Influence of Long-Tailed Alcohols on the Solubilisation of Cationic DHAB Bilayers

Abstract
:Introduction


Results and Discussion
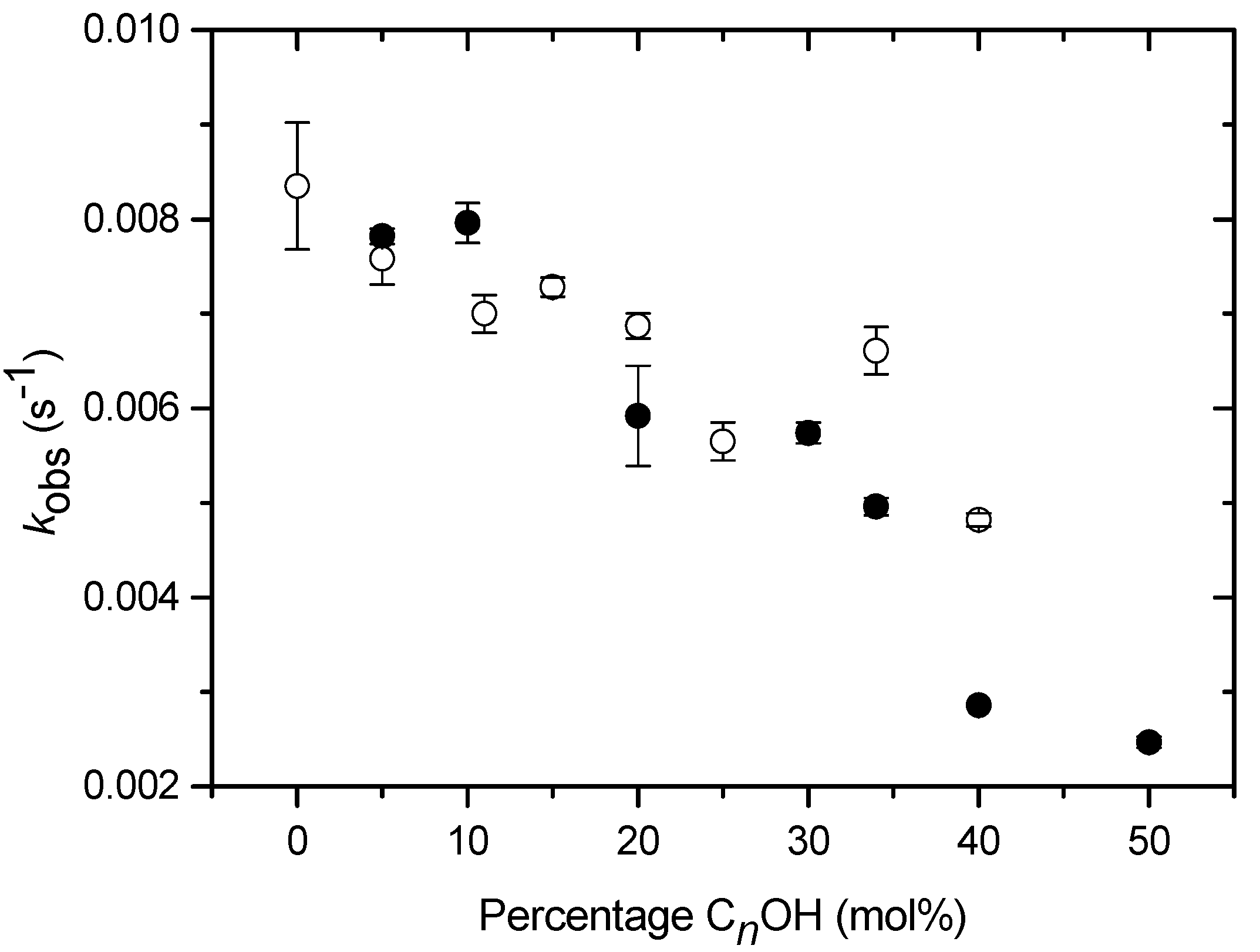
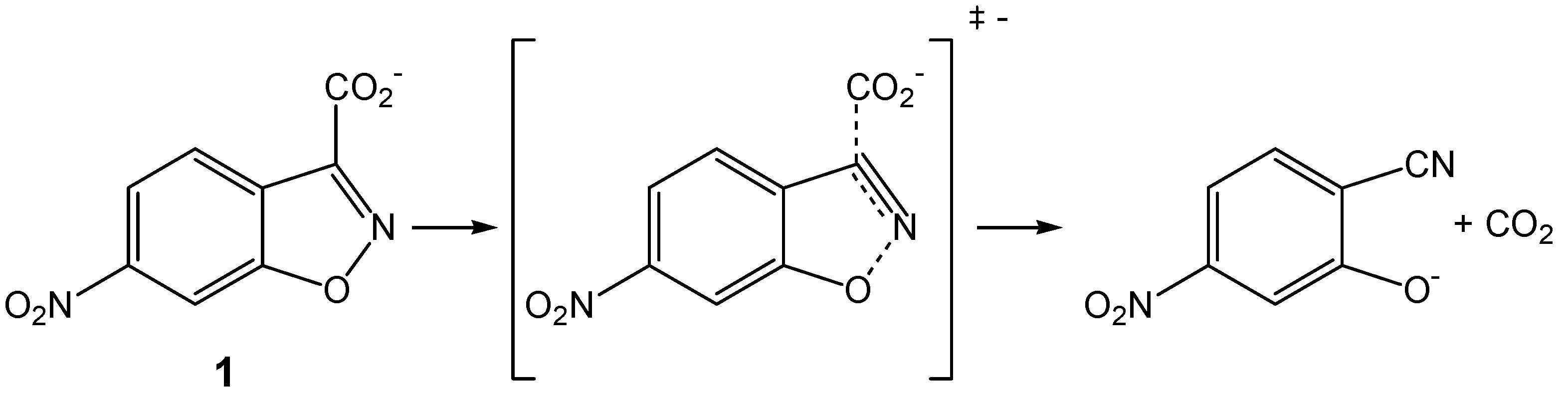
Kinetics

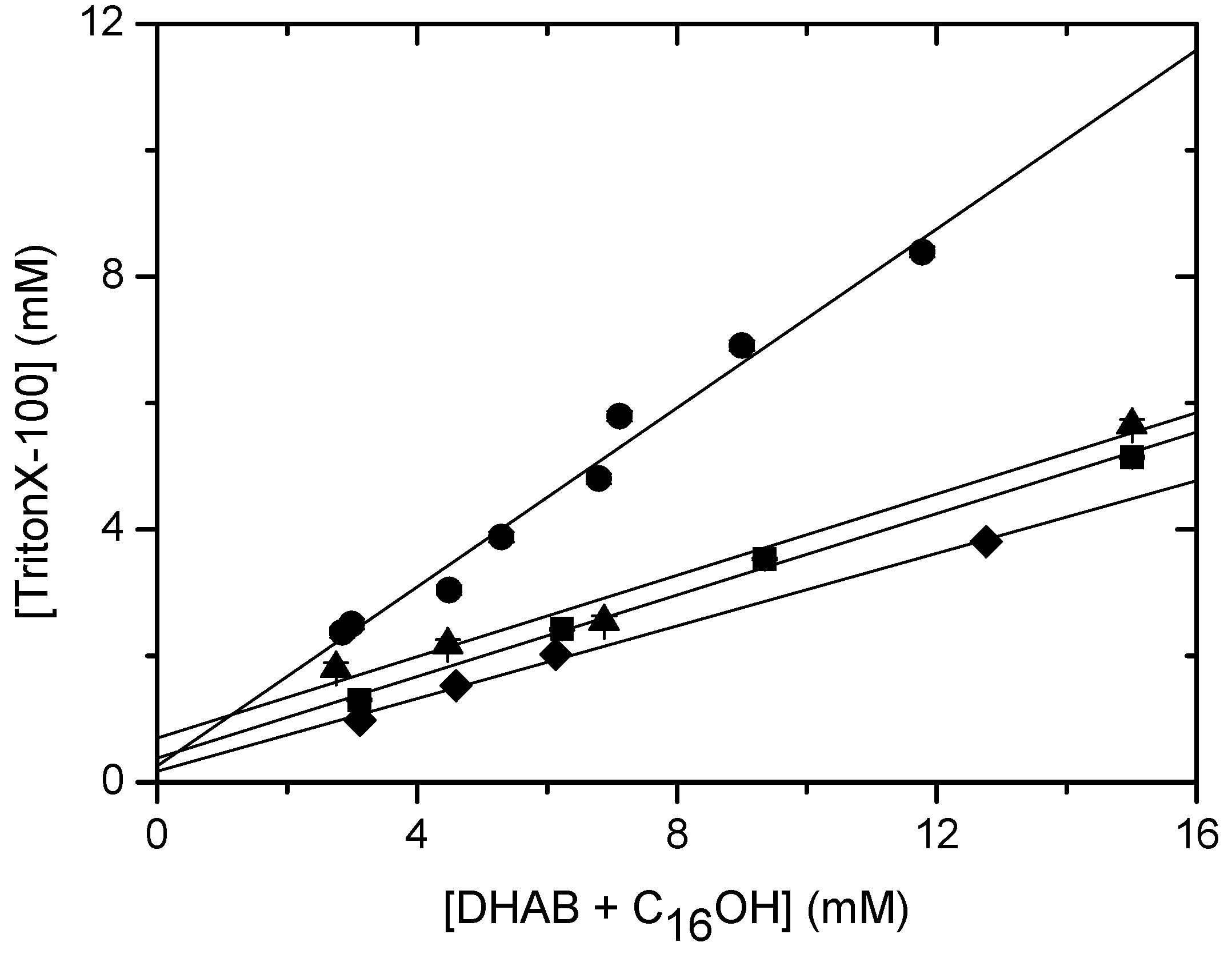
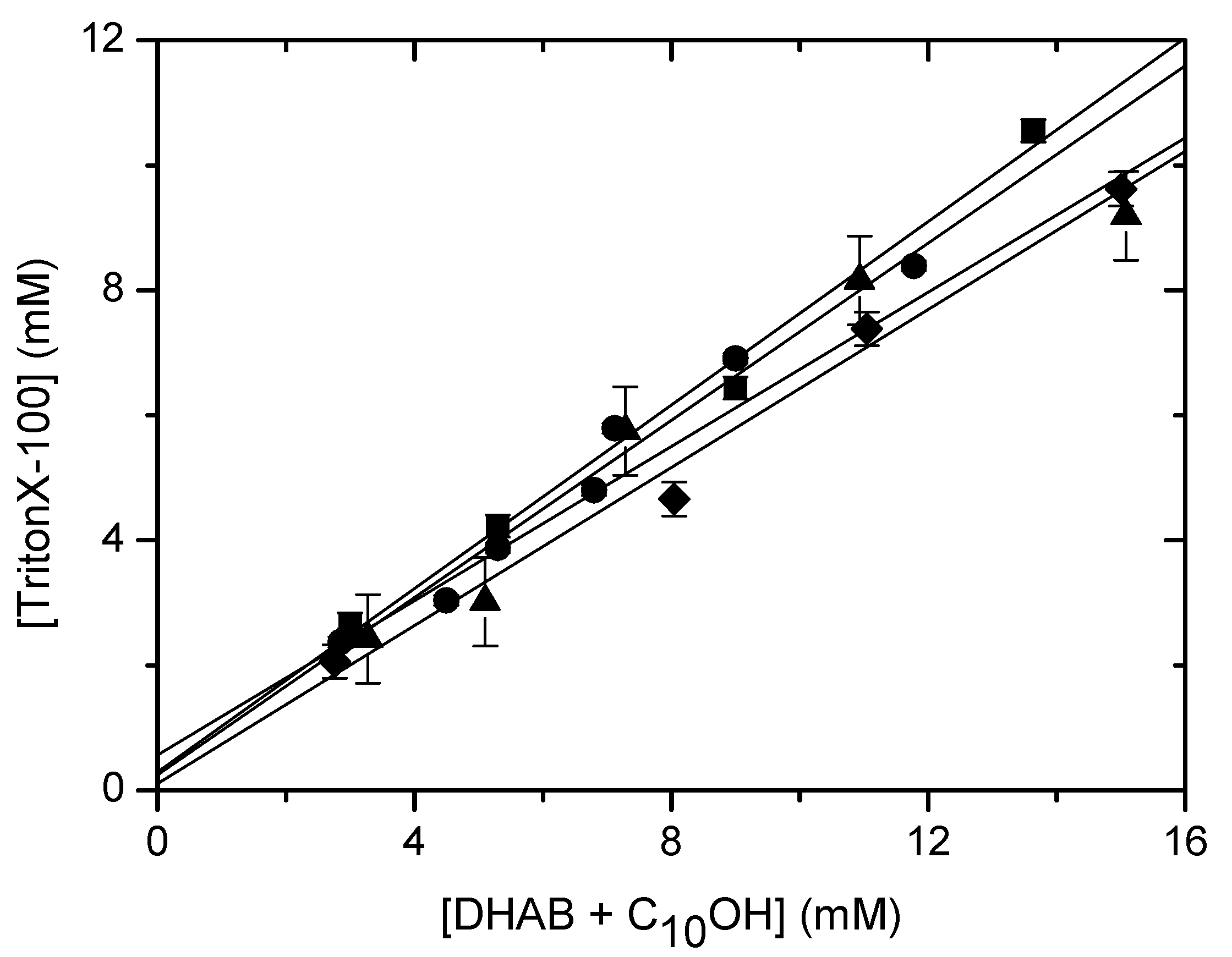
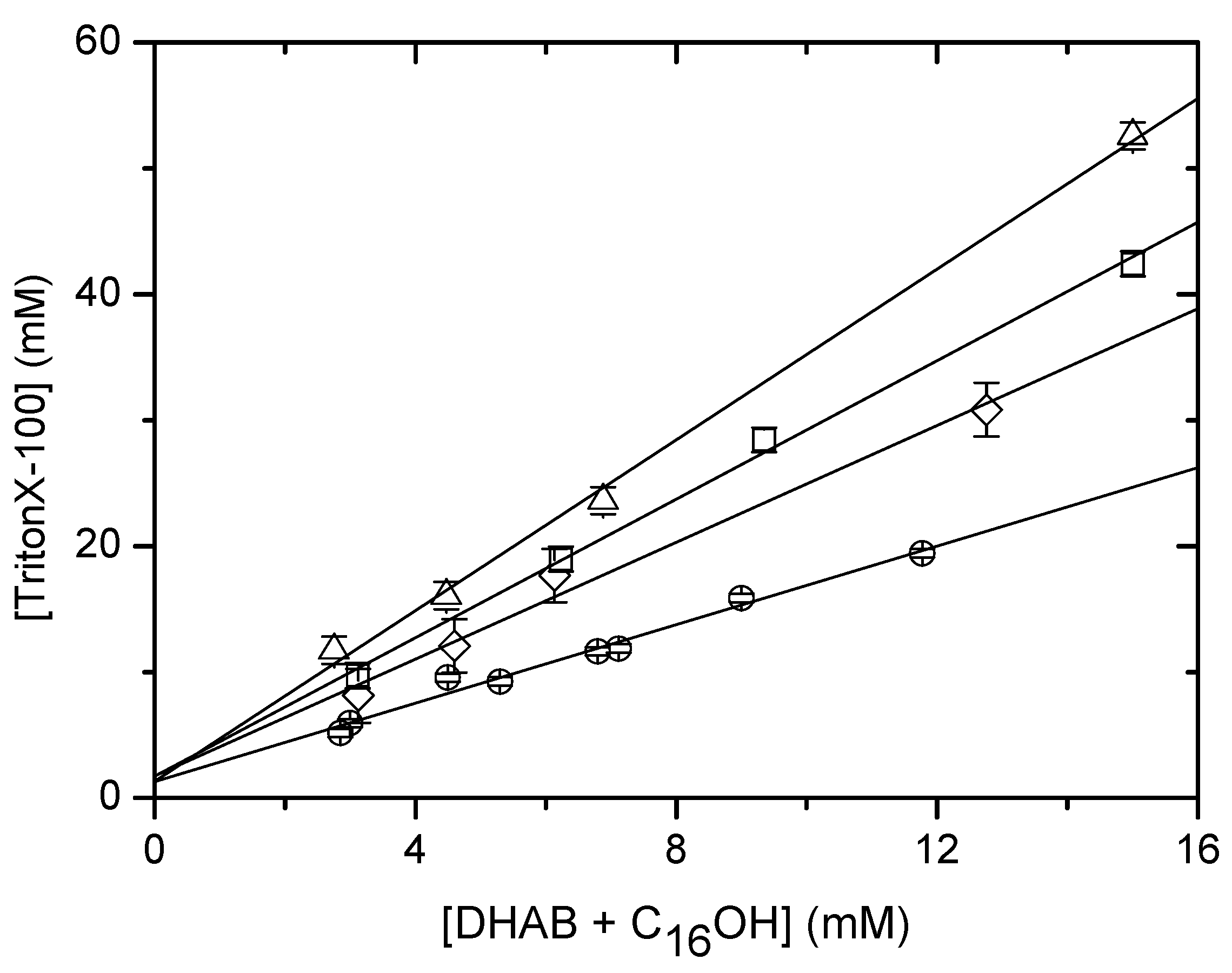
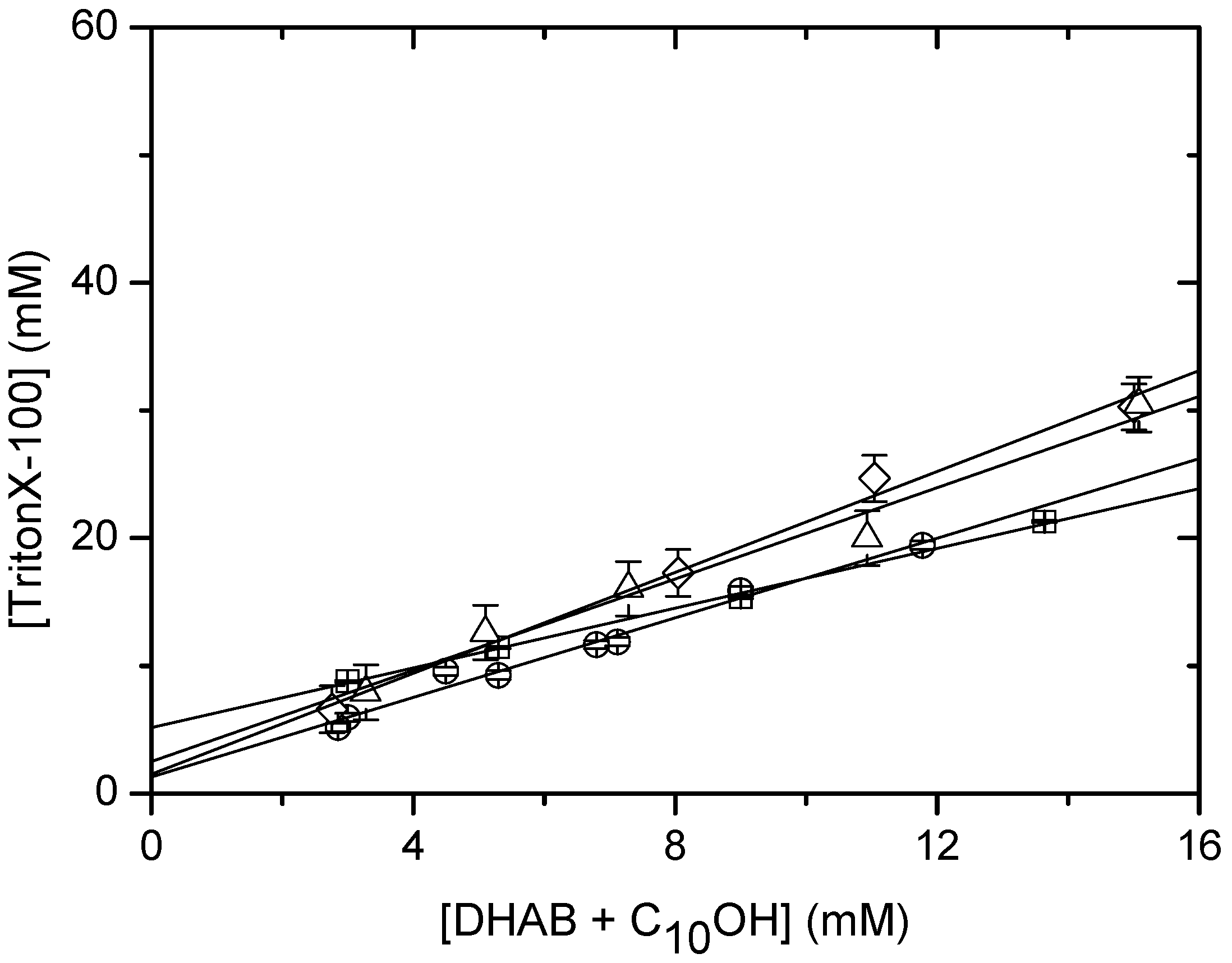
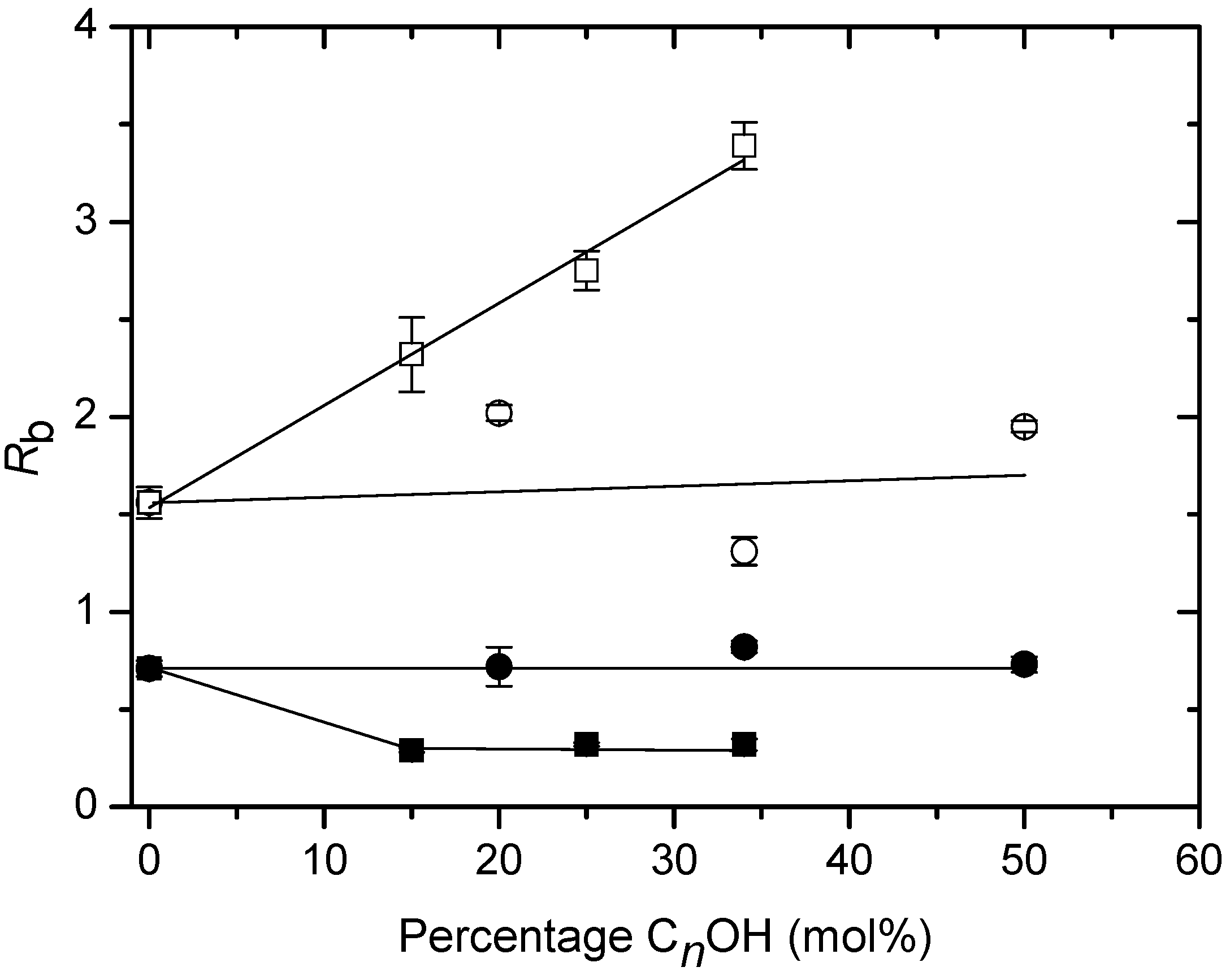
Solubilisation of DHAB/CnOH (n=10,16) mixtures





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
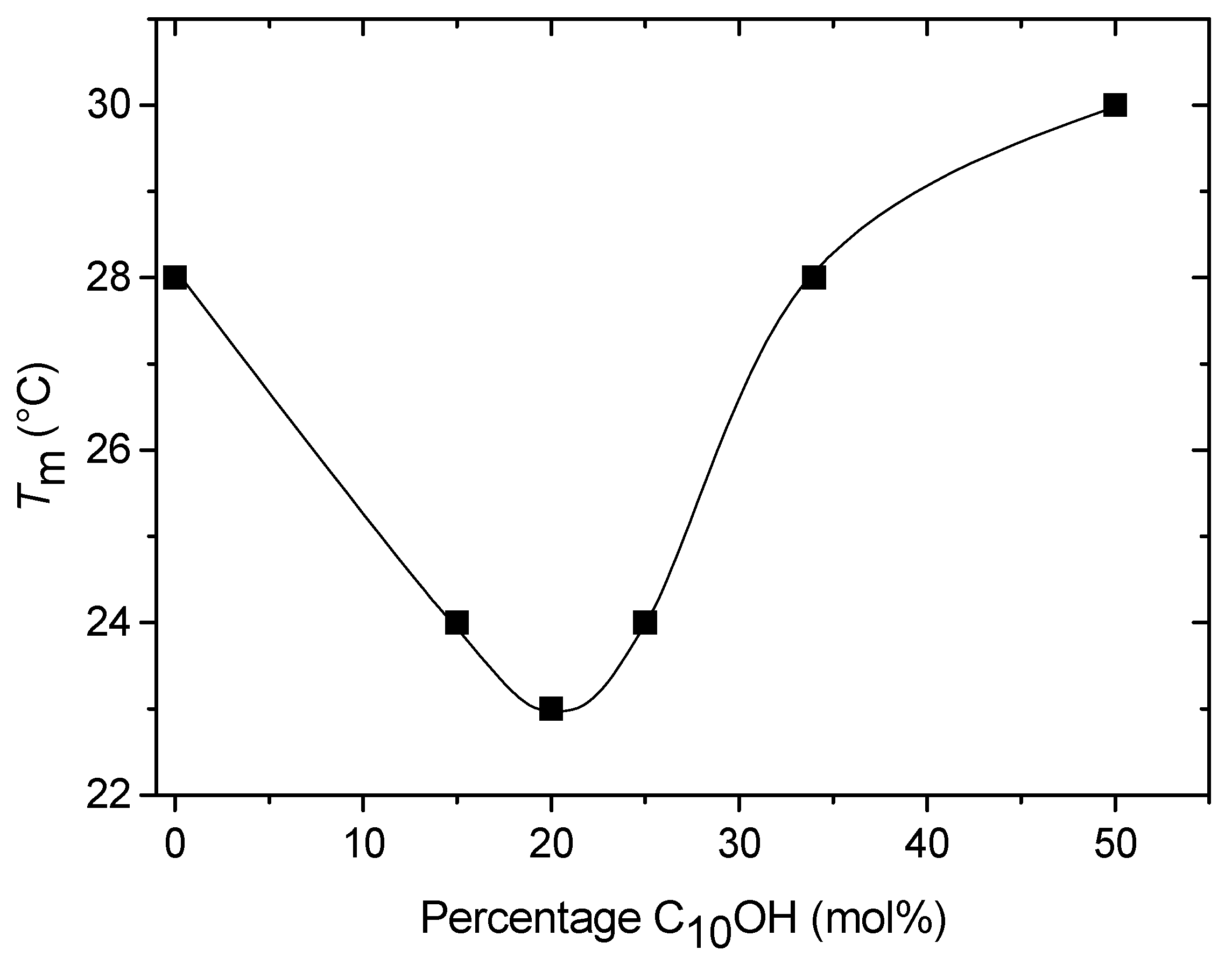
Differential Scanning Calorimetry

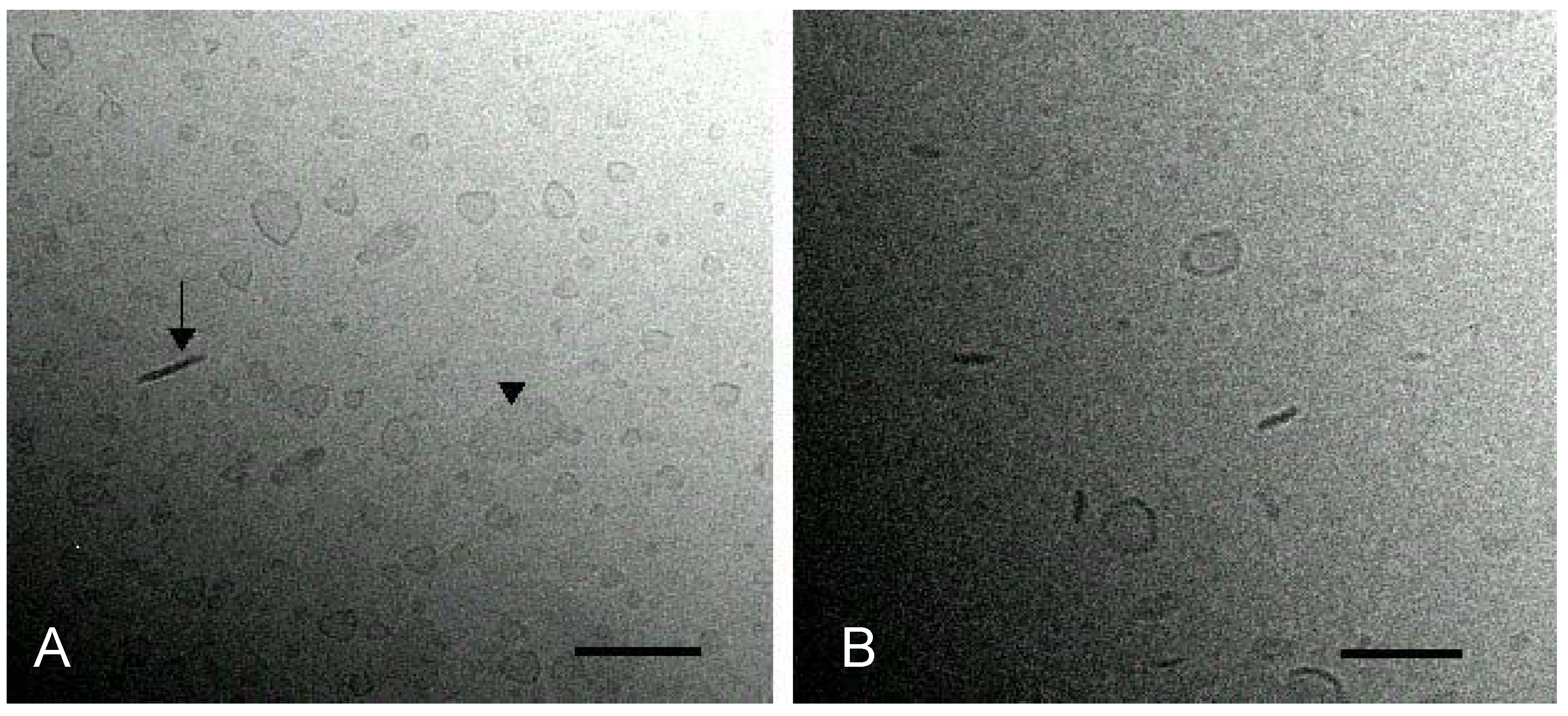
Cryo-electron microscopy

Matching and mismatching tails
Conclusions
Experimental
Materials
Solubilisation measurements
Kinetic measurements
DSC measurements
Cryo-Electron Microscopy
Acknowledgements
References
- Gennis, R. B. Biomembranes: Molecular Structure and Function; Springer-Verlag: New York, 1989; p. 20. [Google Scholar]
- Darnell, J.; Lodish, H.; Baltimore, D. Molecular Cell Biology; Scientific American Books: New York, 1990; p. 491. [Google Scholar]
- Alberts, B.; Bray, D.; Lewis, J.; Raff, M.; Roberts, K.; Watson, J. D. Molecular Biology of the Cell; Garland Publishing: New York, 1994; p. 477. [Google Scholar]
- Ahmed, S. N.; Brown, D. A.; London, E. On the origin of sphingolipid/cholesterol-rich detergent- insoluble cell membranes: Physiological concentrations of cholesterol and sphingolipid induce formation of a detergent- insoluble, liquid-ordered lipid phase in model membranes. Biochemistry 1997, 36, 10944. [Google Scholar]
- Simons, K.; Ikonen, E. Functional rafts in cell membranes. Nature 1997, 387, 569. [Google Scholar]
- London, E.; Brown, D. A. Insolubility of lipids in Triton X-100: physical origin and relationship to sphingolipid/cholesterol membrane domains (rafts). Biochim. Biophys. Acta 2000, 1508, 182. [Google Scholar] [CrossRef]
- Rietveld, A.; Simons, K. The differential miscibility of lipids as the basis for the formation of functional membrane rafts. Biochim. Biophys. Acta 1998, 1376, 467. [Google Scholar]
- Schroeder, R. J.; Ahmed, S. N.; Zhu, Y. Z.; London, E.; Brown, D. A. Cholesterol and sphingolipid enhance the Triton X-100 insolubility of glycosylphosphatidylinositol-anchored proteins by promoting the formation of detergent-insoluble ordered membrane domains. J. Biol. Chem. 1998, 273, 1150. [Google Scholar] [CrossRef]
- Pralle, A.; Keller, P.; Florin, E. L.; Simons, K.; Horber, J. K. H. Sphingolipid-cholesterol rafts diffuse as small entities in the plasma membrane of mammalian cells. J. Cell Biol. 2000, 148, 997. [Google Scholar] [CrossRef]
- Simons, K.; Ikonen, E. Cell biology - How cells handle cholesterol. Science 2000, 290, 1721. [Google Scholar]
- Zubay, G. Biochemistry; Addison-Wesley Publishing Company: Reading (USA), 1983. [Google Scholar]
- Eliasz, A. W.; Chapman, D.; Ewing, D. F. Phospholipid Phase Transitions. Biochim. Biophys. Acta 1976, 448, 220. [Google Scholar] [CrossRef]
- Hui, F. K.; Barton, P. G. Mesomorphic behavior of some phospholipids with aliphatic alcohols and other non-ionic substances. Biochim. Biophys. Acta 1973, 296, 510. [Google Scholar] [CrossRef]
- Lee, A. G. Interactions between anesthetics and lipid mixtures. Normal Alcohols. Biochemistry 1976, 15, 2448. [Google Scholar] [CrossRef]
- Mabrey, S.; Sturtevant, J. M. Incorporation of saturated fatty acid into phosphatidylcholine bilayers. Biochim. Biophys. Acta 1977, 486, 444. [Google Scholar] [CrossRef]
- Jain, M. K.; Wu, N. M. Effects of Small Molecules on the Dipalmitoyl Lecithin Liposomal Bilayer. J. Membrane Biol. 1977, 34, 157. [Google Scholar] [CrossRef]
- Zavoico, G. B.; Chandler, L.; Kutchai, H. Pertubation of egg phosphatidylcholine and dipalmitoylcholine multilamellar vesicles by n-alcohols. A fluorescent probe study. Biochim. Biophys. Acta 1985, 812, 299. [Google Scholar] [CrossRef]
- Campos, A. M.; Abuin, E. B.; Lissi, E. A. Effect of a linear (1-octanol) and a branched (2,6-dimethyl-4- heptanol) alkanol upon the properties of dipalmitoylphosphatidylcholine large unilamellar vesicles. Colloids Surf. A 1995, 100, 155. [Google Scholar]
- Aspée, A.; Lissi, E. A. Solubilization of alkanols in DPPC luvs - dependence on the alkanol concentration and topology. J. Colloid Interface Sci. 1995, 175, 225. [Google Scholar] [CrossRef]
- Blandamer, M. J.; Briggs, B.; Cullis, P. M.; Irlam, K. D.; Kirby, S. D.; Engberts, J. B. F. N. Formation of vesicular bilayers in aqueous solutions containing mixtures of dialkyldimethylammonium bromides. J. Mol. Liq. 1998, 75, 181. [Google Scholar] [CrossRef]
- Kacperska, A. DSC investigations of DDAB, DTAB and DHAB vesicle aqueous solutions in presence of SDS. J. Therm. Anal. 2000, 61, 63. [Google Scholar] [CrossRef]
- Lee, A. G. Lipid phase-transitions and phase-diagrams .2. Mixtures involving lipids. Biochim. Biophys. Acta 1977, 472, 285. [Google Scholar]
- Kemp, D. S.; Paul, K. G. Decarboxylation of benzisoxazole-3-carboxylic acids. Catalysis by extraction of possible relevance to the problem of enzymatic mechanism. J. Am. Chem. Soc. 1970, 92, 2553. [Google Scholar]
- Grate, J. W.; McGill, R. A.; Hilvert, D. Analysis of solvent effects on the decarboxylation of benzisoxazole-3-carboxylate ions using linear solvation energy relatinships: relevance to catalysis in an antibody binding site. J. Am. Chem. Soc. 1993, 115, 8577. [Google Scholar] [CrossRef]
- Talhout, R.; Engberts, J. B. F. N. Self-assembly in mixtures of sodium alkyl sulfates and alkyltrimethylammonium bromides: Aggregation behavior and catalytic properties. Langmuir 1997, 13, 5001. [Google Scholar]
- Brinchi, L.; Germani, R.; Goracci, L.; Savelli, G.; Bunton, C. A. Decarboxylation and dephosphorylation in new gemini surfactants. Changes in aggregate structures. Langmuir 2002, 18, 7821. [Google Scholar]
- Romsted, L. S. Micellization, Solubilization and Microemulsions; Mittal, K. L., Ed.; Plenum Press: New York, 1977; p. 509. [Google Scholar]
- Scarpa, M. V.; Araujo, P. S.; Schreier, S.; Sesso, A.; Oliveira, A. G.; Chaimovich, H.; Cuccovia, I. M. Effect of vesicles of dimethyldioctadecylammonium chloride and phospholipids on the rate of decarboxylation of 6-nitrobenzisoxazole-3-carboxylate. Langmuir 2000, 16, 993. [Google Scholar]
- Schurtenberger, P.; Mazer, N.; Kanzig, W. Micelle to Vesicle Transition in Aqueous-Solutions of Bile-Salt and Lecithin. J. Phys. Chem. 1985, 89, 1042. [Google Scholar] [CrossRef]
- Kragh-Hansen, U.; le Maire, M.; Møller, J. V. The mechanism of detergent solubilization of liposomes and protein-containing membranes. Biophys. J. 1998, 75, 2932. [Google Scholar]
- Johnsson, M.; Wagenaar, A.; Stuart, M. C. A.; Engberts, J. B. F. N. Sugar-based gemini surfactants with pH-dependent aggregation behavior: Vesicle-to-micelle transition, critical micelle concentration, and vesicle surface charge reversal. Biophys 2003, 19, 4609. [Google Scholar]
- Edwards, K.; Almgren, M. Solubilization of lecithin vesicles by C12E8 - structural transitions and temperature effects. J. Colloid Interface Sci. 1991, 147, 1. [Google Scholar] [CrossRef]
- Silvander, M.; Karlsson, G.; Edwards, K. Vesicle solubilization by alkyl sulfate surfactants: A cryo-TEM study of the vesicle to micelle transition. J. Colloid Interface Sci. 1996, 179, 104. [Google Scholar] [CrossRef]
- Wang, G. J.; Engberts, J. B. F. N. Synthesis of hydrophobically and electrostatically modified polyacrylamides and their catalytic effects on the unimolecular decarboxylation of 6-nitrobenzisoxazole-3-carboxylate anion. Langmuir 1995, 11, 3856. [Google Scholar]
- Wang, G. J.; Engberts, J. B. F. N. Synthesis and catalytic properties of non-cross-linked and cross-linked poly(alkylmethyldiallylammonium bromides) having decyl, octyl, and hexyl side-chains. J. Org. Chem. 1995, 60, 4030. [Google Scholar] [CrossRef]
- Wang, G. J.; Engberts, J. B. F. N. Macromolecular flexibility and aggregation tendency of non-cross-linked and crosslinked cationic polysoaps. Eur. Polym. J. 1998, 34, 1319. [Google Scholar]
- Brinchi, L.; di Profio, P.; Germani, R.; Savelli, G.; Spreti, N. Effect of ethanol on micellization and on decarboxylation of 6- nitrobenzisoxazole-3-carboxylate in aqueous cationic micelles. J. Colloid Interface Sci. 2002, 247, 429. [Google Scholar] [CrossRef]
- Caussanel, F.; Andre-Barrès, C.; Lesieur, S.; Rico-Lattes, I. A comparative study of sugar-based surfactants for the solubilization of phosphatidylcholine vesicles. Colloids Surf. B 2001, 22, 193. [Google Scholar]
- Chu, S. C.; Hung, C. H.; Wang, S. C.; Tsao, H. K. Partition thermodynamics of ionic surfactants between phosphatidylcholine vesicle and water phases. J. Chem. Phys. 2003, 119, 3441. [Google Scholar]
- Memoli, A.; Annesini, M. C.; Petralito, S. Surfactant-induced leakage from liposomes: a comparison among different lecithin vesicles. Int. J. Pharm 1999, 184, 227. [Google Scholar]
- Tan, A. M.; Ziegler, A.; Steinbauer, B.; Seelig, J. Thermodynamics of sodium dodecyl sulfate partitioning into lipid membranes. Biophys. J. 2002, 83, 1547. [Google Scholar]
- Carion-Taravella, B.; Lesieur, S.; Chopineau, J.; Lesieur, P.; Ollivon, M. Phase behavior of mixed aqueous dispersions of dipalmitoylphosphatidylcholine and dodecyl glycosides: A differential scanning calorimetry and X-ray diffraction investigation. Langmuir 2002, 18, 325. [Google Scholar]
- Klijn, J. E.; Engberts, J. B. F. N. The Kemp elimination in membrane mimetic reaction media. Probing catalytic properties of cationic vesicles formed from a double-tailed amphiphile and linear long-tailed alcohols or alkyl pyranosides. Org. Biomol. Chem. 2004, 2, 1789. [Google Scholar] [PubMed]
- Hammerström, L.; Velikian, I.; Karlsson, G.; Edwards, K. Cryo-tem evidence - sonication of dihexadecyl phosphate does not produce closed bilayers with smooth curvature. Langmuir 1995, 11, 408. [Google Scholar]
- Andersson, M.; Hammerström, L.; Edwards, K. Effect of bilayer phase-transitions on vesicle structure and its influence on the kinetics of viologen reduction. J. Phys. Chem. 1995, 99, 14531. [Google Scholar] [CrossRef]
- Borsche, W. Über a-Dinitrophenyl-acetessigester und verwandte Verbindungen. Chem. Ber. 1909, 42, 1310. [Google Scholar] [CrossRef]
- Lindemann, H.; Cissée, H. Ringöffnung bei Benz-a,b-isoxazolen. Liebigs Ann. Chem. 1929, 469, 44. [Google Scholar]
- Sample Availability: Available from the authors.
© 2005 by MDPI (http://www.mdpi.org). Reproduction is permitted for noncommercial purposes.
Share and Cite
Linthorst, J.A.; Klijn, J.E.; Engberts, J.B.F.N. Influence of Long-Tailed Alcohols on the Solubilisation of Cationic DHAB Bilayers. Int. J. Mol. Sci. 2005, 6, 104-118. https://doi.org/10.3390/i6010104
Linthorst JA, Klijn JE, Engberts JBFN. Influence of Long-Tailed Alcohols on the Solubilisation of Cationic DHAB Bilayers. International Journal of Molecular Sciences. 2005; 6(1):104-118. https://doi.org/10.3390/i6010104
Chicago/Turabian StyleLinthorst, Johan A., Jaap E. Klijn, and Jan B.F.N. Engberts. 2005. "Influence of Long-Tailed Alcohols on the Solubilisation of Cationic DHAB Bilayers" International Journal of Molecular Sciences 6, no. 1: 104-118. https://doi.org/10.3390/i6010104