
In Silico and In Vitro Exploration of Poziotinib and Olmutinib Synergy in Lung Cancer: Role of hsa-miR-7-5p in Regulating Apoptotic Pathway Marker Genes

Abstract
:1. Introduction
2. Materials and Methods
2.1. In Silico Analysis
2.2. Preparation of Drug Stocks
2.3. Cell Culture
2.4. MTT Cytotoxicity Assay
2.5. Gene Expression Analysis
2.6. Molecular Docking Analysis
2.7. Statistical Analysis
3. Results
3.1. In Silico Analysis Revealed Passive Human Gastrointestinal Absorption of Both Drugs and Downstream Action on the Apoptotic Pathway
3.2. Synergistic Effect of Poziotinib with Olmutinib on Cell Proliferation Activity
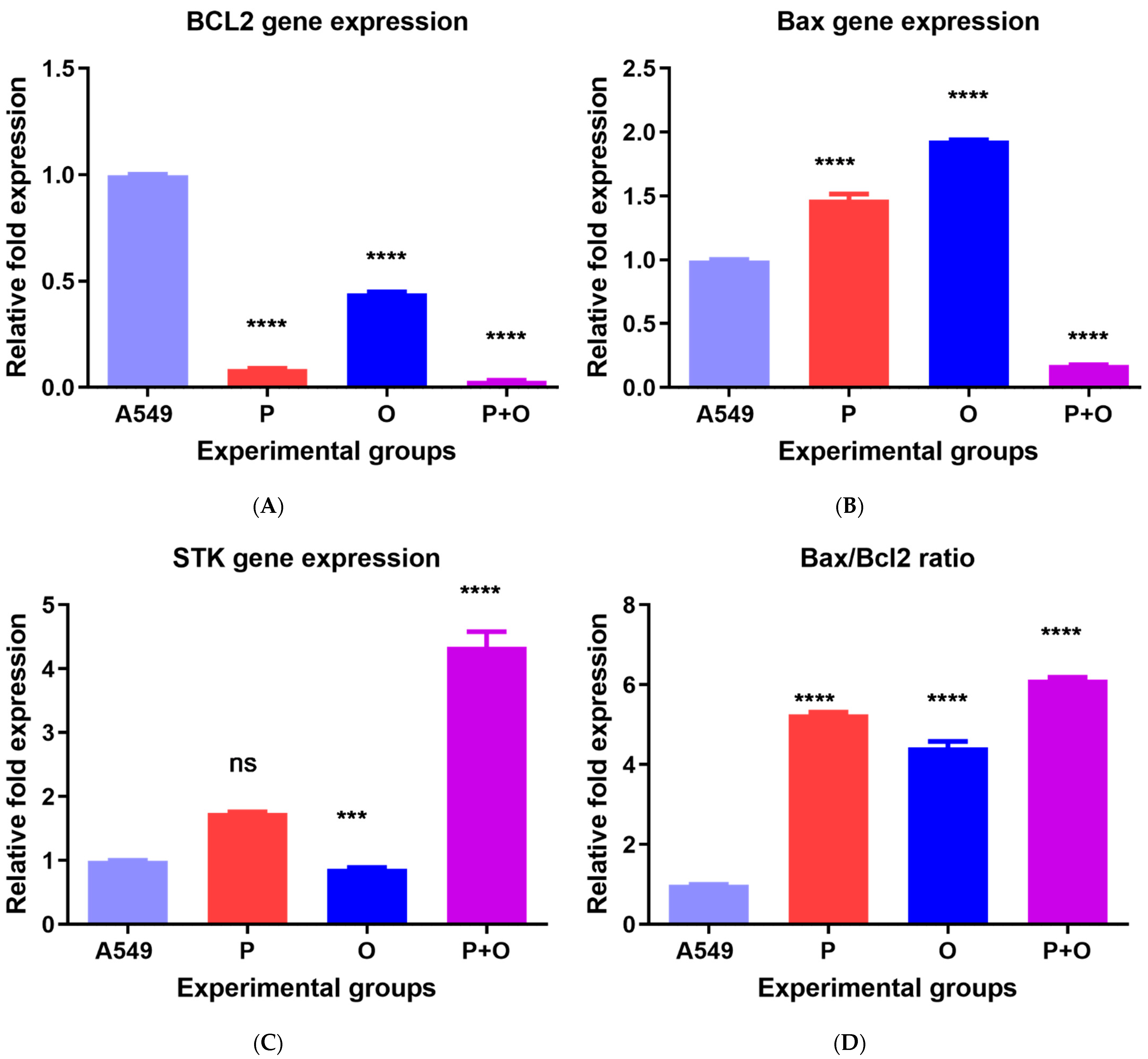
3.3. Gene Expression in Response to Combined Therapy and Olmutinib and Poziotinib Alone
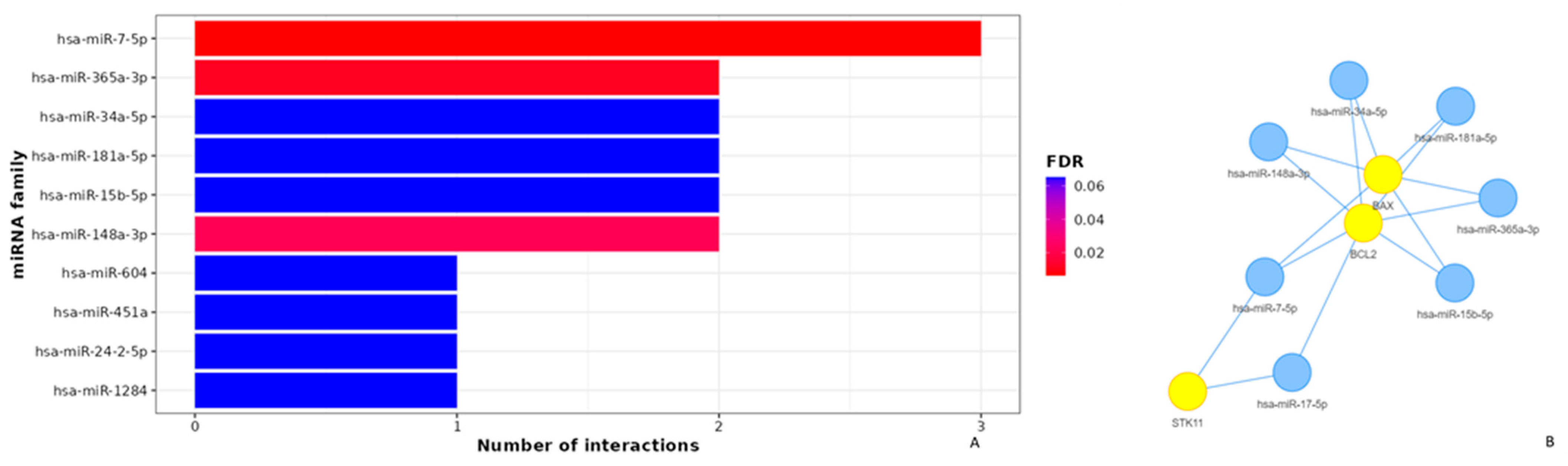
3.4. Apoptotic Effects of Poziotinib and Olmutinib in Lung Cancer Cells Are Mediated by Extrinsic and Intrinsic Apoptotic Pathway Genes through hsa-miR-7-5p
3.5. Molecular Docking Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Thandra, K.C.; Barsouk, A.; Saginala, K.; Aluru, J.S.; Barsouk, A. Epidemiology of lung cancer. Contemp. Oncol. 2021, 25, 45–52. [Google Scholar] [CrossRef]
- Pistritto, G.; Trisciuoglio, D.; Ceci, C.; Garufi, A.; D’Orazi, G. Apoptosis as anticancer mechanism: Function and dysfunction of its modulators and targeted therapeutic strategies. Aging 2016, 8, 603–619. [Google Scholar] [CrossRef] [PubMed]
- Garinet, S.; Wang, P.; Mansuet-Lupo, A.; Fournel, L.; Wislez, M.; Blons, H. Updated Prognostic Factors in Localized NSCLC. Cancers 2022, 14, 1400. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef]
- Lemjabbar-Alaoui, H.; Hassan, O.U.; Yang, Y.W.; Buchanan, P. Lung cancer: Biology and treatment options. Biochim. Biophys. Acta 2015, 1856, 189–210. [Google Scholar] [CrossRef] [PubMed]
- Chevallier, M.; Borgeaud, M.; Addeo, A.; Friedlaender, A. Oncogenic driver mutations in non-small cell lung cancer: Past, present and future. World J. Clin. Oncol. 2021, 12, 217–237. [Google Scholar] [CrossRef]
- Gazdar, A.F. Activating and resistance mutations of EGFR in non-small-cell lung cancer: Role in clinical response to EGFR tyrosine kinase inhibitors. Oncogene 2009, 28 (Suppl. S1), 24–31. [Google Scholar] [CrossRef]
- Remon, J.; Steuer, C.E.; Ramalingam, S.S.; Felip, E. Osimertinib and other third-generation EGFR TKI in EGFR-mutant NSCLC patients. Ann. Oncol. 2018, 29, i20–i27. [Google Scholar] [CrossRef]
- Yamaoka, T.; Kusumoto, S.; Ando, K.; Ohba, M.; Ohmori, T. Receptor Tyrosine Kinase-Targeted Cancer Therapy. Int. J. Mol. Sci. 2018, 19, 3491. [Google Scholar] [CrossRef] [PubMed]
- Meier, D.E.; Brawley, O.W. Palliative care and the quality of life. J. Clin. Oncol. 2011, 29, 2750–2752. [Google Scholar] [CrossRef] [PubMed]
- Alkhezayem, S.; Wani, T.A.; Wakil, S.; Aljuraysi, A.; Zargar, S. Transcriptome analysis of neratinib treated HER2 positive cancer model vs untreated cancer unravels the molecular mechanism of action of neratinib. Saudi Pharm. J. 2020, 28, 963–970. [Google Scholar] [CrossRef]
- Ishqi, H.M.; Sarwar, T.; Husain, M.A.; Rehman, S.U.; Tabish, M. Differentially expressed novel alternatively spliced transcript variant of tumor suppressor Stk11 gene in mouse. Gene 2018, 668, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Zargar, S.; Altwaijry, N.; Wani, T.A.; Alkahtani, H.M. Evaluation of the Possible Pathways Involved in the Protective Effects of Quercetin, Naringenin, and Rutin at the Gene, Protein and miRNA Levels Using In-Silico Multidimensional Data Analysis. Molecules 2023, 28, 4904. [Google Scholar] [CrossRef]
- Wang, S.; Cang, S.; Liu, D. Third-generation inhibitors targeting EGFR T790M mutation in advanced non-small cell lung cancer. J. Hematol. Oncol. 2016, 9, 34. [Google Scholar] [CrossRef] [PubMed]
- Sumbly, V.; Landry, I. Unraveling the Role of STK11/LKB1 in Non-small Cell Lung Cancer. Cureus 2022, 14, e21078. [Google Scholar] [CrossRef]
- Kim, A.; Cohen, M.S. The discovery of vemurafenib for the treatment of BRAF-mutated metastatic melanoma. Expert. Opin. Drug Discov. 2016, 11, 907–916. [Google Scholar] [CrossRef]
- Khunger, A.; Khunger, M.; Velcheti, V. Dabrafenib in combination with trametinib in the treatment of patients with BRAF V600-positive advanced or metastatic non-small cell lung cancer: Clinical evidence and experience. Ther. Adv. Respir. Dis. 2018, 12, 1753466618767611. [Google Scholar] [CrossRef] [PubMed]
- Nagano, T.; Tachihara, M.; Nishimura, Y. Mechanism of resistance to epidermal growth factor receptor-tyrosine kinase inhibitors and a potential treatment strategy. Cells 2018, 7, 212. [Google Scholar] [CrossRef] [PubMed]
- Gridelli, C.; Rossi, A.; Carbone, D.P.; Guarize, J.; Karachaliou, N.; Mok, T.; Petrella, F.; Spaggiari, L.; Rosell, R. Non-small-cell lung cancer. Nat. Rev. Dis. Primers 2015, 1, 15009. [Google Scholar] [CrossRef] [PubMed]
- Jia, C.-Y.; Li, J.-Y.; Hao, G.-F.; Yang, G.-F. A drug-likeness toolbox facilitates ADMET study in drug discovery. Drug Discov. Today 2020, 25, 248–258. [Google Scholar] [CrossRef]
- Daina, A.; Zoete, V. Application of the SwissDrugDesign online resources in virtual screening. Int. J. Mol. Sci. 2019, 20, 4612. [Google Scholar] [CrossRef]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 1997, 23, 3–25. [Google Scholar] [CrossRef]
- Nguyen, H.D.; Jo, W.H.; Hoang, N.H.M.; Kim, M.-S. Curcumin-attenuated TREM-1/DAP12/NLRP3/Caspase-1/IL1B, TLR4/NF-κB pathways, and tau hyperphosphorylation induced by 1, 2-diacetyl benzene: An in vitro and in silico study. Neurotox. Res. 2022, 40, 1272–1291. [Google Scholar] [CrossRef]
- Zargar, S.; Wani, T.A.; Rizwan Ahamad, S. An Insight into Wheat Germ Oil Nutrition, Identification of Its Bioactive Constituents and Computer-Aided Multidimensional Data Analysis of Its Potential Anti-Inflammatory Effect via Molecular Connections. Life 2023, 13, 526. [Google Scholar] [CrossRef] [PubMed]
- Licursi, V.; Conte, F.; Fiscon, G.; Paci, P. MIENTURNET: An interactive web tool for microRNA-target enrichment and network-based analysis. BMC Bioinform. 2019, 20, 545. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef]
- Nikanjam, M.; Kato, S.; Adashek, J.J.; Kurzrock, R. Cetuximab in Patients with Non-Small Cell Lung Cancer and EGFR Exon 20 Insertion Alterations. Clin. Oncol. Case Rep. 2022, 5, 210. [Google Scholar] [PubMed]
- Hasegawa, H.; Yasuda, H.; Hamamoto, J.; Masuzawa, K.; Tani, T.; Nukaga, S.; Hirano, T.; Kobayashi, K.; Manabe, T.; Terai, H.; et al. Efficacy of afatinib or osimertinib plus cetuximab combination therapy for non-small-cell lung cancer with EGFR exon 20 insertion mutations. Lung Cancer 2019, 127, 146–152. [Google Scholar] [CrossRef]
- Heist, R.S.; Christiani, D. EGFR-targeted therapies in lung cancer: Predictors of response and toxicity. Pharmacogenomics 2009, 10, 59–68. [Google Scholar] [CrossRef]
- Seshacharyulu, P.; Ponnusamy, M.P.; Haridas, D.; Jain, M.; Ganti, A.K.; Batra, S.K. Targeting the EGFR signaling pathway in cancer therapy. Expert. Opin. Ther. Targets 2012, 16, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.M.; Lee, K.W.; Oh, D.Y.; Lee, J.S.; Im, S.A.; Kim, D.W.; Han, S.W.; Kim, Y.J.; Kim, T.Y.; Kim, J.H.; et al. Phase 1 Studies of Poziotinib, an Irreversible Pan-HER Tyrosine Kinase Inhibitor in Patients with Advanced Solid Tumors. Cancer Res. Treat. 2018, 50, 835–842. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Huang, Z.; Han, L.; Gong, Y.; Xie, C. Mechanisms and management of 3rd-generation EGFR-TKI resistance in advanced non-small cell lung cancer (Review). Int. J. Oncol. 2021, 59, 90. [Google Scholar] [CrossRef]
- Weigel, M.T.; Meinhold-Heerlein, I.; Bauerschlag, D.O.; Schem, C.; Bauer, M.; Jonat, W.; Maass, N.; Mundhenke, C. Combination of imatinib and vinorelbine enhances cell growth inhibition in breast cancer cells via PDGFR β signalling. Cancer Lett. 2009, 273, 70–79. [Google Scholar] [CrossRef]
- Carneiro, B.A.; El-Deiry, W.S. Targeting apoptosis in cancer therapy. Nat. Rev. Clin. Oncol. 2020, 17, 395–417. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, L.; Yuan, X.; Ou, Y.; Zhu, X.; Cheng, Z.; Zhang, P.; Wu, X.; Meng, Y.; Zhang, L. The Relationship between the Bcl-2/Bax Proteins and the Mitochondria-Mediated Apoptosis Pathway in the Differentiation of Adipose-Derived Stromal Cells into Neurons. PLoS ONE 2016, 11, e0163327. [Google Scholar] [CrossRef]
- Kulsoom, B.; Shamsi, T.S.; Afsar, N.A.; Memon, Z.; Ahmed, N.; Hasnain, S.N. Bax, Bcl-2, and Bax/Bcl-2 as prognostic markers in acute myeloid leukemia: Are we ready for Bcl-2-directed therapy? Cancer Manag. Res. 2018, 10, 403–416. [Google Scholar] [CrossRef]
- Alkaf, A.; Al-Jafari, A.; Wani, T.A.; Alqattan, S.; Zargar, S. Expression of STK11 gene and its promoter activity in MCF control and cancer cells. 3 Biotech 2017, 7, 362. [Google Scholar] [CrossRef]
- Karuman, P.; Gozani, O.; Odze, R.D.; Zhou, X.C.; Zhu, H.; Shaw, R.; Brien, T.P.; Bozzuto, C.D.; Ooi, D.; Cantley, L.C.; et al. The Peutz-Jegher gene product LKB1 is a mediator of p53-dependent cell death. Mol. Cell 2001, 7, 1307–1319. [Google Scholar] [CrossRef] [PubMed]
- Jan, R.; Chaudhry, G.E. Understanding Apoptosis and Apoptotic Pathways Targeted Cancer Therapeutics. Adv. Pharm. Bull. 2019, 9, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Luo, X.; Li, P.; Tan, J.; Wang, X.; Xiang, T.; Ren, G. miR-7-5p suppresses cell proliferation and induces apoptosis of breast cancer cells mainly by targeting REGgamma. Cancer Lett. 2015, 358, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H. MiR-7-5p suppresses tumor metastasis of non-small cell lung cancer by targeting NOVA2. Cell. Mol. Biol. Lett. 2019, 24, 60. [Google Scholar] [CrossRef]
- Lai, J.; Yang, H.; Zhu, Y.; Ruan, M.; Huang, Y.; Zhang, Q. MiR-7-5p-mediated downregulation of PARP1 impacts DNA homologous recombination repair and resistance to doxorubicin in small cell lung cancer. BMC Cancer 2019, 19, 602. [Google Scholar] [CrossRef]
- Peng, J.; Liu, F.; Zheng, H.; Wu, Q.; Liu, S. IncRNA ZFAS1 contributes to the radioresistance of nasopharyngeal carcinoma cells by sponging hsa-miR-7-5p to upregulate ENO2. Cell Cycle 2021, 20, 126–141. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Zhu, Z.; Zhang, Y.; Ji, F.; Zhu, L.; Liu, M.; Deng, Y.; Lv, G.; Li, D.; Zhou, Z.; et al. MiR-7-5p/KLF4 signaling inhibits stemness and radioresistance in colorectal cancer. Cell Death Discov. 2023, 9, 42. [Google Scholar] [CrossRef]
- Fook-Alves, V.L.; de Oliveira, M.B.; Zanatta, D.B.; Strauss, B.E.; Colleoni, G.W. TP53 regulated inhibitor of apoptosis 1 (TRIAP1) stable silencing increases late apoptosis by upregulation of caspase 9 and APAF1 in RPMI8226 multiple myeloma cell line. Biochim. Biophys. Acta 2016, 1862, 1105–1110. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Wang, Q.; Mao, Y.; Gao, W.; Duan, S. Targeting the p53 signaling pathway in cancers: Molecular mechanisms and clinical studies. MedComm 2023, 4, e288. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence of the Gene Expression Primers |
|---|---|
| Stk-11 | F: 5′ GCCGGGACTGACGTGTAGA 3′ |
| R: 5′ CCCAAAAGGAAGGGAAAAACC 3′ | |
| Bcl-2 | F: 5′ AATGGGCAGCCGTTAGGAAA 3′ |
| R: 5′ GCGCCCAATACGACCAAATC 3′ | |
| Bax | F: 5′ GGCCCTTTTGCTTCAGGGTT 3′ |
| R: 5′ GGAAAAAGACCTCTCGGGGG 3′ | |
| Gapdh | F: 5′ ACAGTCAGCCGCATCTTCTT 3′ |
| R: 5′ TTGATTTTGGAGGGATCTCG 3′ |
| Molecule | Olmutinib | Poziotinib |
|---|---|---|
| Canonical SMILES | C=CC(=O)Nc1cccc(c1)Oc1nc(Nc2ccc (cc2)N2CCN(CC2)C)nc2c1scc2 | C=CC(=O)N1CCC(CC1)Oc1cc2c (ncnc2cc1OC)Nc1ccc(c(c1F)Cl)Cl |
| Formula | C26H26N6O2S | C23H21Cl2FN4O3 |
| MW | 486.59 | 491.34 |
| H-bond acceptors | 5 | 6 |
| H-bond donors | 2 | 1 |
| TPSA | 110.86 | 76.58 |
| i Log P | 4.07 | 4.13 |
| X Log P3 | 4.75 | 5.36 |
| W Log P | 4.15 | 5.42 |
| M Log P | 3.03 | 3.76 |
| Silicos-IT Log P | 3.88 | 4.85 |
| Consensus Log P | 3.98 | 4.7 |
| ESOL Log S | −5.77 | −6.16 |
| ESOL Class | Moderately soluble | Poorly soluble |
| Ali Log S | −6.81 | −6.72 |
| GI absorption | High | High |
| BBB permeant | No | No |
| Pgp substrate | Yes | No |
| CYP1A2 inhibitor | No | No |
| CYP2C19 inhibitor | Yes | Yes |
| CYP2C9 inhibitor | Yes | Yes |
| CYP2D6 inhibitor | Yes | Yes |
| CYP3A4 inhibitor | Yes | No |
| Log Kp (cm/s) | −5.9 | −5.49 |
| Lipinski violations | 0 | 0 |
| Ghose violations | 2 | 2 |
| Veber violations | 0 | 0 |
| Egan violations | 0 | 0 |
| Muegge violations | 0 | 1 |
| Bioavailability score | 0.55 | 0.55 |
| PAINS alerts | 1 | 0 |
| Brenk alerts | 1 | 2 |
| Synthetic accessibility | 4.18 | 3.3 |
| Lead-likeness violations | 3 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alamery, S.; AlAjmi, A.; Wani, T.A.; Zargar, S. In Silico and In Vitro Exploration of Poziotinib and Olmutinib Synergy in Lung Cancer: Role of hsa-miR-7-5p in Regulating Apoptotic Pathway Marker Genes. Medicina 2023, 59, 1923. https://doi.org/10.3390/medicina59111923
Alamery S, AlAjmi A, Wani TA, Zargar S. In Silico and In Vitro Exploration of Poziotinib and Olmutinib Synergy in Lung Cancer: Role of hsa-miR-7-5p in Regulating Apoptotic Pathway Marker Genes. Medicina. 2023; 59(11):1923. https://doi.org/10.3390/medicina59111923
Chicago/Turabian StyleAlamery, Salman, Anfal AlAjmi, Tanveer A. Wani, and Seema Zargar. 2023. "In Silico and In Vitro Exploration of Poziotinib and Olmutinib Synergy in Lung Cancer: Role of hsa-miR-7-5p in Regulating Apoptotic Pathway Marker Genes" Medicina 59, no. 11: 1923. https://doi.org/10.3390/medicina59111923