The Potential Role of Cannabinoids in Modulating Serotonergic Signaling by Their Influence on Tryptophan Metabolism


{kind=link}
{kind=link}
Abstract
:1. Introduction


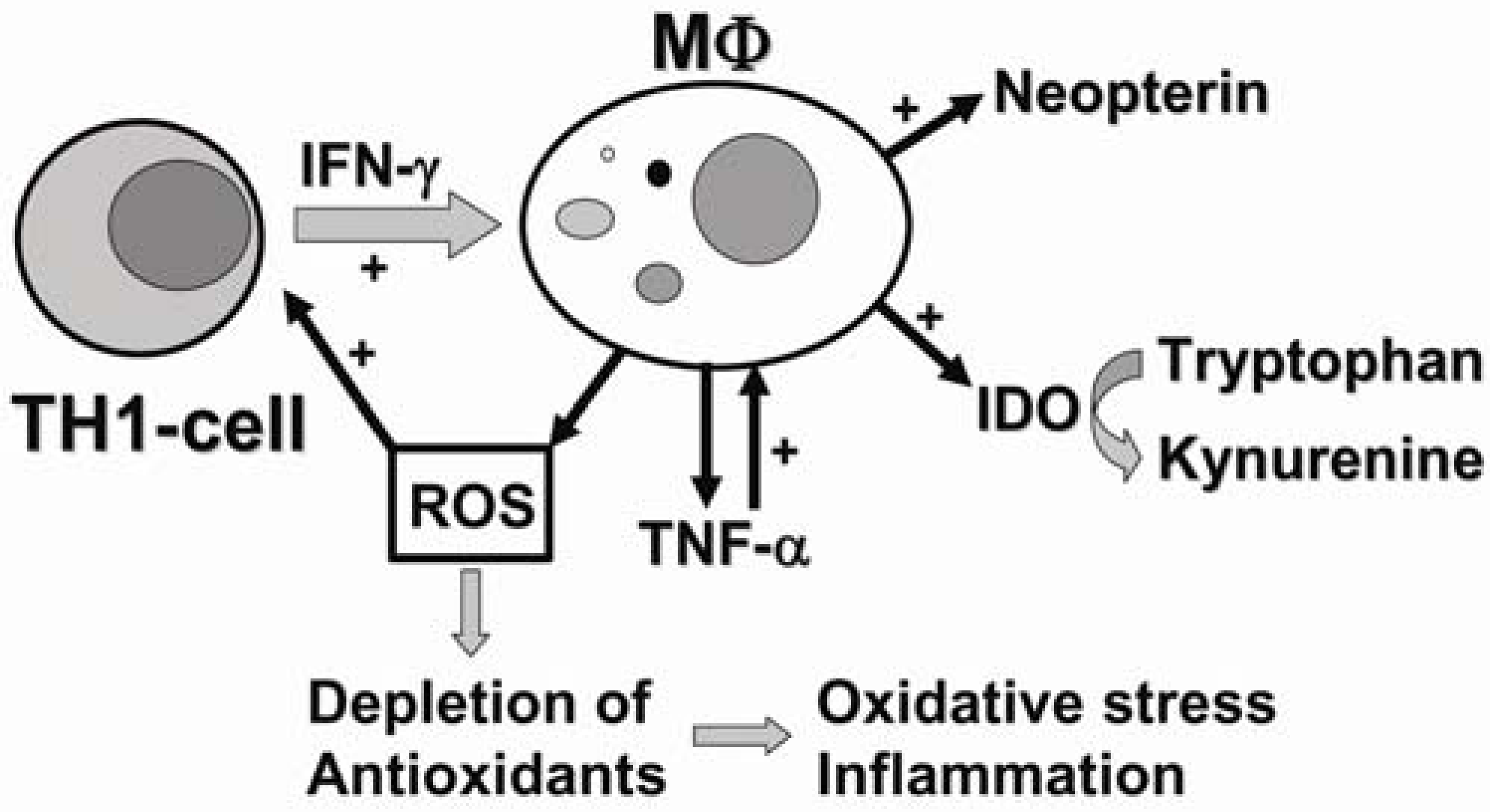
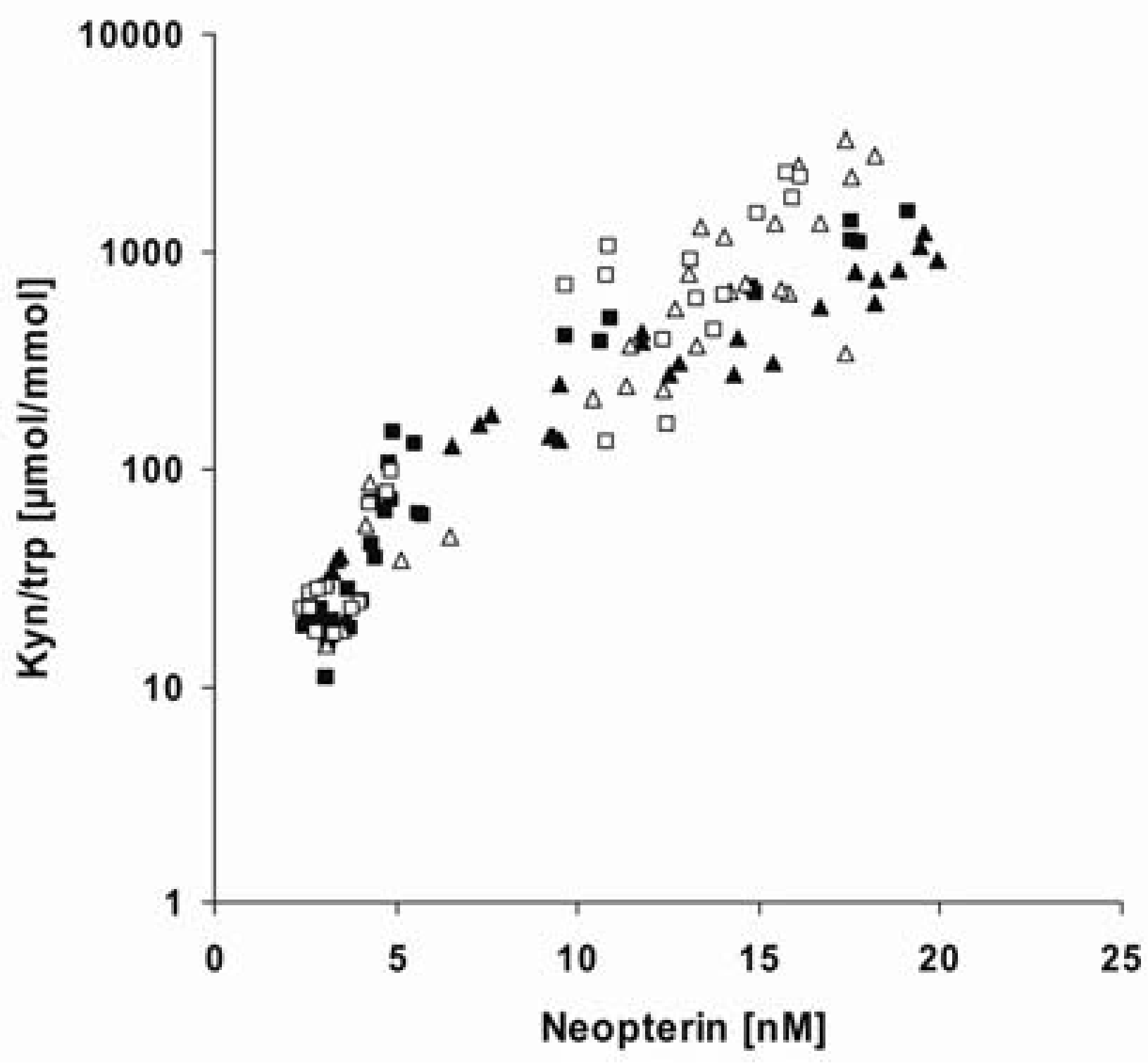
2. The Role of Tryptophan Metabolism in Diseases Associated with Inflammation
3. Involvement of Cannabinoid Receptor Signaling in Depression and Serotonergic Signaling
4. Conclusions
References
- Galiègue, S.; Mary, S.; Marchand, J.; Dussossoy, D.; Carriere, D.; Carayon, P.; Bouaboula, M.; Shire, D.; Le Fur, G.; Casellas, P. Expression of central and peripheral cannabinoid receptors in human immune tissues and leukocyte subpopulations. Eur. J. Biochem. 1995, 232, 54–61. [Google Scholar]
- Klein, T.W.; Newton, C.A.; Friedman, H. Cannabinoids and the immune system. Pain Res. Manag. 2001, 6, 95–101. [Google Scholar]
- Kusher, D.I.; Dawson, L.O.; Taylor, A.C.; Djeu, J.Y. Effect of the psychoactive metabolite of marijuana, delta-9-tetrahydrocannabinol (THC), on the synthesis of tumor necrosis factor by human large granular lymphocytes. Cell Immunol. 1994, 154, 99–108. [Google Scholar]
- McCoy, K.L.; Gainey, D.; Cabral, G.A. delta-9-Tetrahydrocannabinol modulates antigen processing by macrophages. J. Pharmacol. Exp. Ther. 1995, 273, 1216–1223. [Google Scholar]
- Kawakami, Y.; Klein, T.W.; Newton, C.; Djeu, J.Y.; Specter, S.; Friedman, H. Suppression by delta-9-tetrahydrocannabinol of interleukin 2-induced lymphocyte proliferation and lymphokine-activated killer cell activity. Int. J. Immunopharmacol. 1988, 10, 485–488. [Google Scholar]
- Fischer-Stenger, K.; Dove Pettit, D.A.; Cabral, G.A. Delta 9-tetrahydrocannabinol inhibition of tumor necrosis factor-alpha: suppression of posttranslational events. J. Pharmacol. Exp. Ther. 1993, 267, 1558–1565. [Google Scholar]
- Yuan, M.; Kiertscher, S.M.; Cheng, Q.; Zoumalan, R.; Tashkin, D.P.; Roth, M.D. Delta 9-Tetrahydrocannabinol regulates Th1/Th2 cytokine balance in activated human T cells. J. Neuroimmunol. 2002, 133, 124–131. [Google Scholar]
- Srivastava, M.D.; Srivastava, B.I.; Brouhard, B. Delta9 tetrahydrocannabinol and cannabidiol alter cytokine production by human immune cells. Immunopharmacol. 1998, 40, 179–185. [Google Scholar]
- Watzl, B.; Scuderi, P.; Watson, R.R. Marijuana components stimulate human peripheral blood mononuclear cell secretion of interferon-gamma and suppress interleukin-1 in vitro. Int. J. Immunopharmacol. 1991, 13, 1091–1097. [Google Scholar]
- Klein, T.W.; Newton, C.A.; Widen, R.; Friedman, H. The effect of delta-9-tetrahydrocannabinol and 11-hydroxy-delta-9-tetrahydrocannabinol on T-lymphocyte and B-lymphocyte mitogen responses. J. Immunopharmacol. 1985, 7, 451–466. [Google Scholar]
- Pross, S.H.; Nakano, Y.; Widen, R.; McHugh, S.; Newton, C.A.; Klein, T.W.; Friedman, H. Differing effects of delta-9-tetrahydrocannabinol (THC) on murine spleen cell populations dependent upon stimulators. Int. J. Immunopharmacol. 1992, 14, 1019–1027. [Google Scholar]
- Klein, T.W.; Lane, B.; Newton, C.A.; Friedman, H. The cannabinoid system and cytokine network. Proc. Soc. Exp. Biol. Med. 2000, 225, 1–8. [Google Scholar]
- Pertwee, R.G. The Pharmacology and Therapeutic Potential of Cannabidiol. In Cannabinoids; Di Marzo, V., Ed.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2004; pp. 32–83. [Google Scholar]
- Russo, E.B.; Guy, G.W. A tale of two cannabinoids: The therapeutic rationale for combining tetrahydrocannabinol and cannabidiol. Med. Hypotheses 2006, 66, 234–246. [Google Scholar]
- Consroe, P. Brain cannabinoid systems as targets for the therapy of neurological disorders. Neurobiol. Dis. 1998, 5, 534–551. [Google Scholar] [PubMed]
- Guimaraes, F.S.; de Aguiar, J.C.; Mechoulam, R.; Breuer, A. Anxiolytic effect of cannabidiol derivatives in the elevated plus-maze. Gen. Pharmacol. 1994, 25, 161–164. [Google Scholar] [PubMed]
- Zuardi, A.W.; Morais, S.L.; Guimaraes, F.S.; Mechoulam, R. Antipsychotic effect of cannabidiol. J. Clin.Psych. 1995, 56, 485–486. [Google Scholar]
- Hampson, A.J.; Grimaldi, M.; Axelrod, J.; Wink, D. Cannabidiol and (−)-Δ9-tetrahydrocannabinol are neuroprotective antioxidants. Proc. Natl. Acad. Sci. USA 1998, 95, 8268–8273. [Google Scholar]
- Jenny, M.; Santer, E.; Pirich, E.; Schennach, H.; Fuchs, D. Delta9-tetrahydrocannabinol and cannabidiol modulate mitogen-induced tryptophan degradation and neopterin formation in peripheral blood mononuclear cells in vitro. J. Neuroimmunol. 2009, 207, 75–82. [Google Scholar]
- Widner, B.; Werner, E.R.; Schennach, H.; Wachter, H.; Fuchs, D. Simultaneous measurement of serum tryptophan and kynurenine by HPLC. Clin. Chem. 1997, 43, 2424–2426. [Google Scholar]
- Maes, M.; Meltzer, H.Y. The Serotonin Hypothesis of Major Depression. In Psychopharmacoloy: The Fourth Generation of Progress; Bloom, F.E., Kupfer, D.J., Eds.; Raven Press: New York, NY, USA, 1995; pp. 933–944. [Google Scholar]
- Purselle, D.C.; Nemeroff, C.B. Serotonin transporter: A potential substrate in the biology of suicide. Neuropsychopharmacology 2003, 28, 613–619. [Google Scholar]
- Peters, J.C. Tryptophan nutrition and metabolism: an overview. Adv. Exp. Med. Biol. 1991, 294, 345–358. [Google Scholar]
- Stone, T.W.; Darlington, L.G. Endogenous kynurenines as targets for drug discovery and development. Nat. Rev. Drug Discov. 2002, 1, 609–620. [Google Scholar] [PubMed]
- Moffett, J.R.; Namboodiri, M.A. Tryptophan and the immune response. Immunol. Cell Biol. 2003, 81, 247–265. [Google Scholar]
- Taylor, M.W.; Feng, G.S. Relationship between interferon-gamma, indoleamine 2,3-dioxygenase and catabolism. FASEB J. 1991, 5, 2516–2522. [Google Scholar]
- Knox, W.E. The regulation of tryptophan pyrrolase activity by tryptophan. Adv. Enzyme Regul. 1966, 4, 287–297. [Google Scholar]
- Takikawa, O.; Yoshida, R.; Kido, R.; Hayaishi, O. Tryptophan degradation in mice initiated by indoleamine 2,3-dioxygenase. J. Biol. Chem. 1986, 261, 3648–3653. [Google Scholar]
- Byrne, G.I.; Lehmann, L.K.; Landry, G.J. Induction of tryptophan catabolism is the mechanism for gamma-interferon-mediated inhibition of intracellular Chlamydia psittaci replication in T24 cells. Infect. Immun. 1986, 53, 347–351. [Google Scholar]
- Werner, E.R.; Bitterlich, G.; Fuchs, D.; Hausen, A.; Reibnegger, G.; Szabo, G.; Dierich, M.P.; Wachter, H. Human macrophages degrade tryptophan upon induction by interferon-gamma. Life Sci. 1987, 41, 273–280. [Google Scholar]
- Young, S.N.; Leyton, M. The role of serotonin in human mood and social interaction. Insight from altered tryptophan levels. Pharmacol. Biochem. Behav. 2002, 71, 857–865. [Google Scholar]
- Pfefferkorn, E.R. Interferon gamma blocks the growth of Toxoplasma gondii in human fibroblasts by inducing the host cells to degrade tryptophan. Proc. Natl. Acad. Sci. USA 1986, 81, 908–912. [Google Scholar]
- Ozaki, Y.; Edelstein, M.P.; Duch, D.S. Induction of indoleamine 2,3-dioxygenase: a mechanism of the antitumor activity of interferon-gamma. Proc. Natl. Acad. Sci. USA 1988, 85, 1242–1246. [Google Scholar]
- Romagnani, S. Regulation of the T cell response. Clin. Exp. Allergy 2006, 36, 1357–1366. [Google Scholar]
- Wood, K.J.; Sawitzki, B. Interferon gamma: a crucial role in the function of induced regulatory T cells in vivo. Trends Immunol. 2006, 27, 183–187. [Google Scholar]
- Nathan, C.F. Peroxide and pteridine: A hypothesis on the regulation of macrophage antimicrobial activity by interferon gamma. Interferon 1986, 7, 125–143. [Google Scholar]
- Nathan, C.F.; Murray, H.W.; Wiebe, M.E.; Rubin, B.Y. Identification of interferon-gamma as the lymphokine that activates human macrophage oxidative metabolism and antimicrobial activity. J. Exp. Med. 1983, 158, 670–689. [Google Scholar]
- Schreck, R.; Rieber, P.; Baeuerle, P.A. Reactive oxygen intermediates as apparently widely used messengers in the activation of the NF-kappa B transcription factor and HIV-1. EMBO J. 1991, 10, 2247–2258. [Google Scholar]
- Min, K.J.; Jou, I.; Joe, E. Plasminogen-induced IL-1beta and TNF-alpha production in microglia is regulated by reactive oxygen species. Biochem. Biophys. Res. Commun. 2003, 312, 969–974. [Google Scholar]
- Baier-Bitterlich, G.; Wachter, H.; Fuchs, D. Role of neopterin and 7,8-dihydroneopterin in human immunodeficiency virus infection: marker for disease progression and pathogenic link. J. Acquir. Immune. Defic. Syndr. Hum. Retrovirol. 1996, 13, 184–193. [Google Scholar]
- Wirleitner, B.; Neurauter, G.; Schroecksnadel, K.; Frick, B.; Fuchs, D. Interferon-gamma-induced conversion of tryptophan: immunologic and neuropsychiatric aspects. Curr. Med. Chem. 2003, 10, 1581–1591. [Google Scholar]
- Pfefferkorn, E.R.; Eckel, M.; Rebhun, S. Interferon-gamma suppresses the growth of Toxoplasma gondii in human fibroblasts through starvation for tryptophan. Mol. Biochem. Parasitol. 1986, 20, 215–224. [Google Scholar]
- Bodaghi, B.; Goureau, O.; Zipeto, D.; Laurent, L.; Virelizier, J.L.; Michelson, S. Role of IFN-gamma-induced indoleamine 2,3-dioxygenase and inducible nitric oxide synthase in the replication of human cytomegalovirus in retinal pigment epithelial cells. J. Immunol. 1999, 162, 957–964. [Google Scholar]
- de la Maza, L.M.; Peterson, E.M. Dependence of the in vitro antiproliferative activity of recombinant human gamma-interferon on the concentration of tryptophan in culture media. Cancer Res. 1988, 48, 346–350. [Google Scholar]
- Ozaki, Y.; Edelstein, M.P.; Duch, D.S. Induction of indoleamine 2,3-dioxygenase: a mechanism of the antitumor activity of interferon-gamma. Proc. Natl. Acad. Sci. USA 1988, 85, 1242–1246. [Google Scholar]
- Munn, D.H.; Shafizadeh, E.; Attwood, J.T.; Bondarev, I.; Pashine, A.; Mellor, A.L. Inhibition of T-cell proliferation by macrophage tryptophan catabolism. J. Exp. Med. 1999, 189, 1663–1672. [Google Scholar]
- Mellor, A.L.; Munn, D.H. Tryptophan catabolism and T-cell tolerance: immunosuppression by starvation? Immunol. Today 1999, 20, 469–473. [Google Scholar]
- Werner-Felmayr, G.; Werner, E.R.; Fuchs, D.; Hausen, A.; Reibnegger, G.; Wachter, H. Characteristics of interferon induced tryptophan metabolism in human cells in vitro. Biochim. Biophys. Acta 1989, 1012, 140–147. [Google Scholar]
- Werner-Felmayer, G.; Werner, E.R.; Fuchs, D.; Hausen, A.; Reibnegger, G.; Wachter, H. Tumour necrosis factor-alpha and lipopolysaccharide enhance interferon-induced tryptophan degradation and pteridine synthesis in human cells. Biol. Chem. Hoppe Seyler 1989, 370, 1063–1069. [Google Scholar]
- Huber, C.; Batchelor, J.R.; Fuchs, D.; Hausen, A.; Lang, A.; Niederwieser, D.; Reibnegger, G.; Swetly, P.; Troppmair, J.; Wachter, H. Immune response-associated production of neopterin-Release from macrophages primarily under control of interferon-gamma. J. Exp. Med. 1984, 160, 310–316. [Google Scholar]
- Zangerle, R.; Fuchs, D.; Reibnegger, G.; Fritsch, P.; Wachter, H. Markers for disease progression in intravenous drug users infected with HIV-1. AIDS 1991, 5, 985–991. [Google Scholar]
- Murr, C.; Widner, B.; Wirleitner, B.; Fuchs, D. Neopterin as a marker for immune system activation. Curr. Drug Metabol. 2002, 3, 175–187. [Google Scholar]
- Schroecksnadel, K.; Wirleitner, B.; Winkler, C.; Fuchs, D. Monitoring tryptophan metabolism in chronic immune activation. Clin. Chim. Acta 2006, 364, 82–90. [Google Scholar]
- Huang, A.; Fuchs, D.; Widner, B.; Glover, C.; Henderson, D.C.; Allen-Mersh, T.G. Serum tryptophan decrease correlates with immune activation and impaired quality of life in colorectal cancer. Brit. J. Cancer. 2002, 86, 1691–1696. [Google Scholar]
- Capuron, L.; Neurauter, G.; Musselman, D.L.; Lawson, D.H.; Nemeroff, C.B.; Fuchs, D.; Miller, A.H. Interferon-alpha-induced changes in tryptophan metabolism. Relationship to depression and paroxetine treatment. Biol. Psychiatry 2003, 54, 906–914. [Google Scholar]
- Myint, A.M.; Kim, Y.K.; Verkerk, R.; Scharpe, S.; Steinbusch, H.; Leonard, B. Kynurenine pathway in major depression: Evidence of impaired neuroprotection. J. Affect. Disord. 2007, 98, 143–151. [Google Scholar]
- Murr, C.; Widner, B.; Sperner-Unterweger, B.; Ledochowski, M.; Schubert, C.; Fuchs, D. Immune reaction links disease progression in cancer patients with depression. Med. Hypotheses 2000, 55, 137–140. [Google Scholar]
- Widner, B.; Laich, A.; Sperner-Unterweger, B.; Ledochowski, M.; Fuchs, D. Neopterin production tryptophan degradation and mental depression: what is the link? Brain Behav. Immun. 2002, 16, 590–595. [Google Scholar]
- Russo, S.; Kema, I.P.; Fokkema, M.R.; Boon, J.C.; Willemse, P.H.; de Vries, E.G.; den Boer, J.A.; Korf, J. Tryptophan as a link between psychopathology and somatic states. Psychosom. Med. 2003, 65, 665–671. [Google Scholar]
- Hall, W.; Solowij, N. Adverse effects of cannabis. Lancet 1998, 352, 1611–1616. [Google Scholar]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders: DSM-IV, 4th ed.; American Psychiatric Press: Washington, DC, USA, 1994. [Google Scholar]
- Viveros, M.P.; Marco, E.M.; File, S.E. Endocannabinoid system and stress and anxiety responses. Pharmacol. Biochem. Behav. 2005, 81, 331–342. [Google Scholar]
- Patel, S.; Hillard, C.J. Pharmacological evaluation of cannabinoid receptor ligands in a mouse model of anxiety: further evidence for an anxiolytic role for endogenous cannabinoid signaling. J. Pharmacol. Exp. Ther. 2006, 318, 304–311. [Google Scholar]
- Viveros, M.P.; Marco, E.M.; Llorente, R.; Lamota, L. The role of the hippocampus in mediating emotional responses to nicotine and cannabinoids: a possible neural substrate for functional interactions. Behav. Pharmacol. 2007, 18, 375–389. [Google Scholar]
- Rubino, T.; Sala, M.; Viganò, D.; Braida, D.; Castiglioni, C.; Limonta, V.; Guidali, C.; Realini, N.; Parolaro, D. Cellular mechanisms underlying the anxiolytic effect of low doses of peripheral Delta9-tetrahydrocannabinol in rats. Neuropsychopharmacology 2007, 32, 2036–2045. [Google Scholar]
- Onaivi, E.S.; Green, M.R.; Martin, B.R. Pharmacological characterization of cannabinoids in the elevated plus maze. J. Pharmacol. Exp. Ther. 1990, 253, 1002–1009. [Google Scholar]
- Rodriguez de Fonseca, F.; Rubio, P.; Menzaghi, F.; Merlo-Pich, E.; Rivier, J.; Koob, G.F.; Navarro, M. Corticotropin-releasing factor (CRF) antagonist [D-Phe12, Nle21,38,C alpha MeLeu37] CRF attenuates the acute actions of the highly potent cannabinoid receptor agonist HU-210 on defensive-withdrawal behavior in rats. J. Pharmacol. Exp. Ther. 1996, 276, 56–64. [Google Scholar]
- Morales, M. Cannabinoids and the central serotonergic system. In Endocannabinoids: the Brain and the Body's Marijuana and Beyond; Onaivi, E., Sugiura, T., Di Marzo, V., Eds.; Taylor and Francis, Boca Raton: Florida, FL, USA, 2006; pp. 249–260. [Google Scholar]
- Gobbi, G.; Bambico, F.R.; Mangieri, R.; Bortolato, M.; Campolongo, P.; Solinas, M.; Cassano, T.; Morgese, M.G.; Debonnel, G.; Duranti, A.; et al. Antidepressant-like activity and modulation of brain monoaminergic transmission by blockade of anandamide hydrolysis. Proc. Natl. Acad. Sci. USA 2005, 102, 18620–18625. [Google Scholar]
- Hill, M.N.; Gorzalka, B.B. Is there a role for the endocannabinoid system in the etiology and treatment of melancholic depression? Behav. Pharmacol. 2005, 16, 333–352. [Google Scholar]
- Pacher, P.; Kecskemeti, V. Trends in the development of new antidepressants. Is there a light at the end of the tunnel? Curr. Med. Chem. 2004, 11, 925–943. [Google Scholar]
- Martin, M.; Ledent, C.; Parmentier, M.; Maldonado, R.; Valverde, O. Involvement of CB1 cannabinoid receptors in emotional behaviour. Psychopharmacology 2002, 159, 379–387. [Google Scholar]
- Bambico, F.R.; Gobbi, G. The cannabinoid CB1 receptor and the endocannabinoid anandamide: possible antidepressant targets. Expert Opin. Ther. Targets 2008, 12, 1347–1366. [Google Scholar]
- Sagredo, O.; Ramos, J.A.; Fernández-Ruiz, J.; Rodriguez, M.L.; de Miguel, R. Chronic Delta-9-tetrahydrocannabinol administration affects serotonin levels in the rat frontal cortex. Naunyn. Schmiedebergs Arch. Pharmacol. 2006, 372, 313–317. [Google Scholar]
- Griebel, G.; Stemmelin, J.; Scatton, B. Effects of the cannabinoid CB1 receptor antagonist rimonabant in models of emotional reactivity in rodents. Biol. Psychiatry 2005, 57, 261–267. [Google Scholar]
- Steiner, M.A.; Marsicano, G.; Nestler, E.J.; Holsboer, F.; Lutz, B.; Wotjak, C.T. Antidepressant-like behavioral effects of impaired cannabinoid receptor type 1 signaling coincide with exaggerated corticosterone secretion in mice. Psychoneuroendocrinology 2008, 33, 54–67. [Google Scholar]
- Rutkowska, M.; Jachimczuk, O. Antidepressant-like properties of ACEA (arachidonyl-2-chloroethylamide), the selective agonist of CB1 receptors. Acta Pol. Pharm. 2004, 61, 165–167. [Google Scholar]
- Hill, M.N.; Gorzalka, B.B. Pharmacological enhancement of cannabinoid CB1 receptor activity elicits an antidepressant-like response in the rat forced swim test. Eur. Neuropsychopharmacol. 2005, 15, 593–599. [Google Scholar]
- Bortolato, M.; Mangieri, R.A.; Fu, J.; Kim, J.H.; Arguello, O.; Duranti, A.; Tontini, A.; Mor, M.; Tarzia, G.; Piomelli, D. Antidepressant-like activity of the fatty acid amide hydrolase inhibitor URB597 in a rat model of chronic mild stress. Biol. Psychiatry 2007, 62, 1103–1110. [Google Scholar]
- Steffens, M.; Feuerstein, T.J. Receptor-independent depression of DA and 5-HT uptake by cannabinoids in rat neocortex-involvement of Na(+)/K(+)-ATPase. Neurochem. Int. 2004, 44, 529–538. [Google Scholar]
- Molina-Holgado, F.; Molina-Holgado, E.; Leret, M.L.; Gonzalez, M.I.; Reader, T.A. Distribution of indoleamines and [3H]paroxetine binding in rat brain regions following acute or perinatal delta 9-tetrahydrocannabinol treatments. Neurochem. Res. 1993, 18, 1183–1191. [Google Scholar]
- Tzavara, E.T.; Davis, R.J.; Perry, K.W.; Li, X.; Salhoff, C.; Bymaster, F.P.; Witkin, J.M.; Nomikos, G.G. The CB1 receptor antagonist SR141716A selectively increases monoaminergic neurotransmission in the medial prefrontal cortex: implications for therapeutic actions. Br. J. Pharmacol. 2003, 138, 544–553. [Google Scholar]
- Mendiguren, A.; Pineda, J. Effect of the CB(1) receptor antagonists rimonabant and AM251 on the firing rate of dorsal raphe nucleus neurons in rat brain slices. Br. J. Pharmacol. 2009, 158, 1579–1587. [Google Scholar]
- McLaughlin, R.J.; Hill, M.N.; Gorzalka, B.B. Monoaminergic neurotransmission contributes to cannabinoid-induced activation of the hypothalamic-pituitary-adrenal axis. Eur. J. Pharmacol. 2009, 624, 71–76. [Google Scholar]
- Campos, A.C.; Guimarães, F.S. Involvement of 5HT1A receptors in the anxiolytic-like effects of cannabidiol injected into the dorsolateral periaqueductal gray of rats. Psychopharmacology 2008, 199, 223–230. [Google Scholar]
- Resstel, L.B.; Tavares, R.F.; Lisboa, S.F.; Joca, S.R.; Corrêa, F.M.; Guimarães, F.S. 5-HT1A receptors are involved in the cannabidiol-induced attenuation of behavioural and cardiovascular responses to acute restraint stress in rats. Br. J. Pharmacol. 2009, 156, 181–188. [Google Scholar]
- Després, J.P.; Lemieux, I.; Alméras, N. Contribution of CB1 blockade to the management of high-risk abdominal obesity. Int. J. Obes. 2006, 30, 44–52. [Google Scholar]
- Christensen, R.; Kristensen, P.K.; Bartels, E.M.; Bliddal, H.; Astrup, A. Efficacy and safety of the weight-loss drug rimonabant: a meta-analysis of randomised trials. Lancet 2007, 370, 1706–1713. [Google Scholar]
- Moreira, F.A.; Crippa, J.A. The psychiatric side-effects of rimonabant. Rev. Bras. Psiquiatr. 2009, 31, 145–153. [Google Scholar] [Green Version]
- Després, J.P.; Golay, A.; Sjöström, L. Effects of rimonabant on metabolic risk factors in overweight patients with dyslipidemia. N. Engl. J. Med. 2005, 353, 2121–2134. [Google Scholar]
- Van Gaal, L.F.; Scheen, A.J.; Rissanen, A.M.; Rössner, S.; Hanotin, C.; Ziegler, O. Long-term effect of CB1 blockade with rimonabant on cardiometabolic risk factors: two year results from the RIO-Europe Study. Eur. Heart. J. 2008, 29, 1761–1771. [Google Scholar]
- Beyer, C.E.; Dwyer, J.M.; Piesla, M.J.; Platt, B.J.; Shen, R.M.; Rahman, Z.; Chan, K.; Manners, M.T.; Samad, T.A.; Kennedy, J.D.; et al. Depression-like phenotype following chronic CB1 receptor antagonism. Neurobiol. Dis. 2010, 39, 148–155. [Google Scholar]
- Degenhardt, L.; Hall, W.; Lynskey, M. Exploring the association between cannabis use and depression. Addiction 2003, 98, 1493–1504. [Google Scholar]
- Gruber, A.J.; Pope, H.G., Jr.; Brown, M.E. Do patients use marijuana as an antidepressant? Depression 1996, 4, 77–80. [Google Scholar]
- Woolridge, E.; Barton, S.; Samuel, J.; Osorio, J.; Dougherty, A.; Holdcroft, A. Cannabis use in HIV for pain and other medical symptoms. J. Pain Symptom Manage. 2005, 29, 358–367. [Google Scholar]
- Ware, M.A.; Adams, H.; Guy, G.W. The medicinal use of cannabis in the UK: results of a nationwide survey. Int. J. Clin. Pract. 2005, 59, 291–295. [Google Scholar]
- Iwagaki, H.; Hizuta, A.; Tanaka, N.; Orita, K. Decreased serum tryptophan in patients with cancer cachexia correlates with increased serum neopterin. Immunol. Invest. 1995, 24, 467–478. [Google Scholar]
- Dantzer, R.; O'Connor, J.C.; Freund, G.G.; Johnson, R.W.; Kelley, K.W. From inflammation to sickness and depression: when the immune system subjugates the brain. Nat. Rev. Neurosci. 2008, 9, 46–56. [Google Scholar]
- Reilly, J.G.; McTavish, S.F.; Young, A.H. Rapid depletion of plasma tryptophan: a review of studies and experimental methodology. J. Psychopharmacol. 1997, 11, 381–392. [Google Scholar]
- Delgado, P.L.; Price, L.H.; Miller, H.L.; Salomon, R.M.; Aghajanian, G.K.; Heninger, G.R.; Charney, D.S. Serotonin and the neurobiology of depression. Effects of tryptophan depletion in drug-free depressed patients. Arch. Gen. Psychiatry 1994, 51, 865–874. [Google Scholar]
- Ben Amar, M. Cannabinoids in medicine: A review of their therapeutic potential. J. Ethnopharmacol. 2006, 21, 1–25. [Google Scholar]
- Cunha, J.M.; Carlini, E.A.; Pereira, A.E.; Ramos, O.L.; Pimentel, C.; Gagliardi, R.; Sanvito, W.L.; Lander, N.; Mechoulam, R. Chronic administration of cannabidiol to healthy volunteers and epileptic patients. Pharmacology 1980, 21, 175–185. [Google Scholar]
- Consroe, P.; Laguna, J.; Allender, J.; Snider, S.; Stern, L.; Sandyk, R.; Kennedy, K.; Schram, K. Controlled clinical trial of cannabidiol in Huntington's disease. Pharmacol. Biochem. Behav. 1991, 40, 701–708. [Google Scholar]
- Berger, M.; Gray, J.A.; Roth, B.L. The expanded biology of serotonin. Annu. Rev. Med. 2009, 60, 355–366. [Google Scholar]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an Open Access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jenny, M.; Schröcksnadel, S.; Überall, F.; Fuchs, D. The Potential Role of Cannabinoids in Modulating Serotonergic Signaling by Their Influence on Tryptophan Metabolism. Pharmaceuticals 2010, 3, 2647-2660. https://doi.org/10.3390/ph3082647
Jenny M, Schröcksnadel S, Überall F, Fuchs D. The Potential Role of Cannabinoids in Modulating Serotonergic Signaling by Their Influence on Tryptophan Metabolism. Pharmaceuticals. 2010; 3(8):2647-2660. https://doi.org/10.3390/ph3082647
Chicago/Turabian StyleJenny, Marcel, Sebastian Schröcksnadel, Florian Überall, and Dietmar Fuchs. 2010. "The Potential Role of Cannabinoids in Modulating Serotonergic Signaling by Their Influence on Tryptophan Metabolism" Pharmaceuticals 3, no. 8: 2647-2660. https://doi.org/10.3390/ph3082647