Time-varying Brain Potentials and Interhemispheric Coherences of Anterior and Posterior Regions during Repetitive Unimanual Finger Movements

Abstract
:1. Introduction
2. Methods
2.1. Participants
2.2. Variables
2.3. Experimental Design
2.4. Stimulus presentation and key pressing performance
2.5. Experimental Procedure
2.6. Electroencephalogram (EEG) acquisition and ERP recording
2.7. Calculating the coherences
2.8. Statistics
3. Results
3.1. Behavioral results
3.3. A comparison of coherence strength between inter-hemispheric pairs
3.3.1. Early visual phase
3.3.2. Pre-execution, execution, and post-execution phases
3.4. The correlation coefficients between RT and inter-hemispheric coherence pairs
4. Discussion and Conclusions
Acknowledgments
Appendix 1

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coherence pairs | Early visual | Pre-execution | Execution | Post-execution |
|---|---|---|---|---|
| Coh(F3, F4) | .102 | .163 | .175 | .105 |
| Coh(F3, C3) | .177 | .270 | .252 | .126 |
| Coh(F3, C4) | .104 | .192 | .195 | .110 |
| Coh(F3, P3) | .100 | .163 | .161 | .083 |
| Coh(F3, P4) | .086 | .170 | .143 | .073 |
| Coh(F3, O1) | .021 | .055 | .051 | .030 |
| Coh(F3, O2) | .027 | .062 | .076 | .036 |
| Coh(F4, C3) | .126 | .158 | .142 | .095 |
| Coh(F4, C4) | .211 | .283 | .239 | .189 |
| Coh(F4, P3) | .062 | .112 | .098 | .071 |
| Coh(F4, P4) | .088 | .176 | .137 | .100 |
| Coh(F4, O1) | .019 | .038 | .035 | .028 |
| Coh(F4, O2) | .024 | .044 | .056 | .039 |
| Coh(C3,C4) | .223 | .342 | .319 | .259 |
| Coh(C3, P3) | .314 | .454 | .457 | .388 |
| Coh(C3, P4) | .146 | .271 | .254 | .198 |
| Coh(C3, O1) | .041 | .112 | .104 | .090 |
| Coh(C3, O2) | .029 | .094 | .111 | .075 |
| Coh(C4, P3) | .135 | .253 | .232 | .206 |
| Coh(C4, P4) | .394 | .525 | .510 | .510 |
| Coh(C4, O1) | .036 | .097 | .081 | .090 |
| Coh(C4, O2) | .052 | .114 | .131 | .123 |
| Coh(P3, P4) | .229 | .319 | .303 | .264 |
| Coh(P3, O1) | .276 | .304 | .288 | .302 |
| Coh(P3, O2) | .122 | .173 | .176 | .143 |
| Coh(P4, O1) | .112 | .188 | .184 | .175 |
| Coh(P4, O2) | .345 | .359 | .372 | .360 |
| Coh(O1, O2) | .243 | .258 | .250 | .222 |
Appendix 2
| Coherence pairs | Early visual | Pre-execution | Execution | Post-execution | ||||
|---|---|---|---|---|---|---|---|---|
| r | p | r | p | r | p | r | p | |
| Coh(F3, F4) | -.115 | .672 | -.306 | .249 | -.047 | .863 | .021 | .940 |
| Coh(F3, C3) | -.187 | .488 | -.359 | .172 | -.141 | .602 | -.347 | .188 |
| Coh(F3, C4) | -.206 | .444 | -.191 | .478 | -.329 | .213 | -.097 | .721 |
| Coh(F3, P3) | -.053 | .846 | -.247 | .356 | -.188 | .485 | -.209 | .438 |
| Coh(F3, P4) | -.087 | .749 | -.150 | .579 | -.228 | .395 | -.159 | .557 |
| Coh(F3, O1) | -.153 | .571 | -.361 | .170 | -.253 | .345 | -.305 | .251 |
| Coh(F3, O2) | .222 | .408 | -.475 | .063 | -.165 | .542 | -.156 | .564 |
| Coh(F4, C3) | .024 | .931 | -.097 | .721 | -.100 | .713 | -.206 | .444 |
| Coh(F4, C4) | -.053 | .846 | .388 | .137 | -.079 | .770 | -.383 | .144 |
| Coh(F4, P3) | -.215 | .425 | -.074 | .787 | -.174 | .520 | -.197 | .464 |
| Coh(F4, P4) | -.015 | .957 | .282 | .289 | .050 | .854 | -.288 | .279 |
| Coh(F4, O1) | -.477 | .062 | .001 | .996 | -.085 | .753 | -.241 | .368 |
| Coh(F4, O2) | -.397 | .128 | -.271 | .311 | .021 | .940 | -.285 | .284 |
| Coh(C3,C4) | -.418 | .107 | -.306 | .249 | -.585* | .017 | -.300 | .259 |
| Coh(C3, P3) | -.129 | .633 | -.012 | .966 | -.506* | .046 | -.388 | .137 |
| Coh(C3, P4) | -.344 | .192 | -.321 | .226 | -.432 | .094 | -.226 | .399 |
| Coh(C3, O1) | -.353 | .180 | -.524* | .037 | -.438 | .090 | -.397 | .128 |
| Coh(C3, O2) | -.015 | .957 | -.468 | .068 | -.366 | .163 | -.409 | .116 |
| Coh(C4, P3) | -.068 | .803 | -.197 | .464 | -.276 | .300 | -.071 | .795 |
| Coh(C4, P4) | .165 | .542 | .174 | .520 | -.050 | .854 | -.038 | .888 |
| Coh(C4, O1) | -.376 | .151 | -.285 | .284 | -.406 | .119 | -.129 | .633 |
| Coh(C4, O2) | -.159 | .557 | -.444 | .085 | -.282 | .289 | -.168 | .535 |
| Coh(P3, P4) | -.429 | .097 | -.376 | .151 | -.444 | .085 | -.300 | .259 |
| Coh(P3, O1) | -.544* | .029 | -.374 | .154 | -.218 | .418 | -.362 | .169 |
| Coh(P3, O2) | -.374 | .154 | -.388 | .137 | -.385 | .141 | -.394 | .131 |
| Coh(P4, O1) | -.624* | .010 | -.362 | .169 | -.318 | .231 | -.171 | .528 |
| Coh(P4, O2) | -.353 | .180 | -.315 | .235 | -.335 | .204 | -.238 | .374 |
| Coh(O1, O2) | -.088 | .745 | -.082 | .762 | -.432 | .094 | -.156 | .564 |
Appendix 3

References
- Knyazeva, M.; Koeda, T.; Njiokiktjien, C.; Jonkman, E.J.; Kurganskaya, M.; de Sonneville, L.; Vildavsky, V. EEG coherence changes during finger tapping in acallosal and normal children: a study of inter- and intrahemispheric connectivity. Behav. Brain Res. 1997, 89, 243–258. [Google Scholar]
- Knyazeva, M.G.; Kurganskaya, M.E.; Kurgansky, A.V.; Njiokiktjien, C.J.; Vildavsky, V.J. Interhemispheric interaction in children of 7-8: analysis of EEG coherence and finger tapping parameters. Behav. Brain Res. 1994, 61, 47–58. [Google Scholar]
- Manganotti, P.; Gerloff, C.; Toro, C.; Katsuta, H.; Sadato, N.; Zhuang, P.; Leocani, L.; Hallett, M. Task-related coherence and task-related spectral power changes during sequential finger movements. Electroen. Clin. Neuro. 1998, 109, 50–62. [Google Scholar]
- Rissman, J.; Gazzaley, A.; D'Esposito, M. Measuring functional connectivity during distinct stages of a cognitive task. Neuroimage 2004, 23, 752–763. [Google Scholar]
- Stancak, A.; Lucking, C.H.; Kristeva-Feige, R. The size of corpus callosum and functional connectivities of cortical regions in finger and shoulder movements. Cogn. Brain Res. 2002, 13, 61–74. [Google Scholar]
- Bai, O.; Mari, Z.; Vorbach, S.; Hallett, M. Asymmetric spatiotemporal patterns of event-related desynchronization preceding voluntary sequential finger movements: A high-resolution EEG study. Clin. Neurophysiol. 2005, 116, 1213–1221. [Google Scholar]
- Lewis, P.A.; Wing, A.M.; Pope, P.A.; Praamstra, P.; Miall, R.C. Brain activity correlates differentially with increasing temporal complexity of rhythms during initialisation, synchronisation, and continuation phases of paced finger tapping. Neuropsychologia 2004, 42, 1301–1312. [Google Scholar]
- Pfurtscheller, G.; Neuper, C.; Berger, J. Source localization using event-related desynchronization (ERD) within the alpha band. Brain Topogr. 1994, 6, 269–275. [Google Scholar]
- Sailer, A.; Dichgans, J.; Gerloff, C. The influence of normal aging on the cortical processing of a simple motor task. Neurology 2000, 55, 979–985. [Google Scholar]
- Andrew, C.; Pfurtscheller, G. Event-related coherence during finger movement: a pilot study. Biomed. Tech. 1995, 40, 326–332. [Google Scholar]
- Roth, N.; Battig, K. Effects of cigarette smoking upon frequencies of EEG alpha rhythm and finger tapping. Psychopharmacology 1991, 105, 186–190. [Google Scholar]
- Deiber, M.P.; Caldara, R.; Ibanez, V.; Hauert, C.A. Alpha band power changes in unimanual and bimanual sequential movements, and during motor transitions. Clin. Neurophysiol. 2001, 112, 1419–1135. [Google Scholar]
- Kudo, K.; Miyazaki, M.; Kimura, T.; Yamanaka, K.; Kadota, H.; Hirashima, M.; Nakajima, Y.; Nakazawa, K.; Ohtsuki, T. Selective activation and deactivation of the human brain structures between speeded and precisely timed tapping responses to identical visual stimulus: an fMRI study. Neuroimage 2004, 22, 1291–301. [Google Scholar]
- van der Lubbe, R.H.; Wauschkuhn, B.; Wascher, E.; Niehoff, T.; Kompf, D.; Verleger, R. Lateralized EEG components with direction information for the preparation of saccades versus finger movements. Exp. Brain Res. 2000, 132, 163–178. [Google Scholar]
- Babiloni, C.; Babiloni, F.; Carducci, F.; Cincotti, F.; Cocozza, G.; Del Percio, C.; Moretti, D.V.; Rossini, P.M. Human cortical electroencephalography (EEG) rhythms during the observation of simple aimless movements: a high-resolution EEG study. Neuroimage 2002, 17, 559–572. [Google Scholar]
- Babiloni, C.; Del Percio, C.; Babiloni, F.; Carducci, F.; Cincotti, F.; Moretti, D.V.; Rossini, P.M. Transient human cortical responses during the observation of simple finger movements: a high-resolution EEG study. Hum. Brain Map. 2003, 20, 148–157. [Google Scholar]
- Olfield, R. C. The assessment and analysis of handedness: The Edinburgh handedness inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar]
- Ehrenstein, W. H.; Arnold-Schulz-Gahmen, B. E.; Jaschinski, W. Eye preference within the context of binocular function. Graefe's Arch. Clin. Exp. Ophthalmol. 2005, 243, 926–932. [Google Scholar]
- Mendola, J. D.; Conner, I. P. Eye dominance predicts fMRI signals in human retinotopic cortex. Neurosci. Lett. 2007, 414, 30–34. [Google Scholar]
- Meng, L. F.; Lu, C. P.; Chen, B. W.; Chen, C. H. Fatigue induced reversed hemispheric plasticity: A brain electrophysiological study. Lecture Notes in Computer Science: Neural Information Processing 2006, 4232, 65–71. [Google Scholar]
- Meng, L. F.; Lu, C. P.; Chen, C. H. Unskilled finger key pressing and brain coherence. Lecture Notes in Computer Science: Computers Helping People with Special Needs 2006, 4061, 437–441. [Google Scholar]
- Brain Products GmbH. In Brain Vision Analyzer User Manual; 2002; BrainProducts: Munich; pp. 55–56.
- Geisser, S.; Greenhouse, S. On methods in the analysis of profile data. Psychometrika 1959, 24, 95–112. [Google Scholar]
- Calmels, C.; Holmes, P.; Jarry, G.; Hars, M.; Lopez, E.; Paillard, A.; Stam, C. J. Variability of EEG synchronization prior to and during observation and execution of a sequential finger movement. Hum. Brain Map. 2006, 27, 251–266. [Google Scholar]
- Pollok, B.; Gross, J.; Muller, K.; Aschersleben, G.; Schnitzler, A. The cerebral oscillatory network associated with auditorily paced finger movements. Neuroimage 2004, 24, 646–655. [Google Scholar]
- Kristeva, R.; Chakarov, V.; Wagner, M.; Schulte-Monting, J.; Hepp-Reymond, M.C. Is the movement-evoked potential mandatory for movement execution? A high-resolution EEG study in a deafferented patient. Neuroimage 2006, 31, 677–685. [Google Scholar]






| Early visual | Pre-execution | Execution | Post-execution | |
|---|---|---|---|---|
| Mid-line | ||||
| Fz | -0.791 | -0.654 | 0.487 | -1.410 |
| Cz | -1.033 | 0.169 | 0.464 | -2.010 |
| Pz | 0.768 | 0.989 | -0.652 | -2.523 |
| Oz | 3.441 | 0.571 | -1.124 | -0.734 |
| F-value | F(1.246, 18.695) | F(1.634, 24.513) | F(1.991, 29.860) | F(1.522, 22.837) |
| = 23.376; p=.000 | = 6.833; p=.007 | = 11.496; p=.000 | = 15.670; p=.000 | |
| Left | ||||
| F3 | -0.174 | -0.485 | 0.375 | -1.129 |
| C3 | -0.460 | 0.459 | 0.562 | -1.805 |
| P3 | 1.467 | 1.741 | -0.061 | -2.203 |
| O1 | 3.090 | 1.161 | -1.004 | -0.907 |
| F-value | F(1.269, 19.035) | F(1.461, 21.910) | F(1.636,24.534) | F(1.608, 24.124) |
| = 16.094; p=.000 | = 19.628; p=.000 | = 8.637; p=.002 | = 13.453; p=.000 | |
| Right | ||||
| F4 | -0.147 | -0.424 | -0.112 | -1.243 |
| C4 | -0.433 | 1.163 | -0.200 | -2.104 |
| P4 | 1.436 | 2.063 | -1.165 | -2.129 |
| O2 | 3.687 | 1.284 | -1.669 | -0.975 |
| F-value | F(1.113, 16.700) | F(1.919, 28.781) | F(1.840, 27.594) | F(1.414, 21.210) |
| = 16.896; p=001 | = 18.599; p=.000 | =7.127; p=.004 | = 13.543; p=.001 |
| Early visual | Pre-execution | Execution | Post-execution | |||||
|---|---|---|---|---|---|---|---|---|
| Mean Difference | p | Mean Difference | p | Mean Difference | p | Mean Difference | p | |
| Oz vs. Pz | 2.673** | 001 | -0.417 | .250 | -0.473 | .138 | 1.790*** | .000 |
| Oz vs. Cz | 4.474*** | .000 | 0.402 | .403 | -1.588*** | .000 | 1.276** | .001 |
| Oz vs. Fz | 4.232*** | .000 | 1.226* | .034 | -1.612** | .002 | 0.676 | .096 |
| Pz vs. Cz | 1.801*** | .000 | 0.819** | .005 | -1.116*** | .000 | -0.514* | .032 |
| Pz vs. Fz | 1.559** | .001 | 1.643** | .001 | -1.139* | .014 | -1.113** | .003 |
| Cz vs. Fz | -0.243 | .202 | 0.823** | .001 | -0.024 | .925 | -0.600** | .001 |
| Early visual | Pre-execution | Execution | Post-execution | |||||
|---|---|---|---|---|---|---|---|---|
| Mean Difference | p | Mean Difference | p | Mean Difference | p | Mean Difference | p | |
| O1 vs. P3 | 1.623* | .021 | -0.580* | .028 | -0.942** | .002 | 1.296*** | .000 |
| O1 vs. C3 | 3.550** | .001 | 0.702 | .114 | -1.566** | .001 | 0.898** | .005 |
| O1 vs. F3 | 3.265** | .001 | 1.646** | .001 | -1.379* | .011 | 0.222 | .485 |
| P3 vs. C3 | 1.927*** | .000 | 1.282*** | .000 | -0.623* | .014 | -0.398* | .042 |
| P3 vs. F3 | 1.641*** | .000 | 2.226*** | .000 | -0.437 | .231 | -1.074** | .001 |
| C3 vs. F3 | -0.285 | .147 | 0.944*** | .000 | 0.187 | .453 | -0.676*** | .000 |
| Early visual | Pre-execution | Execution | Post-execution | |||||
|---|---|---|---|---|---|---|---|---|
| Mean Difference | p | Mean Difference | p | Mean Difference | p | Mean Difference | p | |
| O2 vs. P4 | 2.251** | .004 | -0.779* | .011 | -.504 | 111 | 1.155*** | .000 |
| O2 vs. C4 | 4.120*** | .000 | 0.121 | .752 | -1.469** | .003 | 1.129*** | .000 |
| O2 vs. F4 | 3.835** | .001 | 1.708** | .002 | -1.556* | .013 | 0.268 | .431 |
| P4 vs. C4 | 1.870*** | .000 | 0.900** | .002 | -.965** | .002 | -0.026 | .873 |
| P4 vs. F4 | 1.584** | .001 | 2.488*** | .000 | -1.053* | .038 | -0.887** | .004 |
| C4vs. F4 | -0.286 | 170 | 1.587*** | 000 | -.088 | .802 | -0.861*** | .000 |
| Early visual | Pre-execution | Execution | Post-execution | |
|---|---|---|---|---|
| Coh(F3, F4) | .102 | .163 | .175 | .105 |
| Coh(C3, C4) | .223 | .342 | .319 | .275 |
| Coh(P3, P4) | .229 | .319 | .303 | .272 |
| Coh(O1,O2) | .243 | .258 | .249 | .230 |
| F-value | F(1.818,27.274) =5.553; p=011 | F(1.816,27.243) =5.167; p=015 | F(2.133,31.991) =3.362; p=.044 | F(1.461,20.459) =5.689; p=017 |
| Early visual | Pre-execution | Execution | Post-execution | |||||
|---|---|---|---|---|---|---|---|---|
| Mean Difference | p | Mean Difference | p | Mean Difference | p | Mean Difference | p | |
| O1-O2 vs. P3-P4 | .014 | .784 | -.061 | 174 | -.053 | .304 | -.042 | .382 |
| O1-O2 vs. C3-C4 | .020 | .697 | -.084 | .193 | -.070 | .254 | -.046 | .434 |
| O1-O2 vs. F3-F4 | .141** | .008 | .095 | .188 | .075 | .270 | .125 | .101 |
| P3-P4 vs. C3-C4 | .005 | .836 | -.023 | .502 | -.016 | .606 | -.003 | .913 |
| P3-P4 vs. F3-F4 | .127*** | .000 | .156** | .002 | .129** | .004 | .167*** | .000 |
| C3-C4 vs. F3-F4 | .121** | .001 | .179** | .001 | .145* | .012 | .170*** | .000 |
| Coherence pairs | Early visual | Pre-execution | Execution | Post-execution | ||||
|---|---|---|---|---|---|---|---|---|
| r | p | r | p | r | p | r | p | |
| Coh(F3, F4) | -.115 | .672 | -.306 | .249 | -.047 | .863 | .021 | .940 |
| Coh(C3,C4) | -.418 | .107 | -.306 | .249 | -.585* | .017 | -.300 | .259 |
| Coh(P3, P4) | -.429 | .097 | -.376 | .151 | -.444 | .085 | -.300 | .259 |
| Coh(O1, O2) | -.088 | .745 | -.082 | .762 | -.432 | .094 | -.156 | .564 |
| Potentials & Coherences | Early visual | Pre-execution | Execution | Post-execution | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L | M | R | L | M | R | L | M | R | L | M | R | |
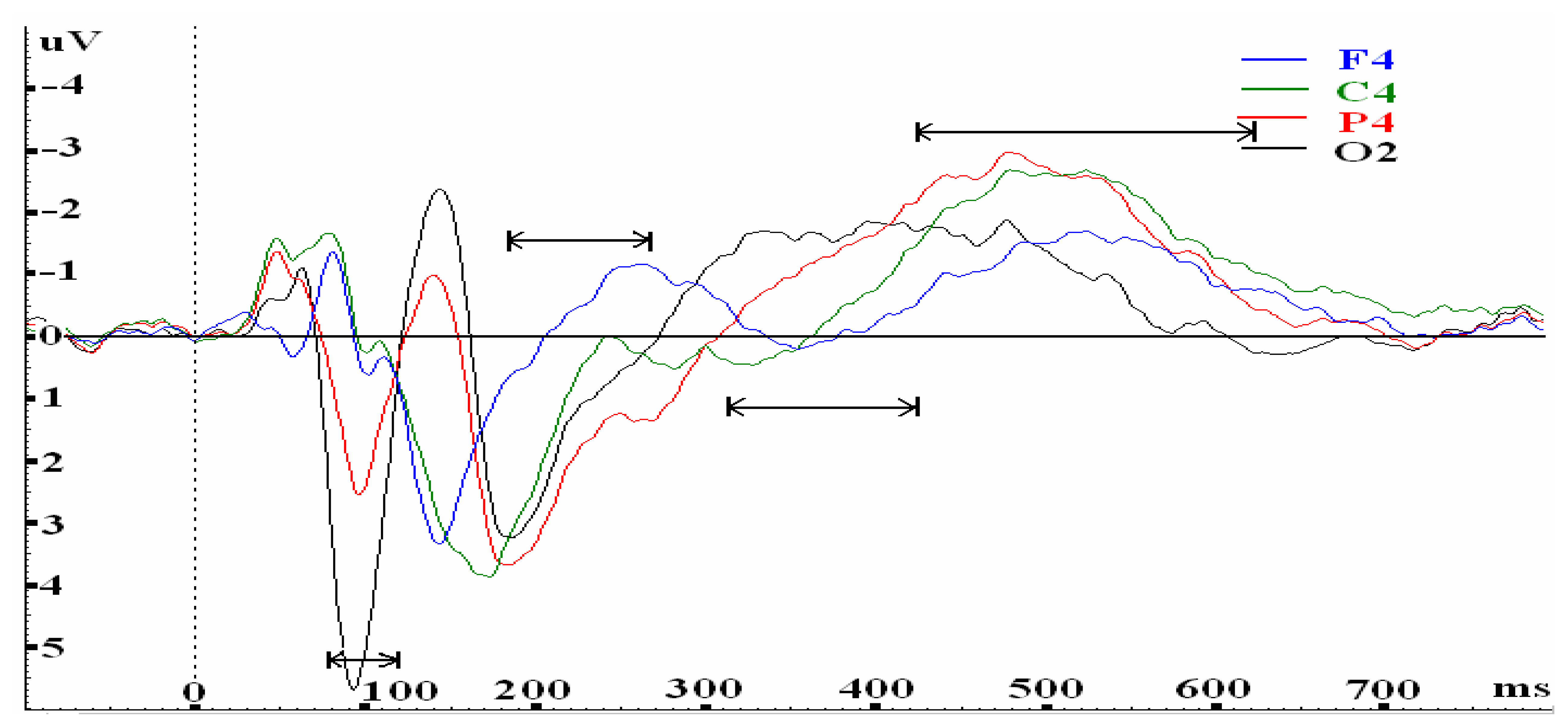
| P75-120 | O1 | Oz | O2 | |||||||||
| N175-260 | F3 | Fz | F4 | |||||||||
| P310-420 | C3 | Fz, Cz | F4, C4 | |||||||||
| N420-620 | P3 | Pz | P4,C4 | |||||||||
| To compare the coherence pairs | Mean Difference | p | To compare the coherence pairs | Mean Difference | p |
|---|---|---|---|---|---|
| O1-O2 vs. P3-P4 | .048 | .243 | P3-P4 vs. C3-C4 | .018 | .554 |
| O1-O2 vs. C3-C4 | .066* | .039 | P3-P4 vs. F3-F4 | .092** | .008 |
| O1-O2 vs. F3-F4 | .141* | .010 | C3-C4 vs. F3-F4 | .074* | .043 |
© 2007 by MDPI ( http://www.mdpi.org). Reproduction is permitted for noncommercial purposes.
Share and Cite
Meng, L.-F.; Lu, C.-P.; Chan, H.-L. Time-varying Brain Potentials and Interhemispheric Coherences of Anterior and Posterior Regions during Repetitive Unimanual Finger Movements. Sensors 2007, 7, 960-978. https://doi.org/10.3390/s7060960
Meng L-F, Lu C-P, Chan H-L. Time-varying Brain Potentials and Interhemispheric Coherences of Anterior and Posterior Regions during Repetitive Unimanual Finger Movements. Sensors. 2007; 7(6):960-978. https://doi.org/10.3390/s7060960
Chicago/Turabian StyleMeng, Ling-Fu, Chiu-Ping Lu, and Hsiao-Lung Chan. 2007. "Time-varying Brain Potentials and Interhemispheric Coherences of Anterior and Posterior Regions during Repetitive Unimanual Finger Movements" Sensors 7, no. 6: 960-978. https://doi.org/10.3390/s7060960