
Metabolomics as a Prospective Tool for Soybean (Glycine max) Crop Improvement


Department of Agriculture and Animal Health, College of Agriculture and Environmental Sciences, University of South Africa, Science Campus, Private Bag x 6, Florida, Johannesburg 1710, South Africa
*
Author to whom correspondence should be addressed.
Curr. Issues Mol. Biol. 2022, 44(9), 4181-4196; https://doi.org/10.3390/cimb44090287
Submission received: 24 July 2022
/
Revised: 28 August 2022
/
Accepted: 7 September 2022
/
Published: 12 September 2022
(This article belongs to the Special Issue Advanced Research in Plant Metabolomics)
Abstract
:Global demand for soybean and its products has stimulated research into the production of novel genotypes with higher yields, greater drought and disease tolerance, and shorter growth times. Genetic research may be the most effective way to continue developing high-performing cultivars with desirable agronomic features and improved nutritional content and seed performance. Metabolomics, which predicts the metabolic marker for plant performance under stressful conditions, is rapidly gaining interest in plant breeding and has emerged as a powerful tool for driving crop improvement. The development of increasingly sensitive, automated, and high-throughput analytical technologies, paired with improved bioinformatics and other omics techniques, has paved the way for wide characterization of genetic characteristics for crop improvement. The combination of chromatography (liquid and gas-based) with mass spectrometry has also proven to be an indisputable efficient platform for metabolomic studies, notably plant metabolic fingerprinting investigations. Nevertheless, there has been significant progress in the use of nuclear magnetic resonance (NMR), capillary electrophoresis, and Fourier-transform infrared spectroscopy (FTIR), each with its own set of benefits and drawbacks. Furthermore, utilizing multivariate analysis, principal components analysis (PCA), discriminant analysis, and projection to latent structures (PLS), it is possible to identify and differentiate various groups. The researched soybean varieties may be correctly classified by using the PCA and PLS multivariate analyses. As metabolomics is an effective method for evaluating and selecting wild specimens with desirable features for the breeding of improved new cultivars, plant breeders can benefit from the identification of metabolite biomarkers and key metabolic pathways to develop new genotypes with value-added features.
1. Introduction
For centuries, mankind is entirely reliant on plants as the main source of nutrients. However, since the world population is increasing at a rapid rate, there is extreme pressure on the harvesting of health and nutritional contributing plants. Therefore, developing and implementing ways to reduce the impact of biotic and abiotic stresses on soybean yield and quality is critical for global food security [1,2].
Soja is a phylogenetic group that comprises wild soybean (Glycine soja), semi-wild soybean (Glycine gracilis), and cultivated soybean (Glycine max). Research has revealed that soybeans grown in the wild can better adapt to a variety of harsh conditions. Semi-wild soybean is a transition type in the Soja evolution, with a physiological metabolism similar to wild soybean and a phenotypic similar to cultivated soybean. Artificial selection and domestication have bred the cultivated soybean from wild soybean with origins from Asia [3].
Soybean, first grown in East Asia millennia years ago, is a vital source of nourishment for people all over the world, and it is widely regarded as a nutritious meal in many Asian countries. Humans have been growing and consuming soybeans for over 5000 years, while soybean oil has only recently become a significant element of our diet [4]. Soybeans are the most important legume and the fourth most important crop in terms of worldwide crop production, after rice, wheat, and maize. Soybeans biosynthesize a range of metabolites that are fundamental in crop yield and abiotic and biotic stress tolerance, disease resistance, seed composition, and flavor enrichment. Owing to the rising demand for soybeans, more output is currently required. Although in 2016, around 340 million metric tons of soybeans were grown globally, the production is influenced by a number of factors, including the availability of macro- and micronutrients as well as temperature of the soil [5]. However, it may become more difficult to obtain sufficient crop yields as the climate continues to change dramatically and soil environments are becoming increasingly more stressful to soybeans. To increase soybean harvest volumes mounting factors, ongoing concerns of environmental pressures such as extreme temperatures, salinity, flooding/drought stress, herbicide induction and the devastating effects of several pathogens, such as bacteria, mold fungi, nematodes, and insects on critical yield loss must be addressed [1,2].
Omics-based interdisciplinary approaches facilitate trait modification/optimization, resulting in optimal and precise design breeding [6]. Here we review a brief overview of the application of metabolomics technologies in crop improvement through genetic modification, their potential for future development, and the consequent assessment of food safety. Razzaq and colleagues (2022) describe the present utilization of advanced metabolomics methods coupled with other OMICS approaches that may be used to: examine the complexities of plant genotype-metabolite-phenotype interactions, facilitating metabolomics-assisted plant breeding for exploring the stress-responsive metabolic markers, uncover the hidden metabolic networks associated with abiotic/biotic stress resistance, and facilitate screening and selection of climate-smart crops at the metabolite level [7]. The fundamental idea underlying metabolic editing is to initially identify the precise genes responsible for key metabolic pathways, then to alter one or more genes associated to those networks.
Food safety is one of the main objections to genetically modified (GM) crops, however these objections should be dispelled by employing the present set of metabolomic technologies as part of a food safety evaluation approach and by using reasonable comparators [8]. Clarke and colleagues (2013) highlight the significance of metabolomics in the safety evaluation of GM crops. One of the world’s most extensive GM crops is a glyphosate-tolerant GM soybean type [9]. Garca-Villalba et al. [9] conducted the initial study on the considerable equivalency of GM soybean using a metabolomic method. Glyphosate binds to and inhibits the activity of EPSPS, an enzyme of the aromatic amino acid biosynthesis pathway (shikimate pathway). The inhibition of EPSPS by glyphosate prevents the plant from synthesizing the aromatic amino acids (phenylalanine, tyrosine, tryptophan) required for protein synthesis. However, certain microorganisms possess a glyphosate-resistant form of 5-enolpyruvoylshikimate-3-phosphate synthetase. The variant utilized in genetically modified crops is often obtained from glyphosate-resistant Agrobacterium tumefaciens strain CP4 (CP4 EPSPS). The ability to spray glyphosate on fields without affecting the crop significantly increased the ability to manage weeds in the field and ultimately increase the soybean yields [10] A separate study by Alberto and colleagues (2012) discovered that amino acid profiles could be used to examine how glyphosate affected both susceptible and resistant soybean lines. HPLC profiles for ten amino acids (Asp, Asn, Gln, Glu, Gly, His, Leu Ser, Thr, Tyr,) were compared in two near isogenic pairs in four varieties of soybean roots. Multivariate analysis utilizing principal component analysis (2D PCA and 3D PCA) enabled various groups to be identified and differentiated based on the genetic origin of the soybean, indicating the amino acid responses on susceptible and resistant types [11]. The resultant GMO soybean variety A3244, is renowned for its exceptional agronomic traits, including several biotic an abiotic-resistance and high yielding property [12]. This classic example highlights the potential of metabolomics-guided breeding in soybean improvement.
Relevance as a Multifunctional Crop
Soybean is the most important worldwide legume crop species worldwide owing to its agro-economic and nutritional value, serving as an essential source of protein and oils (40% and 21% content, respectively) for human consumption, livestock feed, industrial biofuel production, and functional foods [2,13,14]. Animal feed accounts for over 85 percent of global soybean protein meal production whereas the soybean crop is mostly farmed for oil production, with only a small percentage of soybeans consumed directly by humans. Owing to its high oil and protein content, soybean is among the world’s most essential crops as it contributes to 56% of all oilseed production worldwide. Soybeans are also high in vitamins, minerals, phospholipids, saponins, isoflavones, flavonoids, oligosaccharides, edible fiber, free sugars, pterocarpans, phytic acids, peptides, and antioxidant compounds [2,15,16,17]. Although phytochemicals in soybeans are present in small amounts and are not required for normal body function, they confer health benefits and aid in the treatment of a variety of diseases, including cancer, arteriosclerosis, osteoporosis, and metabolic syndrome [2,18,19].
Soybean oil is being developed and marketed as a future fuel source, with attempts being undertaken to enhance soybean-derived biodiesel output. In addition, there is ongoing research where soybean protein-based biodegradable materials are being explored to determine the potential as an alternative for plastic synthesis [13]. In addition, owing to the presence of these phytochemicals such as polyphenols and essential oils, soybean leaves have recently been employed in the cosmetics industry and food products [20]. As a result of soybean–Bradyrhizobium symbiosis, soybean can meet 50–60% of its nitrogen demand, therefore significantly contributing to soil fertility improvement through biological nitrogen fixation [17,21]. Soymeal, the residue remaining after oil extraction, is a key metabolizable energy source and the world’s number one protein source for animal feed. Soybeans are also utilized for the production of adhesives, inks, building materials, and lubricants [22].
Soybean is therefore a valuable crop for agriculture, industry, and food and thus becoming a more common crop species attributing to its diverse uses, and high demand. By 2050, the world’s population will have doubled, necessitating double the current food output; whereas worldwide soybean production is much below what is required [5]. As a result, the agricultural biotechnology community is placing emphasis on the modification of seed-specific output traits of soybeans [23]. The study of functional genomics has had a significant impact in this regard, providing large-scale biological data that can be used to determine how specific processes in an organism are regulated and controlled, a branch of molecular biology (i.e., metabolomics) that utilizes the huge amount of genomic data available to determine gene functions and interactions [22].
2. Metabolomics at the Forefront of Functional Genomic Approaches
Metabolomics refers to a comprehensive modern “omic” approach for analyzing metabolites in a biological system under a specific physiological condition [13,24,25,26,27]. According to the central dogma model, biological information is sequentially transmitted respectively from the genome, transcriptome, proteome, and metabolome (Figure 1). The biochemical phenotype of an organism is represented by the metabolome detail and, thus, a metabolomic investigation unravels the links from the genotype to the phenotype [28,29,30].
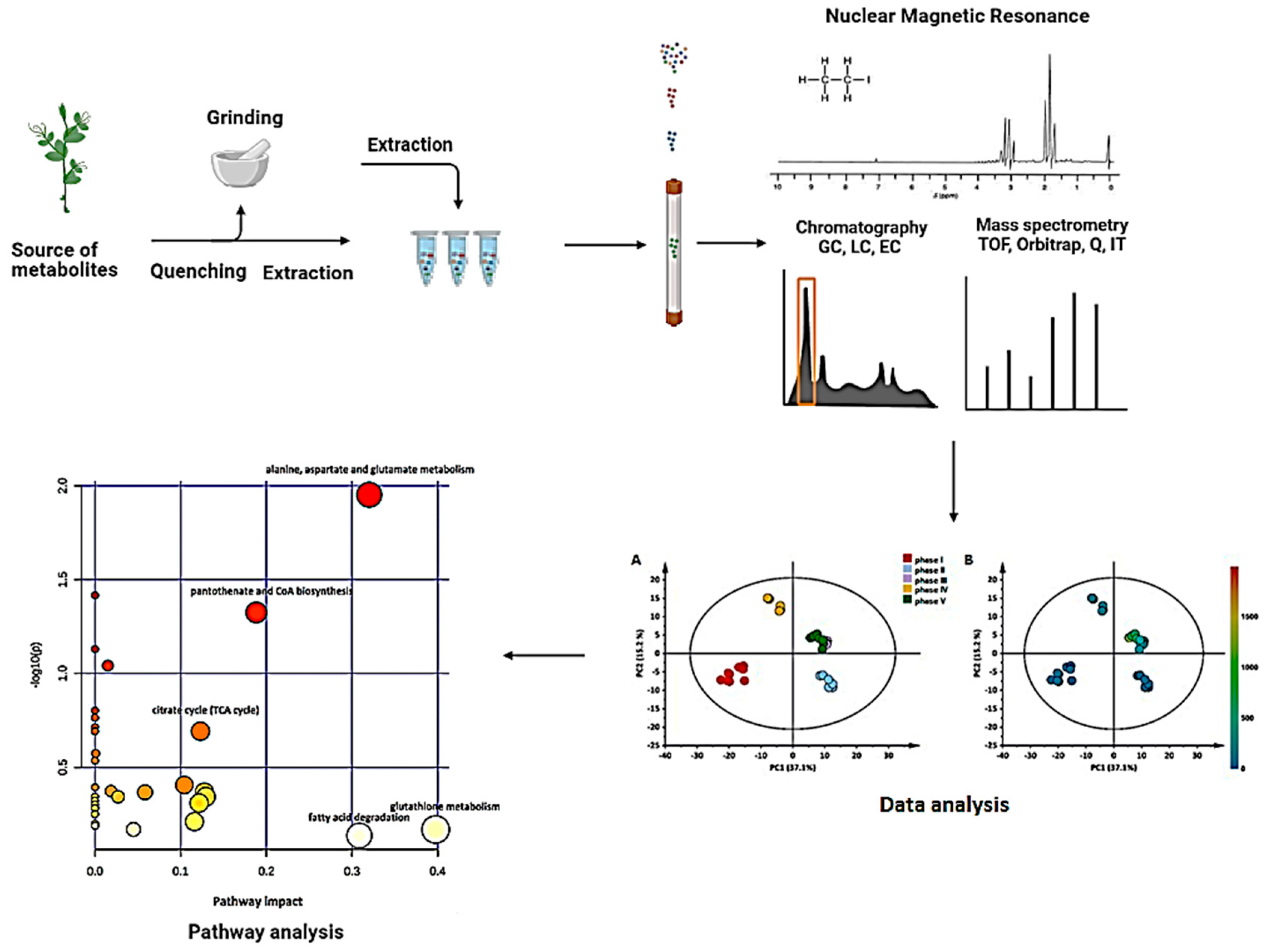
Metabolomics is a multidisciplinary field that includes biology, analytical chemistry, and multivariate statistics. Three main steps are involved in a metabolomics study: sample preparation, data acquisition, and analysis [31] as illustrated in Figure 2.
2.1. Sample Preparation
Sample preparation is the most basic and crucial stage for all plant molecular biological studies since sample integrity dictates the entire outcome of the experiment, i.e., the acquired data and the subsequent biological interpretation. It is critical to keep experimental and biological variance to a minimum to ensure the metabolomic analyses are consistent, robust, and valid [32,33,34]. However, a metabolomic approach generally requires minimal sample preparation relative to the other genomic approaches such as genomics, transcriptomics, proteomics, which are labor intensive [35,36]. However, a diverse set of metabolites with varying physio-chemical complexity and relative abundance poses numerous challenges in plant metabolism [37]. The most common extraction method includes liquid–liquid extraction (LLE), solid-phase extraction (SPE), supercritical fluid extraction (SFE), and microwave aided extraction (MAE) [38,39,40,41,42,43,44,45].
Factors to consider when choosing an extraction technique include selectivity to the widest range of metabolites possible and reproducibility. Regardless of their development, none of these methods can extract the whole metabolome from a biological sample. Each method comes with a built-in bias in favor of a particular class of chemical [37].
2.2. Data Acquisition
There has been significant progress in the development of new technologies for metabolomic platforms, which has resulted in the creation of additional data [46]. Several effective analytical platforms are constantly developed and modified in an attempt to comprehensively include as many secondary metabolites as possible. Thus, in plant metabolomics, chromatography and mass spectrometry are the most common techniques used. The invention of ultra-high performance liquid chromatography (UHPLC) heralded the beginning of innovation in LC-based metabolomic platforms by addressing the problem of poor resolution of data gathered using high-performance liquid chromatography (HPLC). However, substantial advancements have been witnessed in the application of Fourier-transform infrared spectroscopy (FTIR), capillary electrophoresis (CE), and nuclear magnetic resonance (NMR) each with its own set of advantages and disadvantages [2,47,48].
The hyphenation of chromatography (liquid and gas-based) and mass spectrometry has proven to be an indisputably efficient platform utilized in metabolomic studies, particularly plant metabolic fingerprinting investigations because a single run can gather data from two functions [49]. The advantages of mass spectrometry in metabolomic analysis are high sensitivity, repeatability, and adaptability. This platform generates an ion by removing or adding a charge from a neutral species, then measures the m/z (mass–to–charge ratio) of the ions to provide structural information based on the fragmentation pattern obtained. Prior to entering the mass spectrometer, chromatographic separation of unprocessed biological materials further assists metabolite annotation by adding the retention time (Rt) identifier, increases sensitivity, and decreases signal suppression [50,51].
NMR is a spectroscopic technique that makes use of an atom’s spin characteristics to identify and quantify elements. The method is robust and highly selective, albeit limited by low sensitivity. On the other hand, NMR is unrivaled in the annotation of metabolites due to its capacity to provide the structural intricate details about a molecule. An added advantage of NMR is the capacity to provide semi-quantitative information, as the intensity of the acquired signal is directly proportional to the number of nuclear spins [52,53]. To cover a greater spectrum of metabolites, a full global investigation of an organism’s metabolome frequently necessitates the use of parallel analytical platforms [54].
2.3. Data Analysis
Although great strides have been made in the optimization of analytical platforms for data acquisition in metabolomic applications, each platform still has limits. As a result, various steps are conducted post- raw data collection to facilitate metabolite annotation. The initial step is visual data examination followed by data processing and metabolite annotation and, ultimately, biological interpretation [31].
2.3.1. Data Visualization (Pre-Processing and Pre-Treatment)
The visual inspection of graphical information is a vital stage in determining the quality of the raw data and selecting the most appropriate parameters for the successive steps of data processing workflow. However, high-throughput methods generate a large volume of raw data that is not feasible to analyze without automated information technology. As a result, managing these data sets holistically by hand is unfeasible. As a result, a variety of comprehensive software tools and mathematical algorithms for automatic raw data processing have been developed and are all capable of carrying out automated peak picking and other processing functions efficiently [9,55,56,57,58,59]. In this step, raw data are presented in the form of chromatograms and/or spectra as per the various data acquisition mode (Section 2.2).
In the literature, the terms pre-treatment and/or pre-processing are used interchangeably. Essentially, the ultimate purpose of these statistical steps is to eliminate all unrelated factors (experimental and/or analytical) while retaining useful biological information [55,60]. The most common and preferred pre-treatment and pre-processing methods are transformation [9,37], filtration and filling [37,61,62], spectral deconvolution [9,62,63], normalization [24,61], and peak alignment [9,37,61].
2.3.2. Statistical Modelling
The resulting high-throughput raw data matrix is exported to multiple software packages for robust statistical modeling utilizing univariate statistics and multivariate data analysis [9,55]. The most prominent are principal component analysis (PCA), hierarchical cluster analysis (HCA), and partial least squares regression (PLS). PCA is generally the basis for data analysis, wherein pattern recognition model enables the quick display of similarities and differences between sample groups by compressing the multidimensionality of data into a reduced number of variables known as principal components. PCA modeling is thus an unsupervised technique (i.e., without a priori class information) for investigating untargeted metabolic data because it accounts for the overall variance of the dataset provided without the requirement for a priori sample class information [9,64,65,66]. PLS, as a supervised classification model (i.e., with a priori class information), is beneficial when the unsupervised model does not capture the characteristic biomarkers that distinguish between different sample groups. The extension, orthogonal partial least squares/orthogonal projection to latent structures-discriminating analysis (OPLS-DA), explains just the reaction to biological variation, i.e., unique metabolite profiles significantly correlated to the specified response structure [55,66].
2.4. Metabolite Annotation, Pathway Mapping, Network Correlation and Biological Interpretation
In metabolomics, the assignment of appropriate metabolite annotation chemical formulas as well as metabolite annotations is a computationally and analytically challenging task. The lack of standardized experimental settings, as well as the biochemical diversity of metabolites, significantly add to the task’s complexity [67]. Significant progress has been made in the advancements of metabolite annotation databases and user-friendly software resources to overcome these constraints. In this regard, there are various free and well-developed software databases available that provide searches based on precise mass and chemical formulas [67,68,69,70,71,72]. However, in some circumstances, annotation of metabolites based only on precise mass and chemical formula may be insufficient. As a result, it is a critical step to back up computationally generated data with experimental evidence. The list of the tentatively identified metabolites are traditionally presented in the form of a table and/or chemical structures.
The biological interpretation of the overall findings is dependent on the correct assignment of the annotated metabolites’ biological roles. Network modeling and pathway mapping tools enable the comprehension of the biological interactions between metabolites. Accordingly, metabolite profiling enables the interpretation of interconnections that arise primarily through metabolic regulation [13].
3. Application of Metabolomics as a Prospective Tool to Improve Soybean
Metabolomics advancements enable scientists to rapidly map individual metabolites to the genes that encode their metabolic pathways, providing plant scientists with an exceptional chance to thoroughly study and rationally utilize the plethora of metabolites that plants biosynthesize.
Metabolomics is an effective method for measuring biological or physiological reactions to environmental changes, particularly when combined with other profiling technologies such as transcriptomics and proteomics [21]. Although more robust when combined with other “omics” approaches, the knowledge obtained from metabolomics can contribute to the holistic biological profiling of an organism [13].
Metabolomics has a broad range of applications including the annotation of specific genes [29,65], unravelling metabolic pathways [73], evaluation of biomarker products resulting from transgene expression [65] and environmental perturbation in plants [29], clinical diagnostics of diseases, evaluation of environmental research, drug action research [37], plant taxonomic evaluation [29], biotechnological engineering, food nutritional science [37,73]. In the context of this review, metabolomics advancements enable scientists to rapidly map individual metabolites to the genes that encode their metabolic pathways, providing plant scientists with an exceptional chance to thoroughly study and rationally utilize the plethora of metabolites that plants biosynthesize [7].
As an example of the application of metabolomics, previous studies have reported the adaptive responses of soybean to biotic and abiotic stressors, as well as the major primary and secondary metabolites involved in the adaptation and sensing mechanisms as shown in Table 1.
Although significant literature on critical information about the specific metabolite alternations that occur in response to diverse stress circumstances has been documented (Table 1), plant response to biotic and abiotic stress is a complicated and dynamic process. Most of the current research focuses on a single abiotic stress, although in practice, multiple stresses are commonly present. The interaction of these pressures will influence the physiological response of plants. Furthermore, different organs or cultivars of soybean plants respond differently to the same stress, and the plant’s metabolic system is a constantly changing network of interconnections [2,96,97]. Nevertheless, the study of the soybean metabolome paves the way to a better understanding of complex metabolic pathways and stress-associated metabolites. Metabolomics research can, therefore, pave way for the identification of metabolites as biomarkers of various environmental stressors.
Metabolomics-guided plant breeding programs such as mGWAS (Metabolite Genome-Wide Association Study) and mQTL (methylation quantitative trait locus) mGWAS analysis has proven to be critical for dissecting the genetic and metabolic architecture of rice by finding the genes related with natural variation in rice metabolism [7,90,98,99]. Chen and colleagues (2014) used GWAS to detect 6.4 million SNPs from 529 distinct rice strains, and 36 potential genes that regulate the levels of at least 34 recognized primary and secondary metabolites were identified. Here, this technique is essential for performing molecular phenotypic trait mapping for the purpose of rice improvement. Using flow infusion high-resolution mass spectrometry (FIE-HRMS) [98], Yadav et al. (2021) investigated metabolomic fingerprinting of 197 pearl millet inbred lines and identified numerous metabolite characteristics linked to nutritional benefits such lipid metabolism, vitamins, antioxidants, and dietary starch [99]. The wealth of metabolomic -related research on soybean (Table 1) highlights the potential of such metabolomics-guided plant breeding programs to be utilized toward soybean crop improvement.
4. Concluding Remarks and Future Perspectives
Metabolomics, a new and developing field that can predict several biomarkers and characterize the molecular traits involved in physiological processes, is at the forefront of making significant advances in soybean functional genomics research-based methodologies. Thus, the present review provides information on the metabolomics workflow and highlights the prospects of metabolomics in determining key biomarkers associated with mitigating biotic and abiotic stresses to provide valuable information that will guide the soybean breeding programs to produce improved cultivars with value-added features. The principal idea of metabolic editing is to first discover the precise genes responsible for the important metabolic pathways, then change one or more genes connected with those networks. Here, the role of mGWAS in the contribution to the success of genetic modifications and analyses of biomarkers that result in improved soybean yield and stress tolerance has been discussed.
Generating metabolite databases for important crop species under environmental stresses is a time-consuming task. To address these drawbacks, enhancing the resolution and coverage of the metabolome can help to gain a comprehensive understanding of how soybean adapts to biotic and abiotic stress, opening new options for increasing crop yields. Furthermore, although the biochemical and molecular specifics of these pathways are still being worked out, in-depth insights are progressively being achieved through the advancement and development of systems biology strategies. This work provides useful information that may be used in potential metabolic engineering and molecular breeding efforts to improve soybean seed quality and yield in the future. Future investigations may focus on dissecting the metabolome of soybean seeds at different physiological stages, as well as linking the metabolic variations to genomic changes.
Author Contributions
Conceptualization, E.N., K.M. and N.N.; investigation, E.N.; resources, K.M. and N.N.; writing—original draft preparation, E.N.; writing—review and editing, K.M. and N.N.; supervision, K.M. and N.N. All authors have read and agreed to the published version of the manuscript.
Funding
E.N. received funding for the fellowship from the Department of Agriculture and Agriculture and Animal Health, University of South Africa.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Sato, D.; Akashi, H.; Sugimoto, M.; Tomita, M. Metabolomic profiling of the response of susceptible and resistant soybean strains to foxglove aphid, Aulacorthum solani Kaltenbach. J. Chromatogr. B 2013, 925, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Ding, C.; Li, W.; Wang, D.; Cui, D. Applications of Metabolomics in the Research of Soybean Plant under Abiotic Stress; Elsevier Ltd.: Amsterdam, The Netherlands, 2019; ISBN 8657188982820. [Google Scholar]
- Yang, D.; Zhang, J.; Ming-xia, L.; Shi, L. Metabolomics analysis reveals the salt-tolerant mechanism in Glycine soja. J. Plant Growth Regul. 2017, 36, 460–471. [Google Scholar] [CrossRef]
- Muhammad, H.; Sumera, A.K.; Abdul, L.K.; Jae-Ho, S.; Bashir, A.; Shin, D.-H.; Lee, I.-J. Exogenous gibberellic acid reprograms soybean to higher growth and salt stress tolerance. J. Agric. Food Chem. 2010, 58, 7226–7232. [Google Scholar] [CrossRef]
- Shinano, T.; Cheng, W.; Saito, K.; Oikawa, A. Metabolomic analysis of night-released soybean root exudates under high- and low-K conditions. Plant Soil 2020, 456, 259–276. [Google Scholar]
- Oliveira, M.C.; Osipitan, O.A.; Begcy, K.; Werle, R. Cover crops, hormones and herbicides: Priming an integrated weed management strategy. Plant Sci. 2020, 301, 110550. [Google Scholar] [CrossRef]
- Razzaq, A.; Wishart, D.S.; Wani, S.H.; Hameed, M.K.; Mubin, M.; Saleem, F. Advances in metabolomics-driven diagnostic breeding and crop improvement. Metabolites 2022, 12, 511. [Google Scholar] [CrossRef]
- Clarke, J.D.; Alexander, D.C.; Ward, D.P.; Ryals, J.A.; Mitchell, M.W.; Wulff, J.E.; Guo, L. Assessment of genetically modified soybean in relation to natural variation in the soybean seed metabolome. Sci. Rep. 2013, 3, 3082. [Google Scholar] [CrossRef]
- García-Villalba, R.; León, C.; Dinelli, G.; Segura-Carretero, A.; Fernández-Gutiérrez, A.; Garcia-Cañas, V.; Cifuentes, A. Comparative metabolomic study of transgenic versus conventional soybean using capillary electrophoresis-time-of-flight mass spectrometry. J. Chromatogr. A 2008, 1195, 164–173. [Google Scholar] [CrossRef]
- Padgette, S.R.; Re, D.B.; Barry, G.F.; Eichholtz, D.E.; Xavier, D.; Fuchs, R.L.; Kishore, G.M.; Fraley, R.T. New weed control opportunities: Development of soybeans with a Roundup Ready™ gene. In Herbicide–Resistant Crops; CRC Press: Boca Raton, FL, USA, 2018; pp. 53–84. [Google Scholar]
- Alberto, C.; Manuel, J.; Oliveira, L.; Mui, S.; Antunes, R. Physiological effects of glyphosate over amino acid profile in conventional and transgenic soybean (Glycine max). Pestic. Biochem. Physiol. 2012, 102, 134–141. [Google Scholar] [CrossRef]
- Lundry, D.R.; Ridley, W.P.; Meyer, J.J.; Riordan, S.G.; Nemeth, M.A.; Trujillo, W.A.; Breeze, M.L.; Sorbet, R. Composition of Grain, forage, and processed fractions from second-generation glyphosate-tolerant soybean, MON 89788, Is equivalent to that of conventional soybean (Glycine max L.). J. Agric. Food Chem. 2008, 2006, 4611–4622. [Google Scholar] [CrossRef]
- Chaudhary, J.; Deshmukh, R.; Mir, Z.A.; Bhat, J.A. Metabolomics: An Emerging Technology for Soybean Improvement; Springer International Publishing: Berlin/Heidelberg, Germany, 2019; ISBN 9783319923994. [Google Scholar]
- Krishnan, H.B.; Song, B.; Oehrle, N.W.; Cameron, J.C.; Jez, J.M. Impact of overexpression of cytosolic isoform of O-acetylserine sulfhydrylase on soybean nodulation and nodule metabolome. Sci. Rep. 2018, 8, 2367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyung, H.S.; Ryu, W. Metabolomics investigation of flavonoid synthesis in soybean leaves depending on the growth stage. Metabolomics 2014, 10, 833–841. [Google Scholar] [CrossRef]
- Seo, H.S.; Lee, S.; Singh, D.; Shin, H.W.; Cho, S.A.; Lee, C.H. Untargeted metabolite profiling for koji-fermentative bioprocess unravels the effects of varying substrate types and microbial inocula. Food Chem. 2018, 266, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Nahar, K.; Hasanuzzaman, M.; Fujita, M. Heat Stress Responses and Thermotolerance in Soybean; Elsevier Inc.: Amsterdam, The Netherlands, 2016; ISBN 9780128015360. [Google Scholar]
- Gu, E.; Wook, D.; Jang, G.; Hwa, S.; Lee, J.; Bong, S.; Kim, B.; Cho, Y.; Lee, H.; Kim, H. Mass-based metabolomic analysis of soybean sprouts during germination. Food Chem. 2017, 217, 311–319. [Google Scholar] [CrossRef]
- Kwon, Y.S.; Lee, S.; Lee, S.H.; Kim, H.J.; Lee, C.H. Comparative evaluation of six traditional fermented soybean products in East Asia: A metabolomics approach. Metabolites 2019, 9, 183. [Google Scholar] [CrossRef]
- Yun, D.; Kang, Y.; Yun, B.; Kim, E.; Kim, M.; Park, J.S.; Lee, J.; Hong, Y.S. Distinctive metabolism of flavonoid between cultivated and semi-wild soybean unveiled through metabolomics approach. J. Agric. Food Chem. 2016, 64, 5773–5783. [Google Scholar] [CrossRef]
- Bueno, P.C.P.; Lopes, N.P. Metabolomics to characterize adaptive and signaling responses in legume crops under abiotic stresses. ACS Omega 2020, 5, 1752–1763. [Google Scholar] [CrossRef]
- Demers, L.C. Comparative functional genomics characterization of low phytic acid soybeans and virus resistant soybeans. Ph.D. Dissertation, Virginia Polytechnic Institute and State University, Blacksburg, VA, USA, 2020. [Google Scholar]
- Schmidt, M.A.; Herman, E.M. Characterization and functional biology of the soybean aleurone layer. BMC Plant Biol. 2018, 8, 354. [Google Scholar] [CrossRef]
- Yi, L.; Dong, N.; Yun, Y.; Deng, B.; Liu, S.; Zhang, Y.; Liang, Y. Recent advances in chemometric methods for plant metabolomics: A review. Biotechnol. Adv. 2014, 914, 17–34. [Google Scholar] [CrossRef]
- Zampieri, M.; Sekar, K.; Zamboni, N.; Sauer, U. Frontiers of high-throughput metabolomics. Curr. Opin. Chem. Biol. 2017, 36, 15–23. [Google Scholar] [CrossRef]
- Maruyama, Y.; Toya, Y.; Kurokawa, H.; Fukano, Y.; Sato, A.; Umemura, H.; Yamada, K.; Iwasaki, H.; Tobori, N.; Shimizu, H. Characterization of oil-producing yeast Lipomyces starkeyi on glycerol carbon source based on metabolomics and 13C-labeling. Appl. Microbiol. Biotechnol. 2018, 102, 8909–8920. [Google Scholar] [CrossRef]
- Tugizimana, F.; Piater, L.A.; Dubery, I.A. Plant metabolomics: A new frontier in phytochemical analysis. S. Afr. J. Sci. 2013, 109, 18–20. [Google Scholar] [CrossRef]
- Hall, R.D. Plant metabolomics: From holistic hope, to hype, to hot topic. New Phytol. 2006, 169, 453–468. [Google Scholar] [CrossRef] [PubMed]
- Schauer, N.; Fernie, A. Plant metabolomics: Towards biological function and mechanism. Trends Plant Sci. 2006, 11, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.W.; Rutan, S.C. Chemometrics for the analysis of chromatographic data in metabolomics investigations. J. Chemom. 2014, 28, 681–687. [Google Scholar] [CrossRef]
- Djande, C.Y.H.; Pretorius, C.; Tugizimana, F.; Piater, L.A.; Dubery, I.A. Metabolomics: A Tool for Cultivar phenotyping and investigation of grain crops. Agronomy 2020, 10, 831. [Google Scholar] [CrossRef]
- Ranjbar, M.R.N.; Zhao, Y.; Tadesse, M.G.; Wang, Y.; Ressom, H.W. Gaussian process regression model for normalization of LC-MS data using scan-level information. Proteome Sci. 2013, 11, S13. [Google Scholar] [CrossRef]
- Kostidis, S.; Addie, R.D.; Morreau, H.; Mayboroda, O.A.; Giera, M. Quantitative NMR analysis of intra- and extracellular metabolism of mammalian cells: A tutorial. Anal. Chim. Acta 2017, 980, 1–24. [Google Scholar] [CrossRef]
- Nandania, J.; Peddinti, G.; Pessia, A.; Kokkonen, M.; Velagapudi, V. Validation and automation of a high-throughput multitargeted method for semiquantification of endogenous metabolites from different biological matrices using tandem mass spectrometry. Metabolites 2018, 8, 44. [Google Scholar] [CrossRef]
- Fuhrer, T.; Zamboni, N. High-throughput discovery metabolomics. Curr. Opin. Biotechnol. 2015, 31, 73–78. [Google Scholar] [CrossRef]
- Niu, Z.; Zhang, W.; Yu, C.; Zhang, J.; Wen, Y. Trends in analytical chemistry recent advances in biological sample preparation methods coupled with chromatography, spectrometry and electrochemistry analysis techniques. Trends Anal. Chem. 2018, 102, 123–146. [Google Scholar] [CrossRef]
- Antignac, J.; Dervilly-pinel, G.; Bizec, B. Le Basics of mass spectrometry based metabolomics. Proteomics 2014, 14, 2369–2388. [Google Scholar] [CrossRef]
- Jiao, L.; Tao, Y.; Wang, W.; Shao, Y.; Mei, L.; Wang, Q.; Dang, J. Preparative isolation of flavonoid glycosides from Sphaerophysa salsula using hydrophilic interaction solid-phase extraction coupled with two-dimensional preparative liquid chromatography. J. Sep. Sci. 2017, 40, 3808–3816. [Google Scholar] [CrossRef] [PubMed]
- Adegbe, A.A.; Larayetan, R.A.; Omojuwa, T.J. Proximate analysis, physicochemical properties and chemical constituents characterization of Moringa oleifera (Moringaceae) seed oil using GC-MS Analysis. Am. J. Chem. 2016, 6, 23–28. [Google Scholar] [CrossRef]
- Khoza, B.S.; Chimuka, L.; Mukwevho, E.; Steenkamp, P.A.; Madala, N.E. The effect of temperature on pressurised hot water extraction of pharmacologically important metabolites as analysed by UPLC-qTOF-MS and PCA. Evidence-based Complement. Altern. Med. 2014, 2014, 1–9. [Google Scholar] [CrossRef]
- Mohammed, N.K.; Yazid, M.; Manap, A.; Tan, C.P.; Muhialdin, B.J.; Alhelli, A.M.; Shobirin, A.; Hussin, M. The Effects of different extraction methods on antioxidant properties, chemical composition, and thermal behavior of black seed (Nigella sativa L.) oil. Evid. -Based Complementary Altern. Med. 2016, 2016, 6273817. [Google Scholar] [CrossRef]
- Bragagnolo, F.S.; Funari, C.S.; Ibáñez, E.; Cifuentes, A. Metabolomics as a tool to study underused soy parts: In search of bioactive compounds. Foods 2021, 10, 1308. [Google Scholar] [CrossRef]
- Louie, K.B.; Kosina, S.M.; Hu, Y.; Otani, H.; de Raad, M.; Kuftin, A.N.; Mouncey, N.J.; Bowen, B.P.; Northen, T.R. Mass spectrometry for natural product discovery. In Comprehensive Natural Products III; Elsevier Ltd.: Amsterdam, The Netherlands, 2020; pp. 263–306. ISBN 9780124095472. [Google Scholar]
- Buszewski, B.; Rafińska, K.; Cvetanović, A.; Walczak, J. Phytochemical analysis and biological activity of Lupinus luteus seeds extracts obtained by supercritical fluid extraction. Phytochem. Lett. 2019, 30, 338–348. [Google Scholar] [CrossRef]
- Teo, C.C.; Tan, S.N.; Yong, J.W.H.; Hew, C.S.; Ong, E.S. Pressurized hot water extraction (PHWE). J. Chromatogr. A 2010, 1217, 2484–2494. [Google Scholar] [CrossRef]
- Dettmer, K.; Aronov, P.A.; Hammock, B.D. Mass spectrometry-based metabolomics. Mass Spectrom. Rev. 2007, 26, 51–78. [Google Scholar] [CrossRef]
- Porzel, A.; Farag, M.A.; Mülbradt, J.; Wessjohann, L.A. Metabolite profiling and fingerprinting of Hypericum species: A comparison of MS and NMR metabolomics. Metabolomics 2014, 10, 574–588. [Google Scholar] [CrossRef]
- Sehlakgwe, P.F.; Lall, N.; Prinsloo, G. 1H-NMR Metabolomics and LC-MS Analysis to determine seasonal variation in a cosmeceutical plant Leucosidea sericea. Front. Pharmacol. 2020, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Allwood, J.W.; Goodacre, R. An introduction to liquid chromatography—mass spectrometry instrumentation applied in plant metabolomic analyses. Phytochem. Anal. 2010, 21, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.-J.; Schultz, A.W.; Wang, J.; Johnson, C.H.; Yannone, S.M.; Patti, G.J.; Siuzdak, G. Liquid chromatography quadrupole time-of-flight mass spectrometry characterization of metabolites guided by the METLIN database. Nat. Protoc. 2013, 8, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond biomarkers and towards mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef]
- Mahrous, E.A.; Farag, M.A. Two dimensional NMR spectroscopic approaches for exploring plant metabolome: A review. J. Adv. Res. 2015, 6, 3–15. [Google Scholar] [CrossRef]
- Kuhn, S.; Colreavy-donnelly, S. An integrated approach for mixture analysis using MS and NMR techniques. Faraday Discuss 2019, 218, 339–353. [Google Scholar] [CrossRef]
- Tugizimana, F.; Ncube, E.N.; Steenkamp, P.A.; Dubery, I.A. Metabolomics-derived insights into the manipulation of terpenoid synthesis in Centella asiatica cells by methyl jasmonate. Plant Biotechnol. Rep. 2015, 9, 125–136. [Google Scholar] [CrossRef]
- Hantao, L.W.; de Lima Ribeiro, F.A.; Passador, M.M.; Furtado, E.L.; Poppi, R.J.; Gozzo, F.C.; Augusto, F. Metabolic profiling by ultra-performance liquid chromatography-mass spectrometry and parallel factor analysis for the determination of disease biomarkers in Eucalyptus. Metabolomics 2014, 10, 1318–1325. [Google Scholar] [CrossRef]
- Putri, S.P.; Yamamoto, S.; Tsugawa, H.; Fukusaki, E. Current metabolomics. Technological advances. J. Biosci. Bioeng. 2013, 116, 9–16. [Google Scholar] [CrossRef]
- Ernst, M.; Silva, D.B.; Silva, R.R.; Vêncio, R.Z.N.; Lopes, N.P. Mass spectrometry in plant metabolomics strategies: From analytical platforms to data acquisition and processing. Nat. Prod. Rep. 2014, 31, 784. [Google Scholar] [CrossRef] [PubMed]
- Gika, H.G.; Wilson, I.D.; Theodoridis, G.A. The Role of Mass Spectrometry in Nontargeted Metabolomic, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 63, ISBN 0166526X. [Google Scholar]
- Wolfender, J.-L.; Aurelien, G.M.; Bertrand, T.S. Current approaches and challenges for the metabolite profiling of complex natural extracts. J. Chromatogr. A 2015, 1382, 136–164. [Google Scholar] [CrossRef] [PubMed]
- Van Den Berg, R.A.; Hoefsloot, H.C.J.; Westerhuis, J.A.; Smilde, A.K.; Van DerWerf, M.J. Centering, scaling, and transformations: Improving the biological information content of metabolomics data. BMC Genomics 2006, 7, 142. [Google Scholar] [CrossRef] [PubMed]
- Castillo, S.; Gopalacharyulu, P.; Yetukuri, L.; Oresic, M. Algorithms and tools for the preprocessing of LC—MS metabolomics data. Chemom. Intell. Lab. Syst. 2011, 108, 23–32. [Google Scholar] [CrossRef]
- Xu, Y.F.; Lu, W.; Rabinowitz, J.D. Avoiding misannotation of in-source fragmentation products as cellular metabolites in liquid chromatography-mass spectrometry-based metabolomics. Anal. Chem. 2015, 87, 2273–2281. [Google Scholar] [CrossRef]
- Allwood, J.W.; De Vos, R.C.H.; Moing, A.; Deborde, C.; Erban, A.; Kopka, J.; Goodacre, R.; Hall, R.D. Plant metabolomics and its potential for systems biology research. In Methods in Systems Biology; Elsevier Inc.: Amsterdam, The Netherlands, 2011; Volume 500, pp. 299–336. ISBN 9780123851185. [Google Scholar]
- Trygg, J.; Holmes, E.; Lundstedt, T. Chemometrics in metabonomics. J. Proteome Res. 2007, 6, 469–479. [Google Scholar] [CrossRef]
- Allwood, J.W.; Ellis, D.I.; Goodacre, R. Metabolomic technologies and their application to the study of plants and plant–host interactions. Physiol. Plant. 2008, 132, 117–135. [Google Scholar] [CrossRef]
- Saccenti, E.; Hoefsloot, H.C.J.; Smilde, A.K.; Westerhuis, J.A.; Hendriks, M.M.W.B. Reflections on univariate and multivariate analysis of metabolomics data. Metabolomics 2014, 10, 361–374. [Google Scholar] [CrossRef]
- Olivon, F.; Roussi, F.; Litaudon, M.; Touboul, D. Optimized experimental workflow for tandem mass spectrometry molecular networking in metabolomics. Anal. Bioanal. Chem. 2017, 409, 5767–5778. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Furumichi, M.; Morishima, K.; Tanabe, M. New approach for understanding genome variations in KEGG. Nucleic Acids Res. 2019, 47, D590–D595. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y. KEGG Mapper for inferring cellular functions from protein sequences. Protein Sci. 2020, 29, 28–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Hu, B.; Liu, W.; Qin, W.; Wu, H.; Zhang, J.; Yang, C.; Deng, J.; Shu, K.; Du, J.; et al. Metabolomic tool to identify soybean [Glycine max (L.) Merrill] germplasms with a high level of shade tolerance at the seedling stage. Sci. Rep. 2017, 7, 42478. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, A.; Kusano, M. Recent Progress in the development of metabolome databases for plant systems biology. Front. Plant Sci. 2013, 4, 73. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis: Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef]
- Bhalla, R.; Narasimhan, K.; Swarup, S. Metabolomics and its role in understanding cellular responses in plants. Plant Cell Rep. 2005, 24, 562–571. [Google Scholar] [CrossRef]
- Maruyama, K.; Urano, K.; Kusano, M.; Sakurai, T.; Takasaki, H.; Kishimoto, M. Metabolite/phytohormone—gene regulatory networks in soybean organs under dehydration conditions revealed by integration analysis. Plant J. 2020, 103, 197–211. [Google Scholar] [CrossRef]
- Silvente, S.; Sobolev, A.P.; Lara, M. Metabolite adjustments in drought tolerant and sensitive soybean genotypes in response to water stress. PLoS ONE 2012, 7, e38554. [Google Scholar] [CrossRef]
- Wang, X.; Guo, R.; Li, M.; Liu, Y.; Zhao, M.; Fu, H.; Liu, X.; Wang, S. Metabolomics reveals the drought - tolerance mechanism in wild soybean (Glycine soja). Acta Physiol. Plant. 2019, 41, 1–11. [Google Scholar] [CrossRef]
- Díaz-cruz, G.A.; Cassone, B.J. A tale of survival: Molecular defense mechanisms of soybean to overcome soybean mosaic virus infection physiological and molecular plant pathology. A tale of survival: Molecular defense mechanisms of soybean to overcome Soybean Mosaic Virus infection. Physiol. Mol. Plant Pathol. 2017, 102, 79–87. [Google Scholar] [CrossRef]
- Hu, B.-Y.; Yang, C.-Q.; Iqbal, N.; Deng, J.-C.; Zhang, J.; Yang, W.-Y.; Liu, J. Development and validation of a GC–MS method for soybean organ-specific metabolomics. Plant Prod. Sci. 2018, 21, 215–224. [Google Scholar] [CrossRef]
- Coutinho, I.D.; Henning, L.M.M.; Döpp, S.A.; Nepomuceno, A.; Moraes, L.A.C.; Marcolino-Gomes, J.; Richter, C.; Schwalbe, H.; Colnago, L.A. Flooded soybean metabolomic analysis reveals important primary and secondary metabolites involved in the hypoxia stress response and tolerance. Environ. Exp. Bot. 2018, 153, 176–187. [Google Scholar] [CrossRef] [Green Version]
- Kang, W.; Chen, L.; Wang, Y.; Zhu, X.; Liu, X.; Duan, Y. Bacillus simplex treatment promotes soybean defence against soybean cyst nematodes: A metabolomics study using GC-MS. PLoS ONE 2020, 15, e0237194. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, N.; Nakamura, T.; Komatsu, S. Differential responses of microsomal proteins and metabolites in two contrasting cadmium (Cd -accumulating soybean cultivars under Cd stress. Amino Acids 2012, 42, 317–327. [Google Scholar] [CrossRef]
- Rabara, R.C.; Tripathi, P.; Rushton, P.J. Comparative metabolome profile between tobacco and soybean grown under water-stressed conditions. Biomed Res. Int. 2017, 2017, 3065251. [Google Scholar] [CrossRef]
- Das, A.; Rushton, P.J.; Rohila, J.S. Metabolomic profiling of soybeans (Glycine max L.) Reveals the Importance of Sugar and Nitrogen. Plants 2017, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Hu, C.; Hussain, S.; Tan, Q.; Wu, S. Ecotoxicology and environmental safety metabolomics analysis reveals potential mechanisms of tolerance to excess molybdenum in soybean seedlings. Ecotoxicol. Environ. Saf. 2018, 164, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Xu, J.; Wang, X.; Fu, H.; Zhao, M.; Wang, H.; Shi, L. Photosynthetic characteristics and metabolic analyses of two soybean genotypes revealed adaptive strategies to low-nitrogen stress. J. Plant Physiol. 2018, 229, 132–141. [Google Scholar] [CrossRef]
- Jiao, Y.; Bai, Z.; Xu, J.; Zhao, M.; Khan, Y.; Hu, Y.; Shi, L. Metabolomics and its physiological regulation process reveal the salt- tolerant mechanism in Glycine soja seedling roots. Plant Physiol. Biochem. 2018, 126, 187–196. [Google Scholar] [CrossRef]
- Li, M.; Guo, R.; Jiao, Y.; Jin, X.; Zhang, H.; Shi, L. Comparison of salt tolerance in Soja Based on metabolomics of seedling roots. Front. Plant Sci. 2017, 8, 1101. [Google Scholar] [CrossRef]
- Xu, G.; Singh, S.; Barnaby, J.; Buyer, J.; Reddy, V.; Sicher, R. Effects of growth temperature and carbon dioxide enrichment on soybean seed components at different stages of development. Plant Physiol. Biochem. 2016, 108, 313–322. [Google Scholar] [CrossRef]
- Copley, T.R.; Aliferis, K.A.; Kliebenstein, D.J.; Jabaji, S.H. An integrated RNAseq- 1 H NMR metabolomics approach to understand soybean primary metabolism regulation in response to Rhizoctonia foliar blight disease. BMC Plant Biol. 2017, 17, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hrbek, V.; Krtkova, V.; Rubert, J.; Chmelarova, H.; Demnerova, K.; Ovesna, J.; Hajslova, J. Metabolomic strategies based on High-Resolution Mass Spectrometry as a tool for recognition of GMO (MON 89788 Variety) and Non-GMO Soybean: A critical assessment of two complementary methods. Food Anal. Methods 2017, 10, 3723–3737. [Google Scholar] [CrossRef]
- Fu, H.; Guo, R.; Shen, W.Y.; Li, M.X.; Liu, Y.; Zhao, M.L.; Wang, X.X.; Liu, X.Y. Changes in the metabolome of two soybean genotypes under drought stress. Russ. J. Plant Physiol. 2020, 67, 472–481. [Google Scholar] [CrossRef]
- Yang, A.; Kong, L.; Wang, H.; Yao, X.; Xie, F.; Wang, H. Response of soybean root to phosphorus deficiency under sucrose feeding: Insight from morphological and metabolome characterizations. BioMed Res. Int. 2020, 2020, 2148032. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Xu, J.; Guo, R.; Liu, Y.; Wang, S.; Wang, H.; Ullah, A.; Shi, L. Identifying the metabolomics and physiological differences among Soja in the early flowering stage. Plant Physiol. Biochem. 2019, 139, 82–91. [Google Scholar] [CrossRef]
- Salloum, M.S.; Insani, M.; Monteoliva, M.I.; Menduni, M.F.; Silvente, S.; Carrari, F.; Luna, C. Metabolic responses to arbuscular mycorrhizal fungi are shifted in roots of contrasting soybean genotypes. Mycorrhiza 2019, 29, 459–473. [Google Scholar] [CrossRef] [PubMed]
- Pi, E.; Xu, J.; Li, H.; Fan, W.; Zhu, C.; Zhang, T.; Jiang, J.; He, L.; Lu, H.; Wang, H.; et al. Enhanced salt tolerance of rhizobia-inoculated soybean correlates with decreased phosphorylation of the transcription factor GmMYB183 and altered flavonoid biosynthesis enhanced salt tolerance of rhizobia- inoculated soybean correlates with decreased phos. Mol. Cell. Proteom. 2019, 18, 2225–2243. [Google Scholar] [CrossRef]
- John, K.M.M.; Khan, F.; Luthria, D.L.; Matthews, B.; Garrett, W.M.; Natarajan, S. Proteomic and metabolomic analysis of minimax and Williams 82 soybeans grown under two different conditions. J. Food Biochem. 2017, 41, e12404. [Google Scholar] [CrossRef]
- Pastor, V.; Luna, E.; Ton, J.; Flors, V. Primed plants do not forget. Environ. Exp. Bot. 2012, 94, 46–56. [Google Scholar] [CrossRef]
- Chen, W.; Gao, Y.; Xie, W.; Gong, L.; Lu, K.; Wang, W.; Li, Y.; Liu, X. Genome-wide association analyses provide genetic and biochemical insights into natural variation in rice metabolism. Nat. Publ. Gr. 2014, 46, 714–721. [Google Scholar] [CrossRef]
- Yadav, C.B.; Srivastava, R.K.; Mur, L.A.J.; Yadav, R.S. Metabolite diversity and metabolic genome-wide marker traits in pearl millet grains. Cells 2021, 10, 3076. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
A systems biology perspective on the biological information pipeline. The illustration depicts the integrated flow of biological data via the omics system, from the genome to the metabolome. Metabolomics provides a comprehensive overview of an organism’s biochemical and physiological status, and changed metabolomes reflect changes in the genome, transcriptome, and proteome. As a result, the metabolome is regarded as the underlying biochemical layer that reflects all information expressed and regulated across all the omics layers, providing the most direct relationship to the phenotype. Figure created using BioRender (https://biorender.com/).
Figure 1.
A systems biology perspective on the biological information pipeline. The illustration depicts the integrated flow of biological data via the omics system, from the genome to the metabolome. Metabolomics provides a comprehensive overview of an organism’s biochemical and physiological status, and changed metabolomes reflect changes in the genome, transcriptome, and proteome. As a result, the metabolome is regarded as the underlying biochemical layer that reflects all information expressed and regulated across all the omics layers, providing the most direct relationship to the phenotype. Figure created using BioRender (https://biorender.com/).
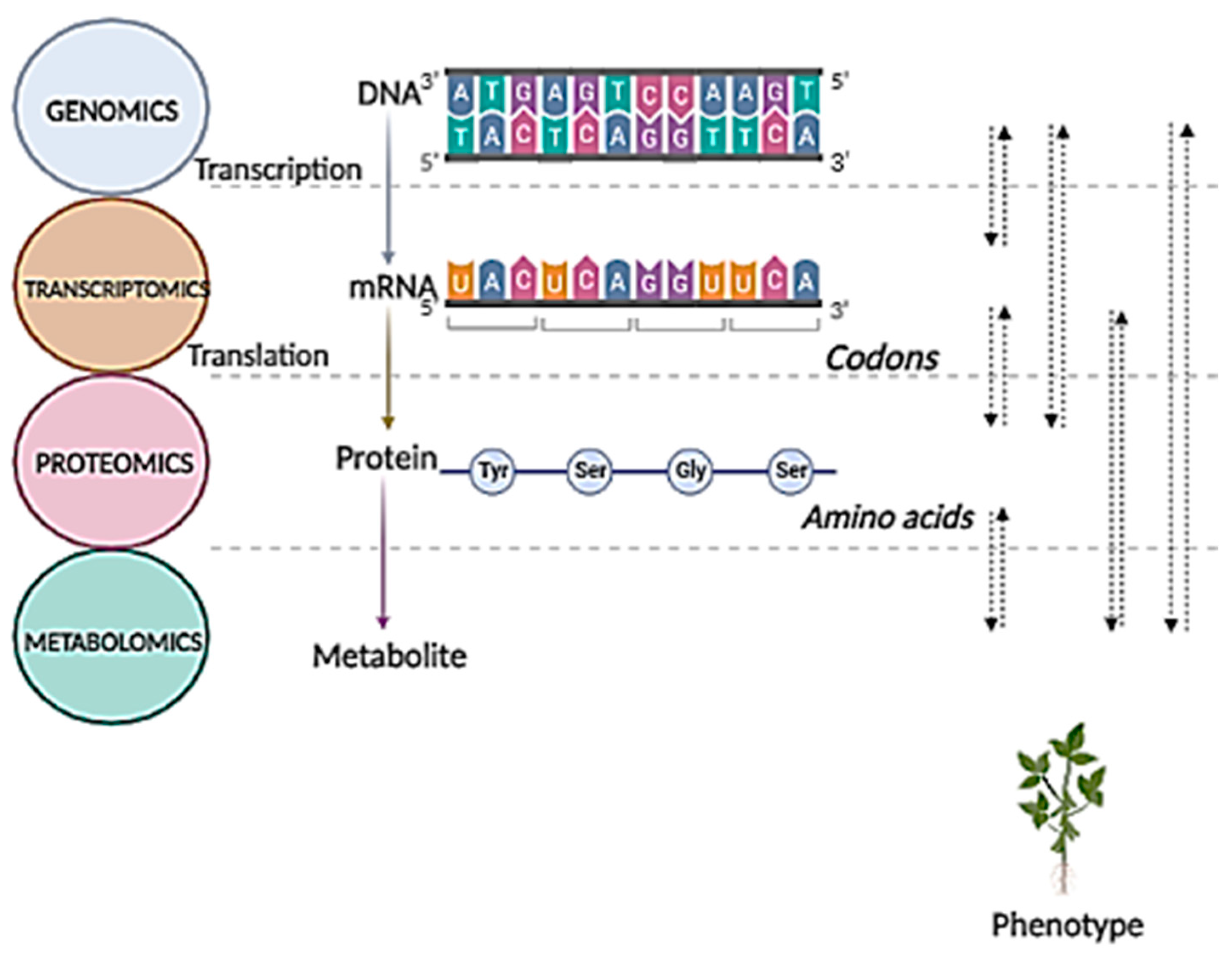
Figure 2.
Workflow of a metabolomics approach. Figure created using BioRender (https://biorender.com/).
Figure 2.
Workflow of a metabolomics approach. Figure created using BioRender (https://biorender.com/).

{kind=link}
{kind=link}
Table 1.
Recent progress in soybean metabolomics studies for identification of key biomarkers to mitigate biotic and abiotic stress tolerance and other growth conditions.
Table 1.
Recent progress in soybean metabolomics studies for identification of key biomarkers to mitigate biotic and abiotic stress tolerance and other growth conditions.
| Objective of the Study | Analytical Platform | Tissue | Other Omics | Main Finding | References |
|---|---|---|---|---|---|
| Analyzed the organ specificity of metabolites and identification of the features of their regulatory networks in dehydrated soybeans | GC-TOF-MS LC-MS | Leaves Stems Roots | Transcriptomics | ABA is the most highly dehydration-inducible phytohormone in plant aerial parts. | [74] |
| Investigated metabolite changes in relation to physiological responses two soybean genotypes with varying drought tolerance | 1H NMR | Leaves Nodule | Markers important for determining water stress response were identified. | [75] | |
| Elucidated the mechanism behind drought tolerance in drought-tolerant wild soybean | GC-MS | Leaves | Drought-stress mechanisms include the accumulation of osmotic chemicals, as well as an increase in energy and secondary antioxidant metabolism. Drought resistance in wild soybeans. | [76] | |
| Described the metabolic changes in soybean leaves ten days after Soybean mosaic virus infection (SMV) | LC-MS/MS | Leaves | Transcriptomics | There were significant changes in amino acid concentrations in connection to viral infection at the metabolomic level. | [77] |
| Investigated the potential organ-specific resistance mechanism of soybean to F. Moniliforme | GC–MS | Seeds Pods | F. Moniliforme disrupted amino acid metabolism in soybean seeds, and metabolic pathways involved to energy conversion in soybean pods responded substantially to fungal infection. | [78] | |
| Examining the responses to flooding stress in roots and leaves of two soybean cultivars (BR4 and Embrapa 45, sensitive and moderately tolerant to flooding stress, respectively). | 1H NMR | Roots Leaves | Different reactions were observed in the roots and leaves, as well as in flood-tolerant and flood-sensitive cultivars. The majority of the molecules that have transformed are associated to carbon and nitrogen metabolism, as well as the phenylpropanoid pathway. | [79] | |
| Two wild soybean types with varying salt tolerance were chosen, and metabolic alterations in response to neutral-salt stress and alkali-salt stress were studied. | GC–MS | Leaf | The salt-tolerant wild soybean modifies amino acid and organic acid metabolism to generate more suitable solutes and promote the TCA cycle to produce more ATP. | [3] | |
| Investigated the metabolic changes in soybean cyst nematodes after treatment with Sneb545Bacillus simplex. Roots of SCN-infected soybeans | GC-MS | Root | Soybeans treated with Sneb545 have certain characteristics of SCN disease-resistant soybeans. | [80] | |
| Investigated Cd absorption and translocation in two different Cd-accumulating soybean cultivars | CE-MS | Roots | Proteomics | In the Enrei cultivar under Cd stress, amino acids linked to Cd-chelating pathways are quite active. | [81] |
| Investigated drought tolerance in tobacco and soybean plants to unravel metabolic pathways affected by increasing dehydration | LC-MS LC-MS/MS GC-MS | Root Leaf | In both species, the accumulation of metabolites is strongly linked to the degree of dehydration. | [82] | |
| Profiled leaf metabolites under control conditions, drought, and heat stress in a controlled setting. | LC-MS GC-MS | Leaves | Drought and heat stress were found to affect metabolites for various cellular processes which regulate carbohydrate metabolism, amino acid metabolism, peptide metabolism, and purine and pyrimidine biosynthesis. | [83] | |
| Investigated changes in the metabolic profiles of leaves and roots of soybean (Glycine max L.) Seedlings cultivated under normal and excess Mo conditions. | LC-MS/MS | Roots Leaves | Mo stress induced only lipid metabolism and salicylic acid buildup in leaves, whilst in roots the ascorbate–glutathione metabolism and flavonoid/isoflavone biosynthesis significantly increased. | [84] | |
| Analyzed of two soybean genotypes at the metabolomic level revealed the mechanism of low-nitrogen tolerance. | GC–MS | Leaves Roots | In order to tolerate low nitrogen, wild soybean synthesizes favorable secondary metabolites under low-nitrogen stress. | [85] | |
| Examined metabolomics features of wild soybean under several forms of salt stress to determine salt-tolerant processes in wild soybean in the field | GC–MS | Roots | Under neutral-salt stress, the salt-tolerant wild soybean showed enhanced amino acid, carbohydrate, and polyol metabolisms, whereas under alkali-salt stress, it showed improved organic acid, amino acid, and tricarboxylic acid metabolisms. | [86] | |
| Explored the salt tolerance-related mechanisms among Soja, wild soybean, semi-wild soybean, and cultivated soybean under two types of salt stress | GC–MS | Roots | Carbon and nitrogen metabolism, as well as the tricarboxylic acid (TCA) cycle and receiver operating properties (particularly phenolic substance metabolism) of seedling roots, were critical for salt stress resistance and demonstrated a steady decreasing trend from wild soybean to cultivated soybean. | [87] | |
| Determined the effects of growth temperature and carbon dioxide enrichment on soybean seed components at different stages of development | GC–MS | Seeds | CO2 (enrichment) treatments significantly changed the composition of early seeds but had little effect on mature seeds. Treatment effects on seed constituents were ranked as follows: Age > Temperature > CO2. | [88] | |
| Characterized the resistance of soybeans to foxglove aphid, Aulacorthum solani Kaltenbach, at the metabolite level. | CE–TOF–MS | Leaves | Differences in the amino acids in the soybean leaves influenced the free amino acids found in the aphids, which might be implicated in aphid resistance. | [1] | |
| Investigated variations in soybean metabolism in response to R. solani infection during early and late disease phases, focusing on the regulation of soybean primary metabolism and oxidative stress tolerance | 1H NMR | Leaves | Transcriptomics | In response to R. solani infection, significant changes in soybean primary metabolism occurred and metabolite levels involved in redox reactions and ROS signaling were also recorded. | [89] |
| Distinguished between genetically modified organisms (Monsanto 89,788 variety) and organic soybeans | DART-HRMS HPLC-HMRS | Seeds | The most important markers were found to be phosphatidylcholines and sugars. | [90] | |
| Compared the response mechanisms of wild and cultivated soybean to water stress | GC–MS | Leaves | Drought tolerance mechanisms included increasing primary metabolism to control osmotic potential, synthesizing desirable secondary metabolites and fatty acids, and maintaining a symbiotic relationship. | [91] | |
| Explored global metabolomic modifications in low-P-tolerant (Liaodou, L13) and low-P-sensitive (Tiefeng 3, T3) soybean genotypes | LC-MS | Root | Metabolite profiles of both genotypes differed in their responses as numbers of metabolites were exclusively and differentially regulated within each genotype. | [92] | |
| Examined the impact of overexpressing OASS on soybean nodulation and nodule metabolome | LC-MS GC-MS | Nodules | There is a slight decrease in the availability of energy metabolites to OASS overexpressing soybean nodules, which is then offset by the breakdown of cellular components to meet the nodule energy metabolism needs. | [14] | |
| Evaluated root exudates of two soybean cultivars grown under low-, normal-, and high-K+ conditions | CE–TOF–MS | Root | Soybean cultivars differ in their capacity to release root metabolites by altering the exudation of certain metabolites for improved adaptability to high- and low-K conditions. | [5] | |
| Investigated the cellular metabolism-related differences among salt-tolerant wild soybean (W2), salt- sensitive wild soybean (W1) and cultivated soybean (C) in the early flowering stage to reveal the adaptive mechanisms. | GC–TOF–MS | Leaf | Carbohydrate and organic acid metabolism were relatively greater, while the amino acid content and secondary metabolism level were lower in C than W1 | [93] | |
| Evaluated the metabolic responses of improved (I-1) and unimproved (UI-4) soybean genotypes after AM root colonization | GC-MS | Roots | The I-1 genotype has lower quantities of isoflavonoids and alpha-tocopherol and greater levels of malondialdehyde, that can affect the soybean-AM symbiosis. | [94] | |
| Investigated secondary metabolites produced when soybean plants were infected by A. Besseyi. | LC–ESI–MS–MS | Root | There were metabolome variations in root defensive chemicals in response to A. Besseyi attack, as indicated by an increase in the level of flavonoids. | [68] | |
| Identify metabolic changes in soybean roots treated with rhizobia inoculation and salt | LC–TOFMS | Root | Phosphoproteomics | Rhizobia symbiosis enables the soybean plant to adapt with the negative consequences of high soil salt, mostly by increasing ROS scavenging activities. | [95] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ncube, E.; Mohale, K.; Nogemane, N. Metabolomics as a Prospective Tool for Soybean (Glycine max) Crop Improvement. Curr. Issues Mol. Biol. 2022, 44, 4181-4196. https://doi.org/10.3390/cimb44090287
AMA Style
Ncube E, Mohale K, Nogemane N. Metabolomics as a Prospective Tool for Soybean (Glycine max) Crop Improvement. Current Issues in Molecular Biology. 2022; 44(9):4181-4196. https://doi.org/10.3390/cimb44090287
Chicago/Turabian StyleNcube, Efficient, Keletso Mohale, and Noluyolo Nogemane. 2022. "Metabolomics as a Prospective Tool for Soybean (Glycine max) Crop Improvement" Current Issues in Molecular Biology 44, no. 9: 4181-4196. https://doi.org/10.3390/cimb44090287