
How Lazy Are Pet Cats Really? Using Machine Learning and Accelerometry to Get a Glimpse into the Behaviour of Privately Owned Cats in Different Households
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Owner and Cat Recruitment
2.2. Data Collection
2.3. Statistical Analysis
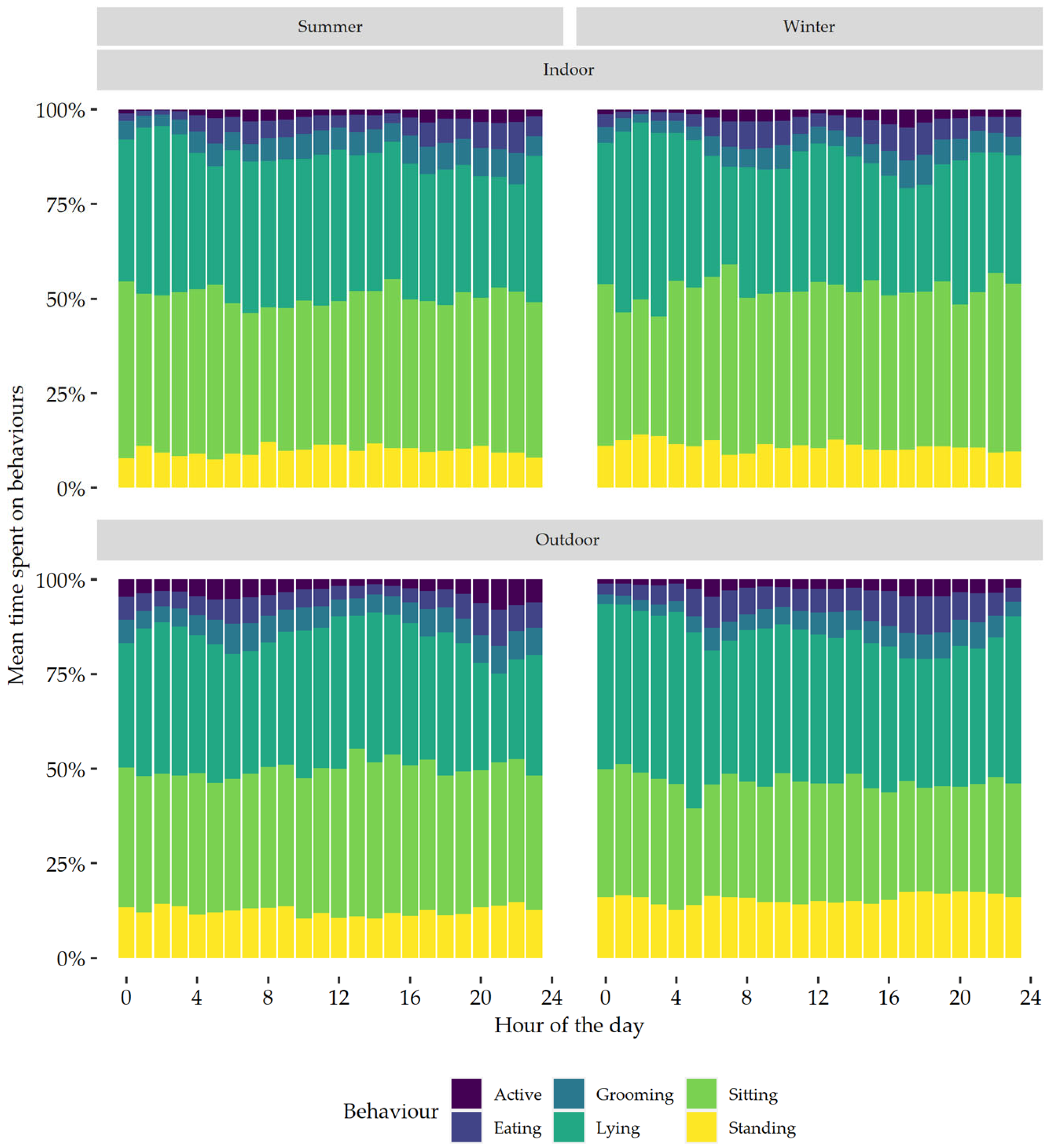
3. Results
3.1. Active
3.2. Eating
3.3. Grooming
3.4. Littering
3.5. Lying
3.6. Scratching
3.7. Sitting
3.8. Standing
3.9. Daily Pattern of Behaviour
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Predictor Variable | Description |
|---|---|
| Mean | Mean, calculated for every second using the raw acceleration data (30 measures per second) |
| Sum | Sum, calculated for every second using the raw acceleration data |
| Minimum (min) | Minimum value of every 30 measures within each second |
| Maximum (max) | Maximum value of every 30 measures within each second |
| Standard deviation (sd) | Measures the spread of the data |
| Skewness | Asymmetry of the distribution |
| Kurtosis | Weight of the tails relative to a normal distribution |
| Vector magnitude (VM) | |
| Overall dynamic body acceleration (ODBA) | |
| Dynamic body acceleration (DBA) 1 |
References
- Companion Animals New Zealand. Companion Animals in New Zealand 2020; Companion Animals New Zealand: Auckland, New Zealand, 2020. [Google Scholar]
- Foreman-Worsley, R.; Farnworth, M.J. A systematic review of social and environmental factors and their implications for indoor cat welfare. Appl. Anim. Behav. Sci. 2019, 220, 104841. [Google Scholar] [CrossRef]
- Judith, L.S.; Candace, C.C. Environmental Aspects of Domestic Cat Care and Management: Implications for Cat Welfare. Sci. World J. 2016, 2016, 6296315. [Google Scholar] [CrossRef]
- Broom, D.M. Indicators of poor welfare. Br. Vet. J. 1986, 142, 524–526. [Google Scholar] [CrossRef]
- Mellor, D.J.; Reid, C.S.W. Concepts of animal well-being and predicting the impact of procedures on experimental animals. In Improving the Well-Being of Animals in the Research Environment; New Zealand Council for the Care of Animals in Research and Teaching (ANZCCART): Glen Osmond, Australia, 1994; pp. 3–18. [Google Scholar]
- Levitis, D.A.; Lidicker, W.Z.; Freund, G. Behavioural biologists don’t agree on what constitutes behaviour. Anim. Behav. 2009, 78, 103–110. [Google Scholar] [CrossRef]
- Vigne, J.D.; Debue, K.; Guilaine, J.; Haye, L.; Gérard, P. Early Taming of the Cat in Cyprus. Science 2004, 304, 259. [Google Scholar] [CrossRef] [PubMed]
- Amat, M.; de la Torre, J.L.R.; Fatjó, J.; Mariotti, V.M.; Van Wijk, S.; Manteca, X. Potential risk factors associated with feline behaviour problems. Appl. Anim. Behav. Sci. 2009, 121, 134–139. [Google Scholar] [CrossRef]
- Sandøe, P.; Nørspang, A.P.; Forkman, B.; Kondrup, S.V.; Lund, T.B.; Bjørnvad, C.R. The burden of domestication: A representative study of welfare in privately owned cats in Denmark. Anim. Welf. 2017, 26, 1–10. [Google Scholar] [CrossRef]
- Adamelli, S.; Marinelli, L.; Normando, S.; Bono, G. Owner and cat features influence the quality of life of the cat. Appl. Anim. Behav. Sci. 2005, 94, 89–98. [Google Scholar] [CrossRef]
- Martin, P.; Bateson, P. Measuring Behaviour: An Introductory Guide, 2nd ed.; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Altmann, J. Observational Study of Behavior: Sampling Methods. Behaviour 1974, 49, 227–267. [Google Scholar] [CrossRef]
- Brown, D.D.; Kays, R.; Wikelski, M.; Wilson, R.; Klimley, A.P. Observing the unwatchable through acceleration logging of animal behavior. Anim. Biotelemetry 2013, 1, 20. [Google Scholar] [CrossRef]
- Watanabe, S.; Izawa, M.; Kato, A.; Ropert-Coudert, Y.; Naito, Y. A new technique for monitoring the detailed behaviour of terrestrial animals: A case study with the domestic cat. Appl. Anim. Behav. Sci. 2005, 94, 117–131. [Google Scholar] [CrossRef]
- Kestler, J.; Wilson, M. Acceleration Derived Feral Cat (Felis catus) Behaviour during Ground Nesting Bird-Breeding Season on the Island of Schiermonnikoog. BSc. Thesis, Van Hall Larenstein, University of Applied Sciences, Leeuwarden, The Netherlands, 2015. [Google Scholar]
- Galea, N.; Murphy, F.; Gaschk, J.L.; Schoeman, D.S.; Clemente, C.J. Quantifying finer-scale behaviours using self-organising maps (SOMs) to link accelerometery signatures with behavioural patterns in free-roaming terrestrial animals. Sci. Rep. 2021, 11, 13566. [Google Scholar] [CrossRef]
- Smit, M.; Ikurior, S.J.; Corner-Thomas, R.A.; Andrews, C.J.; Thomas, D.G. The Use of Triaxial Accelerometers and Machine Learning Algorithms for Behavioural Identification in Domestic Cats (Felis catus): A Validation Study. Sensors 2023, 23, 7165. [Google Scholar] [CrossRef]
- Laflamme, D. Development and validation of a body condition score system for cats: A clinical tool. Feline Pract. 1997, 25, 13–18. [Google Scholar]
- RStudio Team. RStudio: Integrated Development for R, version 4.1.1; RStudio: Boston, MA, USA, 2021. [Google Scholar]
- Pittari, J.; Rodan, I.; Beekman, G.; Gunn-Moore, D.; Polzin, D.; Taboada, J.; Tuzio, H.; Zoran, D. American Association of Feline Practitioners. Senior Care Guidelines. J. Feline Med. Surg. 2009, 11, 763–778. [Google Scholar] [CrossRef]
- Vogt, A.H.; Rodan, I.; Brown, M.; Brown, S.; Buffington, C.A.T.; Forman, M.J.L.; Neilson, J.; Sparkes, A. AAFP-AAHA: Feline Life Stage Guidelines. J. Feline Med. Surg. 2010, 12, 43–54. [Google Scholar] [CrossRef]
- Time and Date. Palmerston North, New Zealand—Sunrise, Sunset, and Daylength. Available online: https://www.timeanddate.com/sun/new-zealand/palmerston-north (accessed on 29 November 2023).
- Andrews, C.J. The Assessment of Activity in Colony-Housed Domestic Cats (Felis catus): A Thesis Presented in Partial Fulfilment of the Requirements for the Degree of Masters of Science in Zoology, Massey University, Palmerston North, New Zealand. Master’s Thesis, Massey University, Palmerston North, New Zealand, 2015. [Google Scholar]
- Smit, M.; Corner-Thomas, R.A.; Weidgraaf, K.; Thomas, D.G. Association of age and body condition with physical activity of domestic cats (Felis catus). Appl. Anim. Behav. Sci. 2022, 248, 105584. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Dogs and Cats; National Academies Press: Washington, DC, USA, 2006. [Google Scholar]
- Palestrini, C.; Minozzi, G.; Mazzola, S.M.; Lopez, A.; Cannas, S. Do intense weather events influence dogs’ and cats’ behavior? Analysis of owner reported data in Italy. Front. Vet. Sci. 2022, 9, 973574. [Google Scholar] [CrossRef] [PubMed]
- Flux, J.E.C. Seventeen years of predation by one suburban cat in New Zealand. N. Z. J. Zool. 2007, 34, 289–296. [Google Scholar] [CrossRef]
- Hansen, C.M. Movements and Predation Activity of Feral and Domestic cats (Felis catus) on Banks Peninsula. Ph.D. Thesis, Lincoln University, Lincoln, New Zealand, 2010. [Google Scholar]
- Krauze-Gryz, D.; Żmihorski, M.; Gryz, J. Annual variation in prey composition of domestic cats in rural and urban environment. Urban Ecosyst. 2017, 20, 945–952. [Google Scholar] [CrossRef]
- Berteselli, G.V.; Regaiolli, B.; Normando, S.; De Mori, B.; Zaborra, C.A.; Spiezio, C. European wildcat and domestic cat: Do they really differ? J. Vet. Behav. 2017, 22, 35–40. [Google Scholar] [CrossRef]
- Piccione, G.; Marafioti, S.; Giannetto, C.; Panzera, M.; Fazio, F. Daily rhythm of total activity pattern in domestic cats (Felis silvestris catus) maintained in two different housing conditions. J. Vet. Behav. Clin. Appl. Res. 2013, 8, 189–194. [Google Scholar] [CrossRef]
- Naik, R.; Witzel, A.; Albright, J.D.; Siegfried, K.; Gruen, M.E.; Thomson, A.; Lascelles, B.D.X.; Price, J. Pilot study evaluating the effect of feeding method on overall activity of neutered indoor pet cats. J. Vet. Behav. 2018, 25, 9–13. [Google Scholar] [CrossRef]
- Andrews, C.J.; Potter, M.A.; Thomas, D.G. Quantification of activity in domestic cats (Felis catus) by accelerometry. Appl. Anim. Behav. Sci. 2015, 173, 17–21. [Google Scholar] [CrossRef]
- Lascelles, B.D.X.; Hansen, B.D.; Thomson, A.; Pierce, C.C.; Boland, E.; Smith, E.S. Evaluation of a digitally integrated accelerometer-based activity monitor for the measurement of activity in cats. Vet. Anaesth. Analg. 2008, 35, 173–183. [Google Scholar] [CrossRef]
- Sordo, L.; Breheny, C.; Tørnqvist-Johnsen, C.; Gunn-Moore, D.A.; Halls, V.; Cotter, A.; Caney, S.M.A. Prevalence of disease and age-related behavioural changes in cats: Past and present. Vet. Sci. 2020, 7, 85. [Google Scholar] [CrossRef] [PubMed]
- Hart, L.A.; Hart, B.L.; Thigpen, A.P.; Willits, N.H.; Lyons, L.A.; Hundenski, S. Compatibility of Cats with Children in the Family. Front. Vet. Sci. 2018, 5, 278. [Google Scholar] [CrossRef]
- Kappen, K.L.; Garner, L.M.; Kerr, K.R.; Swanson, K.S. Effects of photoperiod on food intake, activity and metabolic rate in adult neutered male cats. J. Anim. Physiol. Anim. Nutr. 2014, 98, 958–967. [Google Scholar] [CrossRef]
- Parker, M.; Lamoureux, S.; Challet, E.; Deputte, B.; Biourge, V.; Serra, J. Daily rhythms in food intake and locomotor activity in a colony of domestic cats. Anim. Biotelemetry 2019, 7, 25. [Google Scholar] [CrossRef]
- Walker, W.H.; Meléndez-Fernández, O.H.; Nelson, R.J.; Reiter, R.J. Global climate change and invariable photoperiods: A mismatch that jeopardizes animal fitness. Ecol. Evol. 2019, 9, 10044–10054. [Google Scholar] [CrossRef]
- Hall, C.M.; Bryant, K.A.; Calver, M.C.; Haskard, K.; Major, T.; Bruce, S. Factors determining the home ranges of pet cats: A meta-analysis. Biol. Conserv. 2016, 203, 313–320. [Google Scholar] [CrossRef]
- Lilith, M.; Calver, M.; Garkaklis, M. Roaming habits of pet cats on the suburban fringe in Perth, Western Australia: What size buffer zone is needed to protect wildlife in reserves? Aust. Zool. 2008, 34, 65–72. [Google Scholar] [CrossRef]
- Smit, M. Weekly Data Cats for Home Trial. Available online: https://figshare.com/articles/dataset/How_lazy_are_pet_cats_really_Using_machine_learning_and_accelerometry_to_get_a_glimpse_into_the_behaviour_of_privately_owned_cats_in_different_households/24848292 (accessed on 18 December 2023).





| Training Day | ||||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | Day 7–14 data collection |
| 2 h | 4 h | 6 h | 8 h | 24 h | Off | |
| Variable | Categories |
|---|---|
| Season | Summer (December–February) Winter (June–August) |
| Sex of cat | Entire female Entire male Neutered female Neutered male |
| Age group [20,21] | Kitten (0–6 months) Junior (7 months–2 years) Prime (3–6 years) Mature (7–10 years) Senior (11–14 years) Geriatric (≥15 years) |
| Diet | Dry Wet (e.g., canned, pouched, or raw) Mix (mix of dry and wet foods) |
| Housing | Exclusively indoors Indoors with limited outdoor access (e.g., harnessed walks, catio, or garden access) Indoors with unlimited outdoor access Exclusively outdoors Other |
| Rural vs. urban | Rural Urban |
| Multi-cat vs. single-cat household | Multi Single |
| Presence of at least one dog | No (absent; no dog(s) in household) Yes (present; at least one dog in household) |
| Presence of at least one child (<18 years) | No (absent; no child(ren) in household) Yes (present; at least one child in household) |
| Season | Season | ||||||
|---|---|---|---|---|---|---|---|
| Summer | Winter | Summer | Winter | ||||
| Sex | Housing | ||||||
| Female | 17 | 17 | Indoor 2 | 10 | 10 | ||
| Male | 11 | 11 | Outdoor 2 | 18 | 18 | ||
| Age group | Number of cats in household | ||||||
| Junior 1 | 10 | 10 | One | 12 | 11 | ||
| Prime 1 | 12 | 12 | Two | 9 | 10 | ||
| Mature 1 | 6 | 6 | Three | 7 | 7 | ||
| Coat length | Dogs | ||||||
| Long | 5 | 5 | Absent | 19 | 19 | ||
| Short | 23 | 23 | Present | 9 | 9 | ||
| Environment | Children (<18 years) | ||||||
| Rural | 7 | 7 | Absent | 20 | 18 | ||
| Urban | 21 | 21 | Present | 8 | 10 | ||
| Behaviour | ||||||||
|---|---|---|---|---|---|---|---|---|
| Active | Eating | Grooming | Littering | Lying | Scratching | Sitting | Standing | |
| Season × Sex of the cat | NS | NS | 0.023 | NS | NS | 0.079 | NS | NS |
| Season × Age group | <0.001 | NS | NS | 0.030 | NS | NS | NS | 0.002 |
| Season × Diet | NS | NS | NS | 0.063 | NS | NS | 0.055 | NS |
| Season × Rural vs. urban | NS | <0.001 | NS | NS | NS | 0.013 | NS | NS |
| Season × Housing | <0.001 | NS | NS | NS | NS | NS | NS | NS |
| Season × Multi- vs. single-cat household | NS | NS | NS | NS | 0.064 | NS | NS | NS |
| Season × Dog(s) in household | NS | NS | NS | NS | NS | NS | NS | NS |
| Season × Child(ren) in household | NS | NS | NS | NS | 0.006 | NS | 0.0016 | <0.001 |
| Behaviour | ||||||||
|---|---|---|---|---|---|---|---|---|
| Active | Eating | Grooming | Littering | Lying | Scratching | Sitting | Standing | |
| Season | 0.006 | NS | 0.046 | NS | NS | NS | NS | 0.082 |
| Sex of the cat | NS | 0.017 | NS | NS | 0.049 | NS | 0.090 | NS |
| Age group | <0.001 | 0.066 | 0.057 | NS | NS | NS | NS | NS |
| Diet | NS | 0.017 | NS | NS | 0.015 | NS | <0.001 | NS |
| Rural vs. urban | NS | <0.001 | NS | NS | 0.092 | NS | 0.027 | NS |
| Housing | <0.001 | NS | NS | NS | NS | NS | NS | NS |
| Multi- vs. single-cat household | NS | NS | NS | NS | 0.032 | NS | 0.006 | NS |
| Dog(s) in household | NS | NS | NS | NS | NS | NS | NS | NS |
| Child(ren) in household | NS | 0.051 | 0.090 | NS | 0.005 | NS | 0.002 | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smit, M.; Corner-Thomas, R.A.; Draganova, I.; Andrews, C.J.; Thomas, D.G. How Lazy Are Pet Cats Really? Using Machine Learning and Accelerometry to Get a Glimpse into the Behaviour of Privately Owned Cats in Different Households. Sensors 2024, 24, 2623. https://doi.org/10.3390/s24082623
Smit M, Corner-Thomas RA, Draganova I, Andrews CJ, Thomas DG. How Lazy Are Pet Cats Really? Using Machine Learning and Accelerometry to Get a Glimpse into the Behaviour of Privately Owned Cats in Different Households. Sensors. 2024; 24(8):2623. https://doi.org/10.3390/s24082623
Chicago/Turabian StyleSmit, Michelle, Rene A. Corner-Thomas, Ina Draganova, Christopher J. Andrews, and David G. Thomas. 2024. "How Lazy Are Pet Cats Really? Using Machine Learning and Accelerometry to Get a Glimpse into the Behaviour of Privately Owned Cats in Different Households" Sensors 24, no. 8: 2623. https://doi.org/10.3390/s24082623