocyte PGC-1α gene regulation by ERRα. Biochem. Cell Biol. 2013, 91, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Cantó, C.; Auwerx, J. PGC-1alpha, SIRT1 and AMPK, an energy sensing network that controls energy expenditure. Curr. Opin. Lipidol. 2009, 20, 98–105. [Google Scholar] [CrossRef]
- Wilson, B.J.; Tremblay, A.M.; Deblois, G.; Sylvain-Drolet, G.; Giguère, V. An acetylation switch modulates the transcriptional activity of estrogen-related receptor alpha. Mol. Endocrinol. 2010, 24, 1349–1358. [Google Scholar] [CrossRef]
- Yan, M.; Audet-Walsh, É.; Manteghi, S.; Dufour, C.R.; Walker, B.; Baba, M.; St-Pierre, J.; Giguère, V.; Pause, A. Chronic AMPK activation via loss of FLCN induces functional beige adipose tissue through PGC-1α/ERRα. Genes. Dev. 2016, 30, 1034–1046. [Google Scholar] [CrossRef]
- Ma, L.; Wang, R.; Wang, H.; Zhang, Y.; Zhao, Z. Long-term caloric restriction activates the myocardial SIRT1/AMPK/PGC-1α pathway in C57BL/6J male mice. Food Nutr. Res. 2020, 64. [Google Scholar] [CrossRef]
- Guo, Z.; Wang, M.; Ying, X.; Yuan, J.; Wang, C.; Zhang, W.; Tian, S.; Yan, X. Caloric restriction increases the resistance of aged heart to myocardial ischemia/reperfusion injury via modulating AMPK-SIRT1-PGC1a energy metabolism pathway. Sci. Rep. 2023, 13, 2045. [Google Scholar] [CrossRef] [PubMed]
- Rasbach, K.A.; Gupta, R.K.; Ruas, J.L.; Wu, J.; Naseri, E.; Estall, J.L.; Spiegelman, B.M. PGC-1alpha regulates a HIF2alpha-dependent switch in skeletal muscle fiber types. Proc. Natl. Acad. Sci. USA 2010, 107, 21866–21871. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.M.; Guéant-Rodriguez, R.M.; Pooya, S.; Brachet, P.; Alberto, J.M.; Jeannesson, E.; Maskali, F.; Gueguen, N.; Marie, P.Y.; Lacolley, P.; et al. Methyl donor deficiency induces cardiomyopathy through altered methylation/acetylation of PGC-1α by PRMT1 and SIRT1. J. Pathol. 2011, 225, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Liu, Q.; Li, Y.; Tang, Q.; Wu, T.; Chen, L.; Pu, S.; Zhao, Y.; Zhang, G.; Huang, C.; et al. The diabetes medication canagliflozin promotes mitochondrial remodelling of adipocyte via the AMPK-Sirt1-Pgc-1α signalling pathway. Adipocyte 2020, 9, 484–494. [Google Scholar] [CrossRef] [PubMed]
- Vernier, M.; Dufour, C.R.; McGuirk, S.; Scholtes, C.; Li, X.; Bourmeau, G.; Kuasne, H.; Park, M.; St-Pierre, J.; Audet-Walsh, E.; et al. Estrogen-related receptors are targetable ROS sensors. Genes. Dev. 2020, 34, 544–559. [Google Scholar] [CrossRef] [PubMed]
- Van Waveren, C.; Moraes, C.T. Transcriptional co-expression and co-regulation of genes coding for components of the oxidative phosphorylation system. BMC Genom. 2008, 9, 18. [Google Scholar] [CrossRef] [PubMed]
- Chaltel-Lima, L.; Domínguez, F.; Domínguez-Ramírez, L.; Cortes-Hernandez, P. The Role of the Estrogen-Related Receptor Alpha (ERRa) in Hypoxia and Its Implications for Cancer Metabolism. Int. J. Mol. Sci. 2023, 24, 7983. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Wong, C. Suppressing the activity of ERRα in 3T3-L1 adipocytes reduces mitochondrial biogenesis but enhances glycolysis and basal glucose uptake. J. Cell. Mol. Med. 2009, 13, 3051–3060. [Google Scholar] [CrossRef] [PubMed]
- Tsushida, K.; Tanabe, K.; Masuda, K.; Tanimura, S.; Miyake, H.; Arata, Y.; Sugiyama, H.; Wada, J. Estrogen-related receptor α is essential for maintaining mitochondrial integrity in cisplatin-induced acute kidney injury. Biochem. Biophys. Res. Commun. 2018, 498, 918–924. [Google Scholar] [CrossRef]
- Waadt, R.; Seller, C.A.; Hsu, P.K.; Takahashi, Y.; Munemasa, S.; Schroeder, J.I. Plant hormone regulation of abiotic stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 680–694. [Google Scholar] [CrossRef]
- Liu, H.; Song, S.; Zhang, H.; Li, Y.; Niu, L.; Zhang, J.; Wang, W. Signaling Transduction of ABA, ROS, and Ca2+ in Plant Stomatal Closure in Response to Drought. Int. J. Mol. Sci. 2022, 23, 14824. [Google Scholar] [CrossRef] [PubMed]
- Zocchi, E.; Carpaneto, A.; Cerrano, C.; Bavestrello, G.; Giovine, M.; Bruzzone, S.; Guida, L.; Franco, L.; Usai, C. The temperature-signaling cascade in sponges involves a heat-gated cation channel, abscisic acid, and cyclic ADP-ribose. Proc. Natl. Acad. Sci. USA 2001, 98, 14859–14864. [Google Scholar] [CrossRef] [PubMed]
- Puce, S.; Basile, G.; Bavestrello, G.; Bruzzone, S.; Cerrano, C.; Giovine, M.; Arillo, A.; Zocchi, E. Abscisic acid signaling through cyclic ADP-ribose in hydroid regeneration. J. Biol. Chem. 2004, 279, 39783–39788. [Google Scholar] [CrossRef] [PubMed]
- Takezawa, D.; Komatsu, K.; Sakata, Y. ABA in bryophytes: How a universal growth regulator in life became a plant hormone? J. Plant Res. 2011, 124, 437–453. [Google Scholar] [CrossRef]
- Bruzzone, S.; Ameri, P.; Briatore, L.; Mannino, E.; Basile, G.; Andraghetti, G.; Grozio, A.; Magnone, M.; Guida, L.; Scarfì, S.; et al. The plant hormone abscisic acid increases in human plasma after hyperglycemia and stimulates glucose consumption by adipocytes and myoblasts. FASEB J. 2012, 26, 1251–1260. [Google Scholar] [CrossRef]
- Spinelli, S.; Begani, G.; Guida, L.; Magnone, M.; Galante, D.; D‘Arrigo, C.; Scotti, C.; Iamele, L.; De Jonge, H.; Zocchi, E.; et al. LANCL1 binds abscisic acid and stimulates glucose transport and mitochondrial respiration in muscle cells via the AMPK/PGC-1α/Sirt1 pathway. Mol. Metab. 2021, 53, 101263. [Google Scholar] [CrossRef]
- Magnone, M.; Emionite, L.; Guida, L.; Vigliarolo, T.; Sturla, L.; Spinelli, S.; Buschiazzo, A.; Marini, C.; Sambuceti, G.; De Flora, A.; et al. Insulin-independent stimulation of skeletal muscle glucose uptake by low-dose abscisic acid via AMPK activation. Sci. Rep. 2020, 10, 1454. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, S.; Cossu, V.; Passalacqua, M.; Hansen, J.B.; Guida, L.; Magnone, M.; Sambuceti, G.; Marini, C.; Sturla, L.; Zocchi, E. The ABA/LANCL1/2 Hormone/Receptor System Controls Adipocyte Browning and Energy Expenditure. Int. J. Mol. Sci. 2023, 24, 3489. [Google Scholar] [CrossRef]
- Sturla, L.; Fresia, C.; Guida, L.; Grozio, A.; Vigliarolo, T.; Mannino, E.; Millo, E.; Bagnasco, L.; Bruzzone, S.; De Flora, A.; et al. Binding of abscisic acid to human LANCL2. Biochem. Biophys. Res. Commun. 2011, 415, 390–395. [Google Scholar] [CrossRef]
- Spinelli, S.; Magnone, M.; Guida, L.; Sturla, L.; Zocchi, E. The ABA/LANCL Hormone/Receptor System in the Control of Glycemia, of Cardiomyocyte Energy Metabolism, and in Neuroprotection: A New Ally in the Treatment of Diabetes Mellitus? Int. J. Mol. Sci. 2023, 24, 1199. [Google Scholar] [CrossRef]
- Lai, K.Y.; Galan, S.R.G.; Zeng, Y.; Zhou, T.H.; He, C.; Raj, R.; Riedl, J.; Liu, S.; Chooi, K.P.; Garg, N.; et al. LanCLs Add Glutathione to Dehydroamino Acids Generated at Phosphorylated Sites in the Proteome. Cell 2021, 184, 2680–2695. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, S.; Guida, L.; Passalacqua, M.; Magnone, M.; Cossu, V.; Sambuceti, G.; Marini, C.; Sturla, L.; Zocchi, E. Abscisic Acid and Its Receptors LANCL1 and LANCL2 Control Cardiomyocyte Mitochondrial Function, Expression of Contractile, Cytoskeletal and Ion Channel Proteins and Cell Proliferation via ERRα. Antioxidants 2023, 12, 1692. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Matsuura, T.R.; Wan, S.; Ryba, D.M.; Kim, J.U.; Won, K.J.; Lai, L.; Petucci, C.; Petrenko, N.; Musunuru, K.; et al. A Critical Role for Estrogen-Related Receptor Signaling in Cardiac Maturation. Circ. Res. 2020, 126, 1685–1702. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Xu, H.; Yan, L.; Ye, L.; Zhang, X.; Tan, B.; Yi, Q.; Tian, J.; Zhu, J. PGC-1α promotes mitochondrial respiration and biogenesis during the differentiation of hiPSCs into cardiomyocytes. Genes. Dis. 2020, 8, 891–906. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Tachibana, S.; Lam, K.; Arita, Y.; Khosrowjerdi, S.; Zhang, O.; Liang, A.; Li, R.; Andreyev, A.; Murphy, A.N.; et al. Perm1 promotes cardiomyocyte mitochondrial biogenesis and protects against hypoxia/reoxygenation-induced damage in mice. J. Biol. Chem. 2021, 297, 100825. [Google Scholar]
- Oka, S.I.; Sreedevi, K.; Shankar, T.S.; Yedla, S.; Arowa, S.; James, A.; Stone, K.G.; Olmos, K.; Sabry, A.D.; Horiuchi, A.; et al. PERM1 regulates energy metabolism in the heart via ERRα/PGC-1α axis. Front. Cardiovasc. Med. 2022, 9, 1033457. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Billon, C.; Li, H.; Wilderman, A.; Qi, L.; Graves, A.; Rideb, J.R.D.C.; Zhao, Y.; Hayes, M.; Yu, K.; et al. Novel Pan-ERR Agonists Ameliorate Heart Failure Through Enhancing Cardiac Fatty Acid Metabolism and Mitochondrial Function. Circulation 2024, 149, 227–250. [Google Scholar] [CrossRef]
- Kan, J.; Hu, Y.; Ge, Y.; Zhang, W.; Lu, S.; Zhao, C.; Zhang, R.; Liu, Y. Declined expressions of vast mitochondria-related genes represented by CYCS and transcription factor ESRRA in skeletal muscle aging. Bioengineered 2021, 12, 3485–3502. [Google Scholar] [CrossRef]
- Wattez, J.S.; Eury, E.; Hazen, B.C.; Wade, A.; Chau, S.; Ou, S.C.; Russell, A.P.; Cho, Y.; Kralli, A. Loss of skeletal muscle estrogen-related receptors leads to severe exercise intolerance. Mol. Metab. 2023, 68, 101670. [Google Scholar] [CrossRef]
- Sopariwala, D.H.; Rios, A.S.; Pei, G.; Roy, A.; da Silva, M.T.; Nguyen, H.T.T.; Saley, A.; Van Drunen, R.; Kralli, A.; Mahan, K.; et al. Innately expressed estrogen-related receptors in the skeletal muscle are indispensable for exercise fitness. FASEB J. 2023, 37, e22727. [Google Scholar] [CrossRef]
- Narkar, V.A.; Fan, W.; Downes, M.; Yu, R.T.; Jonker, J.W.; Alaynick, W.A.; Banayo, E.; Karunasiri, M.S.; Lorca, S.; Evans, R.M. Exercise and PGC-1α-independent synchronization of type I muscle metabolism and vasculature by ERRγ. Cell Metab. 2011, 13, 283–293. [Google Scholar] [CrossRef]
- Jager, S.; Handschin, C.; St-Pierre, J.; Spiegelman, B.M. AMP-activated protein kinase (AMPK) action in skeletal muscle via direct phosphorylation of PGC-1alpha. Proc. Natl. Acad. Sci. USA 2007, 104, 12017–12022. [Google Scholar] [CrossRef]
- De Smalen, L.M.; Börsch, A.; Leuchtmann, A.B.; Gill, J.F.; Ritz, D.; Zavolan, M.; Handschin, C. Impaired age-associated mitochondrial translation is mitigated by exercise and PGC-1α. Proc. Natl. Acad. Sci. USA 2023, 120, e2302360120. [Google Scholar] [CrossRef]
- Ahn, J.; Son, H.J.; Seo, H.D.; Há, T.Y.; Ahn, J.; Lee, H.; Shin, S.H.; Jung, C.H.; Jang, Y.J. γ-Oryzanol Improves Exercise Endurance and Muscle Strength by Upregulating PPARδ and ERRγ Activity in Aged Mice. Mol. Nutr. Food Res. 2021, 65, e2000652. [Google Scholar] [CrossRef]
- Billon, C.; Sitaula, S.; Banerjee, S.; Welch, R.; Elgendy, B.; Hegazy, L.; Oh, T.G.; Kazantzis, M.; Chatterjee, A.; Chrivia, J.; et al. Synthetic ERRα/β/γ Agonist Induces an ERRα-Dependent Acute Aerobic Exercise Response and Enhances Exercise Capacity. ACS Chem. Biol. 2023, 18, 756–771. [Google Scholar] [CrossRef]
- Xia, H.; Scholtes, C.; Dufour, C.R.; Guluzian, C.; Giguère, V. ERRα fosters running endurance by driving myofiber aerobic transformation and fuel efficiency. Mol. Metab. 2023, 78, 101814. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.H.; Kim, E.N.; Kim, M.Y.; Chung, S.; Shin, S.J.; Kim, H.W.; Yang, C.W.; Kim, Y.S.; Chang, Y.S.; Park, C.W.; et al. Age-associated molecular changes in the kidney in aged mice. Oxid. Med. Cell. Longev. 2012, 2012, 171383. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.X.; Myakala, K.; Libby, A.E.; Krawczyk, E.; Panov, J.; Jones, B.A.; Bhasin, K.; Shults, N.; Qi, Y.; Krausz, K.W.; et al. Estrogen-Related Receptor Agonism Reverses Mitochondrial Dysfunction and Inflammation in the Aging Kidney. Am. J. Pathol. 2023, 193, 1969–1987. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, P.; Park, J.; Del Pozo, C.H.; Li, L.; Doke, T.; Huang, S.; Zhao, J.; Kang, H.M.; Shrestra, R.; Balzer, M.S.; et al. The Nuclear Receptor ESRRA Protects from Kidney Disease by Coupling Metabolism and Differentiation. Cell Metab. 2021, 33, 379–394. [Google Scholar] [CrossRef]
- Nahar, L.; Delacroix, B.M.; Nam, H.W. The Role of Parvalbumin Interneurons in Neurotransmitter Balance and Neurological Disease. Front. Psychiatry 2021, 12, 679960. [Google Scholar] [CrossRef]
- McMeekin, L.J.; Joyce, K.L.; Jenkins, L.M.; Bohannon, B.M.; Patel, K.D.; Bohannon, A.S.; Patel, A.; Fox, S.N.; Simmons, M.S.; Day, J.J.; et al. Estrogen-related Receptor Alpha (ERRα) is Required for PGC-1α-dependent Gene Expression in the Mouse Brain. Neuroscience 2021, 479, 70–90. [Google Scholar] [CrossRef]
- Tang, Y.; Min, Z.; Xiang, X.J.; Liu, L.; Ma, Y.L.; Zhu, B.L.; Song, L.; Tang, J.; Deng, X.J.; Yan, Z.; et al. Estrogen-related receptor alpha is involved in Alzheimer’s disease-like pathology. Exp. Neurol. 2018, 305, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Lu, Y.; Khan, M.Z.; Anderson, R.M.; McDaniel, L.; Wilson, H.E.; Yin, T.C.; Radley, J.J.; Pieper, A.A.; Lutter, M. Behavioral disturbances in estrogen-related receptor alpha-null mice. Cell Rep. 2015, 11, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Ijichi, N.; Ikeda, K.; Horie-Inoue, K.; Yagi, K.; Okazaki, Y.; Inoue, S. Estrogen-related receptor alpha modulates the expression of adipogenesis-related genes during adipocyte differentiation. Biochem. Biophys. Res. Commun. 2007, 358, 813–818. [Google Scholar] [CrossRef] [PubMed]
- Kubo, M.; Ijichi, N.; Ikeda, K.; Horie-Inoue, K.; Takeda, S.; Inoue, S. Modulation of adipogenesis-related gene expression by estrogen-related receptor gamma during adipocytic differentiation. Biochim. Biophys. Acta 2009, 1789, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Gantner, M.L.; Hazen, B.C.; Eury, E.; Brown, E.L.; Kralli, A. Complementary Roles of Estrogen-Related Receptors in Brown Adipocyte Thermogenic Function. Endocrinology 2016, 157, 4770–4781. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.L.; Hazen, B.C.; Eury, E.; Wattez, J.S.; Gantner, M.L.; Albert, V.; Chau, S.; Sanchez-Alavez, M.; Conti, B.; Kralli, A. Estrogen-Related Receptors Mediate the Adaptive Response of Brown Adipose Tissue to Adrenergic Stimulation. iScience 2018, 2, 221–237. [Google Scholar] [CrossRef] [PubMed]
- Morganstein, D.L.; Wu, P.; Mane, M.R.; Fisk, N.M.; White, R.; Parker, M.G. Human fetal mesenchymal stem cells differentiate into brown and white adipocytes: A role for ERRalpha in human UCP1 expression. Cell Res. 2010, 20, 434–444. [Google Scholar] [CrossRef] [PubMed]
- Emmett, M.J.; Lim, H.W.; Jager, J.; Richter, H.J.; Adlanmerini, M.; Peed, L.C.; Briggs, E.R.; Steger, D.J.; Ma, T.; Sims, C.A.; et al. Histone deacetylase 3 prepares brown adipose tissue for acute thermogenic challenge. Nature 2017, 546, 544–548. [Google Scholar] [CrossRef]
- Bartelt, A.; Heeren, J. Adipose tissue browning and metabolic health. Nat. Rev. Endocrinol. 2014, 10, 24–36. [Google Scholar] [CrossRef]
- Peng, W.; Mu, Y.; Hu, Y.; Li, B.; Raman, J.; Sui, Z. Double Burden of Malnutrition in the Asia-Pacific Region—A Systematic Review and Meta-analysis. J. Epidemiol. Glob. Health 2020, 10, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.; He, P.; Xu, C.Y.; Hou, B.Y.; Qiang, G.F.; Du, G.H. Recent developments in natural products for white adipose tissue browning. Chin. J. Nat. Med. 2020, 18, 803–817. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Yu, L. Natural Bioactive Compounds as Potential Browning Agents in White Adipose Tissue. Pharm. Res. 2021, 38, 549–567. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Choi, J.Y.; Ryu, R.; Lee, J.; Cho, S.J.; Kwon, E.Y.; Lee, M.K.; Liu, K.H.; Rina, Y.; Sung, M.K.; et al. Platycodon grandiflorus Root Extract Attenuates Body Fat Mass, Hepatic Steatosis and Insulin Resistance through the Interplay between the Liver and Adipose Tissue. Nutrients 2016, 8, 532. [Google Scholar] [CrossRef] [PubMed]
- Lugano, R.; Ramachandran, M.; Dimberg, A. Tumor angiogenesis: Causes, consequences, challenges and opportunities. Cell. Mol. Life Sci. 2020, 77, 1745–1770. [Google Scholar] [CrossRef] [PubMed]
- Seo, W.; Yoo, S.; Zhong, Y.; Lee, S.H.; Woo, S.Y.; Choi, H.S.; Won, M.; Roh, T.; Jeon, S.M.; Kim, K.T.; et al. Targeting ERRα promotes cytotoxic effects against acute myeloid leukemia through suppressing mitochondrial oxidative phosphorylation. J. Hematol. Oncol. 2022, 15, 156. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, Y.; Chen, L.; Chen, H.; Qi, H.; Zheng, Y.; Du, Y.; Zhang, L.; Wang, T.; Li, Q. Dihydroartemisinin suppresses glioma growth by repressing ERRα-mediated mitochondrial biogenesis. Mol. Cell. Biochem. 2023, 1–17. [Google Scholar] [CrossRef]
- Park, S.; Safi, R.; Liu, X.; Baldi, R.; Liu, W.; Liu, J.; Locasale, J.W.; Chang, C.Y.; McDonnell, D.P. Inhibition of ERRα Prevents Mitochondrial Pyruvate Uptake Exposing NADPH-Generating Pathways as Targetable Vulnerabilities in Breast Cancer. Cell Rep. 2019, 27, 3587–3601. [Google Scholar] [CrossRef] [PubMed]
- Dings, M.P.G.; van der Zalm, A.P.; Bootsma, S.; van Maanen, T.F.J.; Waasdorp, C.; van den Ende, T.; Liu, D.; Bailey, P.; Koster, J.; Zwijnenburg, D.A.; et al. Estrogen-related receptor alpha drives mitochondrial biogenesis and resistance to neoadjuvant chemoradiation in esophageal cancer. Cell Rep. Med. 2022, 3, 100802. [Google Scholar] [CrossRef]
- Li, X.; Zhang, K.; Hu, Y.; Luo, N. ERRα activates SHMT2 transcription to enhance the resistance of breast cancer to lapatinib via modulating the mitochondrial metabolic adaption. Biosci. Rep. 2020, 40, BSR20192465. [Google Scholar] [CrossRef]
- Fradet, A.; Sorel, H.; Bouazza, L.; Goehrig, D.; Dépalle, B.; Bellahcène, A.; Castronovo, V.; Follet, H.; Descotes, F.; Aubin, J.E.; et al. Dual function of ERRα in breast cancer and bone metastasis formation: Implication of VEGF and osteoprotegerin. Cancer Res. 2011, 71, 5728–5738. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Chang, C.Y.; Safi, R.; Liu, X.; Baldi, R.; Jasper, J.S.; Anderson, G.R.; Liu, T.; Rathmell, J.C.; Dewhirst, M.W.; et al. ERRalpha-regulated lactate metabolism contributes to resistance to targeted therapies in breast cancer. Cell Rep. 2016, 15, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Guo, J.; Zhang, H.; Meng, Q.; Ma, Y.; Lin, R.; Yi, X.; Lu, H.; Bai, X.; Cheng, J. The enhanced expression of estrogen-related receptor α in human bladder cancer tissues and the effects of estrogen-related receptor α knockdown on bladder cancer cells. J. Cell. Biochem. 2019, 120, 13841–13852. [Google Scholar] [CrossRef] [PubMed]
- Tribollet, V.; Cerutti, C.; Géloën, A.; Berger, E.; De Mets, R.; Balland, M.; Courchet, J.; Vanacker, J.M.; Forcet, C. ERRα coordinates actin and focal adhesion dynamics. Cancer Gene Ther. 2022, 29, 1429–1438. [Google Scholar] [CrossRef] [PubMed]
- Scialò, F.; Fernández-Ayala, D.J.; Sanz, A. Role of Mitochondrial Reverse Electron Transport in ROS Signaling: Potential Roles in Health and Disease. Front. Physiol. 2017, 8, 428. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Camacho, I.; Flores-Herrera, O.; Zazueta, C. The relevance of the supramolecular arrangements of the respiratory chain complexes in human diseases and aging. Mitochondrion 2019, 47, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Ramzan, R.; Vogt, S.; Kadenbach, B. Stress-mediated generation of deleterious ROS in healthy individuals-role of cytochrome c oxidase. J. Mol. Med. 2020, 98, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Scarano, N.; Di Palma, F.; Origlia, N.; Musumeci, F.; Schenone, S.; Spinelli, S.; Passalacqua, M.; Zocchi, E.; Sturla, L.; Cichero, E.; et al. New Insights into the LANCL2-ABA Binding Mode towards the Evaluation of New LANCL Agonists. Pharmaceutics 2023, 15, 2754. [Google Scholar] [CrossRef] [PubMed]
- Magnone, M.; Sturla, L.; Guida, L.; Spinelli, S.; Begani, G.; Bruzzone, S.; Fresia, C.; Zocchi, E. Abscisic Acid: A Conserved Hormone in Plants and Humans and a Promising Aid to Combat Prediabetes and the Metabolic Syndrome. Nutrients 2020, 12, 1724. [Google Scholar] [CrossRef]
- Rangwala, S.M.; Li, X.; Lindsley, L.; Wang, X.; Shaughnessy, S.; Daniels, T.G.; Szustakowski, J.; Nirmala, N.R.; Wu, Z.; Stevenson, S.C. Estrogen-related receptor alpha is essential for the expression of antioxidant protection genes and mitochondrial function. Biochem. Biophys. Res. Commun. 2007, 357, 231–236. [Google Scholar] [CrossRef]
- Kim, M.B.; Kim, C.; Hwang, J.K. Standardized Siegesbeckia orientalis L. Extract Increases Exercise Endurance through Stimulation of Mitochondrial Biogenesis. J. Med. Food 2019, 22, 1159–1167. [Google Scholar] [CrossRef]
- Magnone, M.; Spinelli, S.; Begani, G.; Guida, L.; Sturla, L.; Emionite, L.; Zocchi, E. Abscisic Acid Improves Insulin Action on Glycemia in Insulin-Deficient Mouse Models of Type 1 Diabetes. Metabolites 2022, 12, 523. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Scholtes, C.; Dufour, C.R.; Ouellet, C.; Ghahremani, M.; Giguère, V. Insulin action and resistance are dependent on a GSK3β-FBXW7-ERRα transcriptional axis. Nat. Commun. 2022, 13, 2105. [Google Scholar] [CrossRef]
- Billon, C.; Schoepke, E.; Avdagic, A.; Chatterjee, A.; Butler, A.A.; Elgendy, B.; Walker, J.K.; Burris, T.P. A Synthetic ERR Agonist Alleviates Metabolic Syndrome. J. Pharmacol. Exp. Ther. 2024, 388, 232–240. [Google Scholar] [CrossRef]
- Le Page-Degivry, M.T.; Bidard, J.N.; Rouvier, E.; Bulard, C.; Lazdunski, M. Presence of abscisic acid, a phytohormone, in the mammalian brain. Proc. Natl. Acad. Sci. USA 1986, 83, 1155–1158. [Google Scholar] [CrossRef]
- Strum, J.C.; Shehee, R.; Virley, D.; Richardson, J.; Mattie, M.; Selley, P.; Ghosh, S.; Nock, C.; Saunders, A.; Roses, A. Rosiglitazone induces mitochondrial biogenesis in mouse brain. J. Alzheimers Dis. 2007, 11, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Kim, J.; Moon, S.; Choi, B.Y.; Kim, S.; Jeon, H.S.; Suh, S.W.; Kim, Y.M.; Choi, Y.K. Korean Red Ginseng Improves Astrocytic Mitochondrial Function by Upregulating HO-1-Mediated AMPKα-PGC-1α-ERRα Circuit after Traumatic Brain Injury. Int. J. Mol. Sci. 2021, 22, 13081. [Google Scholar] [CrossRef]
- Xie, Z.; Cao, B.Q.; Wang, T.; Lei, Q.; Kang, T.; Ge, C.Y.; Gao, W.J.; Hui, H. LanCL1 attenuates ischemia-induced oxidative stress by Sirt3-mediated preservation of mitochondrial function. Brain Res. Bull. 2018, 142, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Park, W.; Kim, G.J.; Choi, H.S.; Vanacker, J.M.; Sohn, Y.C. Conserved properties of a urochordate estrogen receptor-related receptor (ERR) with mammalian ERRalpha. Biochim. Biophys. Acta 2009, 1789, 125–134. [Google Scholar] [CrossRef]
- Mercurio, S.; Tremolada, P.; Nobile, M.; Fernandes, D.; Porte, C.; Sugni, M. Unraveling estradiol metabolism and involvement in the reproductive cycle of non-vertebrate animals: The sea urchin model. Steroids 2015, 104, 25–36. [Google Scholar]
- Baker, M.E. Trichoplax, the simplest known animal, contains an estrogen-related receptor but no estrogen receptor: Implications for estrogen receptor evolution. Biochem. Biophys. Res. Commun. 2008, 375, 623–627. [Google Scholar] [CrossRef] [PubMed]
- Fresia, C.; Vigliarolo, T.; Guida, L.; Booz, V.; Bruzzone, S.; Sturla, L.; Di Bona, M.; Pesce, M.; Usai, C.; De Flora, A.; et al. G-protein coupling and nuclear translocation of the human abscisic acid receptor LANCL2. Sci. Rep. 2016, 6, 26658. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spinelli, S.; Bruschi, M.; Passalacqua, M.; Guida, L.; Magnone, M.; Sturla, L.; Zocchi, E. Estrogen-Related Receptor α: A Key Transcription Factor in the Regulation of Energy Metabolism at an Organismic Level and a Target of the ABA/LANCL Hormone Receptor System. Int. J. Mol. Sci. 2024, 25, 4796. https://doi.org/10.3390/ijms25094796
Spinelli S, Bruschi M, Passalacqua M, Guida L, Magnone M, Sturla L, Zocchi E. Estrogen-Related Receptor α: A Key Transcription Factor in the Regulation of Energy Metabolism at an Organismic Level and a Target of the ABA/LANCL Hormone Receptor System. International Journal of Molecular Sciences. 2024; 25(9):4796. https://doi.org/10.3390/ijms25094796
Chicago/Turabian StyleSpinelli, Sonia, Maurizio Bruschi, Mario Passalacqua, Lucrezia Guida, Mirko Magnone, Laura Sturla, and Elena Zocchi. 2024. "Estrogen-Related Receptor α: A Key Transcription Factor in the Regulation of Energy Metabolism at an Organismic Level and a Target of the ABA/LANCL Hormone Receptor System" International Journal of Molecular Sciences 25, no. 9: 4796. https://doi.org/10.3390/ijms25094796