Challenges in Complementing Data from Ground-Based Sensors with Satellite-Derived Products to Measure Ecological Changes in Relation to Climate—Lessons from Temperate Wetland-Upland Landscapes


Abstract
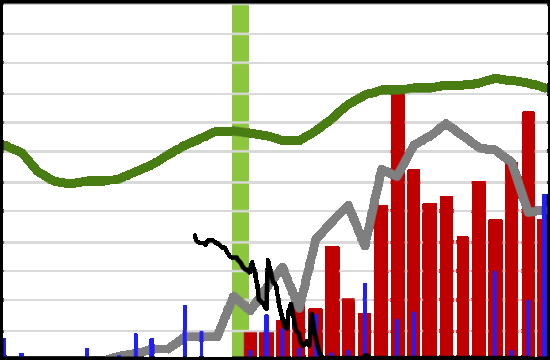
:
1. Introduction
2. Materials and Methods
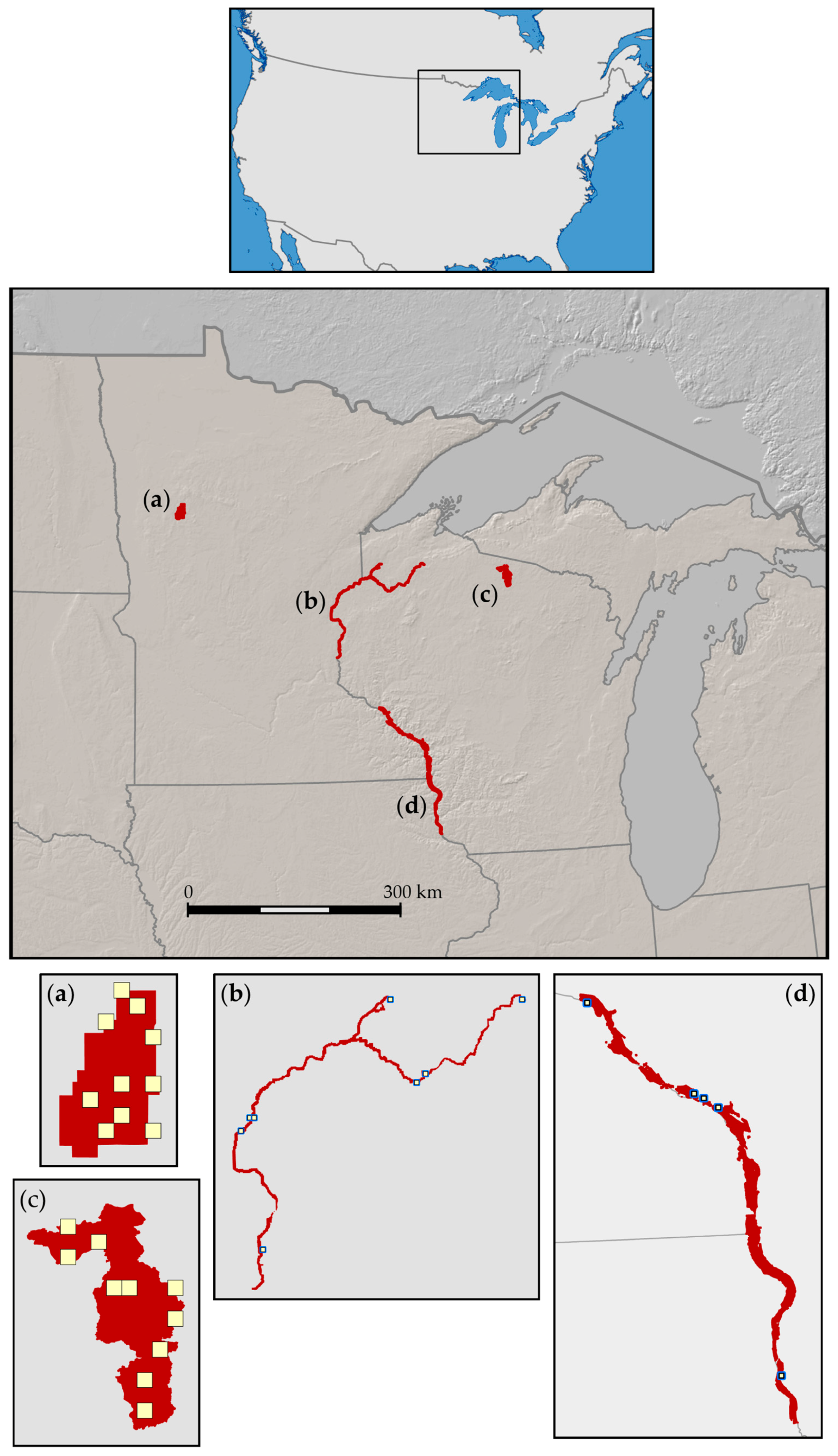
2.1. Study Areas
2.2. In-Situ Measurements
2.3. Data from Weather Stations
2.4. Satellite-Derived Measurements
2.4.1. ET
2.4.2. NDVI
2.4.3. Snow-Off
2.4.4. Integrated Analyses
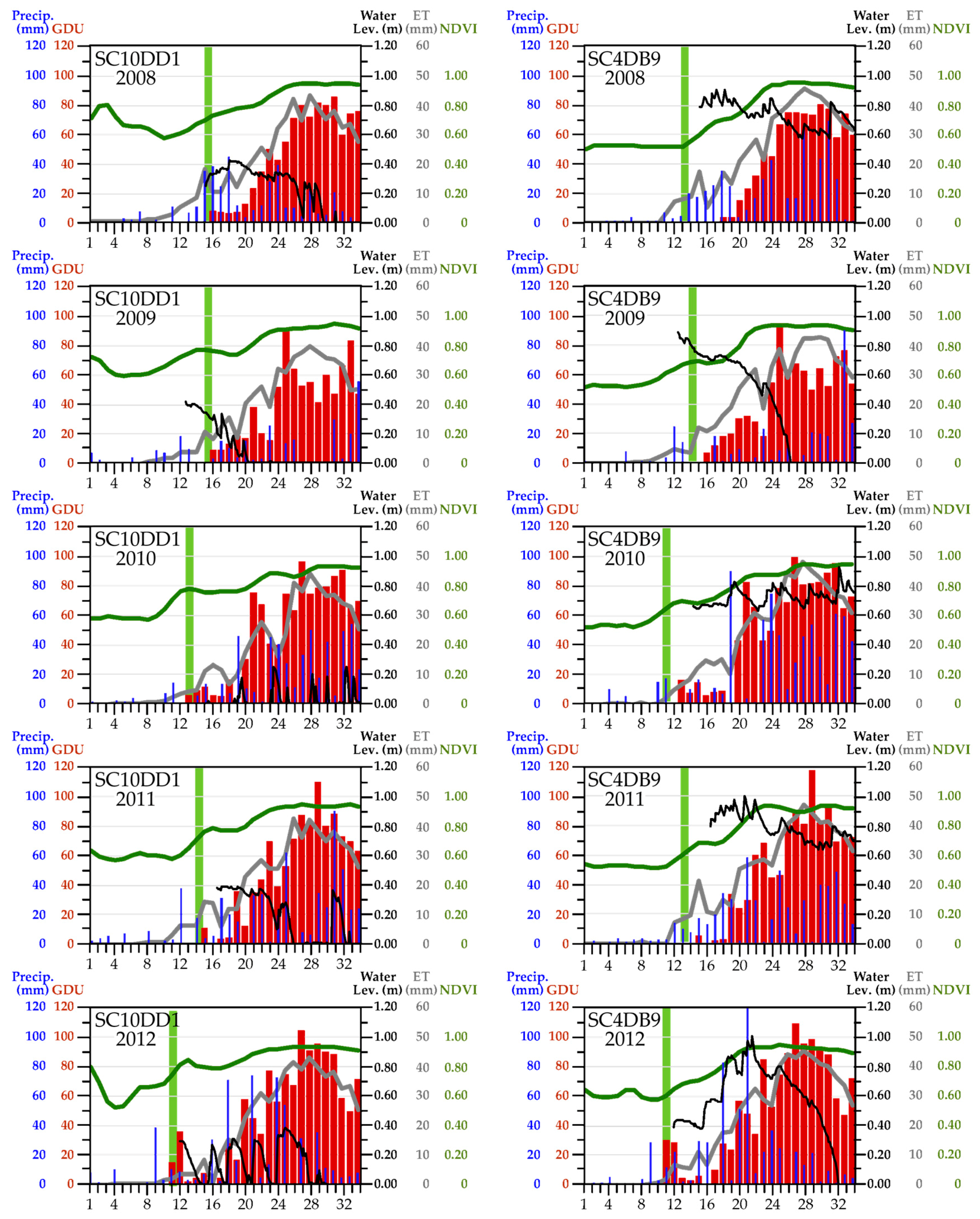
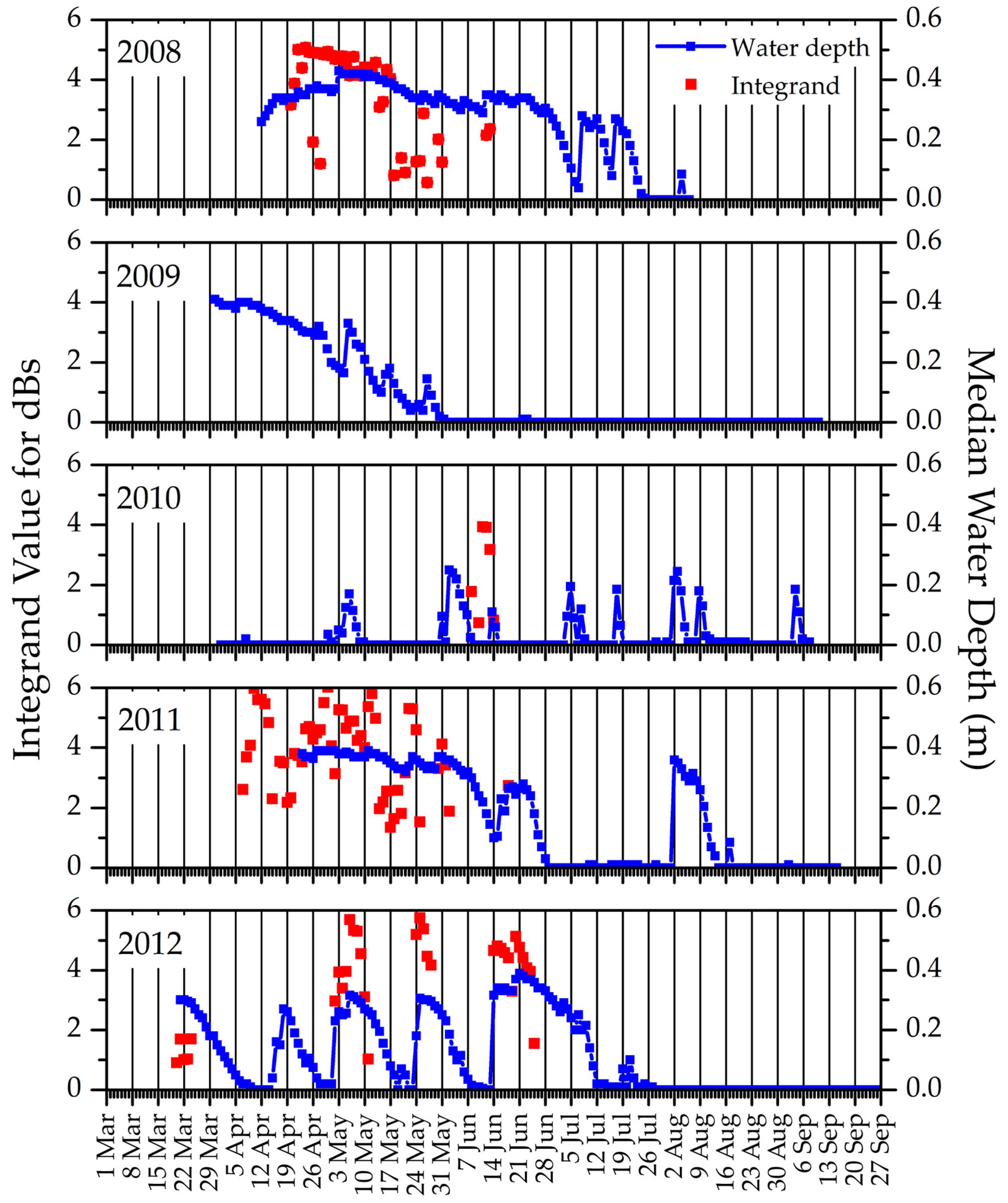
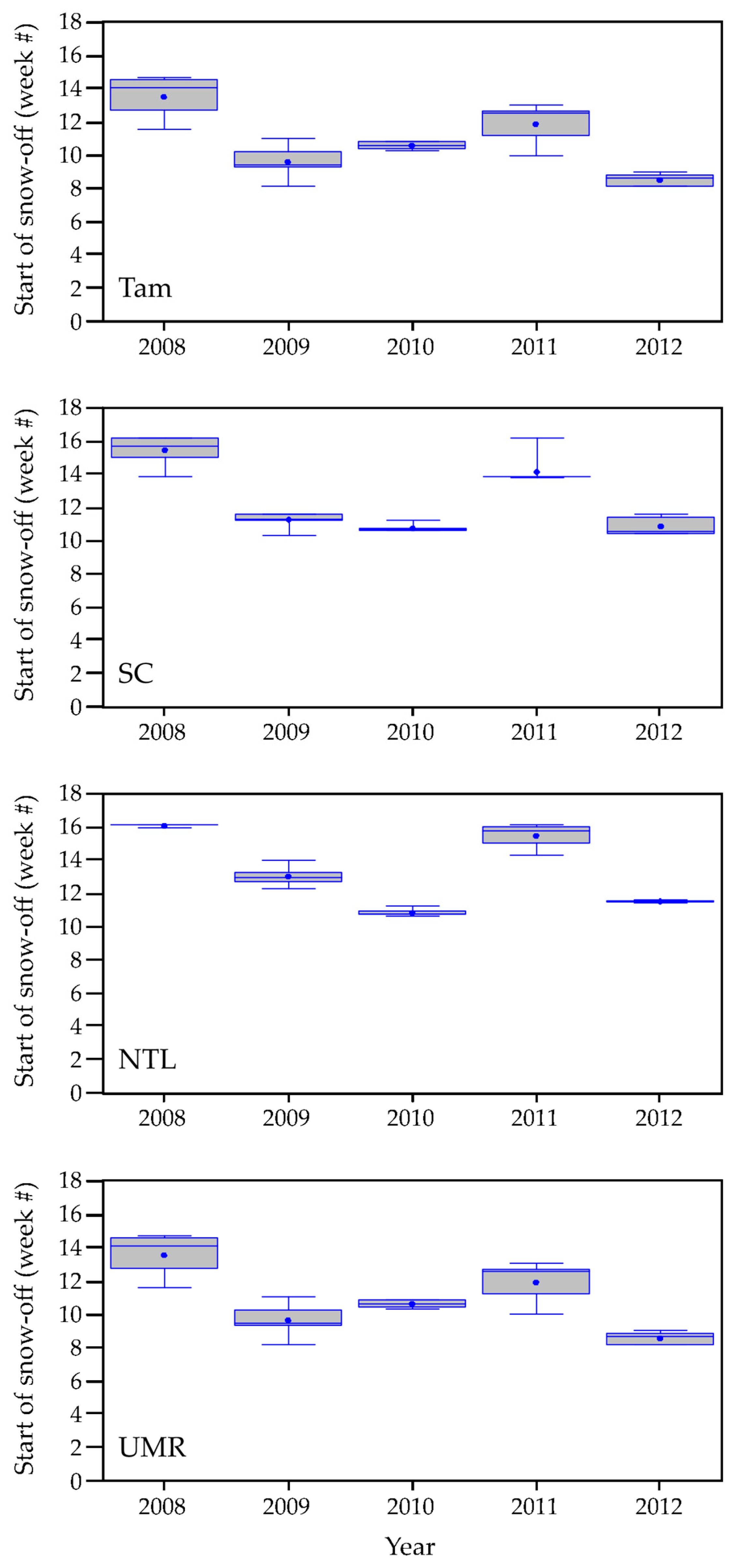
3. Results
3.1. Start of Season
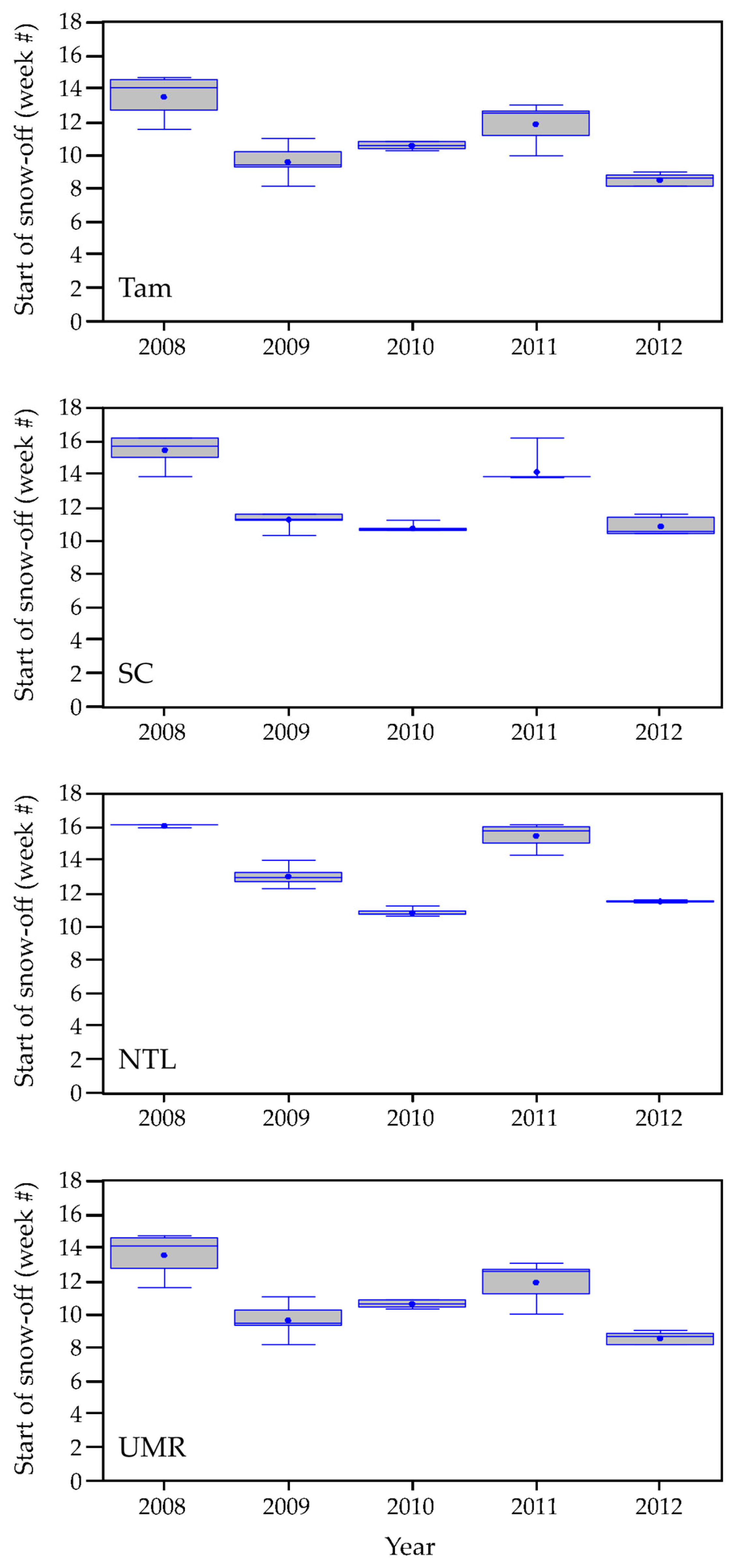
3.1.1. Weather Characteristics, Including Snow Cover
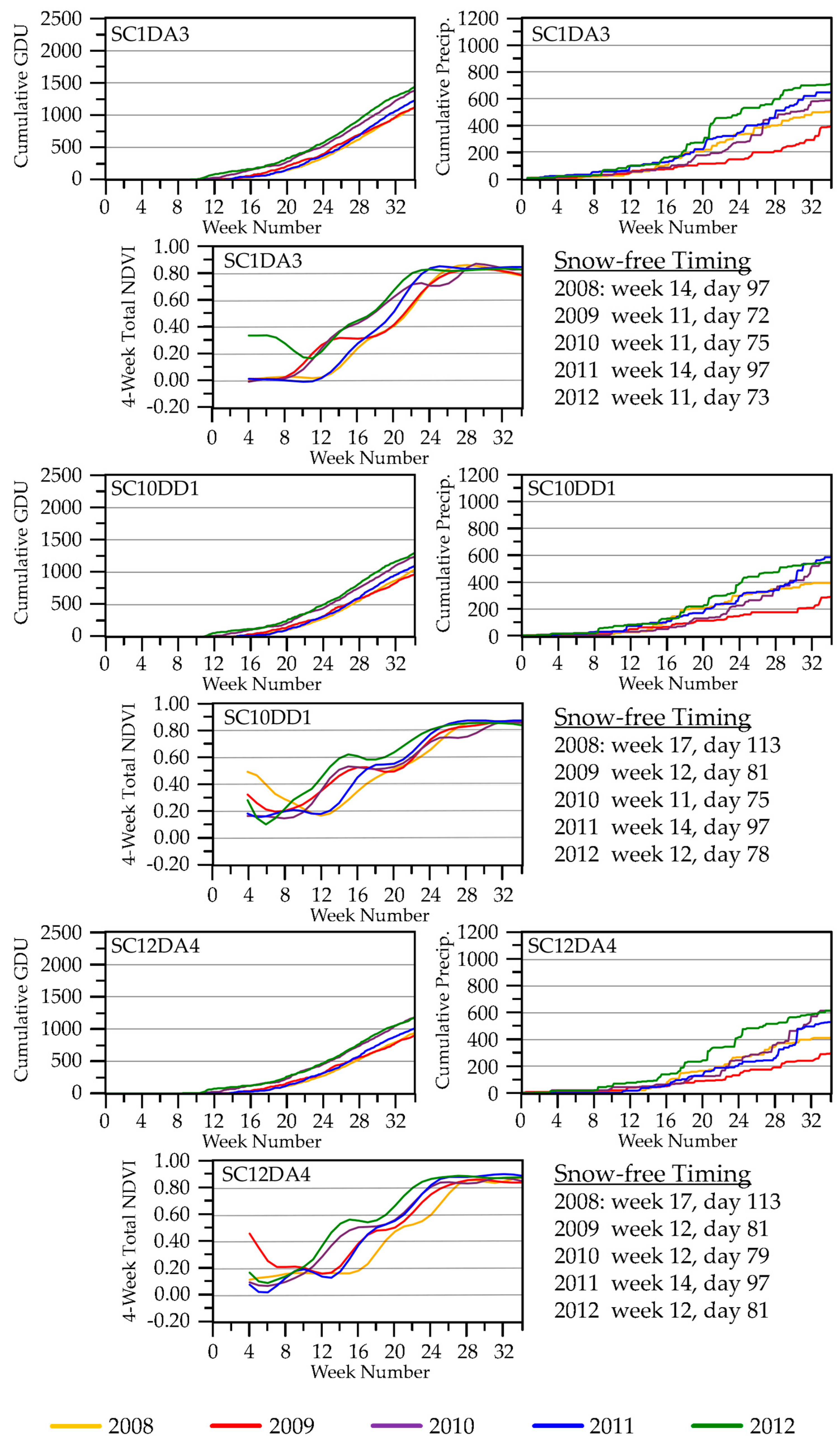
3.1.2. Onset of ET Activity
3.1.3. Vegetation Green-Up
3.2. Seasonal Summaries
3.2.1. Weather
3.2.2. Seasonal ET Dynamics
3.2.3. Seasonal NDVI Dynamics
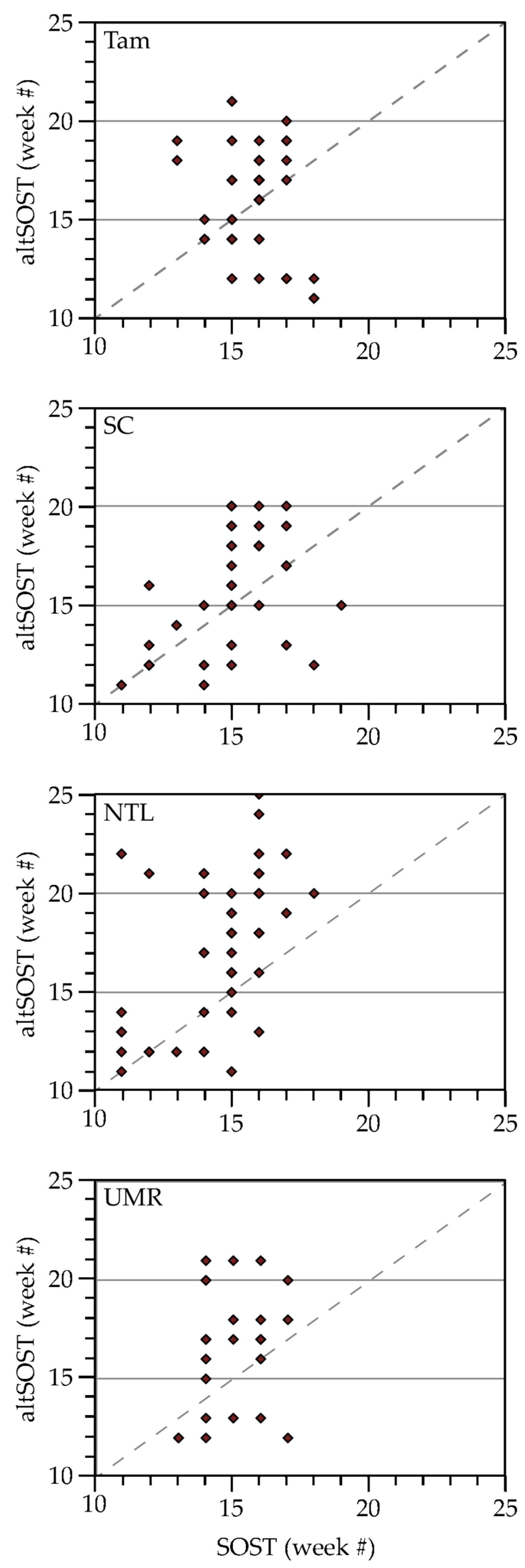
3.2.4. An Integrated Look at the Variables
4. Discussion
4.1. Onset of Growing-Season Conditions
4.2. Tracking Growing-Season Conditions
4.2.1. Seasonal ET Dynamics
4.2.2. Seasonal NDVI Dynamics
4.3. Other Considerations
5. Summary and Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Area | Study Blocks | Weather Station; Type; Location | Latitude Longitude 1 | Records with Missing/Questionable Data and the Mitigation Steps Taken |
|---|---|---|---|---|
| Tam (primary) | All Tam sites | NWS 2 ID 212201; RAWS; Detroit Lakes, MN | 46.84889 −95.84639 | Tmin, Tmax, and P missing for 8–16 April 2009. We substituted with data from Detroit Lakes, MN (station KDTL). |
| Tam (mitigation) | All Tam sites | WS 3 ID KDTL; AWOS; Detroit Lakes, MN | 46.83 −95.89 | |
| SC (primary) | SC1DA3 | NOAA 4 ID USC00212881; GHCND; Forest Lake, MN | 45.3397 −92.9125 | None. |
| SC (primary) | SC4DA3 SC4DAI2 SC4DB9 SC4DBI2 | NWS ID 470602; RAWS; Lind, WI | 45.73972 −92.79556 | Tmin and Tmax missing for 9 April–3 May 2008. We did not substitute any data for these missing temperature records. |
| SC (primary) | SC8DAI1 | NWS ID 470703; RAWS; Minong, WI | 46.13583 −91.98083 | None. |
| SC (primary) | SC10DB1 SC10DD1 | NOAA ID USC00478027; GHCND; Spooner, WI | 45.8236 −91.8761 | Tmin missing for 17 March 2009; Tmin and Tmax missing for all of July 2009; P missing for 23–25 January 2010. We substituted with data from the nearby station in Siren, WI (station KRZN), for July 2009. We did not fill the winter data gap in January 2010. |
| SC (mitigation) | SC10DB1 SC10DD1 | WS ID KRZN; AWOS; Siren, WI | 45.82346 −92.37369 | |
| SC (primary) | SC12DA4 SC12DAI1 | NWS ID 470804; RAWS; Hayward, WI | 46.03111 −91.44900 | P missing for 20–21 February 2012; Tmin and Tmax missing for 20 February–4 March 2012 and 30 March–1 April 2012 and 8–9 April 2012 and 11 April 2012 and 15 April 2012; Tmin, Tmax, and P missing for 1–6 January 2011. We substituted with data from Hayward, WI (station KHYR), for all data gaps. |
| SC (mitigation) | SC12DA4 SC12DAI1 | WS ID KHYR; ASOS; Hayward, WI | 46.0303 −91.4426 | |
| NTL (primary) | TRL1DA1 TRL1DB1 TRL2DA1 TRL2DB1 TRL3DA1 TRL3DB1 TRL3DC1 | NWS ID 471002; RAWS; Woodruff, WI | 45.88972 −89.65222 | P missing from 12–14 May 2011; Tmin and Tmax missing from 11–15 May 2011; P, Tmin, and Tmax missing for 28–29 March 2011. We substituted with data averaged from Arbor Vitae, WI (station KARV), and Minocqua, WI (GHCND:USC00475516), for all data gaps as Woodruff is midway between these two stations. |
| NTL (mitigation) | TRL1DA1 TRL1DB1 TRL2DA1 TRL2DB1 TRL3DA1 TRL3DB1 TRL3DC1 | WS ID KARV; AWOS; Arbor Vitae, WI | 45.9264 −89.7307 | |
| NTL (mitigation) | TRL1DA1 TRL1DB1 TRL2DA1 TRL2DB1 TRL3DA1 TRL3DB1 TRL3DC1 | NOAA ID USC00475516; GHCND; Minocqua, WI | 45.8863 −89.7322 | |
| NTL (primary) | TRL4DA1 TRL4DB1 TRL4DC1 | NWS ID 470302; RAWS; Glidden, WI | 46.14000 −90.00000 | None. |
| UMR (primary) | PSP1 UMRP7 | NOAA ID USC00478589; GHCND; Trempealeau, WI | 43.9994 −91.4378 | |
| UMR (primary) | TrNWRDA1 | NOAA ID USC00472165; GHCND; Dodge, WI | 44.1330 −91.5511 | P, Tmin, and Tmax missing for 31 May 2011, but we did not substitute data for this date. Tmin for 16 January 2009 (−37.8 °C) was noticeably lower than for all other records during 2008–2012; we substituted the Tmin (31.1 °C) recorded at the two nearest stations. |
| UMR (primary) | UMRP4 | NOAA ID USC00470124; GHCND; Alma, WI | 44.32722 −91.91944 | Tmin and Tmax missing for 16 March 2012 and 31 May 2012; Tmax missing for19 July 2012. We did substitute data for missing temperature records because they were sufficiently isolated in time to have little effect on our analyses. |
| UMR (primary) | UMRP10 | NOAA ID USC00476827; GHCND; Prairie du Chien, WI | 43.05150 −91.13490 | Tmin missing for 2 January 2010. We did not substitute data for missing temperature record because it was sufficiently isolated in time to have little effect on our analyses |
Appendix B
| Ending Day-of-Year | Week Number | Day-of-Year When Month Begins | Ending Day-of-Year | Week Number | Day-of-Year When Month Begins |
|---|---|---|---|---|---|
| 7 | 1 | 1 January (day #1) | 189 | 27 | |
| 14 | 2 | 196 | 28 | ||
| 21 | 3 | 203 | 29 | ||
| 28 | 4 | 210 | 30 | ||
| 35 | 5 | 1 February (day #32) | 217 | 31 | 1 August (day #213) |
| 42 | 6 | 224 | 32 | ||
| 49 | 7 | 231 | 33 | ||
| 56 | 8 | 238 | 34 | ||
| 63 | 9 | 1 March (day #60) | 245 | 35 | 1 September (day #244) |
| 70 | 10 | 252 | 36 | ||
| 77 | 11 | 259 | 37 | ||
| 84 | 12 | 266 | 38 | ||
| 91 | 13 | 1 April (day #91) | 273 | 39 | |
| 98 | 14 | 280 | 40 | 1 October (day #274) | |
| 105 | 15 | 287 | 41 | ||
| 112 | 16 | 294 | 42 | ||
| 119 | 17 | 301 | 43 | ||
| 126 | 18 | 1 May (day #121) | 308 | 44 | 1 November (day #305) |
| 133 | 19 | 315 | 45 | ||
| 140 | 20 | 322 | 46 | ||
| 147 | 21 | 329 | 47 | ||
| 154 | 22 | 1 June (day #152) | 336 | 48 | 1 December (day #335) |
| 161 | 23 | 343 | 49 | ||
| 168 | 24 | 350 | 50 | ||
| 175 | 25 | 357 | 51 | ||
| 182 | 26 | 1 July (day #182) | 365 | 52 |
| Day Number | Week Assigned | Day Number | Week Assigned | Day Number | Week Assigned | Day Number | Week Assigned | Day Number | Week Assigned | Day Number | Week Assigned |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1–7 | 1 | 47–53 | 8 | 93–99 | 14 | 139–145 | 21 | 185–191 | 27 | 231–237 | 34 |
| 2–8 | 1 | 48–54 | 8 | 94–100 | 14 | 140–146 | 21 | 186–192 | 27 | 232–238 | 34 |
| 3–9 | 1 | 49–55 | 8 | 95–101 | 14 | 141–147 | 21 | 187–193 | 28 | 233–239 | 34 |
| 4–10 | 1 | 50–56 | 8 | 96–102 | 15 | 142–148 | 21 | 188–194 | 28 | 234–240 | 34 |
| 5–11 | 2 | 51–57 | 8 | 97–103 | 15 | 143–149 | 21 | 189–195 | 28 | 235–241 | 34 |
| 6–12 | 2 | 52–58 | 8 | 98–104 | 15 | 144–150 | 21 | 190–196 | 28 | ||
| 7–13 | 2 | 53–59 | 8 | 99–105 | 15 | 145–151 | 22 | 191–197 | 28 | ||
| 8–14 | 2 | 54–60 | 9 | 100–106 | 15 | 146–152 | 22 | 192–198 | 28 | ||
| 9–15 | 2 | 55–61 | 9 | 101–107 | 15 | 147–153 | 22 | 193–199 | 28 | ||
| 10–16 | 2 | 56–62 | 9 | 102–108 | 15 | 148–154 | 22 | 194–200 | 29 | ||
| 11–17 | 2 | 57–63 | 9 | 103–109 | 16 | 149–155 | 22 | 195–201 | 29 | ||
| 12–18 | 3 | 58–64 | 9 | 104–110 | 16 | 150–156 | 22 | 196–202 | 29 | ||
| 13–19 | 3 | 59–65 | 9 | 105–111 | 16 | 151–157 | 22 | 197–203 | 29 | ||
| 14–20 | 3 | 60–66 | 9 | 106–112 | 16 | 152–158 | 23 | 198–204 | 29 | ||
| 15–21 | 3 | 61–67 | 10 | 107–113 | 16 | 153–159 | 23 | 199–205 | 29 | ||
| 16–22 | 3 | 62–68 | 10 | 108–114 | 16 | 154–160 | 23 | 200–206 | 29 | ||
| 17–23 | 3 | 63–69 | 10 | 109–115 | 16 | 155–161 | 23 | 201–207 | 30 | ||
| 18–24 | 3 | 64–70 | 10 | 110–116 | 17 | 156–162 | 23 | 202–208 | 30 | ||
| 19–25 | 4 | 65–71 | 10 | 111–117 | 17 | 157–163 | 23 | 203–209 | 30 | ||
| 20–26 | 4 | 66–72 | 10 | 112–118 | 17 | 158–164 | 23 | 204–210 | 30 | ||
| 21–27 | 4 | 67–73 | 10 | 113–119 | 17 | 159–165 | 24 | 205–211 | 30 | ||
| 22–28 | 4 | 68–74 | 11 | 114–120 | 17 | 160–166 | 24 | 206–212 | 30 | ||
| 23–29 | 4 | 69–75 | 11 | 115–121 | 17 | 161–167 | 24 | 207–213 | 30 | ||
| 24–30 | 4 | 70–76 | 11 | 116–122 | 17 | 162–168 | 24 | 208–214 | 31 | ||
| 25–31 | 4 | 71–77 | 11 | 117–123 | 18 | 163–169 | 24 | 209–215 | 31 | ||
| 26–32 | 5 | 72–78 | 11 | 118–124 | 18 | 164–170 | 24 | 210–216 | 31 | ||
| 27–33 | 5 | 73–79 | 11 | 119–125 | 18 | 165–171 | 24 | 211–217 | 31 | ||
| 28–34 | 5 | 74–80 | 11 | 120–126 | 18 | 166–172 | 25 | 212–218 | 31 | ||
| 29–35 | 5 | 75–81 | 12 | 121–127 | 18 | 167–173 | 25 | 213–219 | 31 | ||
| 30–36 | 5 | 76–82 | 12 | 122–128 | 18 | 168–174 | 25 | 214–220 | 31 | ||
| 31–37 | 5 | 77–83 | 12 | 123–129 | 18 | 169–175 | 25 | 215–221 | 32 | ||
| 32–38 | 5 | 78–84 | 12 | 124–130 | 19 | 170–176 | 25 | 216–222 | 32 | ||
| 33–39 | 6 | 79–85 | 12 | 125–131 | 19 | 171–177 | 25 | 217–223 | 32 | ||
| 34–40 | 6 | 80–86 | 12 | 126–132 | 19 | 172–178 | 25 | 218–224 | 32 | ||
| 35–41 | 6 | 81–87 | 12 | 127–133 | 19 | 173–179 | 26 | 219–225 | 32 | ||
| 36–42 | 6 | 82–88 | 13 | 128–134 | 19 | 174–180 | 26 | 220–226 | 32 | ||
| 37–43 | 6 | 83–89 | 13 | 129–135 | 19 | 175–181 | 26 | 221–227 | 32 | ||
| 38–44 | 6 | 84–90 | 13 | 130–136 | 19 | 176–182 | 26 | 222–228 | 33 | ||
| 39–45 | 6 | 85–91 | 13 | 131–137 | 20 | 177–183 | 26 | 223–229 | 33 | ||
| 40–46 | 7 | 86–92 | 13 | 132–138 | 20 | 178–184 | 26 | 224–230 | 33 | ||
| 41–47 | 7 | 87–93 | 13 | 133–139 | 20 | 179–185 | 26 | 225–231 | 33 | ||
| 42–48 | 7 | 88–94 | 13 | 134–140 | 20 | 180–186 | 27 | 226–232 | 33 | ||
| 43–49 | 7 | 89–95 | 14 | 135–141 | 20 | 181–187 | 27 | 227–233 | 33 | ||
| 44–50 | 7 | 90–96 | 14 | 136–142 | 20 | 182–188 | 27 | 228–234 | 33 | ||
| 45–51 | 7 | 91–97 | 14 | 137–143 | 20 | 183–189 | 27 | 229–235 | 34 | ||
| 46–52 | 7 | 92–98 | 14 | 138–144 | 21 | 184–190 | 27 | 230–236 | 34 |
| 8-Day Interval | Days of Year | Week Assigned 1 |
|---|---|---|
| 1 | 1–8 | 1 |
| 2 | 9–16 | 2 |
| 3 | 17–24 | 3 |
| 4 | 25–32 | 4 |
| 5 | 33–40 | 6 |
| 6 | 41–48 | 7 |
| 7 | 49–56 | 8 |
| 8 | 57–64 | 9 |
| 9 | 65–72 | 10 |
| 10 | 73–80 | 11 |
| 11 | 81–88 | 12 |
| 12 | 89–96 | 14 |
| 13 | 97–104 | 15 |
| 14 | 105–112 | 16 |
| 15 | 113–120 | 17 |
| 16 | 121–128 | 18 |
| 17 | 129–136 | 19 |
| 18 | 137–144 | 20 |
| 19 | 145–152 | 22 |
| 20 | 153–160 | 23 |
| 21 | 161–168 | 24 |
| 22 | 169–176 | 25 |
| 23 | 177–184 | 26 |
| 24 | 185–192 | 27 |
| 25 | 193–200 | 28 |
| 26 | 201–208 | 30 |
| 27 | 209–216 | 31 |
| 28 | 217–224 | 32 |
| 29 | 225–232 | 33 |
| 30 | 233–240 | 34 |
References
- Coops, N.C.; Hilker, T.; Bater, C.W.; Wulder, M.A.; Nielsen, S.E.; McDermid, G.; Stenhouse, G. Linking ground-based to satellite-derived phenological metrics in support of habitat assessment. Remote Sens. Lett. 2012, 3, 191–200. [Google Scholar] [CrossRef]
- Sessa, R.; Dolman, H. Terrestrial Essential Climate Variables for Climate Change Assessment, Mitigation and Adaptation (GTOS 52); Food and Agriculture Organization of the United Nations: Rome, Italy, 2008; p. 40. [Google Scholar]
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.-M.; Tucker, C.J.; Stenseth, N.C. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends Ecol. Evol. 2005, 20, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Wells, K.D. The Ecology and Behavior of Amphibians; University of Chicago Press: Chicago, IL, USA, 2007. [Google Scholar]
- Hopkins, W.A. Amphibians as models for studying environmental change. ILAR J. 2007, 48, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Pilliod, D.S.; Peterson, C.R. Local and landscape effects of introduced trout on amphibians in historically fishless watersheds. Ecosystems 2001, 4, 322–333. [Google Scholar] [CrossRef]
- Mazzoni, R.; Cunningham, A.A.; Daszak, P.; Apolo, A.; Perdomo, E.; Speranza, G. Emerging pathogen of wild amphibians in frogs (Rana catesbeiana) farmed for international trade. Emerg. Infect. Dis. 2003, 9, 995–998. [Google Scholar] [CrossRef] [PubMed]
- Pounds, J.A.; Bustamante, M.R.; Coloma, L.A.; Consuegra, J.A.; Fogden, M.P.L.; Foster, P.M.; La Marca, E.; Masters, K.L.; Merino-Viteri, A.; Puschendorf, R.; et al. Widespread amphibian extinctions from epidemic diseas driven by global warming. Nature 2006, 439, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Kiesecker, J.M. Synergism between trematode infection and pesticide exposure: A link to amphibian limb deformities in nature? Proc. Natl. Acad. Sci. USA 2002, 99, 9900–9904. [Google Scholar] [CrossRef] [PubMed]
- Hayes, T.B.; Case, P.; Chui, S.; Chung, D.; Haeffele, C.; Haston, K.; Lee, M.; Mai, V.P.; Marjuoa, Y.; Parker, J.; et al. Pesticide mixtures, endocrine disruption, and amphibian declines: Are we underestimating the impact? Environ. Health Perspect. 2006, 114, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, N.S.; Bickford, D.; Diesmos, A.C.; Lee, T.M.; Koh, L.P.; Brook, B.W.; Sekercloglu, C.H.; Bradshaw, C.J.A. Measuring the meltdown: Drivers of global amphibian extinction and decline. PLoS ONE 2008, 3, 8. [Google Scholar] [CrossRef] [PubMed]
- Gallant, A.L.; Klaver, R.W.; Casper, G.S.; Lannoo, M.J. Global rates of habitat loss and implications for amphibian conservation. Copeia 2007, 2007, 967–979. [Google Scholar] [CrossRef]
- Bunn, S.E. Grand challenge for the future of freshwater ecosy*stems. Front. Environ. Sci. 2016, 4, 1–4. [Google Scholar] [CrossRef]
- Dolman, H.; Latham, J.; Sessa, R. Introduction. In Terrestrial Essential Climate Variables for Climate Change Assessment, Mitigation and Adaptation; Food and Agriculture Organization of the United Nations: Rome, Italy, 2008; p. 2. [Google Scholar]
- Millennium Ecosystem Assessment. Ecosystems and Human Well-Being: Wetlands and Water—Synthesis; World Resources Institute: Washington, DC, USA, 2005; p. 68. [Google Scholar]
- Marshall, C.H.; Pielke, R.A.; Steyaert, L.T. Has the conversion of natural wetlands to agricultural land increased the incidence and severity of damaging freezes in south Florida? Mon. Weather Rev. 2004, 132, 2243–2258. [Google Scholar] [CrossRef]
- Frappart, F.; Papa, F.; Malbeteau, Y.; Leon, J.G.; Ramillien, G.; Prigent, C.; Seoane, L.; Seyler, F.; Calmant, S. Surface freshwater storage variations in the Orinoco floodplains using multi-satellite observations. Remote Sens. 2015, 7, 89–110. [Google Scholar] [CrossRef]
- Pekel, J.F.; Cottam, A.; Gorelick, N.; Belward, A.S. High-resolution mapping of global surface water and its long-term changes. Nature 2016, 540, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Tulbure, M.G.; Broich, M. Spatiotemporal dynamic of surface water bodies using Landsat time-series data from 1999 to 2011. ISPRS J. Photogramm. Remote Sens. 2013, 79, 44–52. [Google Scholar] [CrossRef]
- Milly, P.C.D.; Betancourt, J.; Falkenmark, M.; Hirsch, R.M.; Kundzewicz, Z.W.; Letternaier, D.P.; Stouffer, R.J. Stationarity is dead: Whither water management? Science 2008, 319, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Melillo, J.M.; Richmond, T.; Yohe, G.W. (Eds.) Climate Change Impacts in the United States: The Third National Climate Assessment; The US Government Publishing Office: Washington, DC, USA, 2014; p. 841.
- Skidmore, A.K.; Pettorelli, N.; Coops, N.C.; Geller, G.N.; Hansen, M.; Lucas, R.; Mucher, C.A.; O’Connor, B.; Paganini, M.; Pereira, H.M.; et al. Agree on biodiversity metrics to track from space. Nature 2015, 523, 403–405. [Google Scholar] [CrossRef] [PubMed]
- Alcaraz-Segura, D.; Cabello, J.; Paruelo, J. Baseline characterization of major Iberian vegetation types based on the NDVI dynamics. Plant Ecol. 2009, 202, 13–29. [Google Scholar] [CrossRef]
- De Beurs, K.M.; Hennebry, G.M. Land surface phenology, climate variation, and institutional change: Analyzing agricultural land cover change in Kazakhstan. Remote Sens. Environ. 2004, 89, 497–509. [Google Scholar] [CrossRef]
- Reed, B.C.; Brown, J.F.; VanderZee, D.; Loveland, T.R.; Merchant, J.W.; Ohlen, D.O. Measuring phenological variability from satellite imagery. J. Veg. Sci. 1994, 5, 703–714. [Google Scholar] [CrossRef]
- Studer, S.R.; Stöckli, R.; Appenzeller, C.; Vidale, P.L. A comparative study of satellite and ground-based phenology. Int. J. Biometeorol. 2007, 51, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Sadinski, W.; Gallant, A.L.; Roth, M.; Brown, J.; Senay, G.; Brininger, W.; Stoker, J. Multi-year data from satellite- and ground-based sensors show details and scale matter in assessing climate’s effects on wetland surface water, amphibians, and landscape conditions. PLoS ONE 2018. under review. [Google Scholar]
- Butcher, R.D. America’s National Wildlife Refuges, 2nd ed.; Taylor Trade Publishing: Lanham, MD, USA, 2008; pp. 146–147. ISBN 978-1-58979-383-5. [Google Scholar]
- US Geological Survey—Upper Midwest Environmental Sciences Center. Vegetation of Saint Croix National Scenic Riverway; US Geological Survey—Upper Midwest Environmental Sciences Center: La Crosse, WI, USA, 2012.
- Northern Highland-American Legion State Forest. Northern Highland-American Legion State Forest, Draft Master Plan and Environmental Impact Statement; Wisconsin Department of Natural Resources: Madison, WI, USA, 2005; p. 203.
- U.S. Department of the Interior—Fish and Wildlife Service. Trempealeau National Wildlife Refuge Environmental Impact Statement and Comprehensive Conservation Plan; U.S. Fish and Wildlife Service: Trempealeau, WI, USA, 2007; p. 291.
- Wisconsin’s Natural Heritage Inventory Program. Rapid Ecological Assessment for Perrot State Park, Merrick State Park & Whitman Dam Wildlife Area; Wisconsin’s Natural Heritage Inventory Program: Madison, WI, USA, 2012; p. 74.
- Ramsar Convention Secretariat, Wetlands of International Importance (Ramsar Sites). Available online: http://www.ramsar.org/about/wetlands-of-international-importance-ramsar-sites (accessed on 28 September 2017).
- Womach, J. Agriculture: A Glossary of Terms, Programs; The Library of Congress: Washington, DC, USA, 2005.
- McMaster, G.S.; Wilhelm, W. Growing degree-days: One equation, two interpretations. Agric. For. Meteorol 1997, 87, 291–300. [Google Scholar] [CrossRef]
- Qian, B.; Zhang, X.; Chen, K.; Feng, Y.; O’Brien, T. Observed long-term trends for agroclimatic conditions in Canada. J. Appl. Meteorol. Climatol. 2010, 49, 604–618. [Google Scholar] [CrossRef]
- De Beurs, K.M.; Henebry, G.M. Spatio-temporal statistical methods for modelling land surface phenology. In Phenological Research; Hudson, I.L., Keatley, M.R., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 177–208. [Google Scholar]
- Brown, M.E.; de Beurs, K.M.; Marshall, M. Global phenological response to climate change in crop areas using satellite remote sensing of vegetation, humidity and temperature over 26 years. Remote Sens. Environ. 2012, 126, 174–183. [Google Scholar] [CrossRef]
- De Beurs, K.M.; Henebry, G.M. A land surface phenology assessment of the northern polar regions using MODIS reflectance time series. Can. J. Remote Sens. 2010, 36, S87–S110. [Google Scholar] [CrossRef]
- Yang, W.; Yang, L.; Merchant, J.W. An assessment of AVHRR/NDVI-ecoclimatological relations in Nebraska, USA. Int. J. Remote Sens. 1997, 18, 2161–2180. [Google Scholar] [CrossRef]
- Yang, L.; Wylie, B.K.; Tieszen, L.L.; Reed, B.C. An analysis of relationships among climate forcing and time-integrated NDVI of grasslands over the US northern and central Great Plains. Remote Sens. Environ. 1998, 65, 25–37. [Google Scholar] [CrossRef]
- Multi-Resolution Land Characteristics Consortium (MRLC). Available online: https://www.mrlc.gov/index.php (accessed on 6 December 2017).
- Lannoo, M. Amphibian Declines—The Conservation Status of United States Species; University of California Press: Berkeley, CA, USA, 2005. [Google Scholar]
- Dodd, C.K., Jr. Frogs of the United States and Canada; Johns Hopkins University Press: Baltimore, MD, USA, 2014. [Google Scholar]
- Petranka, J.W. Salamanders of the United States and Canada; Smithsonian Institution Press: Washington, DC, USA, 1998. [Google Scholar]
- Risk Assessment—Species Data Matrices: Biological Attributes that May Contribute to Vulnerability. Available online: http://northeastparc.org/risk-assessment (accessed on 14 February 2018).
- Brown, J.F.; Howard, D.; Wylie, B.; Frieze, A.; Ji, L.; Gacke, C. Application-ready expedited MODIS data for operational land surface monitoring of vegetation condition. Remote Sens. 2015, 7, 16226–16240. [Google Scholar] [CrossRef]
- Gallant, A.L. Data Files Supporting the Paper Titled “Challenges in Complementing Data from Ground-Based Sensors with Satellite-Derived Products to Measure Ecological Changes in Relation to Climate—Lessons from Temperate Wetland-Upland Landscapes”. Available online: https://www.sciencebase.gov (accessed on 22 February 2018).
- Senay, G.B.; Bohms, S.; Singh, R.K.; Gowda, P.H.; Velpuri, N.M.; Alemu, H.; Verdin, J.P. Operational evapotranspiration mapping using remote sensing and weather datasets: A new parameterization for the SSEB approach. J. Am. Water Resour. Assoc. 2013, 49, 577–591. [Google Scholar] [CrossRef]
- Wan, Z. Collection-5 MODIS Land Surface Temperature Products User’s Guide. Available online: https://icess.eri.ucsb.edu/modis/LstUsrGuide/usrguide.html (accessed on 6 December 2017).
- Thornton, P.E.; Running, S.W.; White, M.A. Generating surfaces of daily meteorology variables over large regions of complex terrain. J. Hydrol. 1997, 190, 214–251. [Google Scholar] [CrossRef]
- Thornton, P.E.; Thornton, M.M.; Mayer, B.W.; Wilhelmi, N.; Wei, Y.; Cook, R.B. Daymet: Daily Surface Weather on a 1-km grid for North America. Available online: https://daac.ornl.gov/cgi-bin/dsviewer.pl?ds_id=1328 (accessed on 19 December 2017).
- Senay, G.B.; Verdin, J.P.; Lietzow, R.; Melesse, A.M. Global reference evapotranspiration modeling and evaluation. J. Am. Water Resour. Assoc. 2008, 44, 969–979. [Google Scholar] [CrossRef]
- Kanamitsu, M. Description of the NMC Global Data Assimilation and Forecast System. Weather Forecast. 1989, 4, 334–342. [Google Scholar] [CrossRef]
- Velpuri, N.M.; Senay, G.B.; Singh, R.K.; Bohms, S.; Verdin, J.P. A comprehensive evaluation of two MODIS evapotranspiration products over the conterminous United States: Using point and gridded FLUXNET and water balance ET. Remote Sens. Environ. 2013, 139, 35–49. [Google Scholar] [CrossRef]
- FEWS Data Downloads. Available online: https://earlywarning.usgs.gov/fews/datadownloads/Continental%20Africa/Monthly%20ET%20Anomaly (accessed on 28 December 2017).
- eMODIS (EROS Moderate Resolution Imaging Spectroradiometer). Available online: https://lta.cr.usgs.gov/emodis (accessed on 1 April 2015).
- Jenkerson, C.; Maiersperger, T.; Schmidt, G. eMODIS—A User-Friendly Data Source; Open-File Report 2010-1055; US Geological Survey: Sioux Falls, SD, USA, 2010.
- Remote Sensing Phenology. Available online: https://phenology.cr.usgs.gov/ (accessed on 6 December 2017).
- Swets, D.L.; Reed, B.C.; Rowland, J.D.; Marko, S.E. A weighted least-squares approach to temporal NDVI smoothing. In Proceedings of the 1999 ASPRS Annual Conference, Portland, OR, USA, 17–21 May 1999. [Google Scholar]
- Brown, J.F. Start of Season Time Dataset. Available online: https://phenology.cr.usgs.gov/get_data_250e.php (accessed on 1 April 2015).
- White, M.A.; de Beurs, K.M.; Didan, K.; Inouye, D.W.; Richardson, A.D.; Jensen, O.P.; O’Keefe, J.; Zhang, G.; Nemani, R.R.; van Leeuwen, W.J.D.; et al. Intercomparison, interpretation, and assessment of spring phenology in North America estimated from remote sensing for 1982–2006. Glob. Chang. Biol. 2009, 15, 2335–2359. [Google Scholar] [CrossRef]
- Riggs, G.A.; Hall, D.K.; Salomonson, V.V. MODIS Snow Product User Guide for Collection 4 Data Products. Available online: http://modis-snow-ice.gsfc.nasa.gov/?c=sug_main (accessed on 13 January 2017).
- O’Leary, D.S., III; Kellermann, J.L.; Wayne, C. Snowmelt timing, phenology, and growing season length in conifer forests of Crater Lake National Park, USA. Int. J. Biometeorol. 2018, 62, 273–285. [Google Scholar]
- Arismendi, I.; Safeeq, M.; Johnson, S.L.; Dunham, J.B.; Haggerty, R. Increasing synchrony of high temperature and low flow in western Northm American streams: Double trouble for coldwater biota? Hydrobiologia 2013, 712, 61–70. [Google Scholar] [CrossRef]
- Kellermann, J.L.; van Riper, C., III. Detecting mismatches of bird migration stopover and tree phenology in response to changing climate. Oecologia 2015, 178, 1227–1238. [Google Scholar] [CrossRef] [PubMed]
- Thackery, S.J.; Sparks, T.H.; Frederiksen, M.; Burthe, S.; Bacon, P.M.; Bell, J.R.; Botham, M.S.; Brereton, T.M.; Bright, P.W.; Carvalho, L.; et al. Trophic level asynchrony in rates of phenological change for marine, freshwater and terrestrial environments. Glob. Chang. Biol. 2010, 16, 3304–3313. [Google Scholar] [CrossRef]
- Geiger, R. The Climate Near the Ground, 3rd ed.; Harvard University Press: Cambridge, MA, USA, 1950; pp. 26–35. [Google Scholar]
- Walter, H. Vegetation of the Earth in Relation to Climate and the Eco-Physiological Conditions; Springer: New York, NY, USA, 1973; p. 237. ISBN 0340171650, 9780340171653. [Google Scholar]
- Spurr, S.H.; Barnes, B.V. Forest Ecology, 2nd ed.; John Wiley & Sons, Inc.: New York, NY, USA, 1980; p. 687. ISBN 0471068411. [Google Scholar]
- Palmer, W.C. Meteorological Drought; US Department of Commerce, Weather Bureau: Washington, DC, USA, 1965.
- National Oceanic and Atmospheric Administration—National Climate Data Center, National Environmental Satellite, Data, and Information Service. Available online: https://www7.ncdc.noaa.gov/CDO/CDODivisionalSelect.jsp (accessed on 13 December 2017).
- Gallant, A.L.; Kaya, S.G.; White, L.; Brisco, B.; Roth, M.F.; Sadinski, W.; Rover, J. Detecting emergence, growth, and senescence of wetland vegetation with polarimetric synthetic aperture radar (SAR) data. Water 2014, 6, 694–722. [Google Scholar] [CrossRef]
- Devlin, M.J.; da Silva, E.T.; Petus, C.; Wenger, A.; Zeh, D.; Tracey, D.; Alvarez-Romero, J.G.; Brodie, J. Combining in-situ water quality and remotely sensed data across spatial and temporal scales to measure variability in wet season chlorophyll-a: Great Barrier Reef lagoon (Queensland, Australia). Ecol. Process. 2013, 2, 22. [Google Scholar] [CrossRef]
- Huber, R.; Briner, S.; Bugmann, H.; Elkin, C.; Hirschi, C.; Seidl, R.; Snell, R.; Ringling, A. Inter- and transdisciplinary perspective on the integration of ecological processes into ecosystem services analysis in a mountain region. Ecol. Process. 2014, 3, 14. [Google Scholar] [CrossRef]
- Danz, N.P.; Niemi, G.J.; Regal, R.R.; Hollenhorst, T.; Johnson, L.B.; Hanowski, J.A.M.; Axler, R.P.; Ciborowski, J.J.H.; Hrabik, T.; Brady, V.J.; et al. Integrated measures of anthropogenic stress in the U.S. Great Lakes Basin. Environ. Manag. 2007, 39, 631–647. [Google Scholar] [CrossRef] [PubMed]
- Burton, T.M.; Brazner, J.C.; Ciborowski, J.J.H.; Grabas, G.P.; Hummer, J.; Schneider, J.; Uzarski, D.G. Great Lakes Coastal Wetlands Monitoring Plan; Great Lakes Commission: Ann Arbor, MI, USA, 2008. [Google Scholar]
- Halpern, B.S.; Longo, C.; Hardy, D.; McLeod, K.L.; Samhouri, J.F.; Katona, S.K.; Kleisner, K.; Lester, S.E.; O’Leary, J.; Ranelletti, M.; et al. An index to assess the health and benefits of the global ocean. Nature 2012, 488, 615–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andresen, C.G.; Tweedie, C.E.; Lougheed, V.L. Climate and nutrient effects on Arctic wetland plant phenology observed from phenocams. Remote Sens. Environ. 2018, 205, 46–55. [Google Scholar] [CrossRef]
- Yun, K.; Hsiao, J.; Jung, M.P.; Choi, I.T.; Glenn, D.M. Can a multi-model ensemble improve phenology predictions for climate change studies. Ecol. Model. 2017, 362, 54–64. [Google Scholar] [CrossRef]
- Yuan, F.; Wang, C.; Mitchell, M. Spatial patterns of land surface phenology relative to monthly climate variations: US Great Plains. GISci. Remote Sens. 2014, 51, 30–50. [Google Scholar] [CrossRef]
- Zipkin, E.F.; Ries, L.; Reeves, R.; Regetz, J.; Oberhauser, K.S. Tracking climate impacts on the migratory monarch butterfly. Glob. Chang. Biol. 2012, 18, 3039–3049. [Google Scholar] [CrossRef] [PubMed]
- Lillesand, T.M.; Kiefer, R.W. Remote Sensing and Image Interpretation, 3rd ed.; John Wiley & Sons, Inc.: New York, NY, USA, 1994; p. 750. ISBN 0-471-5778309. [Google Scholar]
- Gallant, A.L. The challenges of remote monitoring of wetlands. Remote Sens. 2015, 7, 10938–10950. [Google Scholar] [CrossRef]
- Brisco, B.; Kapfer, M.; Hirose, T.; Tedford, B.; Liu, J. Evaluation of C-band polarization diversity and polarimetry for wetland mapping. Can. J. Remote Sens. 2011, 37, 82–92. [Google Scholar] [CrossRef]
- Brisco, B.; Schmitt, A.; Murnaghan, K.; Kaya, S.; Roth, A. SAR polarimetric change detection for flooded vegetation. Int. J. Digit. Earth 2013, 6, 103–114. [Google Scholar] [CrossRef]
- Corcoran, J.; Knight, J.; Brisco, B.; Kaya, S.; Cull, A.; Murnaghan, K. The integration of optical, topographic, and radar data for wetland mapping in northern Minnesota. Can. J. Remote Sens. 2011, 37, 564–582. [Google Scholar] [CrossRef]
- Wickham, J.D.; Stehman, S.V.; Fry, J.A.; Smith, J.H.; Homer, C.G. Thematic accuracy of the NLCD 2001 land cover for the conterminous United States. Remote Sens. Environ. 2010, 114, 1286. [Google Scholar] [CrossRef]
- Wickham, J.D.; Stehman, S.V.; Gass, L.; Dewitz, J.; Fry, J.A.; Wade, T.G. Accuracy assessment of NLCD 2006 land cover and impervious surface. Remote Sens. Environ. 2013, 130, 294–304. [Google Scholar] [CrossRef]
- Rover, J.; Wylie, B.K.; Ji, L. A self-trained classification technique for producing 30 m percent-water maps from Landsat data. Int. J. Remote Sens. 2010, 31, 2197–2203. [Google Scholar] [CrossRef]
- Adam, E.; Mutanga, O.; Rugege, D. Multispectral and hyperspectral remote sensing for identification and mapping of wetland vegetation: A review. Wetlands Ecol. Manag. 2010, 18, 281–296. [Google Scholar] [CrossRef]
- Jones, J.W. Efficient wetland surface water detection and monitoring via Landsat: Comparison with in situ data from the Everglades Depth Estimation Network. Remote Sens. 2015, 7, 12503–12538. [Google Scholar] [CrossRef]
- Chapman, B.; McDonald, K.; Shimada, M.; Rosenqvist, A.; Schroeder, R.; Hess, L. Mapping regional inundation with spaceborne L-band SAR. Remote Sens. 2015, 7, 5440–5470. [Google Scholar] [CrossRef]
- Westerhoff, R.S.; Kleuskens, M.P.H.; Winsemius, H.C.; Huizinga, H.J.; Brakenridge, G.R.; Bishop, C. Automated global water mapping based on wide-swath orbital synthetic-aperture radar. Hydrol. Earth Syst. Sci. 2013, 17, 651–663. [Google Scholar] [CrossRef]
- Lewis, A.J. Geomorphic and hydrologic applications of active microwave remote sensing. In Principles and Applications of Imaging Radar, 3rd ed.; Henderson, F.M., Lewis, A.J., Eds.; John Wiley & Sons Inc.: New York, NY, USA, 1998; pp. 567–629. [Google Scholar]
- Pekel, J.F.; Cottam, A.; Gorelick, N.; Belward, A.S. Global Surface Water Explorer. Available online: https://global-surface-water.appspot.com (accessed on 7 December 2017).
- Senay, G.B. Modeling landscape evapotranspiration by integrating land surface phenology and a water balance algorithm. Algorithms 2008, 1, 52–68. [Google Scholar] [CrossRef]
- Wu, C.; Peng, D.; Soudani, K.; Siebicke, L.; Gough, C.M.; Arain, M.A.; Bohrer, G.; LaFleur, P.M.; Peichl, M.; Gonsamo, A.; et al. Land surface phenology derived from normalized difference vegetation index (NDVI) at global FLUXNET sites. Agric. For. Meteorol. 2017, 233, 171–182. [Google Scholar] [CrossRef]
- Hmimina, G.; Dufrêne, E.; Pontailler, J.Y.; Delpierre, N.; Aubinet, M.; Caquet, B.; de Grandcourt, A.; Burbane, B.; Flechard, C.; Granier, A.; et al. Evaluation of the potential of MODIS satellite data to predict vegetation phenology in different biomes: An investigation using ground-based NDVI measurements. Remote Sens. Environ. 2013, 132, 145–158. [Google Scholar] [CrossRef]
- Beck, P.S.A.; Atzberger, C.; Hogda, K.A.; Johansen, B.; Skidmore, A.K. Improved monitoring of vegetation dynamics at very high latitudes: A new method using MODIS NDVI. Remote Sens. Environ. 2006, 100, 321–334. [Google Scholar] [CrossRef]
- Atzberger, C.; Klisch, A.; Mattiuzzi, M.; Vuolo, F. Phenological metrics derived over the European continent from NDVI3g data and MODIS time series. Remote Sens. 2013, 6, 257–284. [Google Scholar] [CrossRef]
- Malingreau, J.P. Global vegetation dynamics: Satellite observations over Asia. Int. J. Remote Sens. 1986, 7, 1121–1146. [Google Scholar] [CrossRef]
- Horion, S.; Cornet, Y.; Erpicum, M.; Tychon, B. Studying interactions between climate variability and vegetation dynamic using a phenology based approach. Int. J. Appl. Earth Obs. Geoinf. 2012, 20, 20–32. [Google Scholar] [CrossRef]
- Balzarolo, M.; Vicca, S.; Nguy-Robertson, A.L.; Bonal, D.; Elbers, J.A.; Fu, Y.H.; Grunwald, T.; Horemans, J.A.; Paple, D.; Penuelas, J.; et al. Matching the phenology of net ecosystem exchange and vegetation indices estimated with MODIS and FLUXNET in-situ observations. Remote Sens. Environ. 2016, 174, 290–300. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–214. [Google Scholar] [CrossRef]
- Fensholt, R. Earth observation of vegetation status in the Sahelian and Sudanian West Africa: Comparison of Terra MODIS and NOAA AVHRR satellite data. Int. J. Remote Sens. 2004, 25, 1641–1659. [Google Scholar] [CrossRef]
- Peng, D.; Wu, C.; Li, C.; Zhang, X.; Liu, Z.; Ye, H.; Luo, S.; Liu, X.; Hu, Y.; Fang, B. Spring green-up phenology products derived from MODIS NDVI and EVI: Intercomparison, interpretation and validation using National Phenology Network and AmeriFlux observations. Ecol. Indic. 2017, 77, 323. [Google Scholar] [CrossRef]
- Luo, Z.; Yu, S. Spatiotemporal variability of land surface phenology in China from 2001–2014. Remote Sens. 2017, 9, 65. [Google Scholar] [CrossRef]
- De Beurs, K.M.; Henebry, G.M. Land surface phenology and temperature variation in the International Geosphere-Biosphere Program high-latitude transects. Glob. Chang. Biol. 2005, 11, 779–790. [Google Scholar] [CrossRef]
- Gonsamo, A.; Chen, J.M. Circumpolar vegetation dynamics product for global change study. Remote Sens. Environ. 2016, 182, 13–26. [Google Scholar] [CrossRef]
- Delbart, N.T.; Kergoat, L.; Fedotova, V. Remote sensing of spring phenology in boreal regions: A free of snow-effect method using NOAA-AVHRR and SPOT-VGT data (1982–2004). Remote Sens. Environ. 2006, 101, 52–62. [Google Scholar] [CrossRef]
- Meier, G.A.; Brown, J.F.; Evelsizer, R.J.; Vogelmann, J.E. Phenology and climate relationships in aspen (Populus tremuloides Michx.) forest and woodland communities of southwestern Colorado. Ecol. Indic. 2015, 48, 189–197. [Google Scholar] [CrossRef]
- Beck, P.S.A.; Jonsson, P.; Hogda, K.A.; Karlsen, S.R.; Eklundh, L.; Skidmore, A.K. A ground-validated NDVI dataset for monitoring vegetation dynamics and mapping phenology in Fennoscandia and the Kola Peninsul. Int. J. Remote Sens. 2007, 28, 4311–4330. [Google Scholar] [CrossRef]
- Bottcher, K.; Aurela, M.; Kervinen, M.; Markkanen, T.; Mattila, O.P.; Kolari, P.; Metsamaki, S.; Aalto, T.; Arsian, A.N.; Pulliainen, J. MODIS time-series-derived indicators for the beginning of the growing season in boreal coniferous forest—A comparison with CO2 flux measurements and phenological observations in Finland. Remote Sens. Environ. 2014, 140, 625–638. [Google Scholar] [CrossRef]
- Winter, T.C. (Ed.) Hydrological and Biogeochemical Research in the Shingobee River Headwaters Area, North-Central Minnesota; Water-Resources Investigations Report 96-4215; US Geological Survey: Denver, CO, USA, 1997; p. 213.
- Stark, J.R.; Armstrong, D.S.; Zwilling, D.R. Stream-Aquifer Interactions in the Straight River Area, Becker and Hubbard Counties; Water-Resources Investigations Report 94-4009; US Geological Survey: Mounds View, MN, USA, 1994; p. 93.
- Senay, G.B.; Friedrichs, M.; Singh, R.K.; Velpuri, N.M. Evaluating Landsat 8 evapotranspiration for water use mapping in the Colorado River Basin. Remote Sens. Environ. 2016, 185, 171–185. [Google Scholar] [CrossRef]
- Haynes, J.V.; Senay, G.B. Evaluation of the Relation between Evapotranspiration and Normalized Difference Vegetation Index for Downscaling the Simplified Surface Energy Balance Model; Scientific Investigations Report 2012-5197; US Geological Survey: Mounds View, MN, USA, 2012; p. 8.
- Turner, M.G.; Gardner, R.H. Landscape Ecology in Theory and Practice: Pattern and Process, 2nd ed.; Springer: New York, NY, USA, 2015. [Google Scholar]
- Wang, D.; Morton, D.C.; Masek, J.; Wu, A.; Nagol, J.; Xiong, X.; Levy, R.; Vermote, E.; Wolfe, R.E. Impact of sensor degradation on the MODIS NDVI time series. Remote Sens. Environ. 2012, 119, 55–61. [Google Scholar] [CrossRef]
- Doelling, D.R.; Wu, A.; Xiong, X.; Scarino, B.R.; Bhatt, R.; Haney, C.O.; Morstad, D.; Gopalan, A. The radiometric stability and scaling of Collection 6 Terra- and Aqua-MODIS VIS, NIR, and SWIR spectral bands. IEEE Trans. Geosci. Remote Sens. 2015, 53, 4520–4535. [Google Scholar] [CrossRef]
- Detsch, F.; Otte, I.; Appelhans, T.; Nauss, T. A comparative study of cross-product NDVI dynamics in the Kilimanjaro region—A matter of sensor, degradation calibration, and significance. Remote Sens. 2016, 8, 159–176. [Google Scholar] [CrossRef]
- Brummer, C.; Black, T.A.; Jassal, R.S.; Grant, N.J.; Spittlehouse, D.L.; Chen, B.; Nesic, Z.; Amiro, B.D.; Arain, M.A.; Barr, A.G.; et al. How climate and vegetation type influence evapotranspiration and water use efficiency in Canadian forest, peatland and grassland ecosystems. Agric. For. Meteorol. 2012, 153, 14–30. [Google Scholar] [CrossRef]
- Buyantuyev, A.; Wu, J. Urbanization diversifies land surface phenology in arid environments: Interactions among vegetation, climatic variation, and land use pattern in the Pheonex metropolitan region, USA. Landsc. Urban Plan. 2012, 105, 149–159. [Google Scholar] [CrossRef]
- Cook, B.I.; Shukla, S.P.; Puma, M.J.; Nazarenko, L.S. Irrigation as an historical climate forcing. Clim. Dyn. 2014. [Google Scholar] [CrossRef]
- Diffenbaugh, N.S.; Field, C.B. Changes in ecologically critical terrestrial climate conditions. Nat. Syst. Chang. Clim. 2013, 341, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Forkel, M.; Migliavacca, M.; Thonicke, K.; Reichstein, M.; Schaphoff, S.; Weber, U.; Carvalhais, N. Codominant water control on global interannual variability and trends in land surface phenology and greenness. Glob. Chang. Biol. 2015, 21, 3414–3435. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.Q.; Mao, F.; Sun, H.; Hou, Y.Y. Study of normalized difference vegetation index variation and its correlation with climate factors in the three-river-source region. Int. J. Appl. Earth Obs. Geoinf. 2011, 13, 24–33. [Google Scholar] [CrossRef]
- Karoly, D.J.; Braganza, K.; Stott, P.A.; Arblaster, J.M.; Meehl, G.A.; Broccoli, A.J.; Dixon, K.W. Detection of a human influence on North American climate. Sci. Total Environ. 2003, 302, 1200–1203. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.-L.; Brunsell, N. Assessing regional climate and local landcover impacts on vegetation with remote sensing. Remote Sens. 2013, 5, 4347–4369. [Google Scholar] [CrossRef]
- Mantyka-Pringle, C.S.; Martin, T.G.; Rhodes, J.R. Interactions between climate and habitat loss effects on biodiversity: A systematic review and meta-analysis. Glob. Chang. Biol. 2012, 18, 1239–1252. [Google Scholar] [CrossRef]
- Piao, S.; Tan, J.; Chen, A.; Fu, Y.H.; Ciais, P.; Liu, Q.; Janssens, I.A.; Vicca, S.; Zeng, Z.; Jeong, S.J.; et al. Leaf onset in the northern hemisphere triggered by daytime temperature. Nat. Commun. 2015, 6, 8. [Google Scholar] [CrossRef] [PubMed]
- Norman, S.P.; Hargrove, W.W.; Christie, W.M. Spring and autumn phenological variability across environmental gradients of Great Smoky Mountains National Park, USA. Remote Sens. 2017, 9, 407. [Google Scholar] [CrossRef]
- Doktor, D.; Bondeau, A.; Koslowski, D.; Badeck, F.W. Influence of heterogeneous landscapes on computed green-up dates based on daily AVHRR NDVI observations. Remote Sens. Environ. 2009, 113, 2618–2632. [Google Scholar] [CrossRef]
- Fisher, J.; Mustard, J.; Vadeboncoeur, M. Green leaf phenology at Landsat resolution: Scaling from the field to the satellite. Remote Sens. Environ. 2006, 100, 265–279. [Google Scholar] [CrossRef]
- Butz, P.; Raffelsbauer, V.; Graefe, S.; Peters, T.; Cueva, E.; Holscher, D.; Brauning, A. Tree responses to moisture fluctuations in a neotropical dry forest as potential climate change indicators. Ecol. Indic. 2017, 83, 559–571. [Google Scholar] [CrossRef]
- Belluau, M.; Shipley, B. Predicting habitat affinities of herbaceous dicots to soil wetness based on physiological traits of drought tolerance. Ann. Bot. 2017, 119, 1073–1084. [Google Scholar] [CrossRef] [PubMed]
- Tobin, M.F.; Lopez, O.R.; Kursar, T.A. Responses of tropical understory plants to a severe drought: Tolerance and avoidance of water stress. Biotropica 1999, 31, 570–578. [Google Scholar] [CrossRef]
- Hwang, T.; Gholizadeh, H.; Sims, D.A.; Novick, K.A.; Brzostek, E.R.; Phillips, R.P.; Roman, D.T.; Robeson, S.M.; Rahman, A.F. Capturing species-level drought responses in a temperate deciduous forest using ratios of photochemical reflectance indices between sunlit and shaded canopies. Remote Sens. Environ. 2017, 199, 350–359. [Google Scholar] [CrossRef]
- De Keersmaecker, W.; van Rooijen, N.; Lhermitte, S.; Tits, L.; Schaminee, J.; Coppin, P.; Honnay, O.; Somers, B. Species-rich semi-natural grasslands have a higher resistance but a lower resilience than intensively managed agricultural grasslands in repsonse to climate anomalies. J. Appl. Ecol. 2016, 53, 430–439. [Google Scholar] [CrossRef]
- Moore, G.W.; Edgar, C.B.; Vogel, J.G.; Washington-Allen, R.A.; March, R.G.; Zehnder, R. Tree mortality from an exceptional drought spanning mesic to semiarid ecoregions. Ecol. Appl. 2016, 26, 602–611. [Google Scholar] [CrossRef] [PubMed]
- Swain, S.; Wardlow, B.D.; Narumalani, S.; Rundquist, D.C.; Hayes, M.J. Relationships between vegetation indices and root zone soil moisture under maize and soybean canopies in the US Corn Belt: A comparative study using a close-range sensing approach. Int. J. Remote Sens. 2013, 34, 2814–2828. [Google Scholar] [CrossRef]
- Meinzer, O.E. Plants as Indicators of Ground Water; US Government Printing Office: Washington, DC, USA, 1927; p. 95.
- Andujar, E.; Krakauer, N.Y.; Yi, C.; Kogan, F. Ecosystem drought response timescales from thermal emission versus shortwave remote sensing. Adv. Meteorol. 2017, 2017, 10. [Google Scholar] [CrossRef]
- Vogelmann, J.E.; Gallant, A.L.; Shi, H.; Zhu, Z. Perspectives on monitoring gradual change across the continuity of Landsat sensors using time-series data. Remote Sens. Environ. 2016, 185, 258–270. [Google Scholar] [CrossRef]
- Zhu, Z. Change detection using landsat time series: A review of frequencies, preprocessing, algorithms, and applications. ISPRS J. Photogramm. Remote Sens. 2017, 130, 370–384. [Google Scholar] [CrossRef]
- US Landsat Analysis Ready Data (ARD). Available online: https://landsat.usgs.gov/ard (accessed on 14 December 2017).
- Harmonized Landsat Sentinel-2. Available online: https://hls.gsfc.nasa.gov (accessed on 14 December 2017).
- Turner, W. Sensing biodiversity. Science 2014, 346, 301–302. [Google Scholar] [CrossRef] [PubMed]

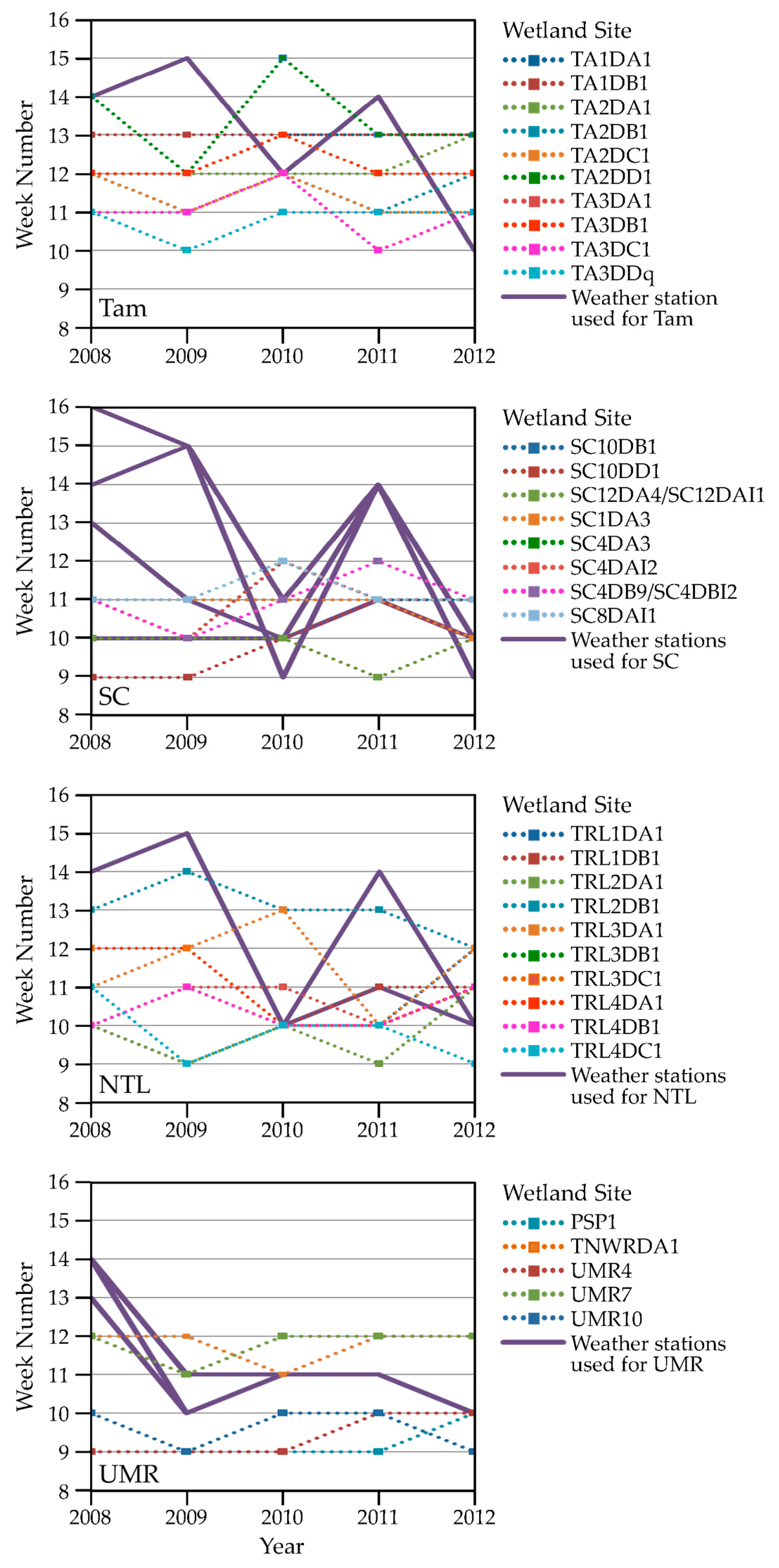



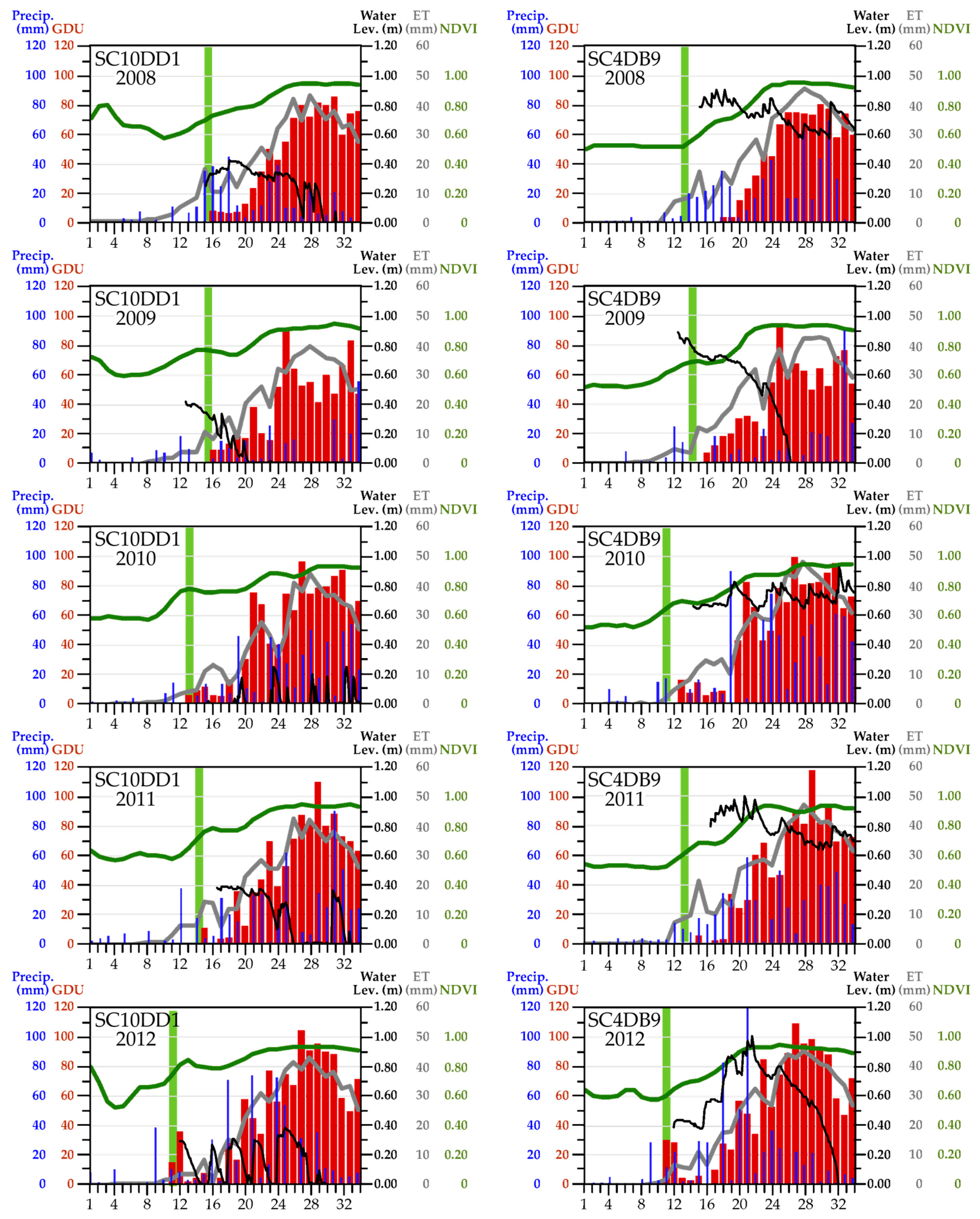
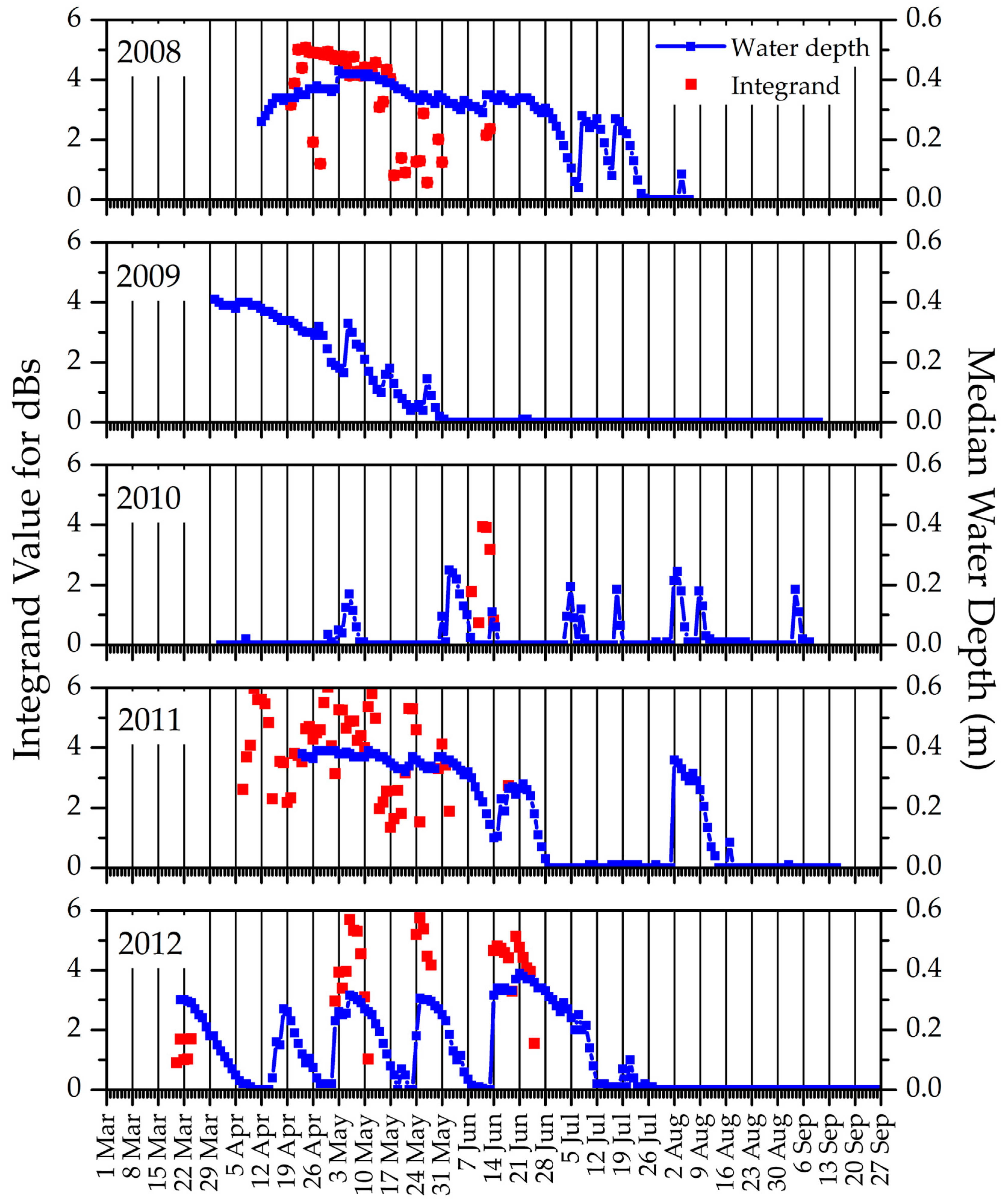
| Study Area | (a) Proportion (and Percentage) of Sites Where Snow Recurred after One Snow-Free Interval | (b) Proportion (and Percentage) of Sites Where Snow Recurred after Two Snow-Free Intervals | (c) Correlation between Timing of Snow-Off at the Cell vs. the Block Scale |
|---|---|---|---|
| Tam 1 | 18/50 (36%) | 2/50 (4%) | 0.98 |
| SC 2 | 10/50 (20%) | 0/50 (0%) | 0.99 |
| NTL 3 | 9/50 (18%) | 2/50 (4%) | 0.97 |
| UMR 4 | 7/25 (28%) | 0/25 (0%) | 0.93 |
| Study Area | SOST | altSOST |
|---|---|---|
| all areas | 0.2076 | 0.4271 |
| Tam 1 | −0.0011 | 0.6456 |
| SC 2 | 0.1701 | 0.3053 |
| NTL 3 | 0.3399 | 0.5007 |
| UMR 4 | 0.1571 | 0.3722 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallant, A.L.; Sadinski, W.; Brown, J.F.; Senay, G.B.; Roth, M.F. Challenges in Complementing Data from Ground-Based Sensors with Satellite-Derived Products to Measure Ecological Changes in Relation to Climate—Lessons from Temperate Wetland-Upland Landscapes. Sensors 2018, 18, 880. https://doi.org/10.3390/s18030880
Gallant AL, Sadinski W, Brown JF, Senay GB, Roth MF. Challenges in Complementing Data from Ground-Based Sensors with Satellite-Derived Products to Measure Ecological Changes in Relation to Climate—Lessons from Temperate Wetland-Upland Landscapes. Sensors. 2018; 18(3):880. https://doi.org/10.3390/s18030880
Chicago/Turabian StyleGallant, Alisa L., Walt Sadinski, Jesslyn F. Brown, Gabriel B. Senay, and Mark F. Roth. 2018. "Challenges in Complementing Data from Ground-Based Sensors with Satellite-Derived Products to Measure Ecological Changes in Relation to Climate—Lessons from Temperate Wetland-Upland Landscapes" Sensors 18, no. 3: 880. https://doi.org/10.3390/s18030880
APA StyleGallant, A. L., Sadinski, W., Brown, J. F., Senay, G. B., & Roth, M. F. (2018). Challenges in Complementing Data from Ground-Based Sensors with Satellite-Derived Products to Measure Ecological Changes in Relation to Climate—Lessons from Temperate Wetland-Upland Landscapes. Sensors, 18(3), 880. https://doi.org/10.3390/s18030880