Illustration of the Structure of Arthropod Assemblages (Collembola and Lepidoptera) in Different Forest Types: An Example in the French Pyrenees

Abstract
: To analyze the impact of management choices on diversity in Pyrenean forests, we selected two ecological indicators: springtails; indicators of long-term responses to perturbation, and moths; which respond quickly to changes in their environment. Our data show that monoculture has a short-term impact on overall diversity and richness of species but with a relative resilience capacity of the forest ecosystem. More precisely, real impacts are visible on dynamics and abundances of certain species, depending on the vertical distribution of the biota and on the composition of soil and forest floor.1. Introduction
European forests have been modified by man for centuries. This has led to extensive ecological changes in terms of diversity but also of soil composition. Current forest management practices (mainly plantation) should now incorporate the recent directives stressed at the Eleventh World Forestry Congress (Antalya, Turkey, 1997) and the resolutions of the Third Ministerial Conference on the Protection of Forests in Europe [1].
Different approaches can be applied to plantations: they can be managed as monocultures or as mixed coniferous and deciduous forests, strict woodlands (only trees of the same age and no undergrowth), also called regular forests, or gardening woodlands (trees of different ages and sizes), also called irregular forests. In an economic context, the various currently applied forest management techniques have the same main goal: transforming forests into “tree making factories” [2].
However, in irregular woodland, trees are selected for timber and the rest of the forest resembles a natural ecosystem with trees of different sizes and ages and a vertical structure of the ecosystem [3]. A strict monoculture is obviously less natural but does provide real economic advantages in tree conformation and growth. However, the expansion of these plantations during the last century could account for pest and pathogen increases as has been shown to occur in central European forests [2-4]. Strict monoculture type management is widespread in the French Pyrenees (depending on various local reports around 50% of the total area of this mountain chain).
Some species living in natural forests are habitat-dependent and can disappear in managed forests. Such losses are difficult to estimate as too few natural reference sites actually exist. Moreover, climatic changes can also influence biodiversity through glaciations, for example [5], which influenced the occurrence of endemic species in alpine chains such as the Pyrenees or the Appalachians [6]. It is enlightening to analyze differences in the impact of forestry treatment between sub-natural (formerly exploited left to reach the climax stage [7] in a natural way) and currently exploited forests. We have to consider endemic or endangered species and must manage to conserve them as laid down in resolutions L2 focusing on endemic species (criteria 4.1b) and vertical structure of forests (criteria 4.2c) [1] and V4: “Importance of forests for biological diversity” [8].
As the disastrous impact of exotic plantations on endemic species has already been clearly shown [9-11], we limited the present study to the impact of forestry practices on strictly indigenous trees.
To study the impact on fauna, one difficulty is to choose a relevant indicator. Rules to choose efficient ecological indicators have been proposed, and involve representativeness, sampling efficiency and require knowledge of systematics, [9,10,12-15]. Moreover, indicators should give information that can be interpreted within the context of natural conditions of locality and forest type [16]. We chose two invertebrate groups: springtails (Collembola) and moths (Lepidoptera, Heterocera), their sampling and taxonomy being well known to us [10-17]. Moreover, these groups are complementary in their function in the ecosystem (decomposers vs. herbivores, respectively). Collembola are dominant hexapods in soils [18] and present high levels of endemicity in the Pyrenees [9]. Lepidoptera are extremely sensitive [19] and respond quickly to changes in habitat regarding botanical composition, fragmentation and vertical structure, [17,20-23]. They have short generation times, and depend on plant species from high to low dispersal ability and high habitat specificity [24], (Table 1).
We revealed large differences between the climax stage and the more recent plantation sites but with high possibilities of resilience. Resilience was defined as the capacity to continue functioning after a perturbation i.e., the ability to return to a former state after an exogenous disturbance [17-25].
We expected that:
Forest management induces changes in the overall cover of the canopy superimposed with presence/absence of a shrub layer, with consequences on the diversity of herbaceous plants (modification of the vertical structure of the forest) and on the abundance and species richness of butterfly communities, especially concerning strictly forest-dependent species. A diminishing richness of specialized and/or endemic species with increasing management is therefore predicable.
As a consequence, the vertical changes mentioned above induce some modifications in the composition of litter which will modify the species composition of springtail communities, affecting both endemic and specialized species.
Consequently, the combination of these two indicators allows the assessment of forestry impact, at different time scales as demonstrated recently for Collembolas [26] and confirmed in Lepidoptera [27,28] and throughout the biota: from tree canopy to soil including vertical forest structure and composition, from herbivores to decomposers. Finally, for the two groups, some endemic, rare, endangered or protected species are of patrimonial interest and their ability to survive in the context of new forestry should be extremely informative from the conservation point of view.
2. Material and Methods
2.1. Study Sites
Two forests differing only in management approach were chosen. They were situated near each other in the mountainous level of vegetation (1300–1450 m), near Campuls Cirque, Bethmale (Central Pyrenees, France).
The sites were exactly 940m apart and separated by a ridge and a stretch of sub-alpine grassland. They had exactly the same exposure slope (deviation from geographic North: plantation: −2°9′15″ and Sub-natural forest: −2°9′41″), topography (slopes identical: 25%), same climate and soil type (pH 4–5, relative humidity 55–75% and C/N = 20).
The two sites were: (i) an irregular/sub-natural forest with Abies alba Mill. and Fagus sylvatica L., which had not been logged for at least 150 years (Latitude 42°51.286′N, longitude 1°3.504′E, Elevation 1,353 m); (ii) a regular/plantation of A. alba about 40 years old, planted after cut of a natural forest of F. sylvatica (L. 42°51.711′ N, 1°3.917′ E, Elevation 1,398 m) [10-17].
Vegetation from these sites is described in connection with springtails, in terms of litter, and moths, in terms of host plants [17-23]. The lists given are not exhaustive but contain the indications necessary to illustrate plant-insect relationships. The floral data have already been published elsewhere [17-23].
2.2. Collecting
2.2.1. Collembola
A total of 40 sample cores were collected in the two forests.
In each forest, four sampling zones were randomly selected. In each zone, five core samples were taken below the forest floor in autumn (1999), the most favorable period for a high diversity of soil fauna [10]. Each core consisted of 250 cm3 of the A horizon taken in the first 10 cm of soil just under the O horizon.
We only considered samples from the A horizon, as its fauna is more directly linked to soil characteristics [29-31] and the A horizon is also less dependent on soil surface heterogeneity.
Arthropods were extracted from soil samples with Berlese-Tullgren funnels at a temperature of 18–23 °C for 7 days. Collembola were identified to species level [32-37] and their abundance recorded.
2.2.2. Moths
Samples were taken using light traps with two 250 W mercury-vapor bulbs connected to a portable generator. Light traps are known to favor the capture of certain species. This can be a problem for obtaining absolute population estimates and for studying moth population dynamics [38], but it is of less concern if the aim is to compare populations through time or between sites [17,23,39,40].
A single sampling point was used for each forest type. Vegetation appeared to be homogeneous around the position of the light traps over a radius of at least 50 m. To avoid biases due to humidity, temperature and moonlight [39] (moonless nights or small new moons were preferred), pair-wise collections were done on the same day, in the two studied sites.
Sampling sessions were carried out during the active period (June–September), once per month over three years (1999–2001). This time span minimizes biases due to annual climatic variations, especially harsh winters that influence most species' pupae.
Moth species determinations were carried out using standard identification handbooks [41-45].
In a former article [17], the same moth database was used but was not processed to analyze the effects of forest management. Some data known for moths will be recalled for comparison with Collembola data.
2.3. Data Treatment
2.3.1. Similarity Index
The Smith-Solow-Preston (SSP) index [46] was calculated with the Matlab Optimisation Toolbox© [47], for data concerning both springtail and moth species, to estimate similarity. Similarity indices were derived from a Bayesian estimate of the Jaccard index, through the use of a delta-beta-binomial model, parameters of which are estimated by a maximum likelihood method. SSP possesses the major advantage of taking into account the number of individuals for each species, yielding a similarity index which includes abundance of each of the species observed. SSP estimates of community species overlap are often 50% larger than Jaccard's non-corrected measure of similarity [46]. The SSP coefficient was estimated using constrained optimization. Constraints defined on both p and q prevent the algorithm from reaching a final solution on non-sense values. The two constraints defined an acceptable range of values for the two parameters (0 < p < 1; 0 < q < 1) outside which the optimization routine was not allowed to wander.
2.3.2. Distance in Species Richness
As this index uses/computes the number of each species present, another measurement is possible during the SSP calculation: Δ = relative distance in species richness between the two communities. As the SSP index gives a realistic overlap between two communities, Δ gives information on the difference in species richness.
2.3.3. Estimation of Diversity
In addition, the Chao 1 index was calculated using EstimateS© 8.20 software [48] (10,000 randomizations). This index was chosen as it gives an estimate of the species richness. Note that Chao 1 can be considered as a minimum value of bulk species richness [49,50]. These results are used in the discussion.
2.4. Endemic Species
The notion of endemic species is dependent on the scale of the study. Some Collembolan species in the Pyrenees possess a very restricted range sometimes limited to a single patch [9-11]. In this study, when speaking of endemic species, we are considering species only present in the central eastern part of the Pyrenees and occurring in the studied type of forest.
2.5. Forest-Specific Species
Lepidoptera are phytophagous insects. Their occurrence directly (but not exclusively) depends on host plant occurrence and possibly density [23]. We assume that the forest community is restricted to moth species having at least one potential host plant in the forest. Species with host plants only on the edge were considered as non-specific (edge management is not necessarily linked to forestry management). Therefore, after determination, species were related to floristic data [17-23] and classified as “forest-dependent species” and all others as “non-forest-dependent species”.
2.6. β Diversity Decomposition and Comparison between Moths and Collembolas
In order to analyze the impact of disturbance on endemic springtail species and moth forest-dependent assemblages, we estimated overall beta diversity (βcc) partitioned by its components: species richness differences (βrich) and species replacement (β−3), following the approach of Carvalho et al. [51]. βcc represent ‘proportional effective species turnover’ [52-53] and correspond to Jaccard dissimilarity measure, whereas β−3 correspond to Williams [54] calculation modified by Cardoso et al. [55]. The index was estimated as:
3. Results and Discussion
3.1. Results
3.1.1. Collembola
A total of 3,962 individuals from 46 species were collected (Table 2).
Thirteen species are endemic with a marked higher abundance in the sub-natural forest, 1,369 individuals vs. 158 in the planted site. The calculation of the estimated difference is only of 3 (Δ = 2.6) additional species occurring in the sub-natural forest (no longer living in the plantation), even though the observed difference is of 7 species missing in the plantation.
Among the 33 non-endemic species, we identified 702 individuals in the sub-natural forest and 1,733 individuals in the plantation. The richness of species belonging to this group was greater in the regular plantation with almost 7 (Δ = 6.6) additional species.
Regarding our Bayesian calculations (Table 3), similarity indices were identical, giving an average of 78% similarity between the two forests independently of the group of species taken into account. Interestingly, the 22% dissimilar species is in favor of the sub-natural forest for endemic species and of the plantation for non-endemic species. A notable difference of 4 species occurred in the relative distance in species richness between endemic and non-endemic species.
3.1.2. Moths
We collected a total of 1,665 moths covering 177 different species. Because the regular forest is a strict fir plantation (with just a few young birch, bramble and bilberry present [23]), it is obvious that many species considered as “forest species” in the sub-natural reference site are also “edge species” for the plantation. In this case, these species were not counted as forest-dependent (Table 4).
Among forest-dependent species, 552 individuals were found: 163 in the planted forest and 389 in the reference sub-natural site; they belonged to 53 different species. A δ value of almost 17 (16.8) in favor of the sub-natural forest is significant with 32% (17/53) of the species found only in this forest.
In the second group, non-forest-dependent species, we found 911 individuals (309 in the regular plantation and 602 in the irregular sub-natural forest) representing 97 species. The δ value is high, 12 (11.6) more species in the sub-natural forest than in the plantation (Table 3).
Bayesian calculations showed a quasi-identical SSP index for the two groups but a third of the species no longer existed in the plantation. The relative difference in species richness was consequently large (almost 5 species) with a higher value for forest-dependent species.
3.2. Comparison of the Decomposition of β Diversity for Collembolas and Moths
βcc values of both springtail assemblages of endemic species and moth forest-dependent species indicate that compositional changes were very similar and relatively high (Table 5). By partitioning these values into βrich and β−3 in the two assemblages we find a similar trend, compositional differences due to richness differences were higher than compositional differences attributable to species replacement. However there are contrasts between communities in the relative contribution of both components of beta-diversity. Whereas in the springtail community the relative contribution of compositional differences determined by species loss accounted for 75% (βrich/βcc), this value is notably higher for moths, 89%. But, when comparing differences determined by replacement (β−3/βcc), the above trend is inverted between collembolas (25%) and moths (11%).
4. Discussion
Losses of plant diversity or capacity of re-colonization, with different types of forest management and with different refuge possibilities, are now well known [58-60]. As proposed by Wülf [61], we now have to evaluate the impact on fauna.
4.1. Collembola
Springtails have little ability to move on the scale of the forest and thus act as indicators of ancient strong forestry impacts [62]. However, they are active participants in litter decomposition and are dependent on tree cover and soil quality and composition [63,64]. Moreover, some endemic species and relict taxa [65] have been described in Pyrenean refuges. For this group we cannot speak about a sociological interest but about a real patrimonial interest.
The endemic community sampled in the sub-natural forest is typical of a community in natural conditions in this type of forest [9-11]. The exact loss of original endemicity is impossible to evaluate here, as no primary forests exist in the area, or in the whole of Europe (except maybe part of the Bialowieża forest in Poland/Belarus). We can just have an idea of what a stable collembolan community could be in the absence of forestry treatment in a mixed forest.
Similarity indices reveal that in the two hexapod communities studied, endemic and non-endemic species show 78% similarity between the two sites. Differences observed between the two forests can be essentially due to the occurrence of beech in the sub-natural forest, which means a different litter/soil (even though some young beeches are present in the plantation, and beech was present in this site 40 years ago). Despite the fact that similarity values are identical between endemic and non-endemic species, they are not equivalent as the 22% difference observed for the sub-natural forest is due to a larger number of endemic species, while in the plantation, it is a combination of loss of some endemic species plus generalist species. Finally the total difference in the number of collembolan species (based on δ values) between the two forests was 9 (6.6 + 2.6 = 9.2). For endemic species, we can note that the value of δ is quite small (2.6 species). Estimation of species richness using the Chao1 index is congruent with the observed richness (13/12 species for sub-natural forest and 6 for plantation). Individual numbers (Table 2) suggest that 7 species are not represented (lost?) in the plantation and one additional species occurs there. The Bayesian approach integrates the idea that while few individuals were collected in the sub-natural forest, their absence from the plantation could in fact result from sampling deficiencies (which would explain the difference between the calculated δ value (2.6) and the observed difference in absolute number of species (7 − 1 = 6)). For example, we cannot confirm that species such as Friesea tolosana are really absent from the plantation or if it is simply rare like in the sub-natural site (1 individual in the sub-natural forest vs. 0 in the plantation). However, for Folsomia sp.1, there is no doubt (384 individuals in the sub-natural forest vs. 0 in the plantation).
Two complementary hypotheses can be put forward to explain this Δ value of 2.6 species (Table 3): (i) the species richness remained relatively constant after commercial forestry; (ii) the ability of collembola to re-colonize sites is low [18-66] as it is restricted to natural recolonization from a sub-natural site (more than a century in the case of the sub-natural site studied here) or artificial reintroduction if species are brought in with planted trees (improbable, even for endemic species as planted trees are generally not grown in natural and local conditions).
For non-endemic taxa, the relative distance in species richness is 6.6 species. For this group, possibilities of re-colonization from other habitats or introduction with transplanted tree soil should be taken into account.
The endemic community sampled in the fir plantation represents the local minimum combination of endemic species present in such forests. This collembolan assemblage can represent the starting point of a possible restructuring of a sub-natural community such as that observed in our “wild” study site. Resilience of collembolan communities is then more dependent on the duration of such strict treatment in the plantation, than on the alternation of “gardened” forests.
Compared to exotic plantations (such as Picea abies in the Pyrenees) [9,10,64,67] the use of indigenous coniferous species creates less disturbance. However, as explained in [68] a less ecosystem-degrading type of forestry practice would be the management of mixed forests with indigenous trees (here firs and beeches) of different ages and sizes.
4.2. Moths
As phytophagous markers, moths were preferred to butterflies (Lepidoptera, Rhopalocera) because the butterfly's ability to fly seems to be primarily determined by solar radiation while moths are more endothermic [69-71]. Butterflies, therefore, have little chance of occurring in temperate forests with a dense canopy although specialized forest butterflies do exist in tropical areas. Moths are able to live and fly in both open and closed habitats. Furthermore, moths are less sensitive to small disturbances of their habitat like partial fragmentation [72].
As with springtails, moths' similarity indices were quite close for the two groups. For non-forest-dependent species the similarity between the two forests was a little higher than for forest-dependent species, which surely means that these species are able to colonize the new ecosystems more quickly than forest-dependent species. An important difference compared to Collembola is that for our two species assemblages, the absolute number of species was higher for the sub-natural forest.
As noted for Collembola communities, the relative distances in species richness values are clearly different. For non-forest-dependent species it can be noted that the δ value is quite high (11.6 species), (Table 3). Estimated species richness using the Chao1 index, is in any case in favor of a sub-estimation/sample of the number of species in the plantation with 9 additional species possible in this forest (Obs: 22/Est.: 31). Anyway, as our sampling was regular (every month) and done on three consecutive years, the estimated average value of forest-dependent species in the plantation is probably optimistic. The difference in the number of species (approximately 12 species) present in the two forests can be assumed to be due to the quasi-absence of deciduous trees in the plantation. In this way, the forest-dependent Δ value (16.8) can be separated into two numbers: around 12 species depending on tree differences and around 5 species really due to other forestry impacts. Two hypotheses can be proposed: (i) a greater effect than the difference in tree composition (deciduous/coniferous) was caused by the real loss in low plant/shrub diversity in addition to differences in the forest structure (no undergrowth due to the strict treatment of the plantation). These differences also lead to highly contrasting habitats with a less diverse floristic composition and an impoverished vertical structure in the strict woodland. As the habitat changes, an impact on habitat-dependent species diversity becomes visible. However, we can assume that in irregular woodland the capacity of moth communities for rapid resilience should increase; (ii) these differences between numbers of species in the two sites are thought to be partly due to the limited ability to move to another habitat type which means that leaving small sub-natural forest sites in the vicinity would be a way to conserve biodiversity.
The most generalist group (non-forest-dependent) is more representative of the capacity of rapid re-colonization depending more on host-plant occurrence than on the forest conformation [73]. However, the non-habitat-dependent species also give information on the re-colonization ability when the forest structure is close to corridors or edges.
4.3. Forest Structure
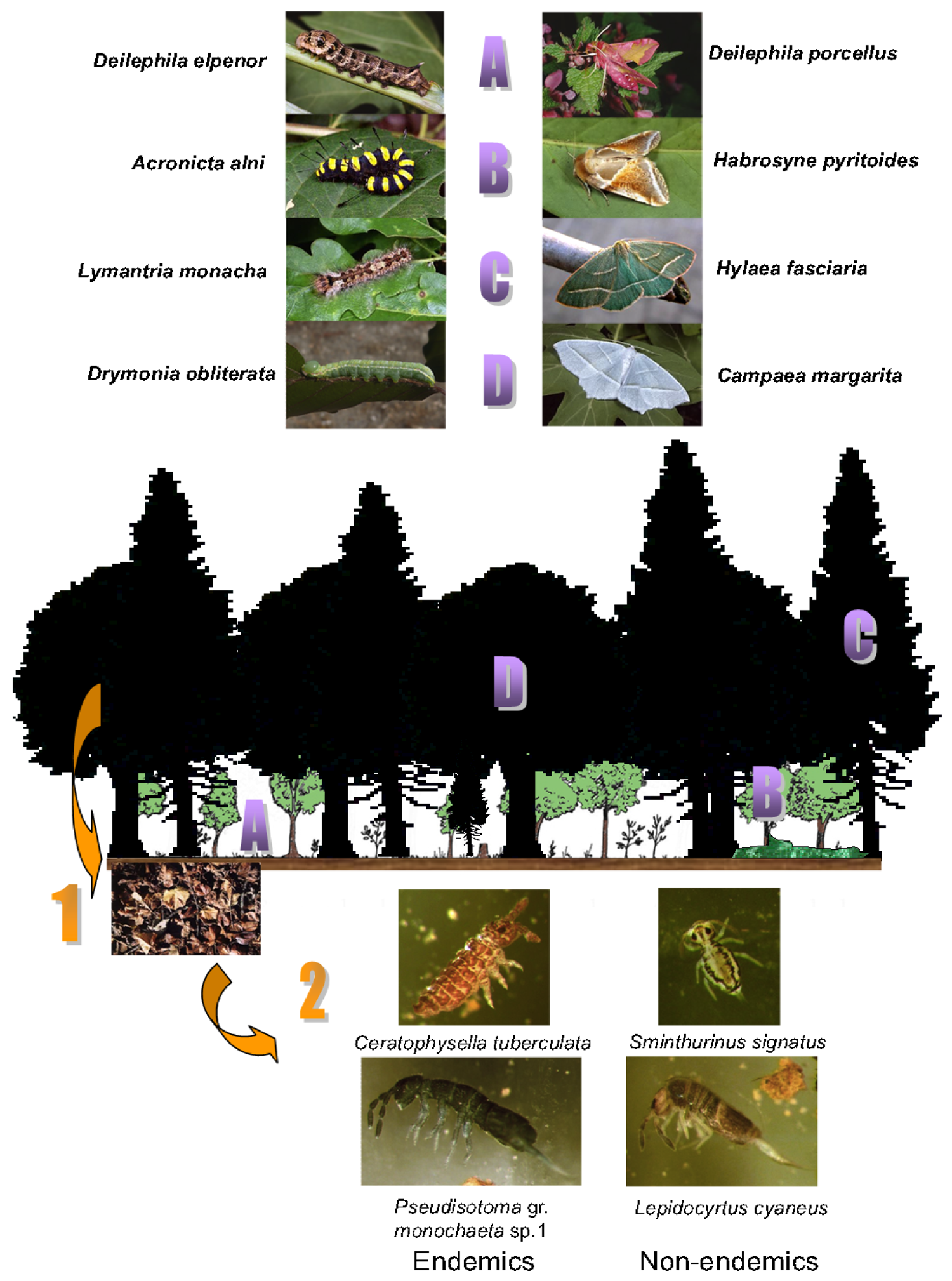
The vertical floristic structure inside the forest is a key point in the present study (Figure 1).
The absence of shrubs has a determinant action on moth assemblages. For example, some species such as Habrosyne pyritoides, Mesoleuca albicillata, and Acronicta auricoma are present only when brambles reach the shrub level (sub-natural forest) and absent (in the plantation) when the brambles stand no taller than herbaceous plants. Some species such as Chloroclysta truncata, living on bilberries are much more abundant in the sub-natural forest when these shrubs are older and higher. Another parameter is the greater mobility of both caterpillars and adults when the horizontal and vertical density of shrubby host-plants is low. During their growth, caterpillars may have to move from one plant to another when the plant density is low and adult females stay close to denser clumps of host plants to lay eggs (leading to lower dispersion).
But these differences in vertical structure not only affect moths as they also cause important changes in litter composition and thus in the physico-chemistry of the soil (porosity, pH, etc.). In the present case, the soil at the two sites could initially be considered as similar, but the input, for 40 years of almost solely fir litter in the plantation then had noticeable effects on collembola assemblages with regression of endemic species and colonization by generalists. A further point of some importance is that below a strict fir forest, most shrubs cannot grow due to the deep shade. This implies that in the plantation all three biodiversity levels studied here are affected: plants, moths and collembola. However, for moths, a resilience capacity is seen in the plantation on the condition that indigenous tree species are planted, and that the edges remain unmanaged [17,27,28]. This resilience may exist for soil characteristics (which affect Collembola) at least, if the duration of the forest management (40 years in our case) is not excessively long. This last assertion should anyway be modulated as these arthropods seem to possess long-term persistence independently of forest management [26].
Analyses of the global beta-diversity show that collembolas and moths respond equally to forest management with similar values around 0.6 showing a marked loss of species. However, when we decomposed the beta-diversity components, effects on species richness were found to be more severe in forest-dependent moth species than in collembolas, which is likely explained by lack of specific host plants and lesser tree coverage. In contrast, in the endemic collembola assemblage there is a higher percentage of species turnover than in the community of moths, suggesting greater resilience (long-term persistence vs. shorter-term persistence, but with higher probabilities, due to dispersion abilities of recolonization by moths). It is important to consider that even when the community of collembola appears more resistant to the change of species (i.e., greater resilience), this does not necessarily mean that the functions of the decomposer food web are left intact in the plantations, since different collembola species may differ from each other in their function within the food web.
5. Conclusions
In this study, we report that, when estimating the effects of the exploitation by man of natural forest landscapes, the use of a single bio-indicator remains useful but a combination of different indicators provides more detailed information on the structure and dynamics of a forest even using mobile organisms such as moths.
A first proposal could be to maintain, independently of the area covered by the managed plot, patches of beech (indigenous deciduous tree) in the plantation forests. However, some rare and endangered species may completely disappear with such management.
A second and preferable proposal would be to limit the size of strictly managed sites and to alternate natural and sub-natural woodland.
Solely based on the present study, which gives a local illustration of the effects of forest management, it is difficult to generalize, but it was shown on a regional scale (Midi-Pyrénées administrative region in France) that a clear positive correlation exists between density of endemic and relictual species of both Collembolas and Lepidopteras and the forest management approach suggested [75].


{kind=link}
| Interests | Springtails | Moths |
|---|---|---|
| Function in ecosystems | Decomposers | Herbivores |
| Interactions | Cover/litter/soil levels | Host-plants/insectivores |
| Sensitivity | Changes in cover or soil quality | Changes in forest structure or host-plants |
| Duration of adaptation or recolonization | Decades to centuries | Seasons to years |
| Conservation and patrimonial interest | Endemic species | Endangered, rare or protected species |
| Taxonomic knowledge and determination scale | Specific | Specific |
| Endemic species | Sub-natural forest | Plantation |
|---|---|---|
| Ceratophysella tuberculata | 30 | 22 |
| Cryptopygus debilis | 639 | 21 |
| Deutonura deficiens | 5 | 4 |
| Folsomia sp.1 | 384 | 0 |
| Friesea tolosana | 1 | 0 |
| Hymenaphorura sp.1 | 1 | 0 |
| Mucrella acuminate | 5 | 0 |
| Onychiurus gr. minutus | 20 | 34 |
| Protachorutes pyreneus | 4 | 0 |
| Pseudisotoma gr. monochaeta sp.1 | 269 | 75 |
| Schaefferia ariegica | 0 | 2 |
| Sminthurinus sp.1 | 8 | 0 |
| Willemia sp.1 | 3 | 0 |
| Observed | 12 | 6 |
| Chao1 | 13 | 6 |
| Springtails | Moths | |||
|---|---|---|---|---|
| Non-endemic | Endemic | Non-forest-dependent | Forest-dependent | |
| SSP | 0.78 | 0.78 | 0.69 | 0.65 |
| Δ | 6.6 | 2.6 | 11.6 | 16.8 |
| Forest dependent species | Sub-natural forest | Plantation | Forest dependent species | Sub-natural forest | Plantation |
|---|---|---|---|---|---|
| Abraxas sylvata | 1 | 0 | Hypena crassalis | 0 | 1 |
| Acronicta alni | 1 | 0 | Hyppa rectilinea | 3 | 0 |
| Acronicta auricoma | 1 | 0 | Idaea contiguaria | 2 | 0 |
| Aethalura punctulata | 2 | 0 | Lacanobia thalassina | 2 | 0 |
| Agrochola circellaris | 1 | 0 | Lymantria monocha | 9 | 1 |
| Alcis jubata | 10 | 6 | Mesoleuca albicillata | 4 | 0 |
| Aplocera praeformata | 16 | 0 | Plagodis dolabraria | 3 | 1 |
| Atolmis rubricollis | 2 | 0 | Plagodis pulveraria | 2 | 0 |
| Cabera pusaria | 1 | 0 | Pseudoips fagana | 12 | 1 |
| Campaea margaritata | 21 | 0 | Pterosoma palpina | 1 | 0 |
| Chloroclysta miata | 1 | 0 | Ptilodon capucina | 9 | 2 |
| Chloroclystis debiliata | 3 | 0 | Pungeleria capreolaria | 4 | 4 |
| Colocasia coryli | 3 | 0 | Selenia tetralunaria | 2 | 0 |
| Cosmia trapezina | 2 | 0 | Semiothisa liturata | 8 | 6 |
| Cyclophora linearia | 21 | 0 | Stauropus fagi | 5 | 2 |
| Diarsia brunnea | 59 | 80 | Thera obeliscata | 2 | 1 |
| Drymomia obliterata | 1 | 0 | Thera variata | 9 | 14 |
| Drymomia querna | 1 | 0 | Thyatira batis | 21 | 1 |
| Ectropis crepuscularia | 14 | 0 | Xanthia aurago | 2 | 4 |
| Eilema complana | 2 | 0 | Xanthia togata | 1 | 1 |
| Eilema lurideola | 5 | 2 | Xanthorhoe designata | 1 | 3 |
| Eilema sororcula | 80 | 0 | Xestia baja | 3 | 1 |
| Eulithis populata | 0 | 8 | Xestia castanea | 1 | 1 |
| Euphyia unangulata | 3 | 0 | Xestia ditrapezium | 5 | 0 |
| Euplexia lucipara | 5 | 6 | Xestia rhomboida | 2 | 0 |
| Gnophos obfuscatus | 2 | 0 | |||
| Habrosyne pyritoides | 4 | 0 | Observed | 51 | 22 |
| Hylaea fasciara | 12 | 19 | Chao1 | 56 | 31 |
| Ecological Indicator | βcc | βrich | β−3 |
|---|---|---|---|
| Springtails: endemic species | 0.61 | 0.46 | 0.15 |
| Moths: forest dependent species | 0.62 | 0.55 | 0.07 |
Acknowledgments
We are very grateful to Marie-Claude Bal-Serin for her help identifying Collembola to the species level and to Louis Deharveng for taxonomic verifications. Thanks to the O.N.F district of Castillon (Ariège) for granting us permission to sample in these forests and for providing information concerning past management choices at these sites.
Appendix
| Non-Endemic species | Non-Endemic species | Non-Endemic species |
|---|---|---|
| Archaphorura sp.1 | Isotomiella cf. paraminor | Paratullbergia callipygos |
| Arrhopalites sp. 1 | Isotomiella minor | Parisotoma notabilis |
| Brachystomella parvula | Isotomurus cf. nebulosus | Proisotoma minuta |
| Ceratophysella armata | Isotomurus prasinus | Protaphorura gr. Armata |
| Deutonura monticola | Lepidocyrtus lanuginosus | Pseudachorutes palmiensis |
| Entomobrya albocincta | Lepidocyrtus cyaneus | Pseudachorutes parvulus |
| Entomobrya sp.1 | Megalothorax incertus | Pseudosinella alba |
| Folsomia manolachei | Megalothorax minimus | Pseudosinella duodecimoculata |
| Folsomia quadrioculata | Mesaphorura gr. Macrochaeta | Sminthurinus signatus |
| Friesea truncate | Micranurida candida | Tomocerus minor |
| Heteromurus major | Oncopodura crassicornis | Vertagopus cinerea |
Appendix
| Non-Forest dependent species | Non-Forest dependent species | Non-Forest dependent species | |
|---|---|---|---|
| Abrostola trigemina | Deilephila porcellus | Mythimna vitellina | |
| Abrostola triplasia | Diacrisia sannio | Nebula tophaceata | |
| Acronicta menyanthidis | Diaphora mendica | Noctua janthina | |
| Acronicta psi | Discestra trifolii | Noctua pronuba | |
| Agrochola cf helvola | Ecliptopera silaceata | Notodonta dromedarius | |
| Agrostis clavis | Ennomos alniaria | Notodonta ziczac | |
| Agrostis ipsilon | Epirrhoe alternata | Nycteola degenerana | |
| Agrostis segetum | Epirrhoe galiata | Odontopera bidentata | |
| Alcis repandata | Epirrhoe rivata | Opisthograptis luteolata | |
| Amphipyra pyramidea | Euchalcia variabilis | Orthosia gothica | |
| Anaplectoides prasina | Eupithecia actaeata | Orthosia stabilis | |
| Apamea crenata | Euxoa nigricans | Pachycnemia hippocastanaria | |
| Apamea maillardi | Euxoa tritici | Panolis flammea | |
| Apamea monoglypha | Hada nana | Paradiarsia glareosa | |
| Arctornis l-nigrum | Heliotis armigera | Peridroma saucia | |
| Autographa bractea | Heliotis peltigera | Phlogophora meticulosa | |
| Autographa gamma | Hoplodrina alsines | Polia nebulosa | |
| Autographa jota | Hoplodrina blanda | Scopula incanata | |
| Autographa pulchrina | Hydriomena furcata | Scopula marginepunctata | |
| Axylia putris | Hypena probiscidalis | Scopula ternata | |
| Biston betularia | Idaea ochrata | Scotopteryx bipunctaria | |
| Blepharita adusta | Lacanobia oleracea | Scotopteryx chenopodiata | |
| Cabera exanthemata | Lampropteryx suffumata | Syngrapha interrogationis | |
| Caradrina morpheus | Lithosia quadra | Tholera cespitis | |
| Ceramica pisi | Lycophotia porphyrea | Triphosa dubitata | |
| Cerapteryx graminis | Lymantria dispar | Xanthorhoe ferrugata | |
| Chloroclysta citrata | Mesapamea secalis | Xanthorhoe fluctuata | |
| Chloroclysta truncata | Mythimna albipuncta | Xanthorhoe montanata | |
| Colostygia pectinataria | Mythimna conigera | Xanthorhoe munitata | |
| Cosmorhoe ocellata | Mythimna ferrago | Xanthorhoe spadicearia | |
| Craniophora ligustri | Mythimna impura | Xestia c-nigrum | |
| Crocota tinctaria | Mythimna unipuncta | Xestia rubi | |
| Deilephila elpenor | |||
References
- General Declarations and Resolutions Adopted at the Third Ministerial Conference in Lisbon, Ministerial Conference on the Protection of Forests in Europe; MCPFE, Lisbon, Portugal; 1998. Available online: http://www.mcpfe.org (accessed on 4 December 2009).
- Failing, L.; Gregory, R. Ten common mistakes in designing biodiversity indicators for forest policy. J. Environ. Manag. 2003, 68, 121–132. [Google Scholar]
- Hartley, M.J. Rationale and methods for conserving biodiversity in plantation forests. For. Ecol. Manag. 2002, 155, 81–95. [Google Scholar]
- Farrell, E.P.; Fuhrer, E.; Ryan, D.; Andersson, F.; Huttl, R.; Piussi, P. European forest ecosystems: building the future on the legacy of the past. For. Ecol. Manag. 2000, 132, 5–20. [Google Scholar]
- Albre, J.; Gers, C.; Legal, L. Molecular phylogeny of the Erebia tyndarus (Lepidoptera, Rhopalocera, Nymphalidae, Satyrinae) species combining CoxII and ND5 mitochondrial genes: A case study of a recent radiation. Mol. Phylogenet. Evol. 2008, 47, 196–210. [Google Scholar]
- Summerville, K.S.; Crist, T.O. Determinants of lepidopteran community composition and species diversity in eastern deciduous forests: Roles of season, eco-region and patch size. Oikos 2003, 100, 134–148. [Google Scholar]
- Ozenda, P. Végétation du Continent Européen; Delachaux and Nieslé: Paris, France, 1994. [Google Scholar]
- MCPFE: Vienna, Austria, 2003. Available online: http://www.mcpfe.org (accessed on 4 December 2009).
- Deharveng, L. Soil collembola diversity, endemism, and reforestation: A case study in the Pyrenees (France). Conserv. Biol. 1996, 10, 74–84. [Google Scholar]
- Cassagne, N.; Bal, M.C.; Gers, C.; Gauquelin, T. Changes in humus properties and Collembolan communities following the replanting of beech forests with spruce. Pedobiologia 2004, 48, 267–276. [Google Scholar]
- Cassagne, N.; Gauquelin, T.; Bal, M.C.; Gers, C. Endemic Collembola, privileged bioindicators of forest management. Pedobiologia 2006, 50, 127–134. [Google Scholar]
- McGeoch, M.A. The selection, testing and application of terrestrial insects as bioindicators. Biol. Rev. 1998, 73, 181–201. [Google Scholar]
- Dale, V.H.; Beyeler, S.C. Challenges in the development and use of ecological indicators. Ecol. Indic. 2001, 1, 3–10. [Google Scholar]
- Büchs, W. Biotic indicators for biodiversity and sustainable agriculture-introduction and background. Agric. Ecosyst. Environ. 2003, 98, 1–16. [Google Scholar]
- Büchs, W. Biodiversity and agri-environmental indicators-general scopes and skills with special reference to the habitat level. Agric. Ecosyst. Environ. 2003, 98, 35–78. [Google Scholar]
- Puumalainen, J.; Kennedy, P.; Folving, S. Monitoring forest biodiversity: A European perspective with reference to temperate and boreal forest zone. J. Environ. Manag. 2003, 67, 5–14. [Google Scholar]
- Luque, C.; Gers, C.; Lauga, J.; Mariano, N.; Wink, M.; Legal, L. Analysis of forestry impacts and biodiversity in two Pyrenean forests through a comparison of moth communities (Lepidoptera, Heterocera). Insect Sci. 2007, 14, 323–338. [Google Scholar]
- Petersen, H.; Luxton, M. A comparative analysis of soil fauna populations and their role in decomposition processes. Oikos 1982, 39, 287–388. [Google Scholar]
- Harper, G.L.; Piyapattanakorn, S.; Goulson, D.; Maclean, N. Isolation of microsatellite markers from the Adonis blue butterfly (Lysandra bellargus). Mol. Ecol. 2000, 9, 1948–1949. [Google Scholar]
- Cappuccino, N.; Martin, M.A. The birch tube-maker Acrobasis betulella in a fragmented habitat: The importance of patch isolation and edges. Oecologia 1997, 110, 69–76. [Google Scholar]
- Zschokke, S.; Dolt, C.; Rusterholz, H.-P.; Oggier, P.; Braschler, B.; Thommen, H.; Lüdin, E.; Erhardt, A.; Baur, B. Short-term responses of plants and invertebrates to experimental small-scale grassland fragmentation. Oecologia 2000, 125, 559–572. [Google Scholar]
- Summerville, K.S.; Boulware, M.J.; Veech, J.A.; Crist, T.O. Spatial variation in species diversity and composition of forest Lepidoptera in eastern deciduous forests of North America. Conserv. Biol. 2003, 17, 1045–1057. [Google Scholar]
- Luque, C.; Legal, L.; Machkour-M'Rabet, S.; Winterton, P.; Gers, C.; Wink, M. Apparent influences of host-plant distribution on the structure and the genetic variability of local populations of purple clay (Diarsa brunea). Biochem. Syst. Ecol. 2009, 37, 6–15. [Google Scholar]
- Hermy, M.; Cornelis, J. Towards a monitoring method and a number of multifaceted and hierarchical biodiversity indicators for urban and suburban parks. Landsc. Urban Plan. 2000, 49, 149–162. [Google Scholar]
- Loranger, G.; Bandyopadhyaya, I.; Razaka, B.; Ponge, J.F. Does soil acidity explain altitudinal sequences in collembolan communities? Soil Biol. Biochem. 2001, 33, 381–393. [Google Scholar]
- Cicconardi, F.; Nardi, F.; Emerson, B.C.; Frati, F.; Fanciulli, P.P. Deep phylogeographic divisions and long-term persistence of forest invertebrates (Hexapoda: Collembola) in the North-Western Mediterranean basin. Mol. Ecol. 2010, 19, 386–400. [Google Scholar]
- Van Halder, I.; Barbaro, L.; Jactel, H. Conserving butterflies in fragmented plantation forests: Are edge and interior habitats equally important? J. Insect Conserv. 2011, 15, 591–601. [Google Scholar]
- Rosin, Z.M.; Mysczko, L.; Skorka, P.; Lenda, M.; Moron, D.; Sparks, T.H.; Tryjanowski, P. Butterfly responses to environmental factors in fragmented calcareous grasslands. J. Insect Conserv. 2011. [Google Scholar] [CrossRef]
- Ponge, J.F. Biocenoses of Collembola in Atlantic temperate grass-woodland ecosystems. Pedobiologia 1993, 37, 223–244. [Google Scholar]
- Ponge, J.F. Emergent properties from organisms to ecosystems: Towards a realistic approach. Biol. Rev. 2005, 80, 403–411. [Google Scholar]
- Chagnon, M.; Pare, D.; Hebert, C. Relationships between soil chemistry, microbial biomass and the collembolan fauna of southern Quebec sugar maple stands. Ecoscience 2007, 307–316. [Google Scholar]
- Gisin, H. Collembolen fauna Europas; Museum d'Histoire Naturelle: Genève, Switzerland, 1960. [Google Scholar]
- Jordana, R.; Arbea, J.I.; Simon, C.; Lucianez, M.J. Fauna Iberica, Vol 8: Collembola Poduromorpha; Museo Nacional de Ciencias Naturales, Consejor Superior de Investigationes Scientificas: Madrid, Spain, 1997. [Google Scholar]
- Bretfield, G. Synopses on Palaearctic Collembola, Vol. 2, Symphypleona; Abhandlungen und Berichte der Naturkundemuseums: Görlitz, Germany, 1999. [Google Scholar]
- Potapov, M. Synopses on Palaeartic Collembola, Vol. 3, Isotomidae; Staatliches Museum fur Naturkunde: Görlitz, Germany, 2001. [Google Scholar]
- Szeptycki, A. Chaetotaxy of the Entomobryidae and Its phylogenetical Significance, Morpho-Systematic Studies on Collembola; Panstwowe Wydawn. Naukowe: Kraków, Poland, 1979. [Google Scholar]
- Thibaud, J.M.; Schulz, H.J.; da Gama Assalino, M.M. Synopses on Palaeartic Collembola, Vol. 4, Hypogastruridae; Staatliches Museum für Naturkunde: Görlitz, Germany, 2004. [Google Scholar]
- Raimondo, S.; Strazanac, J.S.; Butler, L. Comparison of sampling techniques used in studying Lepidoptera population dynamics. Environ. Entomol. 2004, 33, 418–425. [Google Scholar]
- Yela, J.L.; Holyoak, M. Effects of moonlight and meteorological factors on light and bait trap catches of noctuid moths (Lepidoptera: Noctuidae). Environ. Entomol. 1997, 26, 1283–1290. [Google Scholar]
- Luque, C.; Legal, L.; Staudter, H.; Gers, C.; Wink, M. ISSR (Inter Simple Sequence Repeats) as genetic markers in Noctuids (Lepidoptera, Heterocera, Noctuidae, Noctuinae). Hereditas 2002, 136, 251–253. [Google Scholar]
- Carter, D. Les papillons de France et d'Europe; Solar: Barcelona, Spain, 1984. [Google Scholar]
- Reichholf-Riehm, H. Les papillons; Solar: Paris, France, 1984. [Google Scholar]
- Carter, D. Guide des chenilles d'Europe; Delachaux and Niestlé: Lausanne, Switzerland, 1988. [Google Scholar]
- Leraut, P. Les papillons dans leur milieu; Bordas: Paris, France, 1992. [Google Scholar]
- Skinner, B. Colour Identification Guide to Moths of the British Isles; Viking Press: London, UK, 1984. [Google Scholar]
- Smith, W.; Solow, A.; Preston, P. An estimator of species overlap using a modified beta-binomial model. Biometrics 1996, 52, 1472–1477. [Google Scholar]
- MATLAB®. The Language of Technical Computing 5.2. Optimization Toolbox©, 1999. For Use with MATLAB® version 2; The MathWorks, Inc.: Natick, MA, USA, 1998. [Google Scholar]
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples; Version 8. University of Connecticut: Storrs, CT, USA, 2006. Available online: http://purl.oclc.org/estimates (accessed on 10 October 2011).
- Chao, A. Non-parametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Colwell, R.K.; Mao, C.X.; Chang, J. Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology 2004, 85, 2717–2727. [Google Scholar]
- Carvalho, J.C.; Cardoso, P.; Gomes, P. Determining the relative roles of species replacement and species richness differences in generating beta-diversity patterns. Glob. Ecol. Biogeogr. 2011. [Google Scholar] [CrossRef]
- Tuomisto, H. A diversity of beta diversities: Straightening up a concept gone awry. Part 1. Defining beta diversity as a function of alpha and gamma diversity. Ecography 2010, 33, 2–22. [Google Scholar]
- Tuomisto, H. A diversity of beta diversities: Straightening up a concept gone away. Part 2. Quantifying beta diversity and related phenomena. Ecography 2010, 33, 23–45. [Google Scholar]
- Williams, P.H. Mapping variations in the strength and breadth of biogeographic transition zones using species turnover. Proc. R. Soc. B 1996, 263, 579–588. [Google Scholar]
- Cardoso, P.; Borges, P.A.V.; Veech, J.A. Testing the performance of beta diversity measures based on incidence data: the robustness to undersampling. Divers. Distrib. 2009, 15, 1081–1090. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O'Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package; R Package version 2.0-0. University of Oulu: Oulu, Finland, 2011. Available online: http://cran.r-project.org (accessed on 10 October 2011).
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011. Available online: http://www.rproject.org (accessed on 10 October 2011).
- Honnay, O.; Hermy, M.; Coppin, P. Effects of area, age and diversity of forest patches in Belgium on plant species richness, and implications for conservation and reforestation. Biol. Conserv. 1999, 87, 73–84. [Google Scholar]
- Honnay, O.; Hermy, M.; Coppin, P. Impact of habitat quality on forest plant species colonization. For. Ecol. Manag. 1999, 115, 157–170. [Google Scholar]
- Graae, B.J.; Sunde, P.B.; Fritzboger, B. Vegetation and soil differences in ancient opposed to new forests. For. Ecol. Manag. 2003, 177, 179–190. [Google Scholar]
- Wulf, M. Forest policy in the EU and its influence on the plant diversity of woodlands. J. Environ. Manag. 2003, 67, 15–25. [Google Scholar]
- Hutha, V.; Ojala, R. Collembolan communities in deciduous forest of different origin in Finland. Pedobiologia 2006, 31, 83–90. [Google Scholar]
- Chagnon, M.; Pare, D.; Hebert, C.; Camire, C. Effects of experimental liming on collembolan communities and soil microbial biomass in a southern Quebec sugar maple (Acer saccharum Marsh.) stand. Appl. Soil Ecol. 2001, 17, 81–90. [Google Scholar]
- Cassagne, N.; Gers, C.; Gauquelin, T. Relationships between Collembola, soil chemistry and humus types in forest stands (France). Biol. Fertil. Soils 2003, 37, 355–361. [Google Scholar]
- Jeannel, R. Le peuplement des Pyrénées. Rev. Fr. Entomol. 1947, 14, 53–104. [Google Scholar]
- Hopkins, S.P. Biology of Springtails (Insecta: Collembola); Oxford University Press: Oxford, UK, 1997. [Google Scholar]
- Gauquelin, T.; Gers, C.; Deharveng, L. Physico-chemical heterogeneity of superficial soil layers in conifer plantations versus original broad leaf forests in Ariege (Pyrenees, France). Commun. Soil Sci. Plant. 1996, 27, 2361–2380. [Google Scholar]
- Spiecker, H. Silvicultural management in maintaining biodiversity and resistance of forests in Europe-temperate zone. J. Environ. Manag. 2003, 67, 55–65. [Google Scholar]
- Daily, G.C.; Ehrlich, P.R. Nocturnality and species survival. Proc. Natl. Acad. Sci. USA 1996, 93, 11709–11712. [Google Scholar]
- Heinrich, B. Insect Thermoregulation; John Wiley & Sons: New York, NY, USA, 1981. [Google Scholar]
- Tsuji, J.S.; Kingsolver, J.G.; Watt, W.B. Thermal physiological ecology of Colias butterflies in flight. Oecologia 1986, 69, 161–170. [Google Scholar]
- Ehrlich, P.R. Conservation in temperate forests: What do we need to know and do? For. Ecol. Manag. 1996, 85, 9–19. [Google Scholar]
- Monkkonen, M.; Mutanen, M. Occurrence of moths in boreal forest corridors. Conserv. Biol. 2003, 17, 468–475. [Google Scholar]
- Mazzei, P.; Morel, D.; Panfili, R.; Pimpinelli, I.; Reggianti, D. Moths and Butterflies of Europe and North Africa; Private Publisher: Roma, Italy, 2011. Available on line: http://www.leps.it/ (accessed on 16 November 2011).
- Deharveng, L.; Gers, C.; Legal, L.; Bedos, A.; Albre, J.; Faille, A. Invertébrés endémiques et relictuels de la chaîne des pyrénéenne; DIREN: Midi-Pyrénées, France, 2005. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Luque, C.; Legal, L.; Winterton, P.; Mariano, N.A.; Gers, C. Illustration of the Structure of Arthropod Assemblages (Collembola and Lepidoptera) in Different Forest Types: An Example in the French Pyrenees. Diversity 2011, 3, 693-711. https://doi.org/10.3390/d3040693
Luque C, Legal L, Winterton P, Mariano NA, Gers C. Illustration of the Structure of Arthropod Assemblages (Collembola and Lepidoptera) in Different Forest Types: An Example in the French Pyrenees. Diversity. 2011; 3(4):693-711. https://doi.org/10.3390/d3040693
Chicago/Turabian StyleLuque, Carine, Luc Legal, Peter Winterton, Nestor A. Mariano, and Charles Gers. 2011. "Illustration of the Structure of Arthropod Assemblages (Collembola and Lepidoptera) in Different Forest Types: An Example in the French Pyrenees" Diversity 3, no. 4: 693-711. https://doi.org/10.3390/d3040693
APA StyleLuque, C., Legal, L., Winterton, P., Mariano, N. A., & Gers, C. (2011). Illustration of the Structure of Arthropod Assemblages (Collembola and Lepidoptera) in Different Forest Types: An Example in the French Pyrenees. Diversity, 3(4), 693-711. https://doi.org/10.3390/d3040693