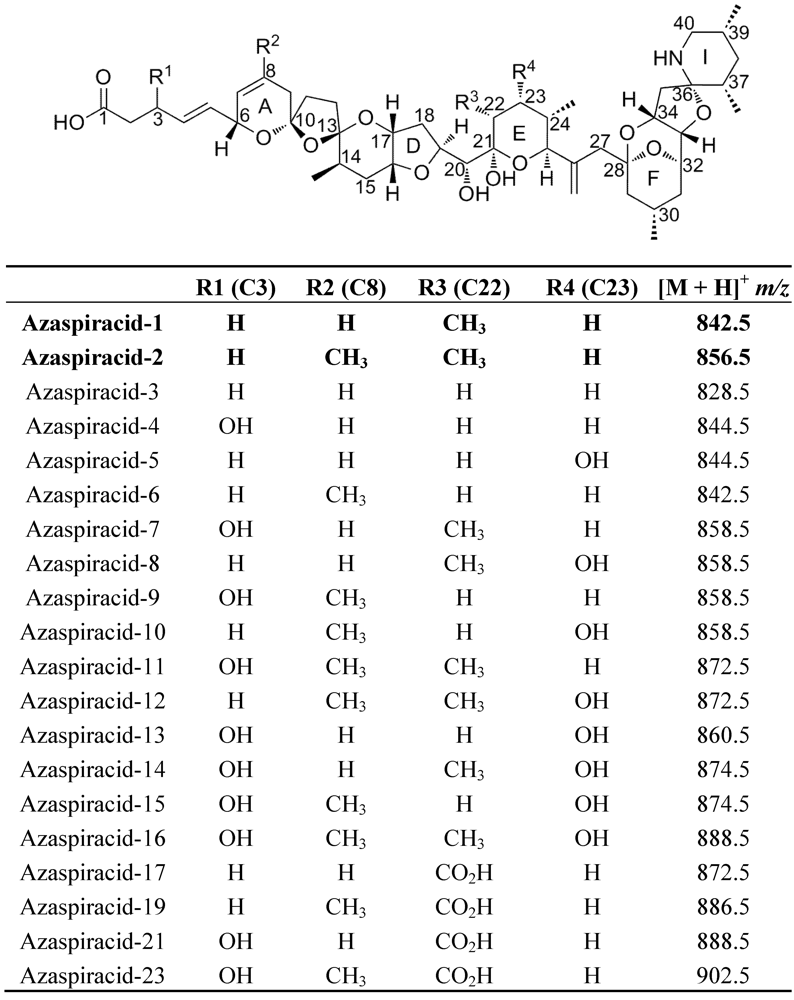
Production and Isolation of Azaspiracid-1 and -2 from Azadinium spinosum Culture in Pilot Scale Photobioreactors

 , , ,
, , , 
Abstract
:1. Introduction


2. Results and Discussion
2.1. Effect of Dilution Rate on A. spinosum and AZA Production in Pilot Scale Photobioreactors

{kind=link}
{kind=link}
{kind=link}
| A. spinosum | n | 0.1 day−1 | 0.15 day−1 | 0.2 day−1 | 0.25 day−1 | 0.3 day−1 | Main factors | Interaction | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| R1 | R2 | R1 | R2 | R1 | R2 | R1 | R2 | R1 | R2 | Dilution rate (D) | Reactor (R) | D–R | ||
| Concentration(×103 cell·mL−1) | 9–22 | 174 ± 6 | 164 ± 4 | 193 ± 6 | 214 ± 3 | 194 ± 8 | 214 ± 7 | 190 ± 6 | 221 ± 5 | 187 ± 5 | 220 ± 4 | p ≤ 0.05 | p ≤ 0.05 | p ≤ 0.05 |
| 0.1 < 0.15 = 0.2 = 0.25 = 0.3 | R1 < R2 | |||||||||||||
| Mean ESD(µm) | 9–22 | 9.81 ± 0.09 | 10.1 ± 0.1 | 9.59 ± 0.15 | 9.9 ± 0.2 | 9.6 ± 0.2 | 10.1 ± 0.2 | 9.29 ± 0.09 | 9.93 ± 0.04 | 9.5 ± 0.1 | 10.02 ± 0.05 | p ≤ 0.05 | p ≤ 0.05 | p ≤ 0.05 |
| 0.25 = 0.3 < 0.15 < 0.2 = 0.1 | R1 < R2 | |||||||||||||
| Cellular volume(×107 µm3·mL−1) | 9–22 | 8.8 ± 0.4 | 8.4 ± 0.3 | 9.2 ± 0.4 | 11.1 ± 0.4 | 9.3 ± 0.6 | 12.0 ± 0.7 | 8.2 ± 0.3 | 11.7 ± 0.2 | 8.5 ± 0.1 | 12.0 ± 0.4 | p ≤ 0.05 | p ≤ 0.05 | p ≤ 0.05 |
| 0.1 < 0.25 = 0.3 = 0.15 < 0.2 | R1 < R2 | |||||||||||||
| AZA1 (fg·cell−1) | 3–5 * | 37 ± 5 | 65 ± 8 | 52 ± 6 | 74 ± 4 | 34 ± 12 | 76 ± 14 | 26 ± 2 | 61 ± 3 | 17 ± 1 | 45 ± 3 | p ≤ 0.05 | p ≤ 0.05 | p ≤ 0.05 |
| AZA2 (fg·cell−1) | 3–5 * | 10 ± 1 | 16 ± 1 | 15 ± 1 | 24 ± 2 | 10 ± 2 | 19 ± 2 | 12 ± 2 | 25 ± 2 | 7 ± 1 | 18 ± 2 | |||
| AZA1 + 2 (fg·cell−1) | 3–5 * | 47 ± 6 | 81 ± 9 | 67 ± 3 | 98 ± 5 | 44 ± 13 | 95 ± 16 | 38 ± 2 | 86 ± 3 | 24 ± 1 | 63 ± 5 | 0.3 < 0.25 = 0.1< 0.2 < 0.15 | R1 < R2 | |
| Cell production (×109 cell·day−1) | n/a | 1.74 ± 0.06 | 1.64 ± 0.04 | 2.90 ± 0.09 | 3.21 ± 0.05 | 3.9 ± 0.2 | 4.3 ± 0.1 | 4.8 ± 0.2 | 5.5 ± 0.1 | 5.6 ± 0.2 | 6.6 ± 0.1 | Cell production = Cell concentration × D | ||
| Toxin production AZA1 + 2 (µg·day−1) | n/a | 82 ± 3 | 134 ± 15 | 193 ± 9 | 314 ± 15 | 170 ± 50 | 406 ± 64 | 180 ± 10 | 475 ± 17 | 134 ± 5 | 415 ± 33 | Toxin production = Cell production × [AZA1 + 2] | ||
2.2. Separation Procedure of A. spinosum from the Culture Medium
2.3. AZA1 and -2 Extraction from the Retentate and Permeate
| Acetone | ACN | DCM | HP-20 | |
|---|---|---|---|---|
| AZA1 + 2 (µg·g−1) | 17.4 ± 0.5 | 18 ± 2 | 17 ± 1 | 17 ± 1 |
| Purity (%) | 0.036 ± 0.002 | 0.07 ± 0.01 | 0.09 ± 0.01 | 0.21 ± 0.03 |
| Acetone Volume/5 g HP-20 | 3 × 5 mL | 3 × 10 mL | 3 × 25 mL | ||||
|---|---|---|---|---|---|---|---|
| AZA yield (µg/mL) | 1.61 ± 0.07 | 2.1 ± 0.1 | 2.19 ± 0.06 | ||||
| Mass HP-20/100 mL concentrate | 1 g | 2.5 g | 5 g | ||||
| AZA yield (µg/mL) | 2.6 ± 0.2 | 2.6 ± 0.1 | 2.1 ± 0.2 | ||||
| Time of contact | 2 h | 6 h | 24 h | 72 h | |||
| AZA yield (µg/mL) | 0.48 ± 0.06 | 0.52 ± 0.03 | 0.61 ± 0.02 | 0.52 ± 0.08 | |||
| HP-20 adsorption efficiency (% after 24 h of contact with the concentrate) | 93.8 ± 0.1 | ||||||
| Desorption procedure (2.5 g HP-20–3 × 7.5 mL acetone) | Soaking using acetone and filtration | Column using acetone | |||||
| Time of soaking (a and b), and flow rate (c) | (a) 5 min | (b) 2 h | (c) 1 mL·min−1 | ||||
| Fraction 1 (%) | 77 ± 3 | 74 ± 5 | 98.2 ± 0.5 | ||||
| Fraction 2 (%) | 21 ± 1 | 22 ± 3 | 1.6 ± 0.8 | ||||
| Fraction 3 (%) | 3.0 ± 0.4 | 3.8 ± 0.2 | 0.2 ± 0.1 | ||||
| AZA yield (µg/mL) | 2.43 ± 0.09 | 2.4 ± 0.1 | 2.58 ± 0.01 | ||||
| Desorption yield (%) | 83 ± 3 | 81 ± 4 | 88.5 ± 0.2 | ||||
| Total extraction yield (%) | 78 ± 3 | 76 ± 4 | 83.1 ± 0.1 | ||||
| Method No. | Method description | % Recovery of total |
|---|---|---|
| (3) | Algal paste | 56 ± 9 |
| (4) | Algal retentate + HP-20 | 54 ± 3 |
| (5) | Algal permeate + SPATT | 21 ± 9 |
| (6) | Algal permeate + SPE | 26 ± 4 |
2.4. Isolation of AZA1 and -2 from A. spinosum Crude Extract
| Step No. | Step | AZA1 (mg) | AZA2 (mg) | Weight (g) | Purity (%) † |
|---|---|---|---|---|---|
| HP-20 resin extract | 12.5 | 3.2 | 3.04 | 0.5 | |
| 1 | Partitioning | 11.2 | 3.0 | 1.32 | 1.1 |
| 2 | Silica gel | 10.2 | 2.8 | 0.17 | 7.6 |
| 3 | Flash (Phenyl-Hexyl) * | 9.7 | 2.4 | 0.01 | >90 |
| 4 | Prep HPLC (C8/C18) | 9.3 | 2.2 | - | >95 |
| % Recovery (steps 1–4) | 75 | 70 |
3. Experimental Section
3.1. Culture Conditions and Measurement
3.2. A. spinosum Analysis
3.3. Solid Phase Extraction Procedure
3.4. Separation of A. spinosum from the Culture Medium

3.5. Extraction of AZA1 and -2 from the Retentate and Permeate
- - The minimum amount of HP-20 resin required for efficient extraction was evaluated (in triplicate). Resin (1, 2.5, or 5 g) was placed in sonicated retentate (100 mL) for 24 h, recovered, packed in a glass column, and eluted with acetone (3 × 3 times the volume of the HP-20 resin), to give final volumes of 9, 22.5, and 45 mL, respectively.
- - The effect of contact time (2, 6, 24 and 72 h) between the sonicated retentate (100 mL) and the HP-20 resin (2.5 g) was tested in triplicate.
- - The volume of solvent required for elution was determined in triplicate experiments where 5 g batches of HP-20 resin were placed in sonicated retentate (100 mL) for 24 h, recovered, packed in a glass column, and eluted using different volumes of acetone ((a) 3 × 5 mL; (b) 3 × 10 mL; (c) 3 × 25 mL).
- - Using the optimized procedures (2.5 g of resin, 24 h contact), adsorption efficiency was assessed by comparing the initial and final amounts of toxin in the sonicated retentate. The following elution procedures were then tested and the overall AZA recoveries determined (Table 3): (a) desorption using three successive soaks of acetone (3 × 7.5 mL) for 5 min; and (b) for 2 h [53]; (c) elution from a glass column (1 mL·min−1) using three successive additions of acetone (3 × 7.5 mL) [55].
3.6. LC-MS/MS Analysis at Ifremer, Nantes
3.7. Analysis at the Marine Institute, Rinville and at NRCC, Halifax
3.7.1. LC-MS/MS Analysis
3.7.2. NMR Spectroscopy
3.8. AZA1 and -2 Isolation from A. spinosum HP-20 Extract
3.9. Reagents
3.10. Statistical Analysis
4. Conclusions
Supplementary Files
Acknowledgments
References
- McMahon, T.; Silke, J. West coast of Ireland winter toxicity of unknown aetiology in mussels. Harmful Algae News 1996, 14, 2. [Google Scholar]
- Satake, M.; Ofuji, K.; Naoki, H.; James, K.J.; Furey, A.; McMahon, T.; Silke, J.; Yasumoto, T. Azaspiracid, a new marine toxin having unique spiro ring assemblies, isolated from Irish mussels, Mytilus edulis. J. Am. Chem. Soc. 1998, 120, 9967–9968. [Google Scholar]
- Nicolaou, K.C.; Koftis, T.V.; Vyskocil, S.; Petrovic, G.; Tang, W.J.; Frederick, M.O.; Chen, D.Y.K.; Li, Y.W.; Ling, T.T.; Yamada, Y.M.A. Total synthesis and structural elucidation of azaspiracid-1. Final assignment and total synthesis of the correct structure of azaspiracid-1. J. Am. Chem. Soc. 2006, 128, 2859–2872. [Google Scholar]
- Ofuji, K.; Satake, M.; McMahon, T.; Silke, J.; James, K.J.; Naoki, H.; Oshima, Y.; Yasumoto, T. Two analogs of azaspiracid isolated from mussels, Mytilus edulis, involved in human intoxication in Ireland. Nat. Toxins 1999, 7, 99–102. [Google Scholar] [CrossRef]
- Ofuji, K.; Satake, M.; McMahon, T.; James, K.J.; Naoki, H.; Oshima, Y.; Yasumoto, T. Structures of azaspiracid analogs, azaspiracid-4 and azaspiracid-5, causative toxins of azaspiracid poisoning in Europe. Biosci. Biotechnol. Biochem. 2001, 65, 740–742. [Google Scholar] [CrossRef]
- Diaz Sierra, M.; Furey, A.; Hamilton, B.; Lehane, M.; James, K.J. Elucidation of the fragmentation pathways of azaspiracids, using electrospray ionisation, hydrogen/deuterium exchange, and multiple-stage mass spectrometry. J. Mass Spectrom. 2003, 38, 1178–1186. [Google Scholar] [CrossRef]
- James, K.J.; Sierra, M.D.; Lehane, M.; Magdalena, A.B.; Furey, A. Detection of five new hydroxyl analogues of azaspiracids in shellfish using multiple tandem mass spectrometry. Toxicon 2003, 41, 277–283. [Google Scholar] [CrossRef]
- Rehmann, N.; Hess, P.; Quilliam, M.A. Discovery of new analogs of the marine biotoxin azaspiracid in blue mussels (Mytilus edulis) by ultra-performance liquid chromatography/tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2008, 22, 549–558. [Google Scholar] [CrossRef]
- McCarron, P.; Kilcoyne, J.; Miles, C.O.; Hess, P. Formation of azaspiracids-3, -4, -6, and -9 via decarboxylation of carboxyazaspiracid metabolites from shellfish. J. Agric. Food. Chem. 2009, 57, 160–169. [Google Scholar] [CrossRef]
- Krock, B.; Tillmann, U.; John, U.; Cembella, A. LC-MS-MS aboard ship: tandem mass spectrometry in the search for phycotoxins and novel toxigenic plankton from the North Sea. Anal. Bioanal. Chem. 2008, 392, 797–803. [Google Scholar] [CrossRef] [Green Version]
- Krock, B.; Tillmann, U.; John, U.; Cembella, A.D. Characterization of azaspiracids in plankton size-fractions and isolation of an azaspiracid-producing dinoflagellate from the North Sea. Harmful Algae 2009, 8, 254–263. [Google Scholar] [CrossRef]
- Tillmann, U.; Elbrachter, M.; Krock, B.; John, U.; Cembella, A. Azadinium spinosum gen. et sp. nov. (Dinophyceae) identified as a primary producer of azaspiracid toxins. Eur. J. Phycol. 2009, 44, 63–79. [Google Scholar] [CrossRef]
- Salas, R.; Tillmann, U.; John, U.; Kilcoyne, J.; Burson, A.; Cantwell, C.; Hess, P.; Jauffrais, T.; Silke, J. The role of Azadinium spinosum (Dinophyceae) in the production of azaspiracid shellfish poisoning in mussels. Harmful Algae 2011, 10, 774–783. [Google Scholar] [CrossRef]
- Hernandez-Becerril, D.U.; Escobae-Morales, S.; Morreno-Gutiérez, S.P.; Baron-Campis, S.A. Two new Records of Potentially toxic Phytoplankton Species from the Mexican Pacific. In Abstract Book of the 14th International Conference Harmful Algae; Hellenic Centre for Marine Research: Hersonissos, Greece, 2010; p. 137. [Google Scholar]
- Akselman, R.; Negri, R.M. Azadinium spinosum Elbrächter et Tillmann (Dinophyceae) is Present and also caused Blooms at the Southwestern Atlantic. In Abstract Book of the 14th International Conference Harmful Algae; Hellenic Centre for Marine Research: Hersonissos, Greece, 2010; p. 132. [Google Scholar]
- Potvin, E.; Jeong, H.J.; Kang, N.S.; Tillmann, U.; Krock, B. First report of the photosynthetic dinoflagellate genus Azadinium in the Pacific Ocean: Morphology and molecular characterization of Azadinium cf. poporum. J. Eukaryot. Microbiol. 2012, 59, 145–156. [Google Scholar] [CrossRef]
- Amzil, Z.; Sibat, M.; Royer, F.; Savar, V. First report on azaspiracid and yessotoxin groups detection in French shellfish. Toxicon 2008, 52, 39–48. [Google Scholar] [CrossRef]
- Vale, P.; Bire, R.; Hess, P. Confirmation by LC-MS/MS of azaspiracids in shellfish from the Portuguese north-western coast. Toxicon 2008, 51, 1449–1456. [Google Scholar] [CrossRef]
- Taleb, H.; Vale, P.; Amanhir, R.; Benhadouch, A.; Sagou, R.; Chafik, A. First detection of azaspiracids in mussels in north west Africa. J. Shellfish Res. 2006, 25, 1067–1070. [Google Scholar]
- James, K.J.; Furey, A.; Lehane, M.; Ramstad, H.; Aune, T.; Hovgaard, P.; Morris, S.; Higman, W.; Satake, M.; Yasumoto, T. First evidence of an extensive northern European distribution of azaspiracid poisoning (AZP) toxins in shellfish. Toxicon 2002, 40, 909–915. [Google Scholar] [CrossRef]
- Ueoka, R.; Ito, A.; Izumikawa, M.; Maeda, S.; Takagi, M.; Shin-Ya, K.; Yoshida, M.; van Soest, R.W.M.; Matsunaga, S. Isolation of azaspiracid-2 from a marine sponge Echinoclathria sp. as a potent cytotoxin. Toxicon 2009, 53, 680–684. [Google Scholar] [CrossRef]
- Alvarez, G.; Uribe, E.; Avalos, P.; Marino, C.; Blanco, J. First identification of azaspiracid and spirolides in Mesodesma donacium and Mulinia edulis from Northern Chile. Toxicon 2010, 55, 638–641. [Google Scholar] [CrossRef]
- Twiner, M.J.; Rehmann, N.; Hess, P.; Doucette, G.J. Azaspiracid shellfish poisoning: A review on the chemistry, ecology, and toxicology with an emphasis on human health impacts. Mar. Drugs 2008, 6, 39–72. [Google Scholar] [CrossRef]
- Tillmann, U.; Elbrachter, M.; John, U.; Krock, B.; Cembella, A. Azadinium obesum (Dinophyceae), a new nontoxic species in the genus that can produce azaspiracid toxins. Phycologia 2010, 49, 169–182. [Google Scholar] [CrossRef]
- Tillmann, U.; Elbrachter, M.; John, U.; Krock, B. A new non-toxic species in the dinoflagellate genus Azadinium: A. poporum sp. nov. Eur. J. Phycol. 2011, 46, 74–87. [Google Scholar] [CrossRef]
- Krock, B.; Tillmann, U.; Voß, D.; Koch, B.P.; Salas, R.; Witt, M.; Potvin, É.; Jeong, H.J. New azaspiracids in Amphidomataceae (Dinophyceae). Toxicon 2012. [Google Scholar] [CrossRef]
- Miles, C.O.; Wilkins, A.L.; Hawkes, A.D.; Jensen, D.J.; Cooney, J.M.; Larsen, K.; Petersen, D.; Rise, F.; Beuzenberg, V.; MacKenzie, A.L. Isolation and identification of a cis-C-8-diol-ester of okadaic acid from Dinophysis acuta in New Zealand. Toxicon 2006, 48, 195–203. [Google Scholar] [CrossRef]
- Abraham, A.; Plakas, S.M.; Wang, Z.H.; Jester, E.L.E.; El Said, K.R.; Granade, H.R.; Henry, M.S.; Blum, P.C.; Pierce, R.H.; Dickey, R.W. Characterization of polar brevetoxin derivatives isolated from Karenia brevis cultures and natural blooms. Toxicon 2006, 48, 104–115. [Google Scholar] [CrossRef]
- Laycock, M.V.; Thibault, P.; Ayer, S.W.; Walter, J.A. Isolation and purification procedures for the preparation of paralytic shellfish poisoning toxin standards. Nat. Toxins 1994, 2, 175–183. [Google Scholar] [CrossRef]
- Loader, J.I.; Hawkes, A.D.; Beuzenberg, V.; Jensen, D.J.; Cooney, J.M.; Wilkins, A.L.; Fitzgerald, J.M.; Briggs, L.R.; Miles, C.O. Convenient large-scale purification of yessotoxin from Protoceratium reticulatum culture and isolation of a novel furanoyessotoxin. J. Agric. Food. Chem. 2007, 55, 11093–11100. [Google Scholar]
- Torigoe, K.; Murata, M.; Yasumoto, T.; Iwashita, T. Prorocentrolide, a toxic nitrogeneous macrocycle from a marine dinoflagellate.Prorocentrum lima. J. Am. Chem. Soc. 1988, 110, 7876–7877. [Google Scholar] [CrossRef]
- Miles, C.O.; Wilkins, A.L.; Stirling, D.J.; MacKenzie, A.L. Gymnodimine C, an isomer of gymnodimine B, from Karenia selliformis. J. Agric. Food. Chem. 2003, 51, 4838–4840. [Google Scholar]
- Selwood, A.I.; Miles, C.O.; Wilkins, A.L.; van Ginkel, R.; Munday, R.; Rise, F.; McNabb, P. Isolation, structural determination and acute toxicity of pinnatoxins E, F and G. J. Agric. Food. Chem. 2010, 58, 6532–6542. [Google Scholar]
- Miles, C.O.; Wilkins, A.L.; Munday, R.; Dines, M.H.; Hawkes, A.D.; Briggs, L.R.; Sandvik, M.; Jensen, D.J.; Cooney, J.M.; Holland, P.T.; et al. Isolation of pectenotoxin-2 from Dinophysis acuta and its conversion to pectenotoxin-2 seco acid, and preliminary assessment of their acute toxicities. Toxicon 2004, 43, 1–9. [Google Scholar] [CrossRef]
- Perez, R.A.; Rehmann, N.; Crain, S.; LeBlanc, P.; Craft, C.; MacKinnon, S.; Reeves, K.; Burton, I.W.; Walter, J.A.; Hess, P.; et al. The preparation of certified calibration solutions for azaspiracid-1,-2, and-3, potent marine biotoxins found in shellfish. Anal. Bioanal. Chem. 2010, 398, 2243–2252. [Google Scholar]
- Kilcoyne, J.; Keogh, A.; Clancy, G.; Leblanc, P.; Burton, I.; Quilliam, M.A.; Hess, P.; Miles, C.O. Improved isolation procedure for azaspiracids from shellfish, structural elucidation of azaspiracid-6, and stability studies. J. Agric. Food. Chem. 2012, 60, 2447–2455. [Google Scholar]
- Rundberget, T.; Sandvik, M.; Larsen, K.; Pizarro, G.M.; Reguera, B.; Castberg, T.; Gustad, E.; Loader, J.I.; Rise, F.; Wilkins, A.L.; Miles, C.O. Extraction of microalgal toxins by large-scale pumping of seawater in Spain and Norway, and isolation of okadaic acid and dinophysistoxin-2. Toxicon 2007, 50, 960–970. [Google Scholar] [CrossRef]
- Hess, P.; McCarron, P.; Quilliam, M.A. Fit-for-purpose shellfish reference materials for internal and external quality control in the analysis of phycotoxins. Anal. Bioanal. Chem. 2007, 387, 2463–2474. [Google Scholar] [CrossRef]
- European Union, Commission Regulation (EU) No 15/2011 of 10 January 2011 Amending Regulation (EC) No 2074/2005 as Regards Recognised Testing Methods for Detecting Marine Biotoxins in Live Bivalve Molluscs; EU: Luxembourg, Luxembourg, 2011; Off. J. Eur. Union L6, 11.1.2011.
- Séchet, V.; Berard, J.B.; Bohec, M.; Bougaran, G.; Carré, C.; Masselin, P.; Truquet, P. Growth and toxicity of Alexandrium catanella isolated from Thau Lagoon (France) Cultured in Stirred Tank Photobioreactors. In Proceeding of the 4th International Conference on Molluscan Shellfish Safety; Villalba, A., Reguera, B., Romalde, J.L., Beiras, R., Eds.; Intergovernmental Oceanographic Commision of UNESCO: Santiago de Compostella, Spain, 2003; pp. 135–144. [Google Scholar]
- Séchet, V.; Berard, J.B.; Bohec, M.; Bougaran, G.; Carré, C.; Masselin, P.; Truquet, P. Continuous-Flow and batch Cultures of Toxic Alexandrium catanella and A. minutum in stirred Tank Bioreactors. In Proceeding of the 10th International Conference on Harmful Algae; Steidinger, K.A., Landsberg, J.H., Tomas, C.R., Vargo, G.A., Eds.; Florida Marine Research Institute, Florida Fish and Wildlife Conservation Commission, Florida Institute of Oceanography, Intergovernmental Oceanographic Commision of UNESCO: St. Pete Beach, FL, USA, 2004; pp. 399–401. [Google Scholar]
- Gallardo Rodriguez, J.J.; Sanchez Miron, A.; Garcia Camacho, F.; Ceron Garcia, M.C.; Belarbi, E.H.; Chisti, Y.; Molina Grima, E. Causes of shear sensitivity of the toxic dinoflagellate Protoceratium reticulatum. Biotechnol. Prog. 2009, 25, 792–800. [Google Scholar] [CrossRef]
- Robinson, J.A. Polyketide synthase complexes: Their structure and function in antibiotic biosynthesis. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1991, 332, 107–114. [Google Scholar] [CrossRef]
- Schembri, M.A.; Neilan, B.A.; Saint, C.P. Identification of genes implicated in toxin production in the cyanobacterium Cylindrospermopsis raciborskii. Environ. Toxicol. 2001, 16, 413–421. [Google Scholar] [CrossRef]
- Medhioub, W.; Sechet, V.; Truquet, P.; Bardouil, M.; Amzil, Z.; Lassus, P.; Soudant, P. Alexandrium ostenfeldii growth and spirolide production in batch culture and photobioreactor. Harmful Algae 2011, 10, 794–803. [Google Scholar] [CrossRef]
- Nagai, S.; Suzuki, T.; Nishikawa, T.; Kamiyama, T. Differences in the production and excretion kinetics of okadaic acid, dinophysistoxin-1, and pectenotoxin-2 between cultures of Dinophysis acuminata and Dinophysis fortii isolated from western Japan. J. Phycol. 2011, 47, 1326–1337. [Google Scholar] [CrossRef]
- Varkitzi, I.; Pagou, K.; Graneli, E.; Hatzianestis, I.; Pyrgaki, C.; Pavlidou, A.; Montesanto, B.; Economou-Amilli, A. Unbalanced N:P ratios and nutrient stress controlling growth and toxin production of the harmful dinoflagellate Prorocentrum lima (Ehrenberg) Dodge. Harmful Algae 2010, 9, 304–311. [Google Scholar] [CrossRef]
- Medhioub, A.; Medhioub, W.; Amzil, Z.; Sibat, M.; Bardouil, M.; Ben Neila, I.; Mezghani, S.; Hamza, A.; Lassus, P. Influence of environmental parameters on Karenia selliformis toxin content in culture. Cah. Biol. Mar. 2009, 50, 333–342. [Google Scholar]
- Pizarro, G.; Paz, B.; Gonzalez-Gil, S.; Franco, J.M.; Reguera, B. Seasonal variability of lipophilic toxins during a Dinophysis acuta bloom in Western Iberia: Differences between picked cells and plankton concentrates. Harmful Algae 2009, 8, 926–937. [Google Scholar] [CrossRef]
- Parker, N.S.; Negri, A.P.; Frampton, D.M.F.; Rodolfi, L.; Tredici, M.R.; Blackburn, S.I. Growth of the toxic dinoflagellate Alexandrium minutum (Dinophyceae) using high biomass culture systems. J. Appl. Phycol. 2002, 14, 313–324. [Google Scholar] [CrossRef]
- Gallardo Rodriguez, J.J.; Sanchez Miron, A.; Garcia Camacho, F.; Ceron Garcia, M.C.; Belarbi, E.H.; Molina Grima, E. Culture of dinoflagellates in a fed-batch and continuous stirred-tank photobioreactors: Growth, oxidative stress and toxin production. Process Biochem. 2010, 45, 660–666. [Google Scholar] [CrossRef]
- Jauffrais, T.; Herrenknecht, C.; Séchet, V.; Sibat, M.; Tillmann, U.; Krock, B.; Kilcoyne, J.; Miles, C.O.; McCarron, P.; Amzil, Z.; et al. Quantitative analysis of azaspiracids in Azadinium spinosum cultures. Anal. Bioanal. Chem. 2012, 403, 833–846. [Google Scholar] [CrossRef]
- MacKenzie, L.; Beuzenberg, V.; Holland, P.; McNabb, P.; Selwood, A. Solid phase adsorption toxin tracking (SPATT): A new monitoring tool that simulates the biotoxin contamination of filter feeding bivalves. Toxicon 2004, 44, 901–918. [Google Scholar]
- Fux, E. Development and evaluation of passive sampling and LC-MS based techniques for the detection and monitoring of lipophilic marine toxins in mesocosm and field studies. Ph.D. Thesis, Dublin Institute of Technology, Dublin, Ireland, 2008. [Google Scholar]
- Fux, E.; Marcaillou, C.; Mondeguer, F.; Bire, R.; Hess, P. Field and mesocosm trials on passive sampling for the study of adsorption and desorption behaviour of lipophilic toxins with a focus on OA and DTX1. Harmful Algae 2008, 7, 574–583. [Google Scholar] [CrossRef]
- Fux, E.; Bire, R.; Hess, P. Comparative accumulation and composition of lipophilic marine biotoxins in passive samplers and in mussels (M. edulis) on the West Coast of Ireland. Harmful Algae 2009, 8, 523–537. [Google Scholar] [CrossRef]
- MacKenzie, L.A. In situ passive solid-phase adsorption of micro-algal biotoxins as a monitoring tool. Curr. Opin. Biotechnol. 2010, 21, 326–331. [Google Scholar] [CrossRef]
- Alfonso, C.; Rehmann, N.; Hess, P.; Alfonso, A.; Wandscheer, C.B.; Abuin, M.; Vale, C.; Otero, P.; Vieytes, M.R.; Botana, L.M. Evaluation of various pH and temperature conditions on the stability of azaspiracids and their importance in preparative isolation and toxicological studies. Anal. Chem. 2008, 80, 9672–9680. [Google Scholar]
- Keller, M.D.; Selvin, R.C.; Claus, W.; Guillard, R.R.L. Media for the culture of oceanic ultraphytoplankton. J. Phycol. 1987, 23, 633–638. [Google Scholar]
- Quilliam, M. A.; Hess, P.; Dell’Aversano, C. Recent Developments in the Analysis of Phycotoxins by Liquid Chromatography-Mass Spectrometry. In Mycotoxins and Phycotoxins in Perspective at the turn of the Millenium: Proceedings of the Xth International IUPAC Symposium on Mycotoxins and Phycotoxins, Guaruja, Brazil; de Koe, W.J., Sampson, R.A., van Egmond, H.P., Gilbert, J., Sabinoan, M., Eds.; IUPAC: Research Triangle Park, NC, USA, 2001; pp. 383–391. [Google Scholar]
- Jauffrais, T.; Marcaillou, C.; Herrenknecht, C.; Truquet, P.; Séchet, V.; Nicolau, E.; Tillmann, U.; Hess, P. Azaspiracid accumulation, detoxification and biotransformation in blue mussels (Mytilus edulis) experimentally fed Azadinium spinosum. Toxicon 2012. [Google Scholar] [CrossRef]
- Samples Availability: Available from the authors.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jauffrais, T.; Kilcoyne, J.; Séchet, V.; Herrenknecht, C.; Truquet, P.; Hervé, F.; Bérard, J.B.; Nulty, C.; Taylor, S.; Tillmann, U.; et al. Production and Isolation of Azaspiracid-1 and -2 from Azadinium spinosum Culture in Pilot Scale Photobioreactors. Mar. Drugs 2012, 10, 1360-1382. https://doi.org/10.3390/md10061360
Jauffrais T, Kilcoyne J, Séchet V, Herrenknecht C, Truquet P, Hervé F, Bérard JB, Nulty C, Taylor S, Tillmann U, et al. Production and Isolation of Azaspiracid-1 and -2 from Azadinium spinosum Culture in Pilot Scale Photobioreactors. Marine Drugs. 2012; 10(6):1360-1382. https://doi.org/10.3390/md10061360
Chicago/Turabian StyleJauffrais, Thierry, Jane Kilcoyne, Véronique Séchet, Christine Herrenknecht, Philippe Truquet, Fabienne Hervé, Jean Baptiste Bérard, Cíara Nulty, Sarah Taylor, Urban Tillmann, and et al. 2012. "Production and Isolation of Azaspiracid-1 and -2 from Azadinium spinosum Culture in Pilot Scale Photobioreactors" Marine Drugs 10, no. 6: 1360-1382. https://doi.org/10.3390/md10061360