Plant Products for Pharmacology: Application of Enzymes in Their Transformations


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
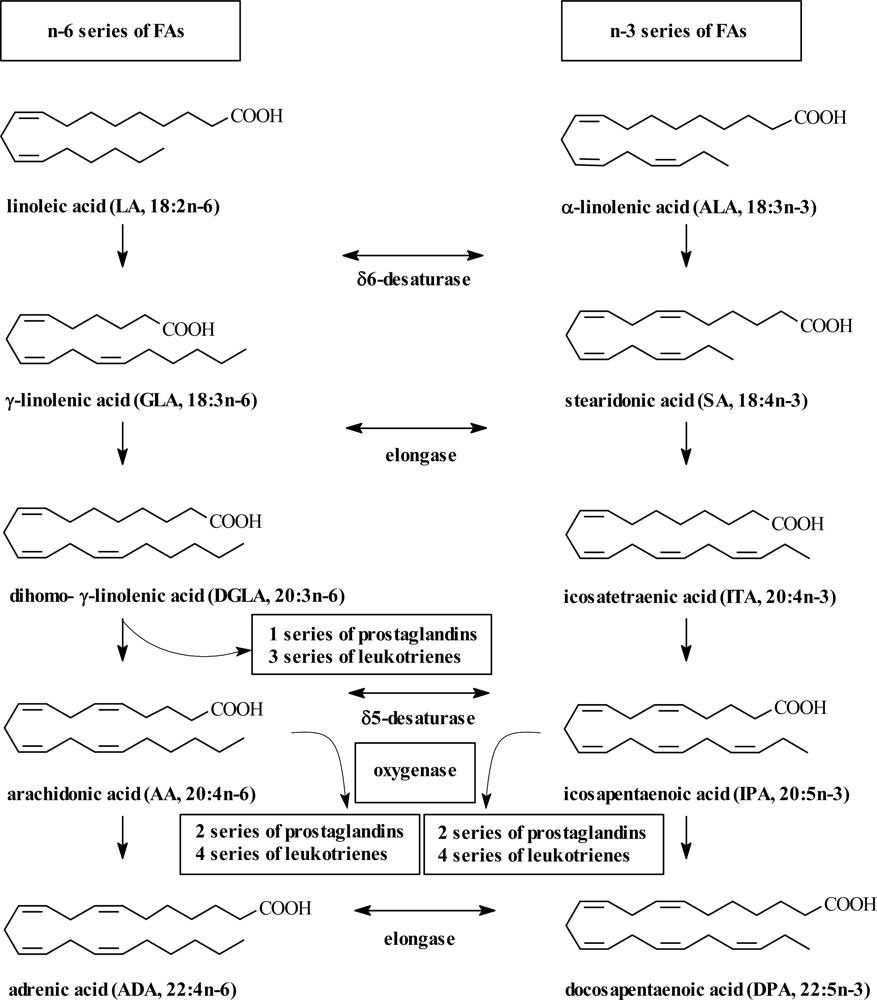
2. PUFA Sources
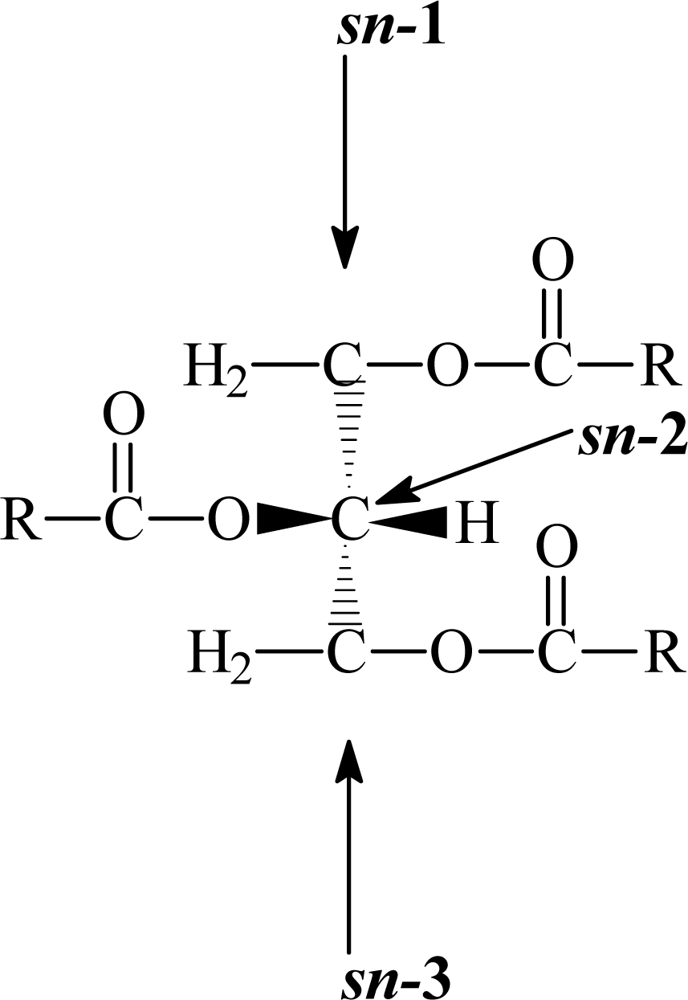
3. Triacylglycerol structure
4. Enzymes acting in lipid modifications
4.1. Lipases
4.2. Lipase structure and catalytic ability
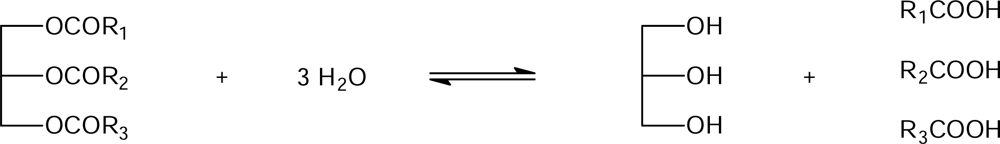
4.3. Lipase-catalyzed hydrolysis under conventional conditions
4.4. Lipase-catalyzed reactions under non-conventional conditions
4.4.1. Properties of supercritical carbon dioxide
4.4.2. Properties of ionic liquids
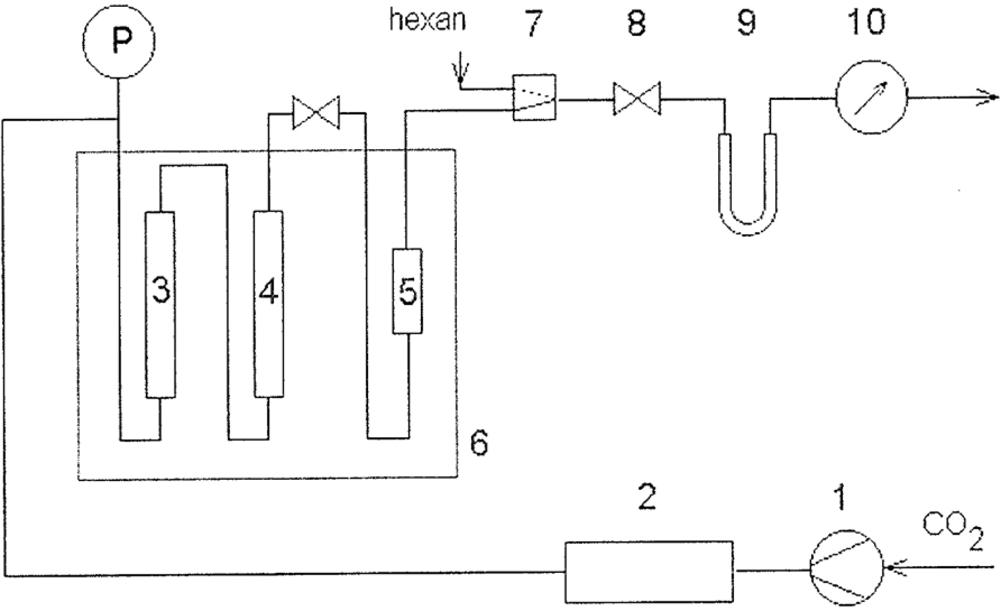
4.4.3. Lipase-catalyzed hydrolysis in supercritical carbon dioxide
4.5. Lipase-catalyzed esterification in organic solvents
4.6. Lipase-catalyzed transesterification and acidolysis under conventional conditions
4.7. Structured lipids
4.7.1. Lipase-catalyzed esterification in supercritical carbon dioxide
4.7.2. Other types of lipase-catalyzed reactions in supercritical carbon dioxide
4.9. Lipase-catalyzed reactions in ionic liquid / supercritical carbon dioxide biphasic systems
5. Conclusions: Importance of the reaction media for sustainability
Acknowledgments
References
- Emken, EA; Adlof, RO; Gulley, RM. Dietary linoleic acid influences desaturation and acylation of deuterium-labeled linoleic and linolenic acids in young adult males. Biochim. Biophys. Acta 1994, 1213, 277–288. [Google Scholar]
- Holub, DJ; Holub, BJ. Omega-3 fatty acids from fish oils and cardiovascular disease. Mol. Cell Biochem 2004, 263, 217–225. [Google Scholar]
- Peet, M; Brind, J; Ramchand, CN; Shah, S; Vankar, GK. Two double blind placebo controlled pilot studies of eicosapentaenoic acid in the treatment of schizophrenia. Schizophr. Res 2001, 49, 243–251. [Google Scholar]
- Simopoulos, AP. Omega-3 fatty acids in health and disease and in growth and development. Am. J. Clin. Nutr 1991, 54, 438–463. [Google Scholar]
- von Schacky, C. Omega-3 fatty acids and cardiovascular disease. Curr. Opin. Clin. Nutr. Metab.Care 2004, 7, 31–136. [Google Scholar]
- Stahl, SM. Essential psychopharmacology. Neuroscientific basis and practical applications, 2nd Ed ed; Cambridge University Press: Cambridge, 2000. [Google Scholar]
- Glen, AIM; Ross, BM. Eicosanoids in the central nervous system. In The Eicosanoids; Curtis-Prior, P, Ed.; Wiley: London, UK, 2004; pp. 210–220. [Google Scholar]
- Carlezon, WA, Jr; Mague, SD; Parow, AM; Stoll, AL; Cohen, BM; Renshaw, PF. Antidepressant-like effects of uridine and omega-3 fatty acids are potentiated by combined treatment in rats. Biol. Psychiatry 2005, 57, 343–350. [Google Scholar]
- Ross, BM; Seguin, J; Sieswerda, LE. Omega-3 fatty acids as treatments for mental illness: Which disorder and which fatty acid? Lipids Health Dis 2007, 6, 21–30. [Google Scholar]
- Xiao, YF; Sigg, DC; Leaf, A. The antiarrhythmic effect of n-3 polyunsaturated fatty acids: Modulation of cardiac ion channels as a potential mechanism. J. Membr. Biol 2005, 206, 141–154. [Google Scholar]
- Wigmore, SJ; Ross, JA; Falconer, JS; Plester, CE; Tisdade, MJ; Carter, DC; Faron, KCH. Effect of PUFA on the progress of cachexia in patiens with pancreatic cancer. Nutrition 1996, 12, 27–30. [Google Scholar]
- Beaubaire, NA; Simon, JE. Production potential of Borago oficinalis L. Acta Horticult 1987, 208, 101–103. [Google Scholar]
- Senanayake, SPJN; Shahidi, F. Enzyme-assisted acidolysis of borage (Borago officinalis L.) and evening primrose (Oenothera biennis L.) oils: Incorporation of omega-3 polyunsaturated fatty acids. J. Agric. Food Chem 1999, 47, 3105–3112. [Google Scholar]
- Stránský, K; Zarevúcka, M; Wimmer, Z. Gas chromatography analysis of blackcurrant oil in relation to its stability. Food Chem 2005, 92, 569–573. [Google Scholar]
- Traitler, H; Wille, HJ; Studer, A. Fractionation of Blackcurrant seed oil. J. Am. Oil Chem. Soc 1988, 65, 755–760. [Google Scholar]
- Wille, HJ; Traitler, H; Lagarde, M. Preparation of stearidonic acid concentrates. Fat Sci. Technol 1991, 93, 362–368. [Google Scholar]
- Traitler, H; Wille, HJ. Isolation of pure fatty acids from fats and oils. Fat Sci. Technol 1992, 94, 506–511. [Google Scholar]
- Syed Rahmatullah, MSK; Shukla, VKS; Mukherjee, KD. Enrichment of γ-linolenic acid from evening primrose oil and borage oil via lipase-catalyzed hydrolysis. J. Am. Oil Chem. Soc 1994, 71, 560–573. [Google Scholar]
- Syed Rahmatullah, MSK; Shukla, VKS; Mukherjee, KD. γ-Linolenic acid concentrates from borage and evening priprose oil via lipase-catalyzed esterification. J. Am. Oil Chem. Soc 1994, 71, 563–577. [Google Scholar]
- Kawashima, H; Sakuradani, E; Kamada, N; Akimoto, K; Konishi, K; Ogawa, J; Shimizu, S. Production of 8,11,14,17-cis-eicosatetraenoic acid (20 : 4 omega-3) by a Delta 5 and Delta 12 desaturase-defective mutant of an arachidonic acid-producing fungus Mortierella alpina 1S-4. J. Am. Oil Chem. Soc 1998, 75, 1495–1500. [Google Scholar]
- Shinmen, Y; Shimizu, S; Akimoto, D; Kawashima, H; Yamada, H. Production of arachidonic acid by Mortierella fungi. Selection of a potent producer and optimization of culture conditions for large-scale production. Appl. Microbiol. Biotechnol 1989, 31, 11–16. [Google Scholar]
- Yongmanitchai, W; Ward, OP. Screening of algae for potential alternative sources of eicosapentaenoic acid. Phytochemistry 1991, 30, 2963–2967. [Google Scholar]
- Nichols, BW; Appleby, RS. The distribution and biosynthesis of arachidonic acid in algae. Phytochemistry 1969, 8, 1907–1915. [Google Scholar]
- Monroig, O; Navarro, JC; Amat, F; Gonzáles, P; Bermejo, A; Hontoria, F. Enrichment of Artemia nauplii in essential fatty acids with different types of liposomes and their use in the rearing of gilthead sea bream (Sparus aurata) larvae. Aquaculture 2006, 251, 491–508. [Google Scholar]
- Zhu, J-J; Shi, J-H; Qian, W-B; Cai, Z-Z; Li, D. Effects of krill oil on serum lipids of hyperlipidemic rats and human SW480 cells. Lipids Health Dis 2008, 7, 30. [Google Scholar]
- Shimada, Y; Sugihara, A; Nakano, H; Kuramoto, T; Nagao, T; Gemba, M; Tominaga, Y. Purification of docosahexaenoic acid by selective esterification of fatty acids from tuna oil with Rhizopus delemar lipase. J. Am. Oil Chem. Soc 1997, 74, 97–101. [Google Scholar]
- Salunkhe, DK; Chavan, JK; Adsule, RN; Kadam, SS. World Oilseeds: Chemistry, Technology, and Utilization; Salunkhe, DK, Van Nostrand, R, Eds.; Springer: New York, USA, 1992. [Google Scholar]
- Gunstone, FD; Ilyas-Qureshi, M. Glyceride studies. Part IV. The component glycerides of ten seed oils containing linoleic acid. J. Am. Oil. Chem. Soc 1965, 42, 961–965. [Google Scholar]
- Gunstone, FD; Hamilton, RJ; Padley, FB; Ilyas-Qureshi, M. Glyceride studies. V. The distribution of unsaturated acyl groups in vegetable triglycerides. J. Am. Oil. Chem. Soc 1965, 42, 965–970. [Google Scholar]
- Biermann, U; Friedt, W; Lang, S; Lühs, W; Machmüller, G; Metzger, JO; Rüsch gen. Klaas, M; Schäfer, J; Schneider, MP. New syntheses with oils and fats as renewable raw materials for the chemical industry. Angew. Chem. Int. Ed. Engl 2000, 39, 2206–2224. [Google Scholar]
- Hayes, DG. Enzyme-catalyzed modification of oilseed materials to produce eco-friendly products. J. Am. Oil Chem. Soc 2004, 81, 1077–1103. [Google Scholar]
- Gandhi, NN. Applications of lipase. J. Am. Oil Chem. Soc 1997, 74, 621–634. [Google Scholar]
- Albasi, C; Bertrand, N; Riba, JP. Enzymatic hydrolysis of sunflower oil in a standardized agitated tank reactor. Bioprocess Eng 1999, 20, 77–81. [Google Scholar]
- Martinez, O; Wilhelm, AM; Riba, JP. Kinetic-study of an enzymatic liquid liquid reaction – the hydrolysis of tributyrin by Candida cylindracea lipase. J. Chem. Tech. Biotechnol 1992, 53, 373–378. [Google Scholar]
- Habulin, M; Knez, Ž. High-pressure enzymatic hydrolysis of oil. Eur. J. Lipid Sci. Technol 2002, 104, 381–386. [Google Scholar]
- Malcata, FX; Reyes, HR; Garcia, HS; Hill, CG, Jr; Amundson, CH. Immobilized lipase reactions for modification of fats and oils – A review. J. Am. Oil Chem. Soc 1990, 67, 890–910. [Google Scholar]
- Bogel-Lukasik, R. Sustainable processes employing ionic liquids for secondary alcohols separation. Monatsh. Chem 2007, 138, 1137–1144. [Google Scholar]
- Dordick, JS. Nonaqueous enzymology. Curr. Opin. Biotechnol 1991, 2, 401–407. [Google Scholar]
- Reverchon, E; De Marco, I. Supercritical fluid extraction and fractionation of natural matters. J. Supercrit Fluids 2006, 38, 146–166. [Google Scholar]
- Klibanov, AM. Improving enzymes by using them in organic solvents. Nature 2001, 409, 241–246. [Google Scholar]
- Schoffers, E; Golebioeski, A; Johnson, CR. Enantioselective synthesis through enzymatic asymmetrization. Tetrahedron 1996, 52, 3769–3826. [Google Scholar]
- Klibanov, AM. Why are enzymes less active in organic solvents than in water? Trends Biotechnol 1997, 15, 97–101. [Google Scholar]
- Schmid, RD; Verger, R. Lipases: Interfacial enzymes with attractive applications. Angew. Chem. Int. Ed. Engl 1998, 37, 1608–1633. [Google Scholar]
- Beisson, F; Tiss, A; Riviere, C; Verger, R. Methods for lipase detection and assay: A critical review. Eur. J. Lipid Sci. Technol 2000, 133–153. [Google Scholar]
- Krishna, HS; Karanth, NG. Lipases and lipase-catalyzed esterification reactions in nonaqueous media. Catal. Rev 2002, 44, 499–591. [Google Scholar]
- Patel, RN. Microbial/enzymatic synthesis of chiral intermediates for pharmaceuticals. Enzyme Microb. Technol 2002, 31, 804–826. [Google Scholar]
- Patel, RN. Biocatalytic synthesis of chiral pharmaceutical intermediate. Food Technol. Biotechnol 2004, 42, 305–325. [Google Scholar]
- Wong, C-H; Whitesides, GM. Enzymes in Synthetic Organic Chemistry; Pergamon: Oxford, UK, 1994. [Google Scholar]
- Dias, SF; Vilas-Boas, L; Cabral, JMS; Fonseca, MMR. Production of ethyl butyrate by Candida rugosa lipase immobilized in polyurethane. Biocatalysis 1991, 5, 21–34. [Google Scholar]
- Macrae, AR. Biocatalysts in Organic Synthesis; Tramper, J, van der Plas, HC, Linko, P, Eds.; Elsevier: Amsterdam, The Netherlands, 1985. [Google Scholar]
- Muderhwa, JM; Pina, M; Graille, J. Interesterification catalyzed by 1–3 regioselective lipases without solvent-valorization of palm oil and its solid fraction. Rev. Fr. Corps Gras 1989, 36, 11–19. [Google Scholar]
- Kyotani, S; Nakashima, T; Izumoto, E; Fukuda, H. Continuous interesterification of oils and fats using dried fungus immobilized in biomass support particles. J. Ferment. Bioeng 1991, 71, 286–288. [Google Scholar]
- Mojovič, L; Siler-Marinkovič, S; Kukič, G; Vunjak-Novakovič, G. Rhizopus arrhizus lipase-catalyzed interesterification of the midfraction of palm oil to a cocoa butter equivalent fat. Enzyme Microb. Technol 1993, 15, 438–443. [Google Scholar]
- Forssell, P; Paruvuori, P; Linko, P; Poutanen, K. Enzymatic transesterification of rapeseed oil and lauric acid in a continuous reactor. J. Am. Oil Chem. Soc 1993, 70, 1105–1109. [Google Scholar]
- Ghazali, HM; Hamidah, S; Che Man, YB. Enzymatic transesterification of palm olein with nonspecific and 1,3-specific lipases. J. Am. Oil Chem. Soc 1995, 72, 633–639. [Google Scholar]
- Soumanou, MM; Bornscheuer, UT; Menge, U; Schmid, RD. Synthesis of structured triglycerides from peanut oil with immobilized lipase. J. Am. Oil Chem. Soc 1997, 74, 427–433. [Google Scholar]
- Ghosh, S; Bhattacharyya, DK. Utilization of high-melting palm stearin in lipase-catalyzed interesterification with liquid oils. J. Am. Oil Chem. Soc 1997, 74, 589–592. [Google Scholar]
- Mukesh, D; Banerji, AA; Newadkar, R; Bevinakatti, HS. Lipase catalysed transesterification of vegetable oils - A comparative study in batch and tubular reactors. Biotechnol. Lett 1993, 15, 77–82. [Google Scholar]
- Xu, X; Balchen, S; Høy, C-E; Adler-Nissen, J. Pilot batch production of specific-structured lipids by lipase-catalyzed interesterification: Preliminary study on incorporation and acyl migration. J. Am. Oil Chem. Soc 1998, 75, 301–308. [Google Scholar]
- Seriburi, V; Akoh, CC. Enzymatic transesterification of triolein and stearic acid and solid fat content of their products. J. Am. Oil Chem. Soc 1998, 75, 511–516. [Google Scholar]
- Mu, H; Xu, X; Høy, C-E. Production of specific-structured triacylglycerols by lipase-catalyzed interesterification in a laboratory-scale continuous reactor. J. Am. Oil Chem. Soc 1998, 75, 1187–1193. [Google Scholar]
- Ferreira-Dias, S; Duarte, CS; Falaschi, V; Marques, SR; Gusmao, JH; da Fonseca, MMR. Stability and Stabilization of Biocatalysts; Ballesteros, A, Plou, FJ, Iborra, JL, Halling, P, Eds.; Elsevier: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Fomuso, LB; Akoh, CC. Enzymatic modification of triolein: Incorporation of caproic and butyric acids to produce reduced-calorie structured lipids. J. Am. Oil Chem. Soc 1997, 74, 269–272. [Google Scholar]
- Xu, X; Skands, ARH; Høy, C-E; Mu, H; Balchen, S; Adler-Nissen, J. Production of specific-structured lipids by enzymatic interesterification: Elucidation of acyl migration by response surface design. J. Am. Oil Chem. Soc 1998, 75, 1179–1186. [Google Scholar]
- Miura, S; Ogawa, A; Konishi, H. A rapid method for enzymatic synthesis and purification of the structured triacylglycerol, 1,3-dilauroyl-2-oleoyl-glycerol. J. Am. Oil Chem. Soc 1999, 76, 927–931. [Google Scholar]
- Adachi, S; Okumura, K; Ota, Y; Mankura, M. Acidolysis of sardine oil by lipase to concentrate eicosapentaenoic and docosahexanoic acids in glycerides. J. Ferment. Bioeng 1993, 75, 259–264. [Google Scholar]
- Yamane, T; Suzuki, T; Hoshino, T. Increasing n-3 polyunsaturated fatty acid content of fish oil by temperature control of lipase-catalyzed acidolysis. J. Am. Oil Chem. Soc 1993, 70, 1285–1287. [Google Scholar]
- Li, Z; Ward, OP. Enzyme catalysed production of vegetable oils containing omega-3 polyunsaturated fatty acid. Biotechnol. Lett 1993, 15, 185–188. [Google Scholar]
- Huang, K-H; Akoh, CC. Lipase-catalyzed incorporation of n-3 polyunsaturated fatty acids into vegetable oils. J. Am. Oil Chem. Soc 1994, 71, 1277–1288. [Google Scholar]
- Lee, K-T; Akoh, CC. Immobilized lipase-catalyzed production of structured lipids with eicosapentaenoic acid at specific positions. J. Am. Oil Chem. Soc 1996, 73, 611–615. [Google Scholar]
- Senanayake, SPJN; Shahidi, F. Enzymatic incorporation of docosahexaenoic acid into borage oil. J. Am. Oil Chem. Soc 1999, 76, 1009–1015. [Google Scholar]
- Jennings, BH; Akoh, C. Enzymatic modification of triacylglycerols of high eicosapentaenoic and docosahexaenoic acids content to produce structured lipids. J. Am. Oil Chem. Soc 1999, 76, 1133–1137. [Google Scholar]
- Haraldsson, G; Thorarensen, A. Preparation of phospholipids highly enriched with n-3 polyunsaturated fatty acids by lipase. J. Am. Oil Chem. Soc 1999, 76, 1143–1149. [Google Scholar]
- Muderhwa, JM; Pina, M; Montet, D; Feuillard, P; Graille, J. 1–3 Regioselective enzymatic interesterification in a melted medium and a continuous reactor – valorization of palm oil. Oleagineux 1989, 44, 36–43. [Google Scholar]
- Zainal, Z; Yusoff, MSA. Enzymatic interesterification of palm stearin and palm kernel olein. J. Am. Oil Chem. Soc 1999, 76, 1003–1008. [Google Scholar]
- Bergstrom, B; Brochmann, HL. Lipases, 1st Ed, Elsevier: Amsterdam, The Netherlands, 1984.
- Quinlan, P; Moore, S. Modification of triglycerides by lipases: Process technology and its application to the production of nutritionally improved fats. INFORM 1993, 4, 580–585. [Google Scholar]
- Brady, L; Brzozowski, AM; Derewenda, ZS; Dodson, E; Dodson, G; Tolley, S; Turkenburg, JP; Christiansen, L; Huge-Jensen, B; Norskov, L; Thim, L; Menge, U. A serine protease triad forms the catalytic centre of a triacylglycerol lipase. Nature 1990, 343, 767–770. [Google Scholar]
- Klibanov, AM. Why are enzymes less active in organic solvents than in water? Trends Biotechnol 1997, 15, 97–101. [Google Scholar]
- Pleiss, J; Fischer, M; Peiker, M; Thiele, C; Schmid, RD. Lipase engineering database. Understanding and exploiting sequence–structure–function relationships. J. Mol. Catal. B: Enzym 2000, 10, 491–508. [Google Scholar]
- Balcao, VM; Paiva, AL; Malcata, FX. Review. Bioreactors with immobilized lipases: State of the art. Enzyme Microb. Technol 1996, 18, 392–416. [Google Scholar]
- Deng, HT; Xu, ZK; Huang, XJ; Wu, J; Seta, P. Adsorption and activity of Candida rugosa lipase on polypropylene hollow fiber membrane modified with phospholipid analogous polymers. Langmuir 2004, 20, 10168–10173. [Google Scholar]
- Homquist, M; Norin, M; Hult, K. The role of arginines in stabilizing the active open-lid conformation of Rhizomucor meihei lipase. Lipids 1993, 28, 721–726. [Google Scholar]
- Villeneuve, P; Muderwha, J; Graille, J; Haas, M. Customizing lipases for biocatalysis: A survey of chemical, physical and molecular biological approaches. J. Mol. Catal. B: Enzym 2000, 9, 113–148. [Google Scholar]
- Holmquist, M; Martinelle, M; Berglund, P; Clausen, IG; Patkar, S; Svendsen, A; Hult, K. Lipases from Rhizomucor miehei and Thermomyces lanuginosa: modification of the lid covering the active site alters enantioselectivity. J. Protein Chem 1993, 12, 749–757. [Google Scholar]
- Gutierrez-Ayesta, C; Carelli, AA; Ferreira, ML. Relation between lipase structures and their catalytic ability to hydrolyse triglycerides and phospholipids. Enzyme Microb. Technol 2007, 41, 35–43. [Google Scholar]
- Pleiss, J; Fisher, M; Schmid, RD. Anatomy of lipase binding sites: the scissile fatty acid binding site. Chem. Phys. Lipids 1998, 93, 67–80. [Google Scholar]
- Bottino, NR; Vandenburg, GA; Reiser, R. Resistance of certain long chain polyunsaturated fatty acids of marine oils to pancreatic lipase hydrolysis. Lipids 1967, 2, 489–493. [Google Scholar]
- Jensen, RG; Gordon, DT. Specificity of Geotrichum candidum lipase with respect to double bond position in triglycerides containing cis-octadecenoic acids. Lipids 1972, 7, 738–741. [Google Scholar]
- Lopez-Martinez, JC; Campra-Madrid, P; Ramirez-Fajardo, A; Esteban-Cerdan, L; Guil-Guerrero, JL. Screening of lipases for enzymatic concentration of gamma-linolenic acid (GLA) from seed oils. J. Food Lipids 2006, 13, 362–374. [Google Scholar]
- Hoshino, T; Yamane, T; Shimizu, S. Selective hydrolysis of fish oil by lipase to concentrate n-3 polyunsaturated fatty acid. Agric. Biol. Chem 1990, 54, 1459–1467. [Google Scholar]
- Tanaka, Y; Hirano, J; Funada, T. Concentration of docosahexaenoic acid glyceride by hydrolysis of fish oil with Candida cylindracea lipase. J. Am. Oil Chem. Soc 1992, 69, 1210–1214. [Google Scholar]
- Basheer, S; Plat, D. Enzymatic modification of sterols using sterol-specific lipase. WO patent 2001-075083, 2001. [Google Scholar]
- Weber, N; Weikamp, P; Mukherjee, KD. Cholesterol-lowering food additives: Lipase-catalyzed preparation of phytosterol and phytostanol esters. Food Res. Intl 2002, 35, 177–181. [Google Scholar]
- Bertinotti, A; Carrea, G; Ottolina, G; Riva, S. Regioselective esterification of polyhydroxylated steroids by Candida antarctica lipase B. Tetrahedron 1994, 50, 13165–13172. [Google Scholar]
- Norinobu, S; Senoo, N; Kaneko, S; Sato, F; Mankura, M. Supercritical preparation of sterol fatty esters with enzyme. Japanese patent 2002-233396, 2002. [Google Scholar]
- Stamatis, H; Sereti, V; Kolisis, FN. Studies on the enzymatic synthesis of lipophilic derivatives of natural antioxidants. J. Am. Oil Chem. Soc 1999, 76, 1505–1510. [Google Scholar]
- Jessop, PG; Ikariya, T; Noyori, R. Homogenous catalysis in supercritical fluids. Chem. Rev 1999, 99, 475–494. [Google Scholar]
- Brennecke, JF; Chateauneuf, JE. Homogenous organic reactions as mechanistic probes in supercritical fluids. Chem. Rev 1999, 99, 433–452. [Google Scholar]
- Baiker, A. Supercritical fluids in heterogenous catalysis. Chem. Rev 1999, 99, 453–474. [Google Scholar]
- Darr, JA; Poliakoff, M. New directions in inorganic and metal-organic coordination chemistry in supercritical fluids. Chem. Rev 1999, 99, 495–542. [Google Scholar]
- Kiran, E; Debenedetti, PG; Peters, CJ. Supercritical Fluids Fundamentals and Applications; Kluwer Academic: Dordrecht, The Netherlands, 2000. [Google Scholar]
- Leitner, W. Supercritical carbon dioxide as a green reaction medium for catalysis. Acc. Chem. Res 2002, 35, 746–756. [Google Scholar]
- Matsuda, T; Watanabe, K; Harada, T; Nakamura, K. Enzymatic reactions in supercritical CO2: carboxylation, asymmetric reduction and esterification. Catal. Today 2004, 96, 103–111. [Google Scholar]
- Kragl, U; Eckstein, M; Kraftzik, N. Biocatalytic reactions in ionic liquids. In Ionic Liquids in Synthesis; Wasserscheid, P, Welton, T, Eds.; Wiley-VCH: Weinheim, Germany, 2003; pp. 336–347. [Google Scholar]
- Seddon, KR. Ionic liquids for clean technology. J. Chem. Technol. Biotechnol 1997, 68, 351–356. [Google Scholar]
- Welton, T. Room-temperature ionic liquids. Solvents for synthesis and catalysis. Chem. Rev 1999, 99, 2071–2084. [Google Scholar]
- Yang, Z; Pan, W. Ionic liquids: Green solvents for nonaqueous biocatalysis. Enzyme Microb. Technol 2005, 37, 19–28. [Google Scholar]
- Chowdhury, S; Mohan, RS; Scott, JL. Reactivity in ionic liquids. Tetrahedron 2007, 63, 2363–2389. [Google Scholar]
- Freemantle, M. Ionic liquids in organic synthesis. Chem. Eng. News 2004, 82, 44–49. [Google Scholar]
- Roberts, NJ; Seago, A; Carey, JS; Freer, R; Preston, C; Lye, GJ. Lipase catalysed resolution of the Lotrafiban intermediate 2,3,4,5-tetrahydro-4-methyl-3-oxo-1H-1,4-benzodiazepine-2-acetic acid methyl ester in ionic liquids: Comparison to the industrial t-butanol process. Green Chem 2004, 6, 475–482. [Google Scholar]
- Lau, RM; Sorgedrager, MJ; Carrea, G; van Rantwijk, F; Secundo, F; Sheldon, RA. Dissolution of Candida antarctica lipase B in ionic liquids: effects on structure and activity. Green Chem 2004, 6, 483–487. [Google Scholar]
- Oliveira, JV; Oliveira, D. Kinetics of the enzymatic alcoholysis of palm kernel oil in supercritical CO2. Ind. Eng. Chem. Res 2000, 39, 4450–4454. [Google Scholar]
- Kamat, SV; Beckman, EJ; Russell, AJ. Enzyme-activity in supercritical fluids. Crit. Rev. Biotechnol 1995, 15, 41–71. [Google Scholar]
- Connstein, W; Hoyer, E; Wartenberg, H. The enzymic decomposition of fat. Chem. Ber 1902, 34, 3988–4006. [Google Scholar]
- Baumann, H; Bühler, M; Fochem, H; Hirsinger, F; Zoebelein, H; Falbe, J. Natural fats and oils - renewable raw materials for the chemical industry. Angew. Chem. Int. Ed. Engl 1988, 27, 41–62. [Google Scholar]
- Moquin, PHL; Temelli, F. Production of monoolein from oleic acid and glycerol in supercritical carbon dioxide media: a kinetic approach. J. Supercrit. Fluids 2008, 44, 40–47. [Google Scholar]
- Promožič, M; Habulin, M; Knez, Ž. Parameter optimization for enzymic hydrolysis of sunflower oil in high-pressure reactors. J. Am. Oil Chem. Soc 2003, 80, 643–646. [Google Scholar]
- Promožič, M; Habulin, M; Knez, Ž. Thermodynamic properties of the enzymatic hydrolysis of sunflower oil in high-pressure reactors. J. Am. Oil Chem. Soc 2003, 80, 785–788. [Google Scholar]
- Promožič, M; Habulin, M; Knez, Ž. Modeling of kinetics for the enzymatic hydrolysis of sunflower oil in a high-pressure reactors. J. Am. Oil Chem. Soc 2005, 82, 543–547. [Google Scholar]
- Zarevúcka, M; Vacek, M; Wimmer, Z; Stránský, K; Koutek, B; Demnerová, K. Enzymic transformations of blackcurrant oil: enrichment with γ-linolenic acid and α-linolenic acid. Chem. Listy 2003, 97, 206–213. [Google Scholar]
- Sovová, H; Zarevúcka, M. Lipase-catalysed hydrolysis of blackcurrant oil in supercritical carbon dioxide. Chem. Eng. Sci 2003, 58, 2339–2350. [Google Scholar]
- Bártlová, M; Bernášek, P; Sýkora, J; Sovová, H. HPLC in reversed phase mode: Tool for investigation of kinetics of blackcurrant seed oil lipolysis in supercritical carbon dioxide. J. Chromatogr. B 2006, 839, 80–84. [Google Scholar]
- Sýkora, J; Bernášek, P; Zarevúcka, M; Kurfürst, M; Sovová, H; Schraml, J. High-performance liquid chromatography with nuclear magnetic resonance detection - A method for quantification of α- and γ-linolenic acids in their mixtures with free fatty acids. J. Chromatogr. A 2007, 1139, 152–155. [Google Scholar]
- Kosugi, Y; Azuma, N. Synthesis of triacylglycerol from polyunsaturated fatty acid by immobilized lipase. J. Am. Oil Chem. Soc 1994, 71, 1397–1403. [Google Scholar]
- Cerdán, LE; Medina, AR; Giménez Giménez, A; Ibánez González, MJ; Molina Grima, E. Synthesis of PUFA enriched triglycerides by lipase catalyzed esterification. J. Am. Oil Chem. Soc 1998, 75, 1329–1337. [Google Scholar]
- Medina, AR; Cerdán, LE; Giménez Giménez, A; Camacho Páez, B; Ibánez González, MJ; Molina Grima, E. Lipase-catalyzed esterification of glycerol and polyunsaturated fatty acids from fish and microalgae oils. J. Biotechnol 1999, 70, 379–391. [Google Scholar]
- Akoh, CC; Moussata, CO. Lipase-catalyzed modification of borage oil: Incorporation of capric and eicosapentaenoic acids to form structured lipids. J. Am. Oil Chem. Soc 1998, 75, 697–701. [Google Scholar]
- McKenna, MC; Hubbard, VS; Aeri, JG. Linoleic acid absorption for lipid supplements in patients with cystic fibrosis with pancreatic insufficiency and in control subjects. J. Pediatr. Gastroenterol. Nutr 1985, 4, 45–48. [Google Scholar]
- Kawashima, A; Shimada, Y; Nagao, T; Ohara, A; Matsuhisa, T; Sugihara, A; Tominaga, Y. Production of structured TAG rich in 1,3- dicapryloyl-2-c-linolenoyl glycerol from borage oil. J. Am. Oil Chem. Soc 2002, 79, 871–877. [Google Scholar]
- Kawashima, A; Shimada, Y; Yamamoto, M; Sugihara, A; Nagao, T; Komemushi, S; Tominaga, Y. Enzymatic synthesis of high-purity structured lipids with caprylic acid at 1,3-positions and polyunsaturated fatty acid at 2-position. J. Am. Oil Chem. Soc 2001, 78, 611–616. [Google Scholar]
- Senanayake, SPJN; Shahidi, F. Chemical and stability of structured lipids from borage (Borago officinalis L.) and evening primrose (Oenothera biennis L.) oils. J. Food Sci 2002, 67, 2038–2045. [Google Scholar]
- Shimada, Y; Suenaga, M; Sugihara, A; Seiichi, S; Tominaga, Y. Continuous production of structured lipids containing clinolenic and caprylic acids by immobilizedRhizopus delemar lipase. J. Am. Oil Chem. Soc 1996, 76, 189–193. [Google Scholar]
- Hamam, F; Shahidi, F. Synthesis of structured lipids via acidolysis of docosahexaenoic acid single cell oil (DHASCO) with capric acid. J. Agric. Food Chem 2004, 52, 2900–2906. [Google Scholar]
- Hamam, F; Shahidi, F. Enzymatic acidolysis of arachidonic acid single cell oil (ARASCO) with capric acid. J. Am. Oil Chem. Soc 2004, 81, 887–892. [Google Scholar]
- Hamam, F; Shahidi, F. Enzymatic incorporation of capric acid into a single cell oil rich in docosahexaenoic acid (DHA) and docosapentaenoic acid (DPA). Food Chem 2005, 91, 583–591. [Google Scholar]
- Iwasaki, Y; Han, JJ; Narita, M; Rosu, R; Yamane, T. Enzymatic synthesis of structured lipids from single cell oil of high docosahexaenoic acid content. J. Am. Oil Chem. Soc 1999, 76, 563–569. [Google Scholar]
- Yang, T; Xu, X; Li, L. Comparison of linoleic and conjugated linoleic acids in enzymatic acidolysis of tristearin. J. Food Lipids 2001, 8, 149–161. [Google Scholar]
- Ronne, TH; Pederson, LS; Xu, X. Triglyceride selectivity of immobilized Thermomyces lanuginose lipase in interesterification. J. Am. Oil Chem. Soc 2005, 82, 737–743. [Google Scholar]
- Tsuzuki, W. Acidolysis between triolein and short-chain fatty acids by lipase in organic solvents. Biosci. Biotechnol. Biochem 2005, 69, 1256–1261. [Google Scholar]
- Paez, BC; Medina, AR; Rubio, FC; Cerdan, LE; Grima, EM. Kinetics of lipase-catalyzed interesterification of triolein and caprylic acid to produce structured lipids. J. Chem. Technol. Biotechnol 2003, 78, 461–470. [Google Scholar]
- Sellappan, S; Akoh, CC. Synthesis of structured lipids by transesterification of trilinolein catalyzed by Lipozyme IM60. J. Agric. Food Chem 2001, 49, 1–9. [Google Scholar]
- Fomus, LB; Akoh, CC. Structured lipids: Lipase-catalyzed interesterification of tricaproin and trilinolein. J. Am. Oil Chem. Soc 1998, 75, 405–410. [Google Scholar]
- Hamam, F; Shahidi, F. Incorporation of selected long-chain fatty acids into trilinolein and trilinolenin. Food Chem 2008, 106, 33–39. [Google Scholar]
- Hamam, F; Shahidi, F. Enzymatic incorporation of selected long-chain fatty acids into triolein. J. Am. Oil Chem. Soc 2007, 84, 533–541. [Google Scholar]
- Hamam, F; Shahidi, F. Acidolysis of tristearin with selected long chain fatty acids. J. Agric. Food Chem 2007, 55, 1955–1960. [Google Scholar]
- Lee, K; Akoh, C. Characterization of enzymatically synthesized structured lipids containing eicosapentaenoic, docosahexaenoic, and caprylic acids. J. Am. Oil Chem. Soc 1998, 75, 495–499. [Google Scholar]
- McNeill, GP; Ackman, RG; Moore, SR. Lipase-catalyzed enrichment of long chain polyunsaturated fatty acids. J. Am. Oil Chem. Soc 1996, 73, 1403–1414. [Google Scholar]
- Schmid, U; Bornscheuer, UT; Soumanou, MM; Mc- Neill, GP; Schmid, RD. Optimization of the reaction conditions in the lipase-catalyzed synthesis of structured triglycerides. J. Am. Oil Chem. Soc 1998, 75, 1527–1531. [Google Scholar]
- Soumanou, MM; Bornscheuer, UT; Schmid, RD. Two step enzymatic reaction for synthesis of pure structured triacylglycerides. J. Am. Oil Chem. Soc 1998, 75, 703–710. [Google Scholar]
- Tsujisaka, Y; Okumura, S; Iwai, M. Glyceride synthesis by four kinds of microbial lipase. Biochim. Biophys. Acta 1977, 489, 415–422. [Google Scholar]
- Okumura, S; Iwai, M; Tsujisaka, Y. Synthesis of various kinds of esters by four microbial lipases. Biochim. Biophys. Acta 1979, 575, 156–165. [Google Scholar]
- Liu, K-J; Chang, H-M; Liu, K-M. Enzymatic synthesis of cocoa butter analog through interesterification of lard and tristearin in supercritical carbon dioxide by lipase. Food Chem 2007, 100, 1303–1311. [Google Scholar]
- Sakurai, H; Pokorný, J. The development and application of novel vegetable oils tailor-made for specific human dietary needs. Eur. J. Lipid Sci. Technol 2003, 105, 769–778. [Google Scholar]
- Jessop, P; Leitner, W. Chemical synthesis using supercritical fluids. Wiley-VCH: Weinheim, 1999. [Google Scholar]
- King, JW; Snyder, JM; Frykman, H; Neese, A. Sterol ester production using lipase-catalyzed reactions in supercritical carbon dioxide. Eur. Food Res. Technol 2001, 212, 566–569. [Google Scholar]
- Gunnlaugsdottir, H; Järemo, M; Sivik, B. Process parameters influencing ethanolysis of cod liver oil in supercritical carbon dioxide. J. Supercrit. Fluids 1998, 12, 85–93. [Google Scholar]
- Overmeyer, A; Schrader-Lippelt, S; Kasche, V; Brunner, G. Lipase-catalysed kinetic resolution of racemates at temperatures from 40 °C to 160 °C in supercritical CO2. Biotechnol. Lett 1999, 21, 65–69. [Google Scholar]
- Habulin, M; Krmelj, V; Knez, Ž. Synthesis of oleic acid esters catalyzed by immobilized lipase. J. Agric. Food Chem 1996, 44, 338–342. [Google Scholar]
- Knez, Ž; Habulin, M. Compressed gases as alternative enzymatic-reaction solvents: A short review. J. Supercrit. Fluids 2002, 23, 29–42. [Google Scholar]
- Leitner, W. Die “bessere Lösung”? Chemische Synthese in überkritischem Kohlendioxid. Chem. Unserer Zeit 2003, 37, 32–38. [Google Scholar]
- King, J; Jackson, M; Temelli, F. Development of industrially-useful synthetic processes in supercritical carbon dioxide. In I Fluidi Supercritici e le Lorno Applicazioni; Kikic, I, Alessi, P, Eds.; Trieste: Italy, 1995; pp. 19–26. [Google Scholar]
- Bornscheuer, U; Kazlauskas, R. Hydrolases in Organic Synthesis: Regio- and Stereoselective Biotransformations; Wiley-VCH: Weinheim, Germany, 1999. [Google Scholar]
- Snyder, J; King, J; Jackson, M. Fat content for nutritional labeling by supercritical fluid extraction and an on-line lipase catalyzed reaction. J. Chromatogr. A 1996, 750, 201–207. [Google Scholar]
- Anderson, M; King, J; Blomberg, L. Synthesis of fatty alcohol mixtures from oleochemicals in supercritical fluids. Green Chem 2000, 2, 230–234. [Google Scholar]
- Nakaya, H; Nakamura, K; Miyawaki, O. Lipase-catalyzed esterification of stearic acid with ethanol, and hydrolysis of ethyl stearate, near the critical point in supercritical carbon dioxide. J. Am. Oil Chem. Soc 2002, 79, 23–27. [Google Scholar]
- Ikushima, Y; Saito, N; Arai, M; Blanch, HW. Activation of lipase triggered by interactions with supercritical carbon dioxide in the near-critical region. J. Phys. Chem 1995, 99, 8941–8944. [Google Scholar]
- Srivastava, S; Madras, G; Modak, J. Esterification of myristic acid in supercritical carbon dioxide. J. Supercrit. Fluids 2003, 27, 55–64. [Google Scholar]
- Gunnlaugsdottir, H; Wannerberger, K; Sivik, B. Alcoholysis and glyceride synthesis with immobilized lipase on controlled-pore glass of varying hydrophobicity in supercritical carbon dioxide. Enzyme Microb. Technol 1998, 22, 360–367. [Google Scholar]
- Laudani, CG; Habulin, M; Primožič, M; Knez, Ž; Della Porta, G; Reverchon, E. Optimisation of n-octyl oleate enzymatic synthesis over Rhizomucor miehei lipase. Bioprocess Biosyst. Eng 2006, 29, 119–127. [Google Scholar]
- Laudani, CG; Habulin, M; Knez, Ž; Della Porta, G; Reverchon, E. Immobilized lipase-mediated long-chain fatty acid esterification in dense carbon dioxide: bench-scale packed-bed reactor study. J. Supercrit. Fluids 2007, 41, 74–81. [Google Scholar]
- Pomier, E; Galy, J; Paolucci-Jeanjean, D; Pina, M; Sarrade, S; Rios, GM. A new reactor design combining enzyme, membrane and SC CO2: Application to castor oil modification. J. Membrane Sci 2005, 249, 127–132. [Google Scholar]
- Liu, K-J; Liu, K-M; Chang, H-M. Biocatalytic synthesis of palmitoyl vanillylamide in supercritical carbon dioxide through amidation of vanillylamide hydrochloride and palmitic anhydride by lipase. Food Chem 2007, 102, 1020–1026. [Google Scholar]
- Hakoda, M; Shiragami, N; Enomoto, A; Nakamura, K. Effect of moisture on enzymatic reaction in supercritical carbon dioxide. Bioprocess Biosyst. Eng 2002, 24, 355–361. [Google Scholar]
- Hlavsová, K; Wimmer, Z; Xanthakis, E; Bernášek, P; Sovová, H; Zarevúcka, M. Lipase activity enhancement by SC-CO2 treatment. Z. Naturforsch 2008, 63B, 779–784. [Google Scholar]
- Eisenmenger, M; Dunford, NT; Eller, F; Tailor, S; Martinez, J. Pilot-scale supercritical carbon dioxide extraction and fractionation of wheat germ oil. J. Am. Oil Chem. Soc 2006, 83, 863–868. [Google Scholar]
- Lau, HLN; Choo, YM; Ma, AN; Chuah, CH. Quality of residual oil from palm-pressed mesocarp fiber (Elaeis guineensis) using supercritical CO2 with and without ethanol. J. Am. Oil Chem. Soc 2006, 83, 893–898. [Google Scholar]
- Üstündag, ÖG; Temelli, F. Column fractionation of canola oil deodorizer distillate using supercritical carbon dioxide. J. Am. Oil Chem. Soc 2007, 84, 953–961. [Google Scholar]
- Nagesha, GK; Manohar, B; Sankar, KU. Enzymatic esterification of free fatty acids of hydrolyzed soy deodorizer distillate in supercritical carbon dioxide. J. Supercrit. Fluids 2004, 32, 137–145. [Google Scholar]
- Frykman, HB; Snyder, JM; King, JW. Screening catalytic lipase activities with an analytical supercritical fluid extractor. J. Am. Oil Chem. Soc 1998, 75, 517–520. [Google Scholar]
- Hernández, FJ; de los Ríos, AP; Gómez, D; Rubio, M; Víllora, G. A new recirculating enzymatic membrane reactor for ester synthesis in ionic liquid/supercritical carbon dioxide biphasic systems. Appl. Catal. B: Environ 2006, 67, 121–126. [Google Scholar]
- Gumí, T; Paolucci-Jeanjean, M; Belleville, M-P; Rios, GM. Enzymatic membrane reactor involving a hybrid membrane in supercritical carbon dioxide. J. Membrane Sci 2007, 297, 98–103. [Google Scholar]
- Wang, L; Weller, CL. Recent advances in extraction of nutraceuticals from plants. Trends Food Sci. Technol 2006, 17, 300–312. [Google Scholar]
- Ghanem, A. Trends in lipase-catalyzed asymmetric access to enantiomerically pure/enriched compounds. Tetrahedron 2007, 63, 1721–1754. [Google Scholar]
- Jurček, O; Wimmerová, M; Wimmer, Z. Selected chiral alcohols: Enzymic resolution and reduction of convenient substrates. Coord. Chem. Rev 2008, 252, 767–781. [Google Scholar]





© 2008 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/). This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zarevúcka, M.; Wimmer, Z. Plant Products for Pharmacology: Application of Enzymes in Their Transformations. Int. J. Mol. Sci. 2008, 9, 2447-2473. https://doi.org/10.3390/ijms9122447
Zarevúcka M, Wimmer Z. Plant Products for Pharmacology: Application of Enzymes in Their Transformations. International Journal of Molecular Sciences. 2008; 9(12):2447-2473. https://doi.org/10.3390/ijms9122447
Chicago/Turabian StyleZarevúcka, Marie, and Zdeněk Wimmer. 2008. "Plant Products for Pharmacology: Application of Enzymes in Their Transformations" International Journal of Molecular Sciences 9, no. 12: 2447-2473. https://doi.org/10.3390/ijms9122447
APA StyleZarevúcka, M., & Wimmer, Z. (2008). Plant Products for Pharmacology: Application of Enzymes in Their Transformations. International Journal of Molecular Sciences, 9(12), 2447-2473. https://doi.org/10.3390/ijms9122447