
2-Styrylchromones: Cytotoxicity and Modulation of Human Neutrophils’ Oxidative Burst
by
 , , , , and
, , , , and
Mariana Lucas
1 ,
,
Marisa Freitas
1,
Marco Zanchetta
2 ,
,
Artur M. S. Silva
2,
Eduarda Fernandes
1,* and
Daniela Ribeiro
1,3,* 1
LAQV, REQUIMTE, Laboratory of Applied Chemistry, Department of Chemical Sciences, Faculty of Pharmacy, University of Porto, 4050-313 Porto, Portugal
2
LAQV, REQUIMTE & Department of Chemistry, Campus de Santiago, University of Aveiro, 3810-193 Aveiro, Portugal
3
Faculty of Agrarian Sciences and Environment, University of the Azores, 9700-042 Angra do Heroísmo, Portugal
*
Authors to whom correspondence should be addressed.
Pharmaceuticals 2022, 15(3), 288; https://doi.org/10.3390/ph15030288
Submission received: 28 January 2022
/
Revised: 21 February 2022
/
Accepted: 21 February 2022
/
Published: 25 February 2022
(This article belongs to the Special Issue Selected Papers from the 7th International Electronic Conference on Medicinal Chemistry (ECMC2021))
Abstract
:Neutrophils are polymorphonuclear leukocytes recruited to sites of acute inflammation, in response to pathogen invasion and tissue injury. The modulation of their activity, especially oxidative burst, may be important to control the inflammatory process. 2-Styrylchromones (2-SC) are derived from chromones and despite their recognized multiple biological activities, their anti-inflammatory and antioxidant properties are still poorly explored. Therefore, in this study, 43 structurally related 2-SC were evaluated concerning their effects on freshly isolated human neutrophils’ viability and oxidative burst. The studied 2-SC were divided into eight groups according to their substitution at C-4′ on B-ring (none, -OH, -OCH3, -OBn, -CH3, and -NO2), existence and location of -Cl on B-ring, and presence of -Br at C-3 on C-ring. Overall, most of the studied 2-SC did not affect neutrophils’ viability, at physiological relevant concentrations. The ones belonging to B group were the most effective (IC50 values < 2 μM), and present one -OH group at C-4′ or a catechol group at C-3′ and C-4′ on B-ring. These substituents seem to play an important role in the modulatory activity of human neutrophils’ oxidative burst. These results reinforce the great potential of 2-SC’s scaffold for the development of new anti-inflammatory agents.
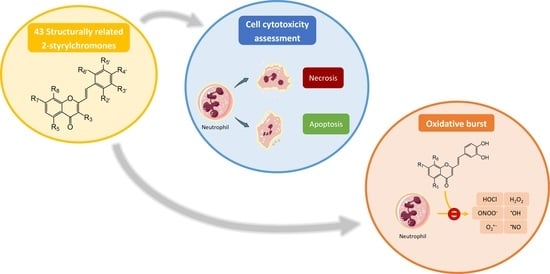
1. Introduction
Inflammation is involved in the development and worsening of several diseases, such as diabetes, rheumatoid arthritis, and cancer [1]. Inflammation is a natural biological process by which the body triggers a non-specific protective response due to harmful stimuli, namely pathogen invasion and tissue injury [1,2]. At the onset of inflammation, cellular signaling occurs, prompting the release of a series of inflammatory mediators. These mediators will increase the permeability of endothelial cells enabling immune cells, such as neutrophils, to access the site of infection or injury [1,2]. Neutrophils belong to the polymorphonuclear leukocytes group, together with basophils and eosinophils, and are highly specialized white blood cells characterized by their rapid migration to the inflammatory local and by their important role in the defense of the organism [3]. At the inflammatory site, neutrophils are activated and trigger a cascade of mechanisms to eliminate the invading agents. One of the main mechanisms by which neutrophils eliminate the invading agents is through the activation of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase complex, with the consequent production of reactive species [4,5]. The reactive species produced during oxidative burst (Figure 1) are essential for the effective antimicrobial and anti-inflammatory activity of neutrophils [3,4]. However, the overproduction of these reactive species, characteristic of the chronic inflammatory diseases, can compromise the endogenous antioxidant defenses, leading to an imbalance between the amount of reactive species produced and their removal, resulting in a phenomenon known as oxidative stress. Consequently, this condition can cause harm to the host, particularly near inflamed tissues, namely through the oxidative damage of biomolecules, such as DNA, lipids, and proteins [2,6].
Neutrophils are immune cells with a short life span, which homeostasis is essentially maintained by constitutive apoptosis upon aging. This type of cell death is a programmed process characterized by several specific cellular changes manifested in the nucleus, cytoplasm, mitochondria, and plasma membrane (Figure 2) [11,12]. Another of the main death mechanisms is necrosis, which is characterized by certain hallmarks as cell swelling, plasma membrane rupture, and release of intracellular content (Figure 2) [13]. During the inflammatory process, there is a delay in apoptosis, which significantly increases the longevity of neutrophils, ensuring their continued presence at the site of inflammation [5,14]. Although these cells are fundamental in the inflammatory process, their activity and survivability must be controlled, to avoid an extended exacerbation of inflammation. Therefore, the development of strategies capable of modulating neutrophils’ activity, namely oxidative burst, has become a central point in the search for anti-inflammatory therapeutic agents [14].
Styrylchromones are a small group of chromone-derived compounds. Featuring an oxygenated heterocyclic structure, the existence of a styryl moiety attached to the chromone core is one of their characteristic features, and the position of this group determines the nomenclature of these compounds [17]. They are scarcely found in nature and are mostly of synthetic origin. 2-Styrylchromones (2-SC) are the most common type of styrylchromones in nature and their synthesis has been commonly described [17]. Over the years, 2-SC have been reported to display several biological effects, namely antiallergic [18], antiviral [19,20,21], antibacterial, antifungal [22,23,24,25], neuroprotective [26,27,28,29], antioxidant [24,30,31], anti-inflammatory [32,33], and antitumoral [34,35,36,37]. Although the chromone core is present in other type of compounds shown to affect human neutrophils’ viability and oxidative burst [38,39], to the best of our knowledge, such effects have not been demonstrated for 2-SC. The present study intends to fill this gap by evaluating the effect of 43 structurally related 2-SC (Figure 3) on neutrophils’ life span and oxidative burst of freshly isolated human neutrophils, establishing, whenever possible, a structure-activity relationship (SAR).
2. Results
2.1. Effect of 2-SC on Neutrophils’ Viability
The effects of each 2-SC were evaluated in terms of total neutrophils’ population percentage, and proportion of viable cells [annexin V (−)/PI (−)], apoptotic cells [annexin V (+)/PI (−)], late apoptotic cells [annexin V (+)/PI (+)], and necrotic cells [annexin V (−)/PI (+)].
At relatively high concentrations, some of the studied 2-SC were cytotoxic. Among the 2-SC of the A group, A2 (12.50 and 25.00 μM), A4 (50.00 μM), A5 (12.50 and 25.00 μM), and A7 (12.50 and 25.00 μM) significantly affected the total neutrophils’ population percentage and additionally, A4 and A5 induced late apoptosis. As an example, Figure 4 displays representative plots of 2-SC A3 and A4, where A3 (75.00 μM) showed no effect on human neutrophils’ viability, and A4 (50.00 μM) affected the total neutrophils’ population percentage and induced late apoptosis. Thus, for 2-SC A4, it was necessary to decrease the tested concentration to find the one that did not affect neutrophils’ viability (25.00 μM).
However, 2-SC in the B group were the most cytotoxic. 2-SC B1 (50.00 μM), B2 (12.50 μM), B4 (12.50 μM), B5 (12.50 and 25.00 μM), and B7 (6.25 μM) affected the total neutrophils’ population percentage, and additionally B7 (0.75–6.25 μM) induced necrosis. 2-SC B10 (12.50 μM) was shown to induce late apoptosis. 2-SC F2 (25.00 μM) induced apoptosis, while H1 (25.00 μM) affected the total neutrophils’ population percentage and induced late apoptosis. The other studied 2-SC were not cytotoxic at the maximum tested concentration.
Figure 5 displays representative examples of the proportion of viable cells, apoptotic cells, late apoptotic cells, and necrotic cells for one 2-SC from each group. 2-SC F2 (25.00 μM) induced apoptosis when compared to the control, while A4 (50.00 μM) and H1 (25.00 μM) induced late apoptosis. The 2-SC B7 (0.75–6.25 μM) of B group significantly induced necrosis, in a concentration-dependent manner, up to the concentration of 0.38 μM, for which no effect on the viability was observed. 2-SC C8 (6.25 μM), D1 (12.50 μM), E2 (12.50 μM), and G1 (12.50 μM) did not affect cells’ viability.
The finding of cytotoxic effects allowed us to accurately select the non-toxic concentrations. Table 1 shows the 2-SC concentrations that did not affect human neutrophils’ viability, i.e., the values of viable cells, apoptotic cells, late apoptotic cells, and necrotic cells that were not statistically different from the control (untreated cells). These concentrations were chosen as the maximal concentrations to be tested in the neutrophils’ oxidative burst assay.
2.2. Effect of 2-SC on Neutrophils’ Oxidative Burst
From all the tested 2-SC, 17 of them were effective inhibitors of neutrophils’ oxidative burst, particularly those belonging to B group, as shown in Table 2. 2-SC B6, B8–B13, and C8 were the most active compounds, presenting IC50 values between 0.7 ± 0.1 (B8) and 1.4 ± 0.2 μM (C8) (Figure 6). For 2-SC from A group, it was only possible to define the percentage of inhibition of 2-SC A3, which reached an inhibitory activity of 53 ± 4% for the concentration of 75.00 μM. The remaining 2-SC in this group were not active, up to the highest tested concentrations (6.25–100.00 μM). Most of the 2-SC from B group inhibited neutrophils’ oxidative burst (IC50 values between 0.7 ± 0.1 and 7.5 ± 0.4 μM), except for B5 and B7 that did not show an inhibitory effect, but the maximal concentrations able to be tested were 6.25 and 0.38 μM, respectively. 2-SC B2 only reached 50 ± 3% of inhibition for the concentration of 6.25 μM. Among 2-SC in C group, 2-SC C8 was the most active, with an IC50 value of 1.4 ± 0.2 μM. 2-SC C4 and C6 also demonstrated activity (IC50 = 24.2 ± 0.3 and 20 ± 2 μM, respectively), whereas C3 and C5 only reached 54 ± 4% and 54 ± 2% of inhibition, respectively, for the concentration of 50.00 μM. 2-SC from D–H groups did not show an effect on the neutrophils’ oxidative burst, up to the maximum tested concentrations (6.25–50.00 μM).
3. Discussion
During the inflammatory response, there is an increase in the production of reactive species by neutrophils, and consequently, an increase in oxygen consumption, leading to an overproduction of reactive oxygen species (ROS) and reactive nitrogen species (RNS)—oxidative burst [7,40]. The modulation of the oxidative burst can occur by scavenging of reactive species and/or inhibition of the enzymes directly involved in the production of reactive species, namely NADPH oxidase complex, superoxide dismutase, and MPO [9].
2-SC have demonstrated various biological activities, including antioxidant and anti-inflammatory [17]. However, to the best of our knowledge, there are no studies in the literature about the effects of 2-SC on human neutrophils’ viability and oxidative burst.
The studied panel of structurally related 2-SC was divided into eight groups considering the presence and the type of substituents (none, -OH, -OCH3, -OBn, -CH3, and -NO2) at C-4′ on B-ring (A to F groups), presence and location of -Cl on B-ring (G group), and presence of -Br at C-3 on C-ring (H group), which allowed to establish a SAR. As mentioned, this study is divided into two parts: the evaluation of the effects of 2-SC on neutrophils’ viability and modulation of the oxidative burst.
In the first part of this work, annexin V/PI flow cytometric assay was applied to assess human neutrophils’ viability. In what concerns neutrophils’ viability, the majority of the studied 2-SC did not affect the viability of these cells, up to the maximum concentration tested for each compound. However, some 2-SC were cytotoxic, affecting the total neutrophils’ population percentage (A2, A4, A5, A7, B1, B2, B4, B5, B7, and H1) and inducing apoptosis (F2), late apoptosis (A4, A5, B10, and H1), or directly necrosis (B7). Currently, in the literature, some of the 2-SC studied in this work are described as being cytotoxic at lower concentrations; however, to the best of our knowledge, there are only studies demonstrating this cytotoxicity in tumor cell lines [34,35,41,42]. In the present work, the concentrations at which the 2-SC showed cytotoxicitywill not have an impact at a physiological level, since they are relatively high (>6.25 μM). Although there are no studies regarding the metabolism and bioavailability of 2-SC, they can be compared with flavonoids, since they are structurally related compounds to 2-SC. It is described in the literature the concentration of flavonoids that can be found in human plasma rarely exceeds 1 μM [43].
In the next phase of this work, a chemiluminescence method, using luminol as a probe, was applied to evaluate the modulation of neutrophils’ oxidative burst. Among the studied 2-SC, the ones from B group were the most effective. Regarding 2-SC from A group, no significant effect in the modulation of the oxidative burst was found. The 2-SC of this group have no substituent at C-4′ on B-ring. Some of the 2-SC belonging to A group (A1–A4) were evaluated by Gomes and co-workers [30] for their scavenging reactive species potential, but were not shown very effective. Ribeiro and co-workers [38] used the same methodology of the present work to assess the modulatory effect of oxidative burst of some flavonoids structurally similar to these 2-SC (A1–A4, A6, and A7); however, there was no significant difference between the activity of flavonoids and 2-SC, since both showed low or no activity.
As mentioned, most of the 2-SC with the best activity belong to B group. These 2-SC have in common the presence of -OH group at C-4′ on B-ring. This substituent in this position in the 2-SC structure (B1) seems to contribute to the activity, when compared with the unsubstituted A1. Nevertheless, it should be noted that within this group, the presence of a catechol group at C-3′ and C-4′ on B-ring seems to be more favorable for the intended activity, as B6 was almost 7.5 times more active than B1. Gomes and co-workers [30,44] showed that some of the 2-SC from this group (B1, B3, B4, B6–B9, B12, and B13) were effective scavengers of reactive species, namely superoxide anion radical (O2•−), hydrogen peroxide (H2O2), hypochlorous acid (HOCl), singlet oxygen (1O2), nitric oxide radical (•NO), and peroxynitrite anion (ONOO−), while B2 was much less active or inactive. Interestingly, the 2-SC that were the most effective in scavenging reactive species (B6, B8, B9, B12, and B13) were also good modulators of the oxidative burst. This suggests that these 2-SC may modulate oxidative burst by scavenging reactive species. Ribeiro and co-workers [38] also evaluated some flavonoids structurally identical to some of the studied 2-SC (B1–B4 and B6–B9), differing only in the presence of the styryl moiety. Overall, the 2-SC showed a better effect than the correspondent flavonoid, which may indicate that the styryl moiety contributes to the observed enhanced effect.
The structures of the 2-SC of B group differ from those of A group mainly by the presence of -OH groups on B-ring. The presence of -OH groups on B-ring of the 2-SC has a greater influence on inhibitory activity than the presence of -OH on A-ring, as can be seen by comparing the activities found for B6 and A4. B6 was active while A4 showed no activity. This observation was also corroborated when -OCH3 groups are present on the A-ring, as can be seen by comparing B8 with B10, B9 with B11, and B12 with B13. These results seem to indicate that the effects of 2-SC are mainly influenced by the presence of -OH group on B-ring, namely the presence of the catechol at C-3′ and C-4′. The importance of the catechol group for the antioxidant activity seems to be related to the formation of a semiquinone, which suffers dismutation and forms a stable ortho-quinone. This process is possible since the -OH groups are highly reactive and able to donate an electron to the reactive species, which stabilizes them due to electron delocalization [45,46].
Among the 2-SC in C group, 2-SC C8 was the most active. C8 has a catechol group at C-7 and C-8 on A-ring, whereas on B-ring it has -OCH3 groups at C-3′ and C-4′. Previously, C8 also demonstrated to have a good scavenging activity of reactive species (O2•−, 1O2, •NO and ONOO−) [44]. C4 and C6 also showed some activity but were about 14 and 17 times less active, respectively, than C8. In fact, C4 and C6 have no -OH group in their structure, unlike C8, demonstrating the importance of -OH groups for this effect. This idea can also be corroborated by comparing the structure of C1 with B1, both with only one substituent present at C-4′ on B-ring: B1 has -OH and C1 has -OCH3, and, as expected, B1 was active while C1 was not active. The same behavior was observed between B6 and C2, where B6 presents a catechol at C-3′ and C-4′ on B-ring, and C2 presents -OCH3 groups at the same positions. When two -OCH3 groups are present on B-ring, namely at C-3′ and C-4′, the presence of one -OCH3 at C-7 on A-ring (C4) seems to better improve the activity when compared to one -OCH3 at C-5 on A-ring (C3). When only -OCH3 groups are present as substituents in 2-SC, the presence of two -OCH3 on A-ring only enhanced the activity if they are on C-7 and C-8, as can be seen by comparing C6 with C5. C6 has -OCH3 at C-7 and C-8 on A-ring and C-3′ and C-4′ on B-ring, while on C5 the -OCH3 groups are located at C-5 and C-7 on A-ring and C-3′ and C-4′ on B-ring. Still, in the presence of two -OCH3 groups on B-ring, varying the type of substituents present on A-ring, namely at C-7 and C-8, it can be concluded that -OH group (C8) is the one that most favors the modulation of the oxidative burst, when compared to the presence of -OCH3 (C6) and -OBn (C7). The presence of -OBn groups on B-ring also did not favor the activity as observed in the results obtained for 2-SC D1–D3. The same behavior was also observed for the 2-SC with -CH3 (E1 and E2) or -NO2 (F1 and F2) on C-4′ on B-ring. The presence of -Cl groups at different positions on B-ring also does not contribute to the activity of G1–G4, even in the presence of -OH groups on C-5 on A-ring (G1, G3, and G4). The simultaneous existence of -Br at C-3 on C-ring and -OCH3 at C-7 on A-ring (H1–H3) also did not seem to favor the modulation of oxidative burst.
Figure 7 shows the chemical structures and structural characteristics of the most active 2-SC in the modulation of human neutrophils’ oxidative burst.
4. Materials and Methods
4.1. Reagents
Histopaque 1077 and 1119, trypan blue solution 0.4%, calcium chloride (CaCl2), Trizma® base, D-glucose, quercetin, magnesium sulfate (MgSO4), Dulbecco’s phosphate buffer saline, without CaCl2 and magnesium chloride (PBS), phorbol-12-myristate-13-acetate (PMA), dimethylsulfoxide (DMSO), and luminol were obtained from Sigma Chemical Co. (St. Louis, MO, USA). Sodium chloride (NaCl) and potassium chloride (KCl) were obtained from VWR Chemicals (Alfragide, Portugal) and Pronalab (Abrunheira, Portugal), respectively. The commercial FITC Annexin V Apoptosis Detection Kit I was obtained from BD Biosciences (Franklin Lakes, NJ, USA). All compounds were synthesized as previously described [44,47,48].
4.2. Equipment
The determination of cells’ yield and viability, through the trypan blue exclusion method, was done on an optic microscope (Nikon Eclipse E200, Nikon Instruments Inc., Melville, NY, USA). The Accuri C6 flow cytometer (BD, Becton, Dickinson and Company, Franklin Lakes, NJ, USA) was used to collect the fluorescence signals for the assessment of 2-SC effects on neutrophils’ viability (apoptosis and necrosis). The chemiluminescent assay for the in the assessment of 2-SC effects on neutrophils’ oxidative burst was performed in a microplate reader (Synergy HT, BIO-TEK Instruments, Inc., Winooski, VT, USA).
4.3. Methods
4.3.1. General Information
All the studied 2-SC and the positive control, quercetin, were dissolved in DMSO (4% in the reactional mixture). Preliminary experiments were performed to verify the clear solubility of the 2-SC under study in the tested experimental conditions and their possible interference effects with the used methodologies. From these assays, the maximum tested concentration was chosen, to avoid any interference with the methodologies used. At least three individual independent experiments were performed in duplicate, for each assay, using between three and nine concentrations.
4.3.2. Human Neutrophils Isolation
Following the Declaration of Helsinki and the approval of patient-related procedures and protocols by the Ethics Committee of Centro Hospitalar do Porto, Portugal, venous blood was collected from healthy human blood donors, after informed consent. The blood was stored into K3EDTA vacuum tubes after antecubital venipuncture of the donors. In order to obtain the isolated neutrophils, the density gradient centrifugation method was applied, as previously reported [38], with modifications. In brief, in a 15 mL polypropylene tube, 3 mL of histopaque 1119 were added followed by 3 mL of histopaque 1077 and finally, 4.5 mL of the collected blood were slowly added. Each tube was centrifuged at 900× g at 20 °C for 30 min, with a medium acceleration and deceleration speed. Following centrifugation, the neutrophils’ portion was harvested, and PBS was added to double the volume. The neutrophils’ suspension was then centrifuged at 850× g at 4 °C for 5 min, using a maximum acceleration and deceleration speed. The supernatant was discarded, and the neutrophils’ pellet was resuspended in PBS and afterward milli-Q water was added to lyse the residual erythrocytes. The tube was gently homogenized and after a few minutes, 3% NaCl was added to restore isotony, and then centrifuged again at 850× g at 4 °C for 5 min, using a maximum acceleration and deceleration speed. The supernatant was discarded, and a neutrophils’ pellet was obtained. After isolation, the neutrophils’ pellet was resuspended in the incubation media, tris-glucose buffer (25 mM Trizma® base, 5.5 mM D-glucose, 140 mM NaCl, 1.26 mM CaCl2, 0.81 mM MgSO4, 5.37 mM KCl, pH = 7.4). Neutrophils’ suspension was kept on ice, under soft shaking, until use. The trypan blue exclusion method was used to determine cells’ yield and viability.
4.3.3. Assessment of the Effect of 2-SC on Neutrophils’ Viability
The effect of the studied 2-SC on neutrophils’ viability was assessed by flow cytometry, after simultaneous staining with FITC Annexin V and propidium iodide (PI) (Figure S1), as previously reported by our research group [49], with modifications. After isolation, neutrophils (1 × 106 cells/mL) were incubated with 2-SC (up to 100.00 μM) or DMSO for 40 min, at 37 °C, in a 48-well plate. At the end of incubation, the content of each well was transferred to conical microtubes and centrifuged at 400× g at 20 °C for 5 min. After discarding the supernatant, the neutrophils’ pellet was resuspended in PBS and centrifuged under the previous conditions. Once the supernatant was removed, the pellet was resuspended in 10× diluted binding buffer, and then PI and annexin V were added. This mixture was incubated in the dark at room temperature for 15 min, and finally, 10× diluted binding buffer was added. In a flow cytometer, the fluorescence signal, of at least 10,000 cells/sample, were collected in logarithmic mode and followed in channels 1 and 3. The green fluorescence corresponding to annexin V FITC was accompanied in channel 1 and plotted as a histogram of FL1 staining, whereas the fluorescence corresponding to PI was monitored in channel 3 and plotted in the same histogram but as FL3 staining. The collected data were analyzed with the BD Accuri™ C6 software. The data analysis was restricted to the neutrophils’ population. Thus, according to the light-scattering properties of neutrophils (in a forward versus side scatter plot), a polygon gate was drawn, where debris and other blood cells were excluded. The effect of 2-SC on neutrophils’ viability was expressed as a relative percentage of apoptotic and/or necrotic cells when compared to the control (without 2-SC).
4.3.4. Assessment of the Effect of 2-SC on Neutrophils’ Oxidative Burst
The effect of 2-SC on the modulation of neutrophils’ oxidative burst was assessed after the stimulation of these cells with PMA by monitoring the oxidation of chemiluminescent probe, luminol, by the generated reactive species (Figure S2), according to a previously described methodology [38]. Briefly, neutrophils (1 × 106 cells/mL) were pre-incubated with luminol (500.00 μM), and 2-SC (0.00–100.00 μM) or DMSO, at the final concentrations indicated. After this pre-incubation, at 37 °C for 5 min, PMA (160.00 nM) was added, and in turn, neutrophils’ stimulation was initiated. Kinetic readings were immediately initiated and followed for 40 min, at 37 °C, in a microplate reader. The arbitrary chemiluminescence value was taken at the peak of the curve, and the results were expressed as the inhibition percentage of luminol oxidation. The maximum tested concentrations of 2-SC in this assay were defined according to the results obtained in the viability assay, where the concentrations that did not affect neutrophils’ viability were defined.
4.3.5. Statistical Analysis
GraphPad Prism 6 software was used to plot the curves of percentage of inhibition versus concentrations of compound, from which the concentration that produces 50% of inhibition (IC50) was determined. GraphPad Prism 6 software was also used to perform all statistical analyses. Statistical comparison among the most active 2-SC was estimated by applying the one-way analysis (ANOVA), followed by the Bonferroni’s multiple comparisons test. In all cases, p-values < 0.05 were considered statistically significant. The results are expressed as mean ± standard error of the mean (SEM).
5. Conclusions
In conclusion, the effect of 2-SC on human neutrophils’ viability and oxidative burst was assessed for the first time in this study, providing promising results. Most of the studied 2-SC did not affect neutrophils’ viability, at physiologically meaningful concentrations. The 2-SC belonging to B group, with only one -OH group at C-4′ or a catechol group at C-3′ and C-4′ on B-ring, were the most effective, which seems to indicate that the presence and the position of these groups play an important role for the modulation of human neutrophil’s oxidative burst. Additionally, the 2-SC with -OCH3 groups at C-3′ and C-4′ on B-ring (C group) also showed modulatory effects, nonetheless not as effective, with exception of C8. One of the possible mechanisms of action of the most active 2-SC found may be through the scavenging of reactive species, since they have already demonstrated this ability in in vitro non-cellular studies.
The obtained results in this study demonstrate that 2-SC are safe to human neutrophils and simultaneously display a scaffold with great potential for the development of new drugs to act on inflammatory processes, especially when a catechol group is present at the B-ring.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/article/10.3390/ph15030288/s1, Figure S1: Annexin V/propidium iodide (PI) staining, Figure S2: Luminol staining.
Author Contributions
Conceptualization, E.F. and D.R.; formal analysis, M.L.; funding acquisition, E.F. and D.R.; investigation, M.L.; methodology, M.L.; project administration, E.F. and D.R.; supervision, A.M.S.S., E.F. and D.R.; validation, E.F. and D.R.; writing—original draft, M.L.; writing—review and editing, M.L., M.F., M.Z., A.M.S.S., E.F., and D.R. All authors have read and agreed to the published version of the manuscript.
Funding
This APC was financially supported by Operational Program Competitiveness and Internationalization supported by FEDER COMPETE POCI-01-0145-FEDER-029253 and National Funds FCT Fundação para a Ciência e Tecnologia PTDC/MED-QUI/29253/2017.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Ethics Committee of Centro Hospitalar Universitário do Porto/Instituto de Ciências Biomédicas Abel Salazar, Oporto, Portugal.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Data are contained within the article.
Acknowledgments
The authors gratefully acknowledge the medical and the nursing staff of the Centro Hospitalar do Porto-Hospital de Santo António Blood Bank for their collaboration in the recruitment of blood donors to participate in the study. Mariana Lucas thanks FCT and ESF (European Social Fund) through Programa Operacional Regional do Norte (NORTE 2020) for her Ph.D. Grant (2021.06746.BD). Marisa Freitas further acknowledges the contract under the Scientific Employment Stimulus-Individual Call (CEEC Individual) 2020.04126.CEECIND/CP1596/CT0006.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Netea, M.G.; Balkwill, F.; Chonchol, M.; Cominelli, F.; Donath, M.Y.; Giamarellos-Bourboulis, E.J.; Golenbock, D.; Gresnigt, M.S.; Heneka, M.T.; Hoffman, H.M.; et al. A guiding map for inflammation. Nat. Immunol. 2017, 18, 826–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Elbim, C.; Lizard, G. Flow cytometric investigation of neutrophil oxidative burst and apoptosis in physiological and pathological situations. Cytom. A 2009, 75A, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C.; Kettle, A.J.; Hampton, M.B. Reactive oxygen species and neutrophil function. Annu. Rev. Biochem. 2016, 85, 765–792. [Google Scholar] [CrossRef] [PubMed]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef]
- Chatterjee, S. Chapter Two—Oxidative Stress, Inflammation, and Disease. In Oxidative Stress and Biomaterials; Dziubla, T., Butterfield, D.A., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 35–58. [Google Scholar]
- Amulic, B.; Cazalet, C.; Hayes, G.L.; Metzler, K.D.; Zychlinsky, A. Neutrophil function: From mechanisms to disease. Annu. Rev. Immunol. 2012, 30, 459–489. [Google Scholar] [CrossRef]
- Herb, M.; Schramm, M. Functions of ROS in Macrophages and Antimicrobial Immunity. Antioxidants 2021, 10, 313. [Google Scholar] [CrossRef]
- Freitas, M.; Lima, J.L.F.C.; Fernandes, E. Optical probes for detection and quantification of neutrophils’ oxidative burst. A review. Anal. Chim. Acta 2009, 649, 8–23. [Google Scholar] [CrossRef]
- Fang, F.C. Antimicrobial reactive oxygen and nitrogen species: Concepts and controversies. Nat. Rev. Microbiol. 2004, 2, 820–832. [Google Scholar] [CrossRef]
- El Kebir, D.; Filep, J.G. Modulation of neutrophil apoptosis and the resolution of inflammation through β2 Iintegrins. Front. Immunol. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.R.; Loison, F. Constitutive neutrophil apoptosis: Mechanisms and regulation. Am. J. Hematol. 2008, 83, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Figueroa, E.; Álvarez-Carrasco, P.; Ortega, E.; Maldonado-Bernal, C. Neutrophils: Many ways to die. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Liew, P.X.; Kubes, P. The neutrophil’s role during health and disease. Physiol. Rev. 2019, 99, 1223–1248. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Kabakov, A.E.; Gabai, V.L. Cell death and survival assays. In Chaperones: Methods and Protocols; Methods in Molecular Biology; Calderwood, S.K., Prince, T.L., Eds.; Humana Press: New York, NY, USA, 2018; pp. 107–127. [Google Scholar]
- Santos, C.M.M.; Silva, A.M.S. An overview of 2-styrylchromones: Natural occurrence, synthesis, reactivity and biological properties. Eur. J. Org. Chem. 2017, 2017, 3115–3133. [Google Scholar] [CrossRef]
- Doria, G.; Romeo, C.; Forgione, A.; Sberze, P.; Tibolla, N.; Corno, M.L.; Cruzzola, G.; Cadelli, G. Antiallergic agents. III. Substituted trans-2-ethenyl-4-oxo-4H-1-benzopyran-6-carboxylic acids. Eur. J. Med. Chem. 1979, 14, 347–351. [Google Scholar] [CrossRef]
- Desideri, N.; Conti, C.; Mastromarino, P.; Mastropaolo, F. Synthesis and anti-rhinovirus activity of 2-styrylchromones. Antivir. Chem. Chemother. 2000, 11, 373–381. [Google Scholar] [CrossRef]
- Desideri, N.; Mastromarino, P.; Conti, C. Synthesis and evaluation of antirhinovirus activity of 3-hydroxy and 3-methoxy 2-styrylchromones. Antivir. Chem. Chemother. 2003, 14, 195–203. [Google Scholar] [CrossRef]
- Conti, C.; Mastromarino, P.; Goldoni, P.; Portalone, G.; Desideri, N. Synthesis and anti-rhinovirus properties of fluoro-substituted flavonoids. Antivir. Chem. Chemother. 2005, 16, 267–276. [Google Scholar] [CrossRef]
- Ujwala, B.; Priyadarsini, P.; Rao, V.M. Synthesis and bio-activity evaluation of 2-styrylchromones. Int. J. Pharma Bio Sci. 2013, 4, 199–206. [Google Scholar]
- Nikam, M.D.; Mahajan, P.S.; Damale, M.G.; Sangshetti, J.N.; Chate, A.V.; Dabhade, S.K.; Gill, C.H. Novel amalgamation of 2-styrylchromones and 1,2,4-triazole: Synthesis, antimicrobial evaluation and docking study. Lett. Drug. Des. Discov. 2015, 12, 650–660. [Google Scholar] [CrossRef]
- Rao, V.M.; Ujwala, B.; Priyadarsini, P.; Krishna Murthy, P. Synthesis, antioxidant and antimicrobial activity of three new 2-styrylchromones and their analogues. Der Pharma Chem. 2016, 8, 1–6. [Google Scholar]
- Momin, M.; Ramjugernath, D.; Chenia, H.; Koorbanally, N.A. Synthesis and evaluation of novel fluorinated 2-styrylchromones as antibacterial agents. J. Chem. 2013, 2013, 436758. [Google Scholar] [CrossRef] [Green Version]
- Takao, K.; Endo, S.; Nagai, J.; Kamauchi, H.; Takemura, Y.; Uesawa, Y.; Sugita, Y. 2-Styrylchromone derivatives as potent and selective monoamine oxidase B inhibitors. Bioorganic Chem. 2019, 92, 103285. [Google Scholar] [CrossRef]
- Yoon, J.S.; Lee, M.K.; Sung, S.H.; Kim, Y.C. Neuroprotective 2-(2-phenylethyl)chromones of Imperata cylindrica. J. Nat. Prod. 2006, 69, 290–291. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Qiao, L.; Xie, D.; Yuan, Y.; Chen, N.; Dai, J.; Guo, S. 2-(2-Phenylethyl)chromones from Chinese eaglewood. Phytochemistry 2012, 76, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.J.; Jung, H.A.; Min, B.S.; Choi, J.S. Anticholinesterase and β-site amyloid precursor protein cleaving enzyme 1 inhibitory compounds from the heartwood of Juniperus chinensis. Chem. Pharm. Bull. 2015, 63, 955–960. [Google Scholar] [CrossRef] [Green Version]
- Gomes, A.; Fernandes, E.; Silva, A.M.S.; Santos, C.M.M.; Pinto, D.C.G.A.; Cavaleiro, J.A.S.; Lima, J.L.F.C. 2-Styrylchromones: Novel strong scavengers of reactive oxygen and nitrogen species. Bioorganic Med. Chem. 2007, 15, 6027–6036. [Google Scholar] [CrossRef]
- Takamatsu, S.; Hodges, T.W.; Rajbhandari, I.; Gerwick, W.H.; Hamann, M.T.; Nagle, D.G. Marine natural products as novel antioxidant prototypes. J. Nat. Prod. 2003, 66, 605–608. [Google Scholar] [CrossRef] [Green Version]
- Gomes, A.; Capela, J.P.; Ribeiro, D.; Freitas, M.; Silva, A.M.S.; Pinto, D.C.G.A.; Santos, C.M.M.; Cavaleiro, J.A.S.; Lima, J.L.F.C.; Fernandes, E. Inhibition of NF-kB activation and cytokines production in THP-1 monocytes by 2-styrylchromones. Med. Chem. 2015, 11, 560–566. [Google Scholar] [CrossRef]
- Takamatsu, S.; Nagle, D.G.; Gerwick, W.H. Secondary metabolites from marine Cyanobacteria and Algae inhibit LFA-1/ICAM-1 mediated cell adhesion. Planta Med. 2004, 70, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Lu, P.-J.; Yang, C.-N.; Hulme, C.; Shaw, A.Y. Structure-activity relationship study of growth inhibitory 2-styrylchromones against carcinoma cells. Med. Chem. Res. 2013, 22, 2385–2394. [Google Scholar] [CrossRef]
- Momoi, K.; Sugita, Y.; Ishihara, M.; Satoh, K.; Kikuchi, H.; Hashimoto, K.; Yokoe, I.; Nishikawa, H.; Seiichiro, F.; Sakagami, H. Cytotoxic activity of styrylchromones against human tumor cell lines. In Vivo 2005, 19, 157–163. [Google Scholar]
- Shaw, A.Y.; Chang, C.-Y.; Liau, H.-H.; Lu, P.-J.; Chen, H.-L.; Yang, C.-N.; Li, H.-Y. Synthesis of 2-styrylchromones as a novel class of antiproliferative agents targeting carcinoma cells. Eur. J. Med. Chem. 2009, 44, 2552–2562. [Google Scholar] [CrossRef]
- Lee, K.Y.; Nam, D.H.; Moon, C.S.; Seo, S.H.; Lee, J.Y.; Lee, Y.S. Synthesis and anticancer activity of lavendustin A derivatives containing arylethenylchromone substituents. Eur. J. Med. Chem. 2006, 41, 991–996. [Google Scholar] [CrossRef]
- Ribeiro, D.; Freitas, M.; Tomé, S.M.; Silva, A.M.S.; Porto, G.; Fernandes, E. Modulation of human neutrophils’ oxidative burst by flavonoids. Eur. J. Med. Chem. 2013, 67, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Freitas, M.; Ribeiro, D.; Tomé, S.M.; Silva, A.M.S.; Fernandes, E. Synthesis of chlorinated flavonoids with anti-inflammatory and pro-apoptotic activities in human neutrophils. Eur. J. Med. Chem. 2014, 86, 153–164. [Google Scholar] [CrossRef]
- El-Benna, J.; Hurtado-Nedelec, M.; Marzaioli, V.; Marie, J.-C.; Gougerot-Pocidalo, M.-A.; Dang, P.M.-C. Priming of the neutrophil respiratory burst: Role in host defense and inflammation. Immunol. Rev. 2016, 273, 180–193. [Google Scholar] [CrossRef]
- Yang, C.-H.; Yang, Y.; Liu, J.-H. Platachromone A–D: Cytotoxic 2-styrylchromones from the bark of Platanus×acerifolia (Aiton) Willd. Phytochem. Lett. 2013, 6, 387–391. [Google Scholar] [CrossRef]
- Uesawa, Y.; Nagai, J.; Shi, H.; Sakagami, H.; Bandow, K.; Tomomura, A.; Tomomura, M.; Endo, S.; Takao, K.; Sugita, Y. Quantitative structure–cytotoxicity relationship of 2-styrylchromones. Anticancer Res. 2019, 39, 6489. [Google Scholar] [CrossRef] [Green Version]
- Scalbert, A.; Williamson, G. Dietary intake and bioavailability of polyphenols. J. Nutr. 2000, 130, 2073S–2085S. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.; Neuwirth, O.; Freitas, M.; Couto, D.; Ribeiro, D.; Figueiredo, A.G.P.R.; Silva, A.M.S.; Seixas, R.S.G.R.; Pinto, D.C.G.A.; Tomé, A.C.; et al. Synthesis and antioxidant properties of new chromone derivatives. Bioorg. Med. Chem. 2009, 17, 7218–7226. [Google Scholar] [CrossRef] [PubMed]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef]
- Gomes, A.; Fernandes, E.; Garcia, M.B.Q.; Silva, A.M.S.; Pinto, D.C.G.A.; Santos, C.M.M.; Cavaleiro, J.A.S.; Lima, J.L.F.C. Cyclic voltammetric analysis of 2-styrylchromones: Relationship with the antioxidant activity. Bioorg. Med. Chem. 2008, 16, 7939–7943. [Google Scholar] [CrossRef]
- Santos, C.M.M.; Silva, A.M.S.; Cavaleiro, J.A.S. Synthesis of new hydroxy-2-styrylchromones. Eur. J. Org. Chem. 2003, 2003, 4575–4585. [Google Scholar] [CrossRef]
- Santos, C.M.M.; Silva, A.M.S.; Cavaleiro, J.A.S. Efficient syntheses of new polyhydroxylated 2,3-diaryl-9H-xanthen-9-ones. Eur. J. Org. Chem. 2009, 2009, 2642–2660. [Google Scholar] [CrossRef]
- Freitas, M.; Costa, V.M.; Ribeiro, D.; Couto, D.; Porto, G.; Carvalho, F.; Fernandes, E. Acetaminophen prevents oxidative burst and delays apoptosis in human neutrophils. Toxicol. Lett. 2013, 219, 170–177. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Mechanisms involved in production of reactive species in neutrophils’ oxidative burst. The nicotinamide adenine dinucleotide phosphate (NADPH) oxidase complex is activated, its cytoplasmic components migrate to the cellular membrane, and the molecular oxygen is reduced to the superoxide anion radical (O2•−), which in turn is the precursor to hydrogen peroxide (H2O2) and other reactive species [3,4,7,8]. Most of the H2O2 formed is consumed by myeloperoxidase (MPO), which catalyzes the formation of hypochlorous acid (HOCl), in the presence of H2O2 and by oxidation of the halide Cl−, since this ion exists in high concentrations in the body [4,7,8,9]. In the cytoplasm, L-arginine can be converted into L-citrulline and nitric oxide radical (•NO) by the action of the inducible nitric oxide synthase (iNOS). Consequently, •NO can react with O2•− and form peroxynitrite anion (ONOO−) [8,9,10]. •OH, hydroxyl radical; SOD, superoxide dismutase.
Figure 1.
Mechanisms involved in production of reactive species in neutrophils’ oxidative burst. The nicotinamide adenine dinucleotide phosphate (NADPH) oxidase complex is activated, its cytoplasmic components migrate to the cellular membrane, and the molecular oxygen is reduced to the superoxide anion radical (O2•−), which in turn is the precursor to hydrogen peroxide (H2O2) and other reactive species [3,4,7,8]. Most of the H2O2 formed is consumed by myeloperoxidase (MPO), which catalyzes the formation of hypochlorous acid (HOCl), in the presence of H2O2 and by oxidation of the halide Cl−, since this ion exists in high concentrations in the body [4,7,8,9]. In the cytoplasm, L-arginine can be converted into L-citrulline and nitric oxide radical (•NO) by the action of the inducible nitric oxide synthase (iNOS). Consequently, •NO can react with O2•− and form peroxynitrite anion (ONOO−) [8,9,10]. •OH, hydroxyl radical; SOD, superoxide dismutase.

Figure 2.
Mechanisms involved in cell death (apoptotic and necrotic pathways). During necrosis, the cell swells, which consequently leads to the loss of membrane integrity and the release of intracellular contents, and subsequently the disintegration of the cell. The apoptotic process is characterized by condensation of chromatin and cytoplasm, fragmentation of the cell nucleus, and formation of apoptotic bodies. In addition, changes in cell structure occur [13,15,16].
Figure 2.
Mechanisms involved in cell death (apoptotic and necrotic pathways). During necrosis, the cell swells, which consequently leads to the loss of membrane integrity and the release of intracellular contents, and subsequently the disintegration of the cell. The apoptotic process is characterized by condensation of chromatin and cytoplasm, fragmentation of the cell nucleus, and formation of apoptotic bodies. In addition, changes in cell structure occur [13,15,16].

Figure 3.
Chemical structures of the studied 2-SC and of the positive control, quercetin. 2-SC were divided into eight groups according to the type of substituents at C-4′ on B-ring (A to F groups), existence and location of -Cl on B-ring (G group), and presence of -Br at C-3 on C-ring (H group).
Figure 3.
Chemical structures of the studied 2-SC and of the positive control, quercetin. 2-SC were divided into eight groups according to the type of substituents at C-4′ on B-ring (A to F groups), existence and location of -Cl on B-ring (G group), and presence of -Br at C-3 on C-ring (H group).

Figure 4.
Effect of 2-SC A3 and A4 on human neutrophils’ viability. Representative flow cytometry plots of forward scatter area (FSC-A, xx axis) / side scatter area (SSC-A, yy axis) histogram (A,C,E,G,I), and annexin V (xx axis) / propidium iodide (PI, yy axis) histogram (B,D,F,H,J). (A,B,E,F): control (without 2-SC); (C,D): 2-SC A3 (75.00 μM); and (G–J): 2-SC A4 (25.00 and 50.00 μM).
Figure 4.
Effect of 2-SC A3 and A4 on human neutrophils’ viability. Representative flow cytometry plots of forward scatter area (FSC-A, xx axis) / side scatter area (SSC-A, yy axis) histogram (A,C,E,G,I), and annexin V (xx axis) / propidium iodide (PI, yy axis) histogram (B,D,F,H,J). (A,B,E,F): control (without 2-SC); (C,D): 2-SC A3 (75.00 μM); and (G–J): 2-SC A4 (25.00 and 50.00 μM).

Figure 5.
Representative examples of the effect of 2-SC on neutrophils’ viability, for different concentrations: A4 (25.00 and 50.00 μM), B7 (0.38 and 6.25 μM), C8 (6.25 μM), D1 (12.50 μM), E2 (12.50 μM), F2 (12.50 and 25.00 μM), G1 (12.50 μM), and H1 (12.50 and 25.00 μM). Results are expressed as the percentages (%) of viable cells [annexin V (−)/PI (−)], apoptotic cells [annexin V (+)/PI (−)], late apoptotic cells [annexin V (+)/PI (+)], and necrotic cells [annexin V (−)/PI (+)]; and presented as mean ± SEM (n ≥ 3).
Figure 5.
Representative examples of the effect of 2-SC on neutrophils’ viability, for different concentrations: A4 (25.00 and 50.00 μM), B7 (0.38 and 6.25 μM), C8 (6.25 μM), D1 (12.50 μM), E2 (12.50 μM), F2 (12.50 and 25.00 μM), G1 (12.50 μM), and H1 (12.50 and 25.00 μM). Results are expressed as the percentages (%) of viable cells [annexin V (−)/PI (−)], apoptotic cells [annexin V (+)/PI (−)], late apoptotic cells [annexin V (+)/PI (+)], and necrotic cells [annexin V (−)/PI (+)]; and presented as mean ± SEM (n ≥ 3).

Figure 6.
Graphical representation of IC50 values of active 2-SC and the positive control, quercetin (μM, mean ± SEM, n ≥ 3). Same letters indicate that IC50 values are not statistically different and different letters indicate that IC50 values are statistically different from each other (p < 0.05).
Figure 6.
Graphical representation of IC50 values of active 2-SC and the positive control, quercetin (μM, mean ± SEM, n ≥ 3). Same letters indicate that IC50 values are not statistically different and different letters indicate that IC50 values are statistically different from each other (p < 0.05).

Figure 7.
Chemical structures of the most active 2-SC in the modulation of neutrophils’ oxidative burst.
Figure 7.
Chemical structures of the most active 2-SC in the modulation of neutrophils’ oxidative burst.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Higher 2-SC concentrations (μM) that did not affect human neutrophils’ viability, according to the annexin V/PI assay.
Table 1.
Higher 2-SC concentrations (μM) that did not affect human neutrophils’ viability, according to the annexin V/PI assay.
| 2-SC | Concentration (μM) | 2-SC | Concentration (μM) |
|---|---|---|---|
| A1 | 75.00 | C2 | 25.00 |
| A2 | 6.25 | C3 | 50.00 |
| A3 | 75.00 | C4 | 37.50 |
| A4 | 25.00 | C5 | 75.00 |
| A5 | 6.25 | C6 | 37.50 |
| A6 | 100.00 | C7 | 25.00 |
| A7 | 6.25 | C8 | 6.25 |
| A8 | 25.00 | D1 | 12.50 |
| B1 | 25.00 | D2 | 12.50 |
| B2 | 6.25 | D3 | 12.50 |
| B3 | 25.00 | E1 | 25.00 |
| B4 | 6.25 | E2 | 12.50 |
| B5 | 6.25 | F1 | 6.25 |
| B6 | 12.50 | F2 | 12.50 |
| B7 | 0.38 | G1 | 12.50 |
| B8 | 25.00 | G2 | 12.50 |
| B9 | 12.50 | G3 | 25.00 |
| B10 | 6.25 | G4 | 25.00 |
| B11 | 12.50 | H1 | 12.50 |
| B12 | 12.50 | H2 | 50.00 |
| B13 | 12.50 | H3 | 12.50 |
| C1 | 25.00 |
Table 2.
Chemical structures and inhibition of human neutrophils’ oxidative burst by the tested 2-SC and the positive control, quercetin.
Table 2.
Chemical structures and inhibition of human neutrophils’ oxidative burst by the tested 2-SC and the positive control, quercetin.
| 2-SC | R5 | R7 | R8 | R2′ | R3′ | R4′ | R6′ | Inhibitory Activity * (% ± SEM) or IC50 (μM, mean ± SEM) | |
|---|---|---|---|---|---|---|---|---|---|
| A1 |  | - | - | - | - | - | - | - | <30% 75.00µM |
| A2 | OH | - | - | - | - | - | - | <30% 6.25µM | |
| A3 | - | OH | - | - | - | - | - | 53 ± 4 % 75.00µM | |
| A4 | OH | OH | - | - | - | - | - | <30% 25.00µM | |
| A5 | OCH3 | - | - | - | - | - | - | <30% 6.25µM | |
| A6 | - | - | - | - | OH | - | - | <30% 100.00µM | |
| A7 | OH | - | - | - | OH | - | - | <30% 6.25µM | |
| A8 | OH | - | - | OH | - | - | - | <30% 25.00µM | |
| B1 |  | - | - | - | - | - | - | - | 7.5 ± 0.4 |
| B2 | OH | - | - | - | - | - | - | 50 ± 3 % 6.25µM | |
| B3 | - | OH | - | - | - | - | - | 4.1 ± 0.3 | |
| B4 | OH | OH | - | - | - | - | - | 3.1 ± 0.3 | |
| B5 | OH | OCH3 | - | - | - | - | - | <30% 6.25µM | |
| B6 | - | - | - | - | OH | - | - | 1.0 ± 0.1 | |
| B7 | OH | - | - | - | OH | - | - | <30% 0.38µM | |
| B8 | - | OH | - | - | OH | - | - | 0.7 ± 0.1 | |
| B9 | OH | OH | - | - | OH | - | - | 0.8 ± 0.1 | |
| B10 | - | OCH3 | - | - | OH | - | - | 1.0 ± 0.1 | |
| B11 | OCH3 | OCH3 | - | - | OH | - | - | 0.8 ± 0.1 | |
| B12 | - | OH | OH | - | OH | - | - | 0.9 ± 0.3 | |
| B13 | - | OCH3 | OCH3 | - | OH | - | - | 1.0 ± 0.2 | |
| C1 |  | - | - | - | - | - | - | - | <30% 25.00µM |
| C2 | - | - | - | - | OCH3 | - | - | <30% 25.00µM | |
| C3 | OCH3 | - | - | - | OCH3 | - | - | 54 ± 4 % 50.00µM | |
| C4 | - | OCH3 | - | - | OCH3 | - | - | 24.2 ± 0.3 | |
| C5 | OCH3 | OCH3 | - | - | OCH3 | - | - | 54 ± 2 % 50.00µM | |
| C6 | - | OCH3 | OCH3 | - | OCH3 | - | - | 20 ± 2 | |
| C7 | - | OBn | OBn | - | OCH3 | - | - | <30% 25.00µM | |
| C8 | - | OH | OH | - | OCH3 | - | - | 1.4 ± 0.2 | |
| D1 |  | - | - | - | - | OBn | - | - | <30% 12.50µM |
| D2 | OCH3 | OCH3 | - | - | OBn | - | - | <30% 12.50µM | |
| D3 | - | OCH3 | OCH3 | - | OBn | - | - | <30% 12.50µM | |
| E1 |  | - | - | - | - | - | - | - | <30% 6.25µM |
| E2 | OH | - | - | - | - | - | - | <30% 12.50µM | |
| F1 |  | - | - | - | - | - | - | - | <30% 6.25µM |
| F2 | OH | - | - | - | - | - | - | <30% 12.50µM | |
| G1 |  | OH | - | - | - | Cl | - | - | <30% 12.50µM |
| G2 | - | - | - | - | - | Cl | - | <30% 12.50µM | |
| G3 | OH | - | - | - | Cl | Cl | - | <30% 25.00µM | |
| G4 | OH | - | - | Cl | - | - | Cl | <30% 25.00µM | |
| H1 |  | - | - | - | - | - | - | - | <30% 12.50µM |
| H2 | OCH3 | - | - | - | - | - | - | <30% 50.00µM | |
| H3 | - | - | - | - | OCH3 | OCH3 | - | <30% 12.50µM | |
 | Positive control Quercetin | 0.8 ± 0.1 | |||||||
* The percentage of inhibition is expressed for the highest concentration (in superscript) that could be tested under the assay conditions to avoid interferences with the methodology.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lucas, M.; Freitas, M.; Zanchetta, M.; Silva, A.M.S.; Fernandes, E.; Ribeiro, D. 2-Styrylchromones: Cytotoxicity and Modulation of Human Neutrophils’ Oxidative Burst. Pharmaceuticals 2022, 15, 288. https://doi.org/10.3390/ph15030288
AMA Style
Lucas M, Freitas M, Zanchetta M, Silva AMS, Fernandes E, Ribeiro D. 2-Styrylchromones: Cytotoxicity and Modulation of Human Neutrophils’ Oxidative Burst. Pharmaceuticals. 2022; 15(3):288. https://doi.org/10.3390/ph15030288
Chicago/Turabian StyleLucas, Mariana, Marisa Freitas, Marco Zanchetta, Artur M. S. Silva, Eduarda Fernandes, and Daniela Ribeiro. 2022. "2-Styrylchromones: Cytotoxicity and Modulation of Human Neutrophils’ Oxidative Burst" Pharmaceuticals 15, no. 3: 288. https://doi.org/10.3390/ph15030288
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.