1. Introduction
Idiopathic pulmonary fibrosis is an irreversible and fatal interstitial lung disease with a high mortality rate, increasing annual incidence, and extremely poor prognosis for patients, with a survival time of 3–5 years after diagnosis [
1]. The current coronavirus disease 2019 (COVID-19) pandemic has been gradually controlled, but the consequent emergence of complications, such as pulmonary fibrosis, has caused widespread concern. According to the World Health Organization, more than 562 million people have been infected with COVID-19 worldwide and more than 6.3 million people have died [
2]. Most mild and moderate cases of COVID-19 can recover, but some severely ill patients still have persistent hypoxemia, and chest imaging shows the characteristic pattern of fibrosis [
3]. Complications of pulmonary fibrosis can occur in some patients, even after discharge from the hospital [
4,
5,
6,
7]. Therefore, development of efficient therapeutic drugs and treatment modalities to reduce pulmonary fibrosis is the key to effectively and completely curing patients with COVID-19. The process of pulmonary fibrosis formation can be considered an uncontrolled wound-healing response, which is ultimately manifested as massive proliferation and aggregation of myofibroblasts and excessive deposition of extracellular matrix in the alveoli and interstitium of the lung, leading to a reduction in lung volume, thickening of alveoli septa, and a loss of lung tissue elasticity [
8]. However, the pathogenesis of fibrosis is not completely clear, and the only antifibrotic drugs currently approved for marketing are pirfenidone and nintedanib, which have narrow indications, limited duration of administration, and some side effects. These drugs are not widely used in clinical practice [
9]. Therefore, there is an urgent need for efficient therapeutic drugs and treatment methods to improve the current status of pulmonary fibrosis treatment.
The interstitium is composed of a fibrous network and matrix, which is located between cells, substantive functional tissues, and organs, forming a multi-level network structure from a loose fibrous network to a dense fibrous stroma structure, making the body a flat and connected organic whole [
10,
11]. The structure and composition of the interstitium in different organs vary but are similar to some extent, consisting of flowing interstitial fluid and a complex network of collagen bundles [
12]. In addition to nutritional and mechanical support functions, the interstitial system has efficient connectivity pathways, which can serve as transportation channels for cancer cells, immune cells, and microorganisms, and is involved in pathophysiological processes, such as cancer cell metastasis, fibrosis, and edema [
12,
13]. These channels may be a potential conduit for damaging substances and pro-fibrotic signaling molecules, whereas interstitial cells may be precursors of myofibroblasts and play an important role in the formation of fibrosis. Therefore, the interstitium may play a crucial role in the occurrence and development of pulmonary fibrosis. However, the importance of the interstitium is neglected in the diagnosis and treatment of the disease. The combination of interstitial pathways with disease prevention and treatment will bring new technical means and application methods to scientific research and clinical practice. Intervaginal space injection (ISI) can deliver nanoparticles to various organs without relying on blood circulation, confirming that the interstitial system can serve as a long-range and efficient transportation pathway, providing a potentially efficient option for targeted drug delivery [
14,
15,
16,
17]. Therefore, the introduction of ISI as a novel drug delivery method in the treatment of pulmonary fibrosis may play a crucial role in the behavior of interstitial myofibroblasts and in improving the interstitial microenvironment.
Hydroxysafflor yellow A (HSYA), a compound with a monochalcone glycoside structure, is the most effective water-soluble ingredient of safflower that has been widely used in the treatment of cardiovascular diseases because of its effective anticoagulant, anti-myocardial ischemic, and vasorelaxant effects [
18,
19,
20,
21]. Studies have shown that HSYA inhibits lipopolysaccharide-induced proliferation, migration, and invasion of non-small-cell lung cancer cells by inhibiting the PI3K and AKT signaling pathways [
22] and TGF-β1-induced activation of human lung fibroblasts in vitro [
23]. HSYA has been shown to have anti-inflammatory, antioxidant, and apoptosis-inhibiting effects [
24], and it has potential applications in the treatment of dementia, Parkinson’s disease, and tumors.
Herein, hydroxysafflor yellow A phytosome (HYAP), as an anti-pulmonary fibrosis therapeutic agent, was prepared to improve the lipid solubility and bioavailability of HSYA, to treat bleomycin-induced pulmonary fibrosis in mice by ISI. This therapeutic strategy alleviates the severity of pulmonary fibrosis, elaborates the mechanism of interstitial treatment, and provides a novel therapeutic approach for the treatment of pulmonary fibrosis.
3. Discussion
ISI is a novel drug delivery method for injecting drugs or nanoparticles into inter-stitial pathways using the ankle or carpal tunnel as ISI sites, which contain tendons, arteries, veins, and nerves. This transport pathway allows gold nanoparticles (AuNPs) to reach the central nervous system without crossing the blood–brain barrier, providing a new avenue for more effective treatment of central nervous system diseases [
15]. Subsequently, the injection of malaria parasites through ISI better mimics the cutaneous phase of the natural malaria infection process, providing a new perspective on the mechanism of recurrence and enabling antimalarial treatment through this approach [
16]. A disordered imbalance of macrophages and fibroblasts is the major regulator of pulmonary fibrosis. However, the origin of fibrotic macrophages and fibroblasts is not clear. Interstitial resident fibroblasts and macrophages are most likely fibrogenic precursor cells recruited from interstitial channels to the site of injury following stimulation of the lung interstitium. We introduced ISI into the treatment of pulmonary interstitial fibrosis to suppress cell recruitment at the injury site through interstitial therapy, which in turn suppresses inflammation and accelerates recovery.
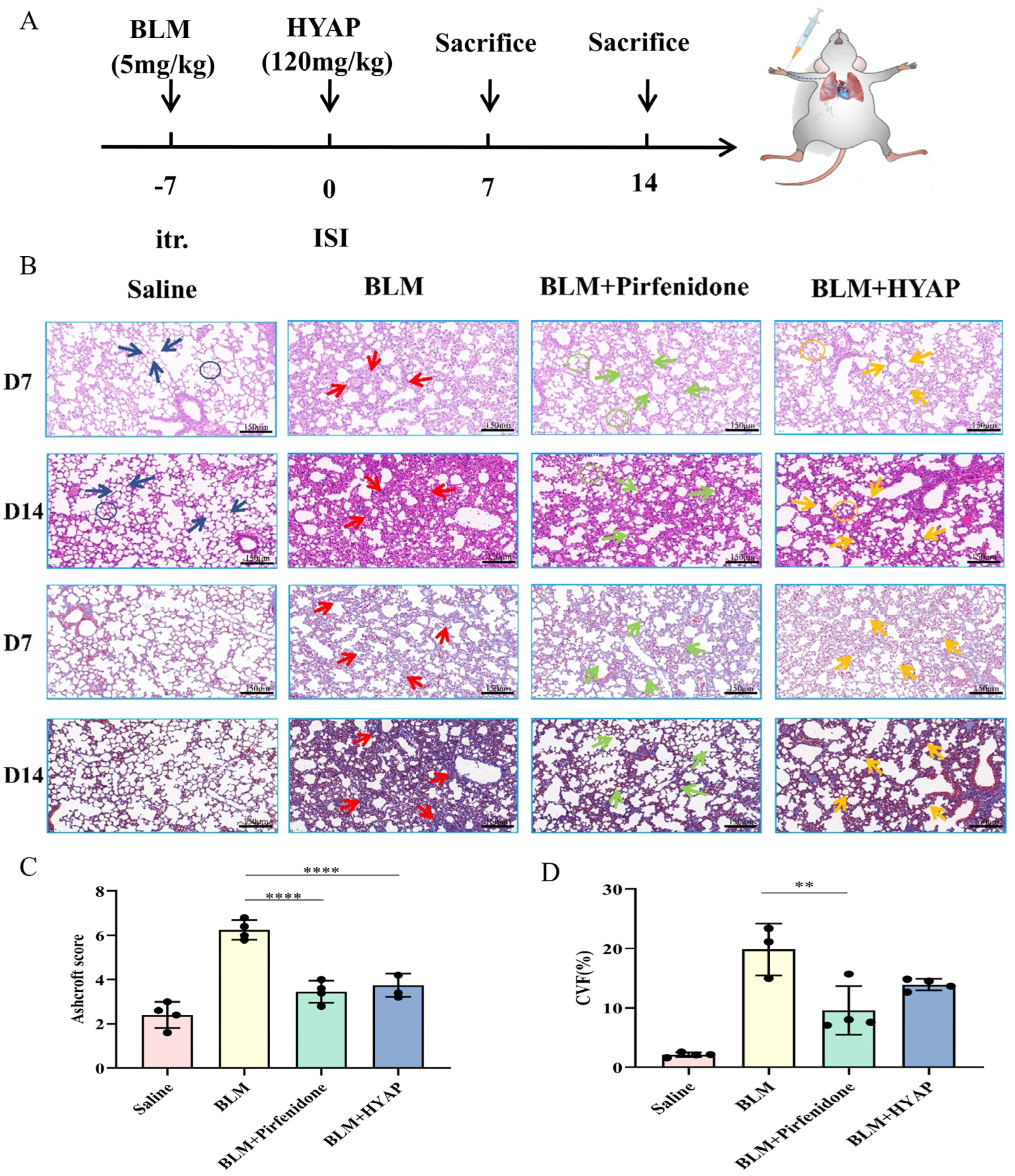
HSYA has antioxidant, anti-inflammatory, and anti-cancer activities and a wide range of pharmacological effects on the cardiovascular, hematological, and nervous systems. However, it exhibits poor lipid solubility and low bioavailability when orally administered. We selected HSYA as the target drug for interstitial treatment of antifibrosis and prepared phytosomes to improve the lipid solubility of HSYA, thereby improving its bioavailability. The prepared HYAP had a uniform particle size, good stability, and a good, sustained effect. Subsequently, we compared the different administration concentrations. When the administration concentration of HYAP was 60 mg/kg, the degree of pulmonary fibrosis did not increase after 14 days of administration, and the alveolar structure tended to recover; however, the pharmacological effect was not obvious. The recovery of the alveolar structure increased significantly, and collagen accumulation decreased significantly, with increasing administration concentration. Combined with the clinical dosage, we selected 120 mg/kg for the treatment of the mouse model of pulmonary fibrosis.
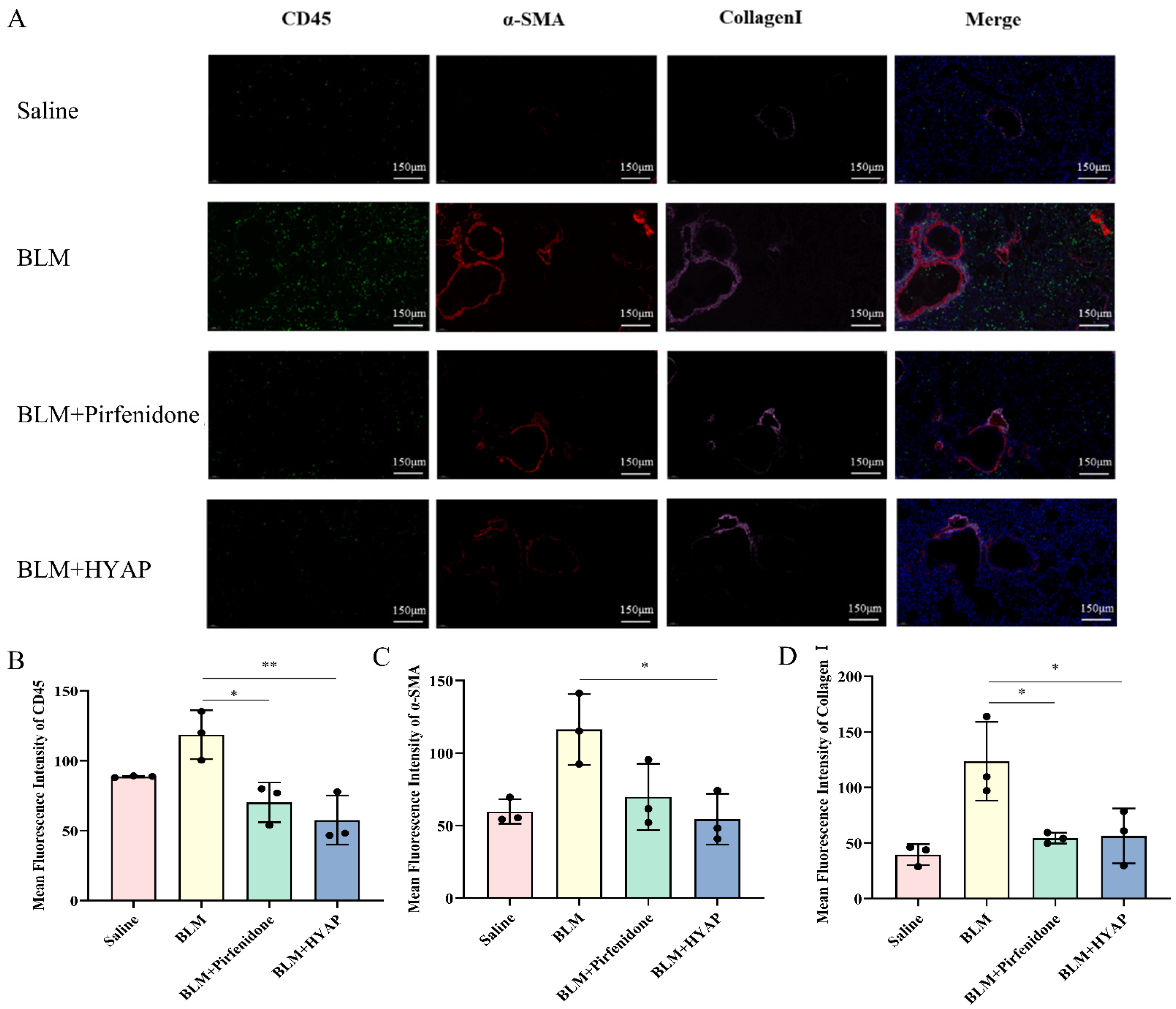
Repair of lung tissue after injury is a complex phenomenon involving intricate mechanisms. In general, wound healing undergoes three dynamic and interrelated stages that overlap over time: inflammatory, proliferative, and remodeling phases of maturation and scar formation [
25]. After repetitive injury, alveolar epithelial cells undergo damage, apoptosis, and capillary damage, leading to clot formation [
26]. The initial source of cytokine in the wound is platelets present in the clot, which provide a variety of factors that stimulate the recruitment of neutrophils and macrophages [
25]. Among these cytokines, TGF-β1 is a potent inducer of myofibroblast differentiation, which can also affect the balance between matrix metalloproteinases (MMPs) and their inhibitors (TIMPs), thereby favoring matrix deposition [
27]. In the present study, the plasma levels of TGF-β1 increased significantly in the BLM group, whereas those of TGF-β1 were restored to normal levels in the BLM + HYAP group, thereby reducing the stimulation of fibroblasts and synthesis of extracellular matrix. The proliferative phase includes angiogenesis, which is essential for tissue repair because it provides vascular perfusion to the wound and delivers oxygen and nutrients, thereby promoting cell proliferation. On the other hand, fibroblasts in the injured tissue are activated and differentiate into myofibroblasts. Although α-SMA is also expressed in smooth-muscle cells and pericytes, it remains the most reliable phenotypic marker for myofibroblasts [
28]. In the present study, using immunofluorescence, we confirmed the strong upregulation of α-SMA expression in the BLM group, whereas the expression levels of α-SMA in lung tissue decreased significantly in the BLM + HYAP group, suggesting that the proliferation and differentiation of fibroblasts were significantly inhibited by treatment with HYAP via ISI.
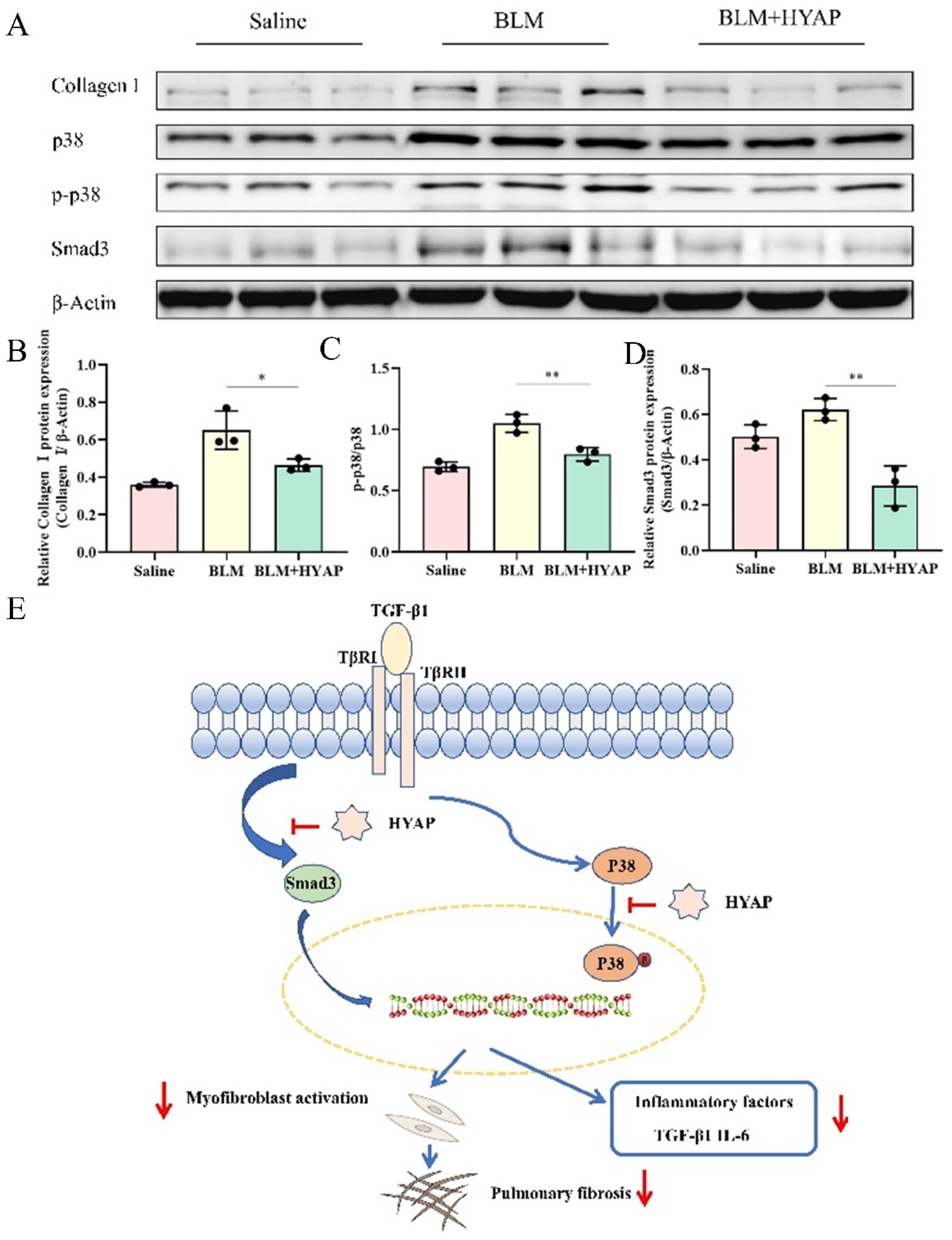
The role of the interstitium in pulmonary fibrosis has rarely been described. It has been shown that the subcutaneous interstitium is a major reservoir of fibroblasts, endothelial cells, and macrophages. In deep injury, fibroblasts in the subcutaneous interstitium direct their local complex matrix into the wound, coordinating with the coagulation cascade response and forming a prefabricated matrix that subsequently develops into mature scar tissue [
29]. The interstitial microenvironment plays an important role in disease development. By adding wound fluid to the culture environment, the function of fibroblasts in the normal dermis may be altered to resemble fibroblasts in injured tissue [
30]. Similarly, in the process of pulmonary fibrosis, fibroblasts and macrophages stored in the interstitium and interstitial fluid may play crucial roles in the occurrence and development of fibrosis. Therefore, it is important for us to try to identify an effective treatment for pulmonary fibrosis from an interstitial perspective. We attempted to identify an effective way to treat pulmonary fibrosis from an interstitial perspective. Using ISI in the upper limbs of mice, HYAP reached the corresponding organs through a non-vascular pathway, while inhibiting the proliferation and differentiation of fibroblasts in the interstitium by suppressing TGF-β/Smad signaling and further reducing collagen synthesis and secretion. HYAP entered the interstitial microenvironment through ISI and blocked the MAPK-p38 signaling pathway, possibly by inhibiting the activation of p38, and consequently, reducing the number of inflammatory factors in the interstitial fluid, thereby affecting fibroblast function and attenuating the progression of pulmonary fibrosis. However, whether ISI can play a more effective role in the treatment of pulmonary fibrosis needs to be further explored, but as far as the results are concerned, ISI has achieved good therapeutic results relative to oral positive drugs, proving the effectiveness of this treatment method.
4. Materials and Methods
4.1. Reagents
HSYA (C27H32O16; MW, 612.53; 98% Purity; Solarbio, Beijing, China), lecithin (Aladdin, Shanghai, China), bleomycin (BLM Biofeng, Shanghai, China), pirfenidone (Aladdin, Shanghai, China), ELISA kits (for TGF-β1 and IL-6; Cloud-Clone, Wuhan, China), 4% paraformaldehyde (Solarbio, Beijing, China), hematoxylin and eosin staining kit (beyotime, Shanghai, China), Weigert hematoxylin staining solution (Solarbio, Beijing, China), Ponceau S and Magenta staining solution (Solarbio, Beijing, China), phosphomolybdic acid solution (Solarbio, Beijing, China), Aniline blue solution (Solarbio, Beijing, China), DAPI (Solarbio, Beijing, China), RIPA buffer (Thermo Fisher Scientific, Waltham, USA), antibodies (for p38, p-p38, Smad3, collagen I, β-actin (Servicebio, Wuhan, China), anti-mouse IgG (Abcam, Cambridge, UK), anti-rabbit IgG (Abcam, Cambridge, UK)), BCA Protein Assay Reagent Kit (beyotime, Shanghai, China), and primary antibodies (CD45, α-SMA, and collagen I (Abcam, Cambridge, UK)), FITC-conjugated goat anti-rabbit IgG (Abcam, Cambridge, UK), CY3-conjugated goat anti-rat IgG (Abcam, Cambridge, UK), and CY5-conjugated goat anti-mouse IgG (Abcam, Cambridge, UK), were purchased.
4.2. Experimental Animals and Pulmonary Fibrosis Model Establishment
C57 BL/6J mice (male, 20 ± 2 g, 6 weeks old) were purchased from Beijing Vital River Laboratory Animal Technology (Beijing, China). C57 BL/6J mice were randomly divided into four groups: saline, BLM, BLM + pirfenidone, and BLM + HYAP groups. The experimental animals were anesthetized by inhalation of isoflurane, and the gas flow rate of the anesthesia machine was adjusted to 300 mL/min. After the anesthetic agent filled the anesthesia box, the animals were put into the anesthesia box with an anesthetic concentration of 3% and anesthesia time of 3 min. After the mice were completely anesthetized, the anesthesia mask was switched to continue the anesthesia on the experimental animals, followed by the next operation. The skin of the neck of the mice was dissected to expose the trachea, and 0.05 mL of BLM was injected at a dose of 5 mg/kg. An equal volume of saline was administered to the saline-treated group. This study was approved by the Institutional Animal Care and Use Committee (approval number: NCNST21-2104-0605).
4.3. Preparation of HYAP
The thin-film hydration method was used to prepare HYAP. HSYA powder and soybean lecithin were placed in a round-bottomed flask at a mass ratio of 1:3, ethanol was used as the solvent, and the mass concentration of HSYA was adjusted to 2 mg/kg. The samples were placed on a constant temperature magnetic stirrer and incubated at 40 °C for 2 h. After the reaction was complete, the unreacted precipitate was removed by suction filtration, and the organic solvent was removed by rotary evaporation under reduced pressure at 45 °C. An appropriate amount of chloroform and cholesterol (the mass ratio of phospholipid and cholesterol was 5:1) was added, shaken slightly to dissolve it, and any insoluble precipitates were removed via filtration. Continued rotary evaporation under reduced pressure at 45 °C formed a phytosome film on the bottom of the bottle. An appropriate amount of the saline was added to hydrate the phospholipid film for 30 min at 60 °C, followed by sonication for 15 min in a bath sonicator at 42 kHz and 100 W. The resulting phytosomes were subsequently extruded 10 times through a 100 nm polycarbonate porous membrane using an extruder (Avestin, Ottawa, ON, Canada).
4.4. Characterization of HYAP
4.4.1. Size, Zeta-Potential, and Morphological Characterization of HYAP
The size, polydispersity index, and zeta-potential of HYAP were measured by DLS (Malvern, London, UK) in triplicate at 25 °C. The morphology was characterized by TEM (Hitachi, Tokyo, Japan). A drop of diluted HYAP was placed on a carbon-coated grid and air-dried, and a drop of 1% uranyl acetate was deposited onto the grid for 3 min. Excess uranyl acetate stain was removed from the edge with filter paper, dried overnight at room temperature, and used for TEM.
4.4.2. Measurement of HYAP Compounding Rate, Encapsulation Rate, and Release Rate In Vitro
Phospholipids and HYAP are soluble in chloroform, so the compounding rate of HSYA with phospholipids can be measured by dissolving HYAP in chloroform. The finished compounded solution was dissolved by rotary evaporation out of anhydrous ethanol, and the appropriate amount of chloroform was added. The free HSYA precipitated because it was insoluble in chloroform, the precipitate was removed, and the filtrate was broken by rotary evaporation with the addition of quantitative methanol ultrasonication. The content of the compound HSYA was measured at 403 nm using UV (Perkin Elmer, Boston, MA, USA).
A certain amount of HYAP squeezed by the extruder was placed in an ultrafiltration tube with a MWCO of 10,000 Da, centrifuged at 15,000 r/min for 30 min, and the nanoparticles were trapped in the inner tube, then the nanoparticles were collected and added to the quantitative methanol ultrasonication to break the emulsion.
The compounding rate and encapsulation rate were calculated using the following equations:
The sustained release rate of the drug was determined by dialysis sampling and analyzed by UV. Using PBS as the release medium, stirring at 37 °C, 3 mL of HYAP was precisely measured in a pretreated dialysis bag. Samples (3 mL) were taken from the release medium at 0.5, 1, 2, 4, 6, 8, 10, 12, 24, 36, and 48 h and supplemented with equal volumes of PBS. The samples were filtered through a 0.22 μm microporous membrane and used for UV analysis. Absorbance values were measured at 403 nm and cumulative release rates were calculated.
4.4.3. Ultraviolet Spectroscopy Analysis
The HSYA, phospholipids, physical mixture of HSYA and phospholipids, and HYAP were dissolved in methanol as the solvent and methanol was used as the blank, then the absorbance was measured in the wavelength range of 200–600 nm and the curves were plotted.
4.4.4. Fourier Transform Infrared Spectroscopy Analysis
The mixture of HSYA and phospholipids, phospholipids, and HYAP with KBr was ground at 1:100, pressed into tablets, and then measured by FT-IR at wavelengths of 4000–500 cm−1.
4.5. Drug Treatment
Seven days after BLM injection, pulmonary fibrosis model mice were randomly divided into three groups of six mice each. The BLM + pirfenidone group was intragastrically treated with pirfenidone solution (200 mg/kg), daily for 14 days. The BLM + HYAP group was treated with HYAP in 0.1 mL of saline solution (120 mg/kg) by ISI in the right upper limb for 14 consecutive days, once a day. The chosen ISI injection point was the carpal tunnel of the wrist bone. The carpal tunnel is composed of the flexor retinaculum and carpal groove, inside which are the superficial and deep flexor tendons, flexor tendon sheath, flexor hallucis longus tendon and its sheath, and the median nerve and vessels running through it. The injection point was a convergent point for tendons, vessels, and nerve fibers connected to the fascia surrounding them. The saline and BLM groups were injected with an equal amount of saline via ISI in the right upper limb for 14 consecutive days, once a day.
4.6. CT Imaging Analysis
On day 14 of administration, mice were randomly selected from the saline, BLM, BLM + pirfenidone, and BLM + HYAP groups, and changes in lung structure during the establishment of fibrosis and drug treatment were recorded using CT in vivo imaging. CT was preheated for 15 min. The setting parameters were as follows: voltage, 90 kV; imaging current, 180 µA; real-time observation current, 180 µA; FOV, 24 mm; scanning program, 4.5 min; scanning angle, 360. The anesthesia machine was connected to an oxygen bottle, isoflurane was added, the gas source valve was adjusted at the front end of the oxygen flowmeter so that the output gas reached the required flow rate (300 mL/min), and the mice were placed in the anesthesia box. The anesthetized mice were fixed on the instrument cavity operating table, and the chamber was closed for scanning and analysis of the structure.
4.7. Histopathological Analysis
The CO2 asphyxiation method was adopted to execute the animal, whereby the animal was placed into the euthanasia box and CO2 was infused into the box at a rate of 20% of the volume of the box per minute replacement. It was confirmed that the animal was not moving and not breathing, the CO2 was turned off after the pupils were dilated, and then the animal was observed for 2 min to ensure that it was dead. Then, the left lower lung tissues were collected from each group on days 7 and 14 after administration and then fixed in 4% paraformaldehyde for 72 h. The samples were subsequently dehydrated using a series of gradients of ethanol, made transparent with xylene, and then embedded in paraffin. The samples were then cut into thin slices using an automated microtome, mounted on slides, and stored at room temperature for hematoxylin and eosin (HE) and Masson trichrome staining. After dewaxing, hydration, staining, dehydration, and sealing, pathological analysis was performed. Semi-quantitative analysis of pulmonary fibrosis was performed with reference to the Ashcroft scoring criteria, and the collagen volume fraction of Masson trichrome staining was calculated for different groups using Image J software.
4.8. Immunofluorescence
The largest lobe of the left lung was fixed with 4% paraformaldehyde, sliced using a frozen microtome, and stained with immunofluorescence. The sections were incubated overnight at 4 °C after adding the primary antibodies against CD45, α-SMA, and collagen I (1:50), and the slices were washed three times with TBST and incubated with FITC-labeled secondary antibody (1:200), CY3-labeled secondary antibody (1:300), and CY5-labeled secondary antibody (1:400) at room temperature for 3 h. The nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI) for 3 min, washed three times with TBST, dehydrated with gradient alcohol, made transparent with xylene, and sealed. Images were detected and collected by a slice scanner: DAPI glows blue by UV excitation wavelength 330–380 nm and emission wavelength 420 nm, FITC glows green by excitation wavelength 465–495 nm and emission wavelength 515–555 nm, CY3 glows red by excitation wavelength 510–560 nm and emission wavelength 590 nm, and CY5 glows pink by excitation wavelength 608–648 nm and emission wavelength 672–712 nm (CY5 was originally red, however to distinguish it from CY3, we set it to pink light). The staining results were analyzed using the Image J software.
4.9. ELISA
On day 14 of treatment, blood was collected from the ocular venous sinus and placed in blood collection tubes with EDTA. The upper plasma was collected after centrifugation at 4 °C and 3000 rpm for 5 min. ELISA kits were used to quantify the levels of TGF-β1 and IL-6 in the plasma according to the manufacturer’s instructions.
4.10. Western Blotting
A section (100 mg) of the lower lobe of the right lung was cut into small pieces on ice. After adding 1 mL of RIPA lysis solution and 10 μL of PMSF (100 mM), the sample was ground until there was no visible tissue and was left standing for 30 min or until complete lysis. The cells were centrifuged at 10,000× g at 4 °C for 10 min, and the supernatant was kept at −80 °C. Protein concentration was determined using the BCA method. Equal amounts of total protein were separated via sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to a nitrocellulose membrane using the semi-dry transfer method. After being sealed with 5% skim milk powder for 1 h at room temperature, the membrane was incubated with primary antibody diluted in phosphate-buffered saline overnight at 4 °C. After washing thrice with TBST, the proteins were incubated with horseradish peroxide-coupled IgG diluted in PBS at room temperature for 1 h. The proteins collagen I, p38, p-p38, and Smad3 were detected using enhanced chemiluminescence reagents, and the optical density values of the target bands were determined.
4.11. Statistical Analysis
Group comparisons were performed using the GraphPad Prism (version 9.0) software (GraphPad Software, San Diego, CA, USA). Ordinary one-way ANOVA was used for comparisons between more than two groups. Statistical significance was set at p < 0.05. * p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




