
TRPV3 in Drug Development

 and
and
Abstract
:1. Introduction
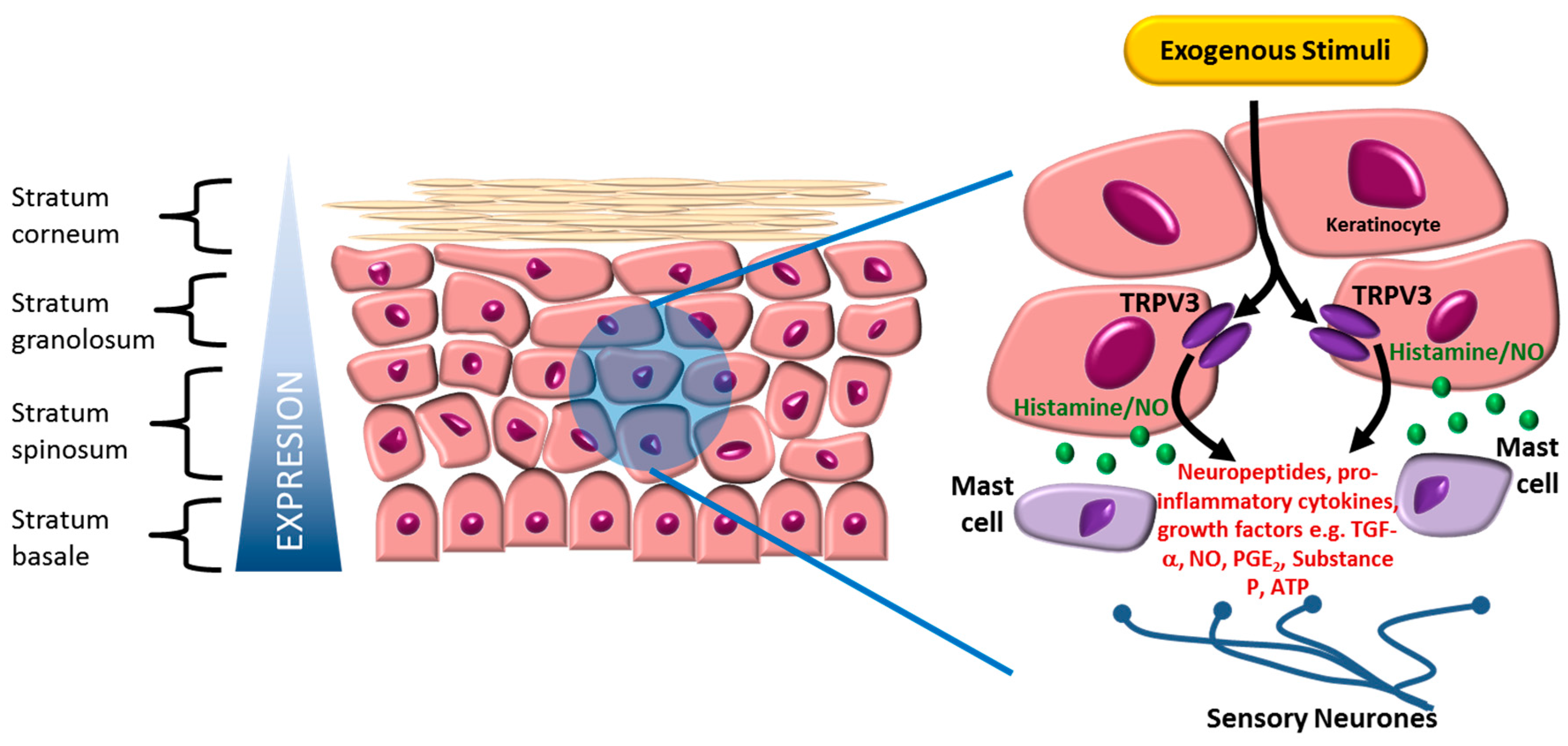
2. Expression and Function of TRPV3
3. Insights from Genetic Mutations
4. TRPV3 Indications
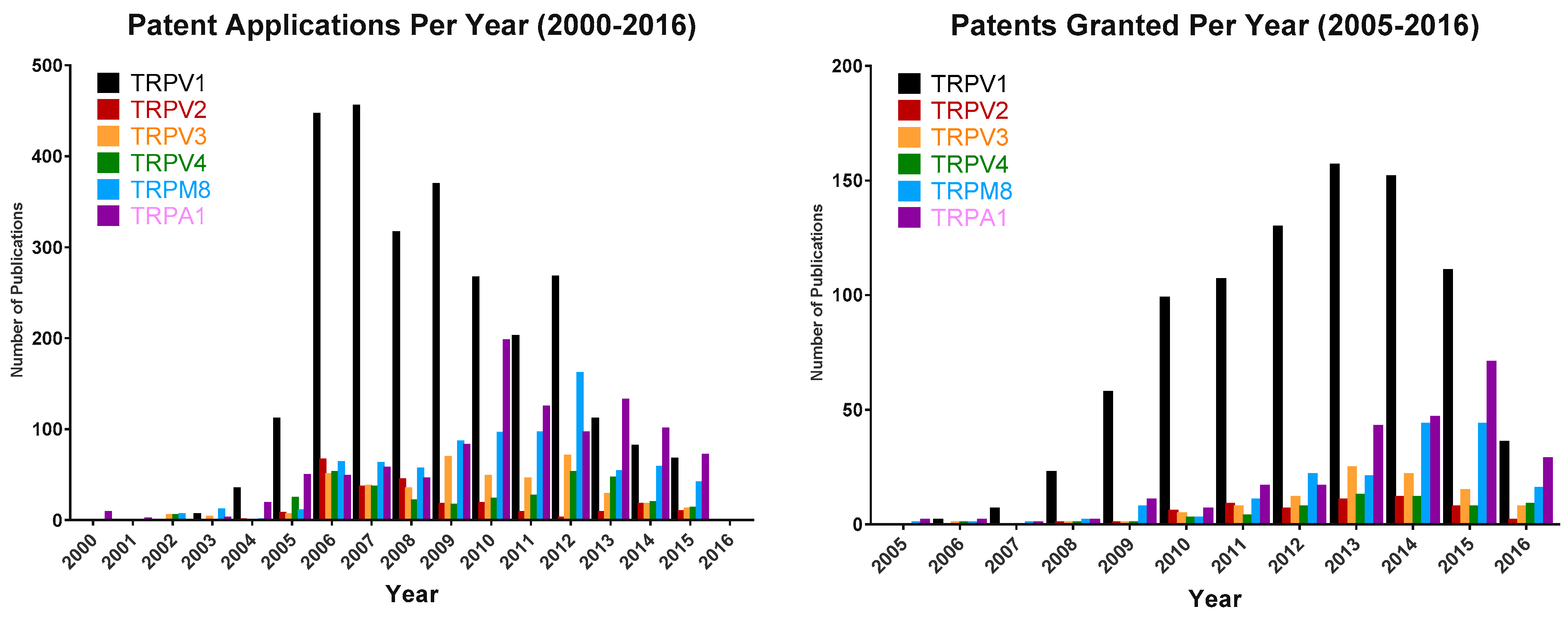
5. TRPV3 Drug Development Overview
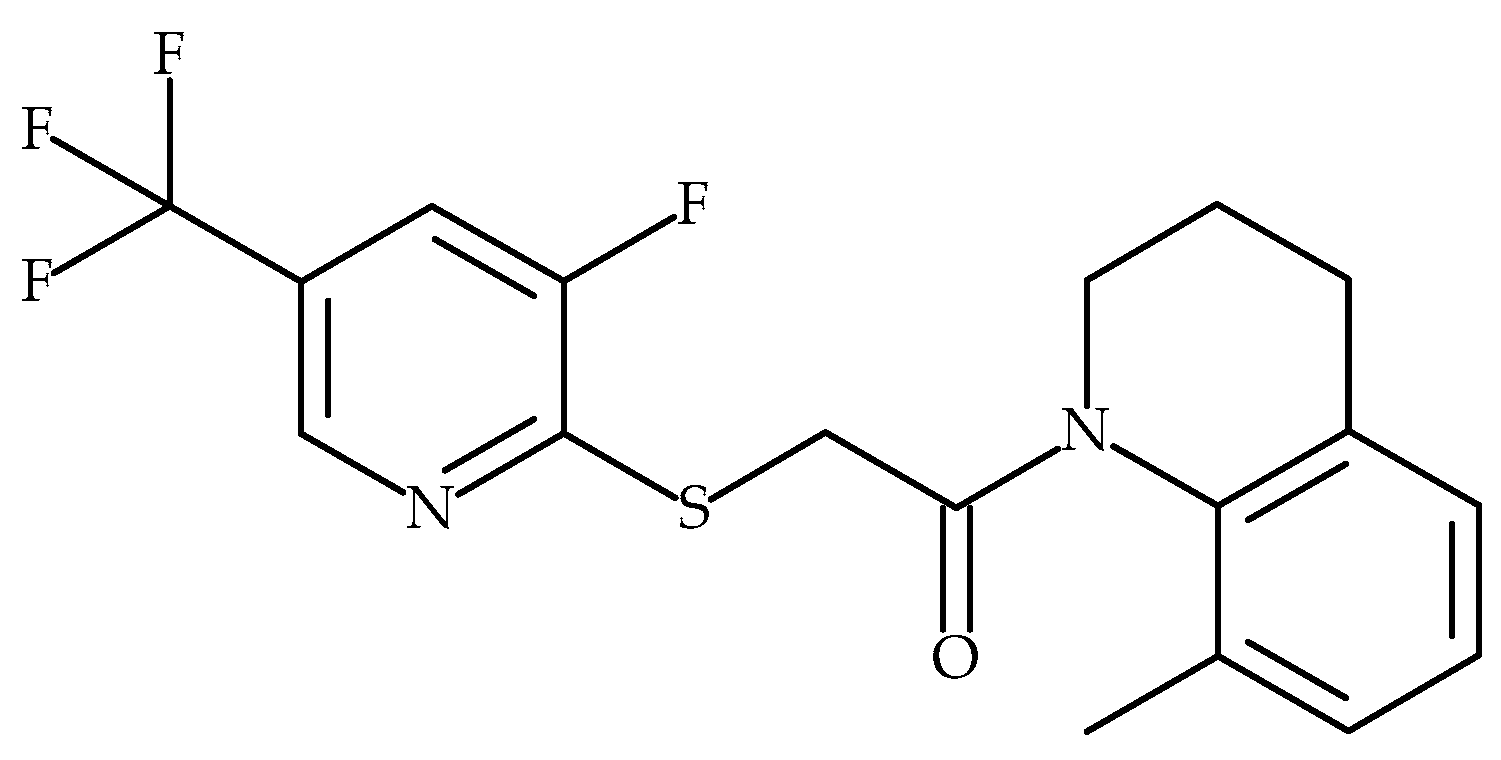
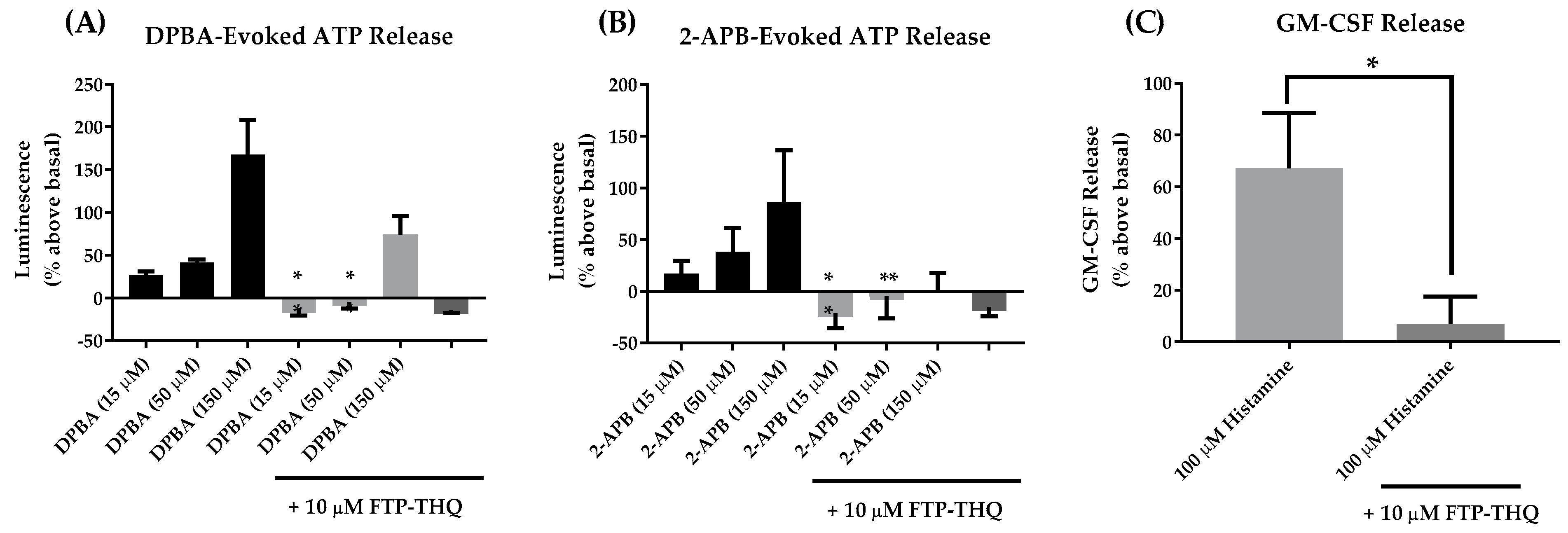
5.1. Glenmark Pharmaceuticals Ltd.
5.2. Hydra Biosciences Inc.
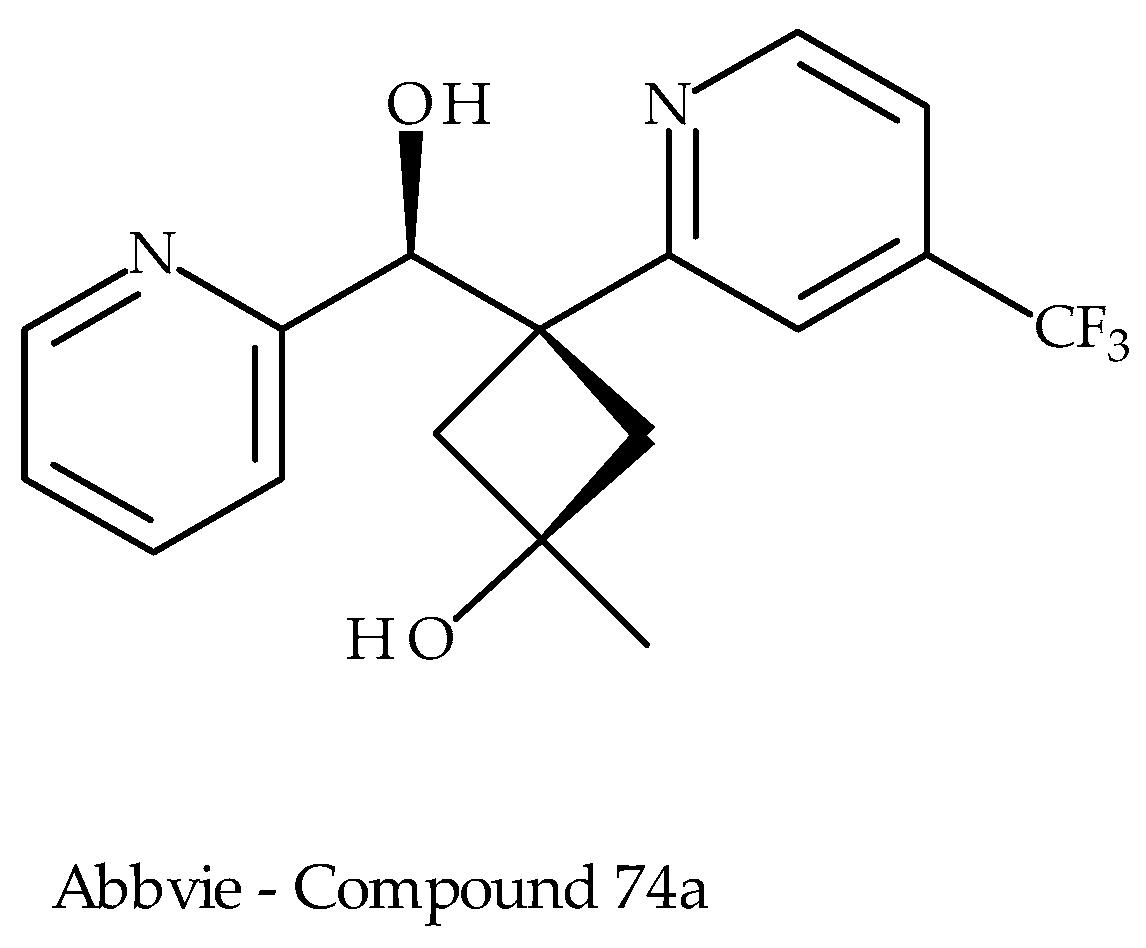
5.3. Abbvie Inc.
5.4. Miscellaneous
6. Conclusions and Outlook
Supplementary Materials
Acknowledgments
Conflicts of Interest
References
- Peier, A.M.; Reeve, A.J.; Andersson, D.A.; Moqrich, A.; Earley, T.J.; Hergarden, A.C.; Story, G.M.; Colley, S.; Hogenesch, J.B.; McIntyre, P.; et al. A heat-sensitive TRP channel expressed in keratinocytes. Science 2002, 296, 2046–2049. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Ramsey, I.S.; Kotecha, S.A.; Moran, M.M.; Chong, J.A.; Lawson, D.; Ge, P.; Lilly, J.; Silos-Santiago, I.; Xie, Y.; et al. TRPV3 is a calcium-permeable temperature-sensitive cation channel. Nature 2002, 418, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.D.; Gunthorpe, M.J.; Kelsell, R.E.; Hayes, P.D.; Reilly, P.; Facer, P.; Wright, J.E.; Jerman, J.C.; Walhin, J.-P.; Ooi, L.; et al. TRPV3 is a temperature-sensitive vanilloid receptor-like protein. Nature 2002, 418, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Yang, F.; Takanishi, C.L.; Zheng, J. Thermosensitive TRPV channel subunits coassemble into heteromeric channels with intermediate conductance and gating properties. J. Gen. Physiol. 2007, 129, 16. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Yang, F.; Liu, S.; Colton, C.K.; Wang, C.; Cui, Y.; Cao, X.; Zhu, M.X.; Sun, C.; Wang, K.; et al. Heteromeric heat-sensitive transient receptor potential channels exhibit distinct temperature and chemical response. J. Biol. Chem. 2012, 287, 7279–7288. [Google Scholar] [CrossRef] [PubMed]
- Hellwig, N.; Albrecht, N.; Harteneck, C.; Gunter, S.; Schaefer, M. Homo- and heteromeric assembly of TRP channel subunits. J. Cell. Sci. 2005, 118, 917–928. [Google Scholar] [CrossRef] [PubMed]
- Grandl, J.; Hu, H.; Bandell, M.; Bursulaya, B.; Schmidt, M.; Petrus, M.; Patapoutian, A. Pore region of TRPV3 ion channel is specifically required for heat activation. Nat. Neurosci. 2008, 11, 1007–1013. [Google Scholar] [CrossRef] [PubMed]
- Xiao, R.; Tang, J.; Wang, C.; Colton, C.K.; Tian, J.; Zhu, M.X. Calcium plays a central role in the sensitization of TRPV3 channel to repetitive stimulations. J. Biol. Chem. 2008, 283, 6162–6174. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Grandl, J.; Bandell, M.; Petrus, M.; Patapoutian, A. Two amino acid residues determine 2-APB sensitivity of the ion channels TRPV3 and TRPV4. Proc. Natl. Acad. Sci. USA 2009, 106, 1626–1631. [Google Scholar] [CrossRef] [PubMed]
- Phelps, C.B.; Wang, R.R.; Choo, S.S.; Gaudet, R. Differential regulation of TRPV1, TRPV3, and TRPV4 sensitivity through a conserved binding site on the ankyrin repeat domain. J. Biol. Chem. 2010, 285, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Doerner, J.F.; Hatt, H.; Ramsey, I.S. Voltage- and temperature-dependent activation of TRPV3 channels is potentiated by receptor-mediated PI(4,5)P2 hydrolysis. J. Gen. Physiol. 2011, 137, 271–288. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Stewart, R.; Berdeaux, R.; Hu, H. Tonic inhibition of TRPV3 by Mg2+ in mouse epidermal keratinocytes. J. Investig. Dermatol. 2012, 132, 2158–2165. [Google Scholar] [CrossRef] [PubMed]
- Sherkheli, M.A.; Vogt-Eisele, A.K.; Weber, K.; Hatt, H. Camphor modulates TRPV3 cation channels activity by interacting with critical pore-region cysteine residues. Pak. J. Pharm. Sci. 2013, 26, 431–438. [Google Scholar] [PubMed]
- Liu, B.; Yao, J.; Zhu, M.X.; Qin, F. Hysteresis of gating underlines sensitization of TRPV3 channels. J. Gen. Physiol. 2011, 138, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Vogt-Eisele, A.K.; Weber, K.; Sherkheli, M.A.; Vielhaber, G.; Panten, J.; Gisselmann, G.; Hatt, H. Monoterpenoid agonists of TRPV3. Br. J. Pharmacol. 2007, 151, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Bang, S.; Yoo, S.; Yang, T.-J.; Cho, H.; Hwang, S.W. Farnesyl pyrophosphate is a novel pain-producing molecule via specific activation of TRPV3. J. Biol. Chem. 2010, 285, 19362–19371. [Google Scholar] [CrossRef] [PubMed]
- Moqrich, A.; Hwang, S.W.; Earley, T.J.; Petrus, M.J.; Murray, A.N.; Spencer, K.S.R.; Andahazy, M.; Story, G.M.; Patapoutian, A. Impaired thermosensation in mice lacking TRPV3, a heat and camphor sensor in the skin. Science 2005, 307, 1468–1472. [Google Scholar] [CrossRef] [PubMed]
- Grubisha, O.; Mogg, A.J.; Sorge, J.L.; Ball, L.-J.; Sanger, H.; Ruble, C.L.A.; Folly, E.A.; Ursu, D.; Broad, L.M. Pharmacological profiling of the TRPV3 channel in recombinant and native assays. Br. J. Pharmacol. 2014, 171, 2631–2644. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Delling, M.; Jun, J.C.; Clapham, D.E. Oregano, thyme and clove-derived flavors and skin sensitizers activate specific TRP channels. Nat. Neurosci. 2006, 9, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.-Z.; Xiao, R.; Wang, C.; Gao, N.; Colton, C.K.; Wood, J.D.; Zhu, M.X. Potentiation of TRPV3 channel function by unsaturated fatty acids. J. Cell. Physiol. 2006, 208, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Yang, F.; Zheng, J.; Wang, K. Intracellular proton-mediated activation of TRPV3 channels accounts for the exfoliation effect of alpha-hydroxyl acids on keratinocytes. J. Biol. Chem. 2012, 287, 25905–25916. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Yang, P.; Qin, P.; Lu, Y.; Li, X.; Tian, Q.; Li, Y.; Xie, C.; Tian, J.-B.; Zhang, C.; et al. Selective potentiation of 2-APB-induced activation of TRPV1–3 channels by acid. Sci. Rep. 2016, 6, 20791. [Google Scholar] [CrossRef] [PubMed]
- Bang, S.; Yoo, S.; Yang, T.-J.; Cho, H.; Hwang, S.W. Isopentenyl pyrophosphate is a novel antinociceptive substance that inhibits TRPV3 and TRPA1 ion channels. Pain 2011, 152, 1156–1164. [Google Scholar] [CrossRef] [PubMed]
- Bang, S.; Yoo, S.; Yang, T.J.; Cho, H.; Hwang, S.W. 17(R)-resolvin D1 specifically inhibits transient receptor potential ion channel vanilloid 3 leading to peripheral antinociception. Br. J. Pharmacol. 2012, 165, 683–692. [Google Scholar] [CrossRef] [PubMed]
- de Petrocellis, L.; Orlando, P.; Moriello, A.S.; Aviello, G.; Stott, C.; Izzo, A.A.; di Marzo, V. Cannabinoid actions at TRPV channels: Effects on TRPV3 and TRPV4 and their potential relevance to gastrointestinal inflammation. Acta Physiol. 2012, 204, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.-K.; Guler, A.D.; Caterina, M.J. Biphasic currents evoked by chemical or thermal activation of the heat-gated ion channel, TRPV3. J. Biol. Chem. 2005, 280, 15928–15941. [Google Scholar] [CrossRef] [PubMed]
- Grandl, J.; Kim, S.E.; Uzzell, V.; Bursulaya, B.; Petrus, M.; Bandell, M.; Patapoutian, A. Temperature-induced opening of TRPV1 ion channel is stabilized by the pore domain. Nat. Neurosci. 2010, 13, 708–714. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.E.; Patapoutian, A.; Grandl, J. Single residues in the outer pore of TRPV1 and TRPV3 have temperature-dependent conformations. PLoS ONE 2013, 8, e59593. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Zhu, M.X. TRPV3. Handb. Exp. Pharmacol. 2014, 222, 273–291. [Google Scholar] [PubMed]
- Huang, S.M.; Lee, H.; Chung, M.-K.; Park, U.; Yu, Y.Y.; Bradshaw, H.B.; Coulombe, P.A.; Walker, J.M.; Caterina, M.J. Overexpressed transient receptor potential vanilloid 3 ion channels in skin keratinocytes modulate pain sensitivity via prostaglandin E2. J. Neurosci. 2008, 28, 13727–13737. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, M.; Yoshioka, T.; Matsutani, T.; Hikita, I.; Suzuki, M.; Oshima, I.; Tsukahara, K.; Arimura, A.; Horikawa, T.; Hirasawa, T.; et al. Association of a mutation in TRPV3 with defective hair growth in rodents. J. Investig. Dermatol. 2006, 126, 2664–2672. [Google Scholar] [CrossRef] [PubMed]
- Lonsdale, J.; Thomas, J.; Salvatore, M.; Phillips, R.; Lo, E.; Shad, S.; Hasz, R.; Walters, G.; Garcia, F.; Young, N.; et al. The genotype-tissue expression (GTEx) project. Nat. Genet. 2013, 45, 580–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aijima, R.; Wang, B.; Takao, T.; Mihara, H.; Kashio, M.; Ohsaki, Y.; Zhang, J.-Q.; Mizuno, A.; Suzuki, M.; Yamashita, Y.; et al. The thermosensitive TRPV3 channel contributes to rapid wound healing in oral epithelia. FASEB J. 2015, 29, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Ueda, T.; Yamada, T.; Ugawa, S.; Ishida, Y.; Shimada, S. TRPV3, a thermosensitive channel is expressed in mouse distal colon epithelium. Biochem. Biophys. Res. Commun. 2009, 383, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Ueda, T.; Ugawa, S.; Ishida, Y.; Imayasu, M.; Koyama, S.; Shimada, S. Functional expression of transient receptor potential vanilloid 3 (TRPV3) in corneal epithelial cells: involvement in thermosensation and wound healing. Exp. Eye Res. 2010, 90, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.E.; Chirila, A.M.; Schrank, B.R.; Kauer, J.A. Loss of interneuron LTD and attenuated pyramidal cell LTP in TRPV1 and TRPV3 KO mice. Hippocampus 2013, 23, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Moussaieff, A.; Rimmerman, N.; Bregman, T.; Straiker, A.; Felder, C.C.; Shoham, S.; Kashman, Y.; Huang, S.M.; Lee, H.; Shohami, E.; et al. Incensole acetate, an incense component, elicits psychoactivity by activating TRPV3 channels in the brain. FASEB J. 2008, 22, 3024–3034. [Google Scholar] [CrossRef] [PubMed]
- Gallego-Sandín, S.; Rodríguez-García, A.; Alonso, M.T.; García-Sancho, J. The endoplasmic reticulum of dorsal root ganglion neurons contains functional TRPV1 channels. J. Biol. Chem. 2009, 284, 32591–32601. [Google Scholar] [CrossRef] [PubMed]
- Lotteau, S.; Ducreux, S.; Romestaing, C.; Legrand, C.; Van Coppenolle, F. Characterization of Functional TRPV1 Channels in the Sarcoplasmic Reticulum of Mouse Skeletal Muscle. PLoS ONE 2013, 8, e58673. [Google Scholar] [CrossRef] [PubMed]
- Lo, I.C.; Chan, H.C.; Qi, Z.; Ng, K.L.; So, C.; Tsang, S.Y. TRPV3 channel negatively regulates cell cycle progression and safeguards the pluripotency of embryonic stem cells. J. Cell. Physiol. 2016, 231, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Guatteo, E.; Chung, K.K.H.; Bowala, T.K.; Bernardi, G.; Mercuri, N.B.; Lipski, J. Temperature sensitivity of dopaminergic neurons of the substantia nigra pars compacta: involvement of transient receptor potential channels. J. Neurophysiol. 2005, 94, 3069–3080. [Google Scholar] [CrossRef] [PubMed]
- Singh, U.; Kumar, S.; Shelkar, G.P.; Yadhav, M.; Kokare, D.M.; Goswami, C.; Lechan, R.M.; Singru, P.S. Transient receptor potential vanilloid (TRPV3) in the ventral tegmental area of rat: Role in modulation of the mesolimbic-dopamine reward pathway. Neuropharmacology 2016, 110, 198–210. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.-K.; Lee, H.; Mizuno, A.; Suzuki, M.; Caterina, M.J. 2-aminoethoxydiphenyl borate activates and sensitizes the heat-gated ion channel TRPV3. J. Neurosci. 2004, 24, 5177–5182. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.-K.; Lee, H.; Mizuno, A.; Suzuki, M.; Caterina, M.J. TRPV3 and TRPV4 mediate warmth-evoked currents in primary mouse keratinocytes. J. Biol. Chem. 2004, 279, 21569–21575. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Jin, J.; Hu, L.; Shen, D.; Dong, X.-P.; Samie, M.A.; Knoff, J.; Eisinger, B.; Liu, M.-L.; Huang, S.M.; et al. TRP channel regulates EGFR signaling in hair morphogenesis and skin barrier formation. Cell 2010, 141, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T.; Petrus, M.J.; Dubin, A.E.; Patapoutian, A. TRPV3 regulates nitric oxide synthase-independent nitric oxide synthesis in the skin. Nat. Commun. 2011, 2, 369. [Google Scholar] [CrossRef] [PubMed]
- Borbíró, I.; Lisztes, E.; Tóth, B.I.; Czifra, G.; Oláh, A.; Szöllosi, A.G.; Szentandrássy, N.; Nánási, P.P.; Péter, Z.; Paus, R.; et al. Activation of transient receptor potential vanilloid-3 inhibits human hair growth. J. Investig. Dermatol. 2011, 131, 1605–1614. [Google Scholar] [CrossRef] [PubMed]
- Mandadi, S.; Sokabe, T.; Shibasaki, K.; Katanosaka, K.; Mizuno, A.; Moqrich, A.; Patapoutian, A.; Fukumi-Tominaga, T.; Mizumura, K.; Tominaga, M. TRPV3 in keratinocytes transmits temperature information to sensory neurons via ATP. Pflugers Arch. Eur. J. Physiol. 2009, 458, 1093–1102. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Sakamoto, T.; Tiwari, V.; Kim, Y.-S.; Yang, F.; Dong, X.; Güler, A.D.; Guan, Y.; Caterina, M.J. Selective keratinocyte stimulation is sufficient to evoke nociception in mice. Pain 2015, 156, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Radtke, C.; Vogt, P.M.; Devor, M.; Kocsis, J.D. Keratinocytes acting on injured afferents induce extreme neuronal hyperexcitability and chronic pain. Pain 2010, 148, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.M.; Li, X.; Yu, Y.; Wang, J.; Caterina, M.J. TRPV3 and TRPV4 ion channels are not major contributors to mouse heat sensation. Mol. Pain 2011, 7, 37. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B.; Bíró, T.; Owsianik, G. TRPV3: Time to decipher a poorly understood family member! J. Physiol. 2014, 592, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.S.; Cho, S.I.; Choi, M.G.; Choi, Y.H.; Kwak, I.S.; Park, C.W.; Kim, H.O. Increased expression of three types of transient receptor potential channels (TRPA1, TRPV4 and TRPV3) in burn scars with post-burn pruritus. Acta Derm. Venereol. 2015, 95, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Sulk, M.; Seeliger, S.; Aubert, J.; Schwab, V.D.; Cevikbas, F.; Rivier, M.; Nowak, P.; Voegel, J.J.; Buddenkotte, J.; Steinhoff, M. Distribution and expression of non-neuronal transient receptor potential (TRPV) ion channels in rosacea. J. Investig. Dermatol. 2012, 132, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Scott, V.E.; Patel, H.; Wetter, J.; Edlmayer, R.; Neelands, T.; Miller, L.; Huang, S.; Gauld, S.; Todorovic, V.; Gomtsian, A.; et al. Defining a mechanistic link between TRPV3 activity and psoriasis through IL-1α and EGFR signaling pathways. J. Investig. Dermatol. 2016, 136, S94. [Google Scholar] [CrossRef]
- Gopinath, P.; Wan, E.; Holdcroft, A.; Facer, P.; Davis, J.B.; Smith, G.D.; Bountra, C.; Anand, P. Increased capsaicin receptor TRPV1 in skin nerve fibres and related vanilloid receptors TRPV3 and TRPV4 in keratinocytes in human breast pain. BMC Women’s Health 2005, 5, 2. [Google Scholar] [CrossRef] [PubMed]
- Brash, A.R. Arachidonic acid as a bioactive molecule. J. Clin. Investig. 2001, 107, 1339–1345. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, T.; Imura, K.; Asakawa, M.; Suzuki, M.; Oshima, I.; Hirasawa, T.; Sakata, T.; Horikawa, T.; Arimura, A. Impact of the Gly573Ser substitution in TRPV3 on the development of allergic and pruritic dermatitis in mice. J. Investig. Dermatol. 2009, 129, 714–722. [Google Scholar] [CrossRef] [PubMed]
- Xiao, R.; Tian, J.; Tang, J.; Zhu, M.X. The TRPV3 mutation associated with the hairless phenotype in rodents is constitutively active. Cell Calcium 2008, 43, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, M.; Sakai, A.; Suzuki, H.; Akimoto, T. Analysis of TRPV3 function in pain sensation using a gain-of-function mutant rat. Exp. Anim. 2015, 64, S69–S135. [Google Scholar]
- Duchatelet, S.; Hovnanian, A. Olmsted syndrome: clinical, molecular and therapeutic aspects. Orphanet. J. Rare Dis. 2015, 10, 33. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B.; Biro, T. TRPV3: A “more than skinny” channel. Exp. Dermatol. 2013, 22, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Moran, M.M.; Chong, J.; Fanger, C.A.; Ripka, A.; Larsen, G.R.; Zhen, X.; Underwood, D.J.; Weigele, M. Compounds for Modulating TRPV3 Function. WO2008033564A1, 2008. [Google Scholar]
- Reilly, R.M.; Kym, P.R. Analgesic potential of TRPV3 antagonists. Curr. Top. Med. Chem. 2011, 11, 2210–2215. [Google Scholar] [CrossRef] [PubMed]
- Joshi, N. The TRPV3 Receptor as a Pain Target: A therapeutic promise or just some more new biology? Open Drug Discov. J. 2010, 2, 89–97. [Google Scholar] [CrossRef]
- Huang, S.M.; Chung, M.-K. Targeting TRPV3 for the Development of Novel Analgesics. Open Pain J. 2013, 6, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Gomtsyan, A.R.; Schmidt, R.G.; Bayburt, E.K.; Gfesser, G.A.; Voight, E.A.; Daanen, J.F.; Schmidt, D.L.; Cowart, M.D.; Liu, H.; Altenbach, R.J.; et al. Synthesis and pharmacology of (pyridin-2-yl)methanol derivatives as novel and selective transient receptor potential vanilloid 3 antagonists. J. Med. Chem. 2016, 59, 4926–4947. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.A.; Fanger, C.M.; Larsen, G.R.; Lumma, W.C.; Moran, M.M.; Ripka, A.; Underwood, D.J.; Weigele, M.; Zhen, X. Compounds for Modulating TRPV3 Function. US20060270688A1, 2006. [Google Scholar]
- Lin, Z.; Chen, Q.; Lee, M.; Cao, X.; Zhang, J.; Ma, D.; Chen, L.; Hu, X.; Wang, H.; Wang, X.; et al. Exome sequencing reveals mutations in TRPV3 as a cause of Olmsted syndrome. Am. J. Hum. Genet. 2012, 90, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Lai-Cheong, J.E.; Sethuraman, G.; Ramam, M.; Stone, K.; Simpson, M.A.; McGrath, J.A. Recurrent heterozygous missense mutation, p.Gly573Ser, in the TRPV3 gene in an Indian boy with sporadic Olmsted syndrome. Br. J. Dermatol. 2012, 167, 440–442. [Google Scholar] [CrossRef] [PubMed]
- Eytan, O.; Fuchs-Telem, D.; Mevorach, B.; Indelman, M.; Bergman, R.; Sarig, O.; Goldberg, I.; Adir, N.; Sprecher, E. Olmsted syndrome caused by a homozygous recessive mutation in TRPV3. J. Investig. Dermatol. 2014, 134, 1752–1754. [Google Scholar] [CrossRef] [PubMed]
- Ni, C.; Yan, M.; Zhang, J.; Cheng, R.; Liang, J.; Deng, D.; Wang, Z.; Li, M.; Yao, Z. A novel mutation in TRPV3 gene causes atypical familial Olmsted syndrome. Sci. Rep. 2016, 6, 21815. [Google Scholar] [CrossRef] [PubMed]
- Kariminejad, A.; Barzegar, M.; Abdollahimajd, F.; Pramanik, R.; McGrath, J.A. Olmsted syndrome in an Iranian boy with a new de novo mutation in TRPV3. Clin. Exp. Dermatol. 2014, 39, 492–495. [Google Scholar] [CrossRef] [PubMed]
- Duchatelet, S.; Guibbal, L.; de Veer, S.; Fraitag, S.; Nitschke, P.; Zarhrate, M.; Bodemer, C.; Hovnanian, A. Olmsted syndrome with erythromelalgia caused by recessive transient receptor potential vanilloid 3 mutations. Br. J. Dermatol. 2014, 171, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Duchatelet, S.; Pruvost, S.; de Veer, S.; Fraitag, S.; Nitschke, P.; Bole-Feysot, C.; Bodemer, C.; Hovnanian, A. A new TRPV3 missense mutation in a patient with Olmsted syndrome and erythromelalgia. JAMA Dermatol. 2014, 150, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto-Kasai, E.; Yasui, K.; Shichijo, M.; Sakata, T.; Yoshioka, T. Impact of TRPV3 on the development of allergic dermatitis as a dendritic cell modulator. Exp. Dermatol. 2013, 22, 820–824. [Google Scholar] [CrossRef] [PubMed]
- Lipski, J.; Park, T.I.H.; Li, D.; Lee, S.C.W.; Trevarton, A.J.; Chung, K.K.H.; Freestone, P.S.; Bai, J.-Z. Involvement of TRP-like channels in the acute ischemic response of hippocampal CA1 neurons in brain slices. Brain Res. 2006, 1077, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Moussaieff, A.; Yu, J.; Zhu, H.; Gattoni-Celli, S.; Shohami, E.; Kindy, M.S. Protective effects of incensole acetate on cerebral ischemic injury. Brain Res. 2012, 1443, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Facer, P.; Casula, M.A.; Smith, G.D.; Benham, C.D.; Chessell, I.P.; Bountra, C.; Sinisi, M.; Birch, R.; Anand, P. Differential expression of the capsaicin receptor TRPV1 and related novel receptors TRPV3, TRPV4 and TRPM8 in normal human tissues and changes in traumatic and diabetic neuropathy. BMC Neurol. 2007, 7, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lingam, V.S.P.; Chaudhari, S.S.; Thomas, A.; Khairatkar-Joshi, N.; Kattige, V.G. Fused Pyrimidineone Compounds as TRPV3 Modulators. U.S. Patent 20090286811, 19 November 2009. [Google Scholar]
- Lingam, V.S.P.; Thomas, A.; Phatangare, S.K.; Mindhe, A.S.; Khatik, J.Y.; Khairatkar-Joshi, N.; Kattige, V.G. Fused Imidazole Derivatives as TRPV3 Antagonist. U.S. Patent 20110257193, 20 October 2011. [Google Scholar]
- Lingam, V.S.P.; Chaudhari, S.S.; Thomas, A.; Khairatkar-Joshi, N.; Kattige, V.G. Fused Pyrimidineone Compounds as TRPV3 Modulators. U.S. Patent 20120115886, 10 May 2012. [Google Scholar]
- Lingam, V.S.P.; Thomas, A.; Dattaguru, A.M.; Khatik, J.Y.; Khairatkar-Joshi, N.; Kattige, V.G. Fused Pyramine Derivatives As TRPV3 Modulators. U.S. Patent 20100292254, 18 November 2010. [Google Scholar]
- Lingam, V.S.P.; Thomas, A.; Gharat, L.A.; Ukirde, D.V.; Phatangare, S.K.; Mindhe, A.S.; Khairatkar-Joshi, N.; Kattige, V.G. Chromane Derivatives As TRPV3 Modulators. U.S. Patent 20100311778, 9 December 2010. [Google Scholar]
- Bayburt, E.K.; Clapham, B.; Cox, P.B.; Daanen, J.F.; Gomtsyan, A.R.; Kort, M.E.; Kym, P.R.; Voight, E.A. Novel TRPV3 Modulators. U.S. Patent 20130150409, 13 June 2013. [Google Scholar]
- Bayburt, E.K.; Clapham, B.; Cox, P.B.; Daanen, J.F.; Dart, M.J.; Gfesser, G.A.; Gomtsyan, A.R.; Kort, M.E.; Kym, P.R.; Schmidt, R.G.; et al. Novel TRPV3 Modulators. U.S. Patent 20130131036, 23 May 2009. [Google Scholar]
- Moussaieff, A.; Shohami, E.; Kashman, Y.; Fride, E.; Schmitz, M.L.; Renner, F.; Fiebich, B.L.; Munoz, E.; Ben-Neriah, Y.; Mechoulam, R. Incensole acetate, a novel anti-inflammatory compound isolated from Boswellia resin, inhibits nuclear factor-kappa B activation. Mol. Pharmacol. 2007, 72, 1657–1664. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.M. TRPV3 Agonists for the Treatment of Skin Conditions. U.S. Patent 20150250699, 10 September 2015. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Potential Roles | Summary of Evidence | Reference(s) |
|---|---|---|
| Olmsted Syndrome | Several independent clinical reports identify mutations in the TRPV3 gene as a cause of gain-of-function mutations and recessive Olmsted’s Syndrome. Characteristic features include palmoplantar keratoderma, periorificial hyperkeratotic lesions and alopecia. Less common presentations include digit constriction, onychodystrophy and pruritus | [66,69,70,71,72,73] |
| Olmsted Syndrome with Erythromelalgia | Clinical data of three patients whose disease presentation included intense flares of inflammation, itching, burning pain, vasodilatation, and redness of the extremities consistent with erythromelalgia. Whole exome sequencing identified a de novo heterozygous missense mutation within TRPV3, p.Leu673Phe. | [74,75] |
| Pruitic and Atopic Dermatitis | Clinical data suggest TRPV3 expression is increased in lesional skin in patients with atopic dermatitis. Preclinical data suggest DS-Nh mice develop allergic and pruritic dermatitis | [58,76] |
| Psoriasis | Clinical data suggest TRPV3 expression is significantly increased in psoriatic lesions, and that these channels are functional in keratinocytes isolated from lesioned skin. A novel antagonist of TRPV3 dose-dependently inhibited 2APB/carvacrol induced IL-1α release keratinocytes. Similarly, inhibition of EGFR signaling was observed with the antagonist. Inhibitors of both IL-1α release and EGFR signaling have previously attenuated psoriatic symptoms and thus a linkage with TRPV3 is suggested | [55] |
| Wound Healing | Pre-clinical data suggest higher expression of TRPV3 in mouse oral epithelia versus skin, and expression was upregulated in wounded oral epithelial tissue. TRPV3 activation promoted oral epithelial cell proliferation, which was diminished in TRPV3 knockout mice. Subsequent knock out profiling in a molar tooth extraction model suggest oral wound closure was delayed | [33] |
| Burn/Post-burn pruritus | Clinical data suggest increased TRPV3 expression in the epidermis of burn scars with pruritus | [29,53] |
| Hair growth | Pre-clinical data suggest TRPV3 agonists eugenol and 2-aminoethoxydiphenyl borate inhibited hair shaft elongation, suppressed proliferation, and induced apoptosis in human organ-cultured hair follicles. Similarly, functional effects of TRPV3 activation in human ORS keratinocytes were demonstrated as on-target via siRNA | [47] |
| Skin Barrier Formation | Pre-clinical data suggest TRPV3 forms a direct complex with transglutaminases, thereby regulating growth factor signaling for the formation of the skin barrier | [45] |
| Rosacea | Clinical data suggest increased TRPV3 expression in epidermal keratinocytes, and dermal labeling was observed in a subset of immune cells and fibroblasts in erythematotelangiectatic rosacea and phymatous rosacea-affected skin. Increased gene expression was also observed in patients with phymatous rosacea | [54] |
| Cerebral Ischemia | Pre-clinical data suggest the TRPV3 agonist incensole acetate protects against ischemic neuronal damage and reperfusion injury in mice. Reduced infarct volumes, inhibition of TNF-α, IL-1β and TGF-β expression, and NF-κB activation were demonstrated as on-target using TRPV3 knock-out mice | [77,78] |
| Mastalgia | Clinical data suggest increased expression of TRPV3 in basal keratinocytes that correlated with disease score | [56] |
| Traumatic Peripheral Nerve Injury | Clinical data suggest increased TRPV3 expression in the DRG neurons of patients with DRG avulsion injury, in the peripheral nerve proximal to the site of brachial plexus injury. However, a decrease in TRPV3 expression was observed in the skin of patients with diabetic neuropathy | [3,79] |
| Cold- and Heat-evoked Pain | Pre-clinical data showed TRPV3 knockout mice have impaired responses to noxious heat. WBN/Kob-Ht rats, which have a TRPV3 gain-of-function mutation, showed an increased sensitivity to noxious heat and cold stimuli. Multiple antagonists have shown efficacy in inflammatory insult-induced hypersensitivity and nerve ligation-induced hypersensitivity | [45,60,64,65] |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Broad, L.M.; Mogg, A.J.; Eberle, E.; Tolley, M.; Li, D.L.; Knopp, K.L. TRPV3 in Drug Development. Pharmaceuticals 2016, 9, 55. https://doi.org/10.3390/ph9030055
Broad LM, Mogg AJ, Eberle E, Tolley M, Li DL, Knopp KL. TRPV3 in Drug Development. Pharmaceuticals. 2016; 9(3):55. https://doi.org/10.3390/ph9030055
Chicago/Turabian StyleBroad, Lisa M., Adrian J. Mogg, Elizabeth Eberle, Marcia Tolley, Dominic L. Li, and Kelly L. Knopp. 2016. "TRPV3 in Drug Development" Pharmaceuticals 9, no. 3: 55. https://doi.org/10.3390/ph9030055