
Fucoidan from Fucus vesiculosus Protects against Alcohol-Induced Liver Damage by Modulating Inflammatory Mediators in Mice and HepG2 Cells


Abstract
:
1. Introduction
2. Results and Discussion
2.1. Changes in Organ Weight in the Presence or Absence of Fucoidan for Alcohol-Fed Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vehicle | 25% (w/v) Alcohol | Fucoidan 30 mg/kg | Fucoidan 60 mg/kg | |||
|---|---|---|---|---|---|---|
| - | Fucoidan 30 mg/kg | Fucoidan 60 mg/kg | ||||
| (n = 6) | (n = 6) | (n = 6) | (n = 6) | (n = 6) | (n = 6) | |
| BW (g) | 32.7 5 ± 0.92 | 34.15 ± 0.73 | 33.60 ± 0.70 | 33.85 ± 1.20 | 34.01 ± 1.00 | 34.45 ± 0.67 |
| Liver (%) | 5.13 ± 0.16 | 5.24 ± 0.11 # | 5.65 ± 0.23 * | 5.52 ± 0.12 * | 5.59 ± 0.05 # | 5.31 ± 0.03 # |
| Spleen (%) | 0.27 ± 0.02 | 0.27 ± 0.09 | 0.27 ± 0.01 | 0.32 ± 0.02 * | 0.35 ± 0.06 # | 0.32 ± 0.01 # |
| Thymus (%) | 0.24 ± 0.01 | 0.21 ± 0.01 # | 0.23 ± 0.09 * | 0.23 ± 0.03 * | 0.24 ± 0.08 # | 0.25 ± 0.13 # |
2.2. Protective Effects of Fucoidan on Serum Aspartate Aminotransferase and Alanine Aminotransferase in Alcohol-Induced Murine Liver Damage

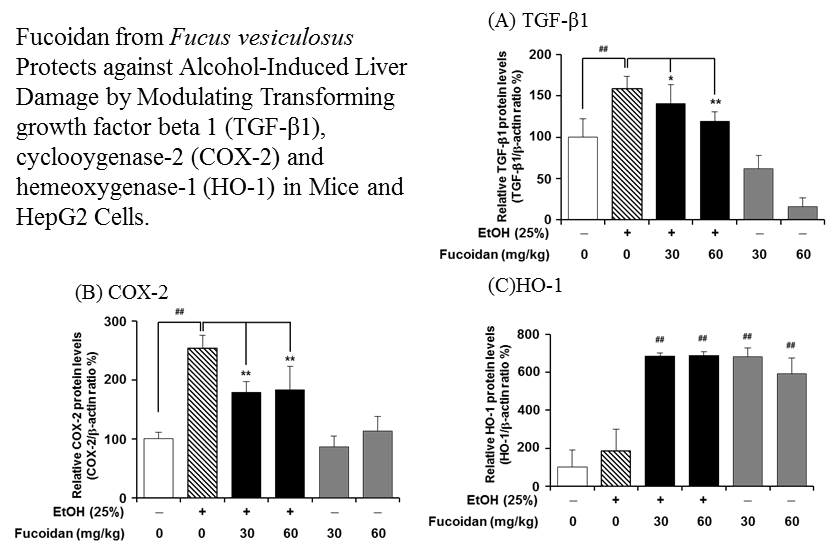
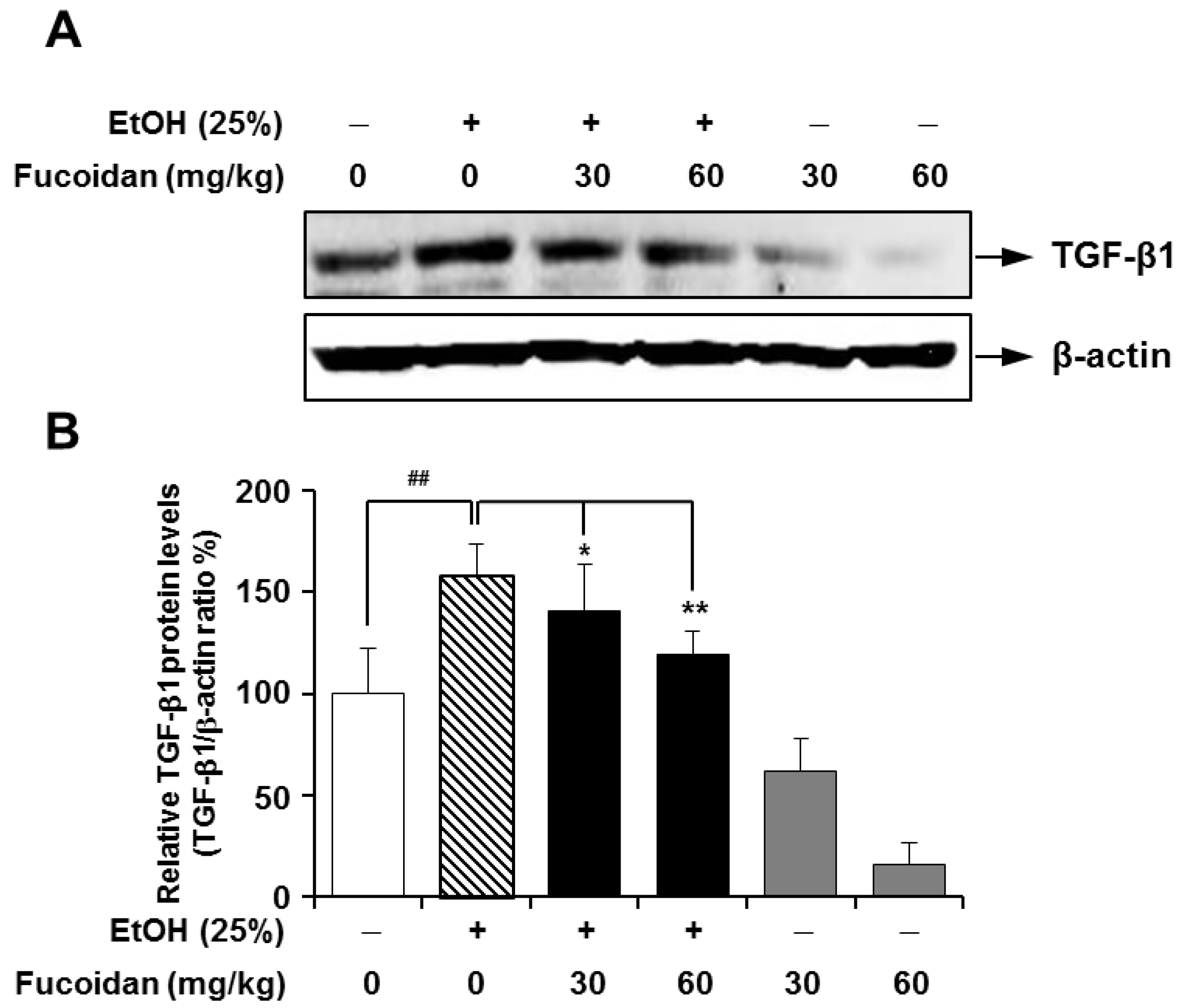
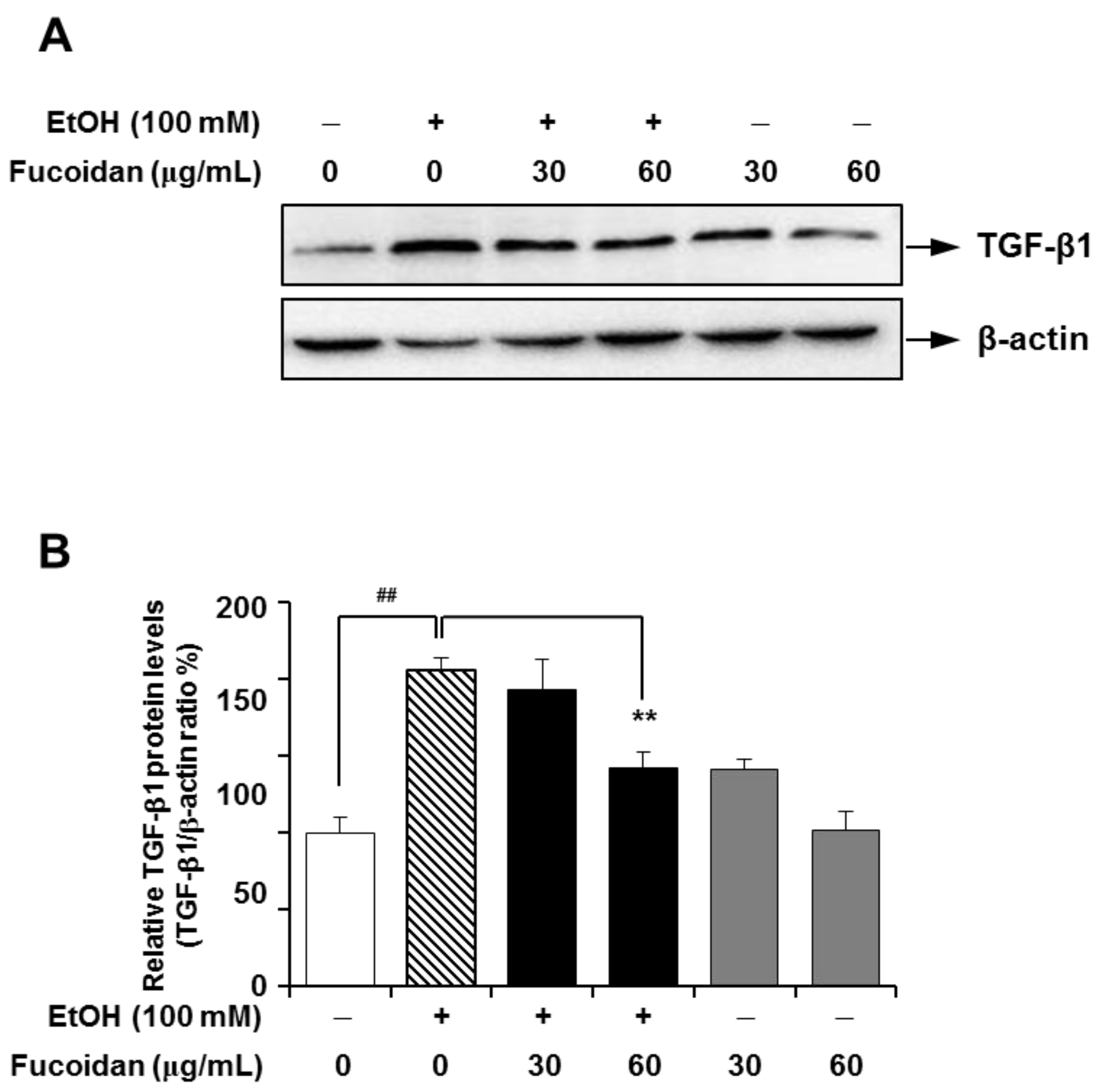
2.3. Fucoidan Suppresses the Alcohol-Induced Expression of TGF-β1 in the Murine Liver and HepG2 Cells


2.4. Fucoidan Suppresses NO Production Following Exposure to Alcohol

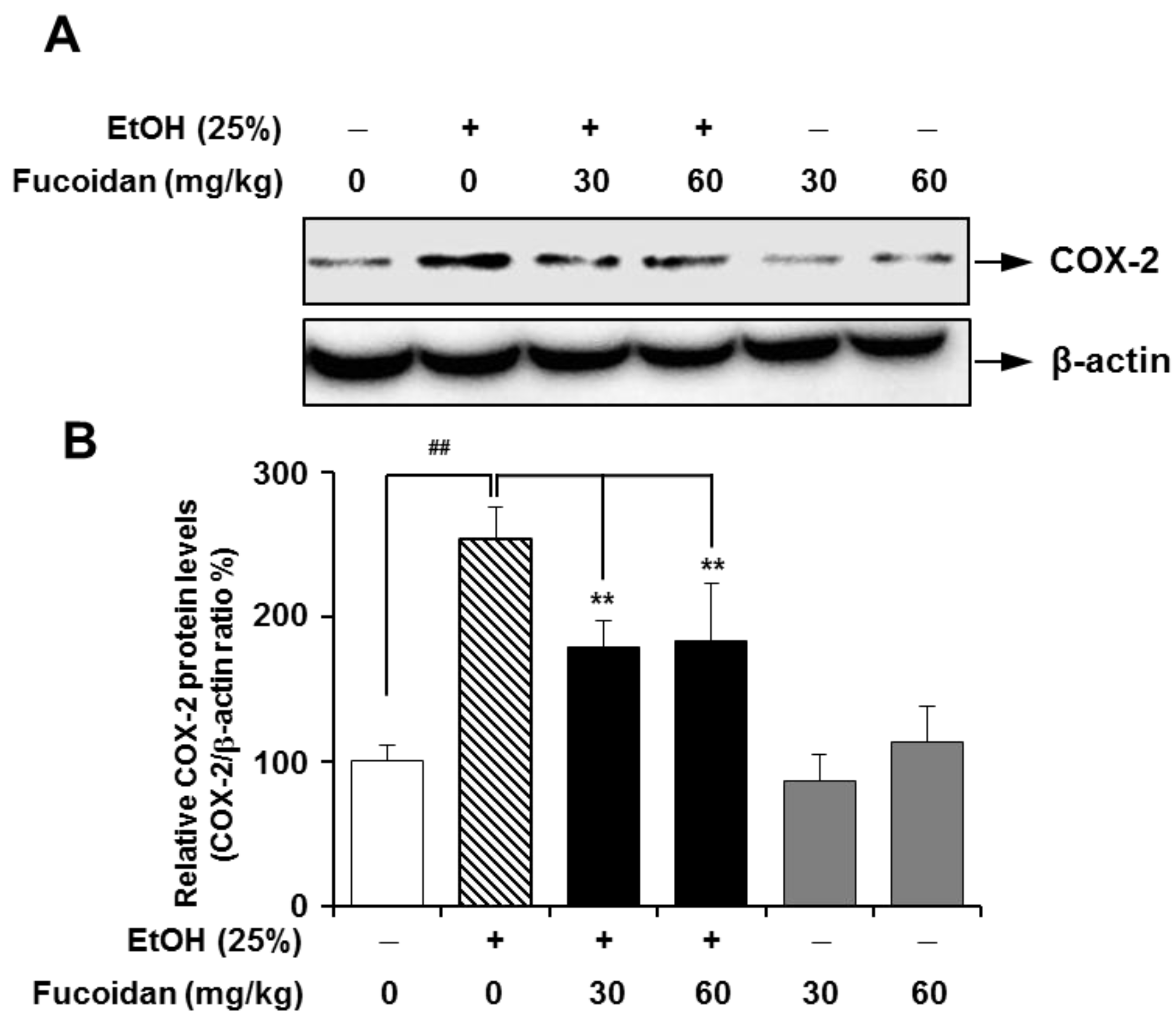
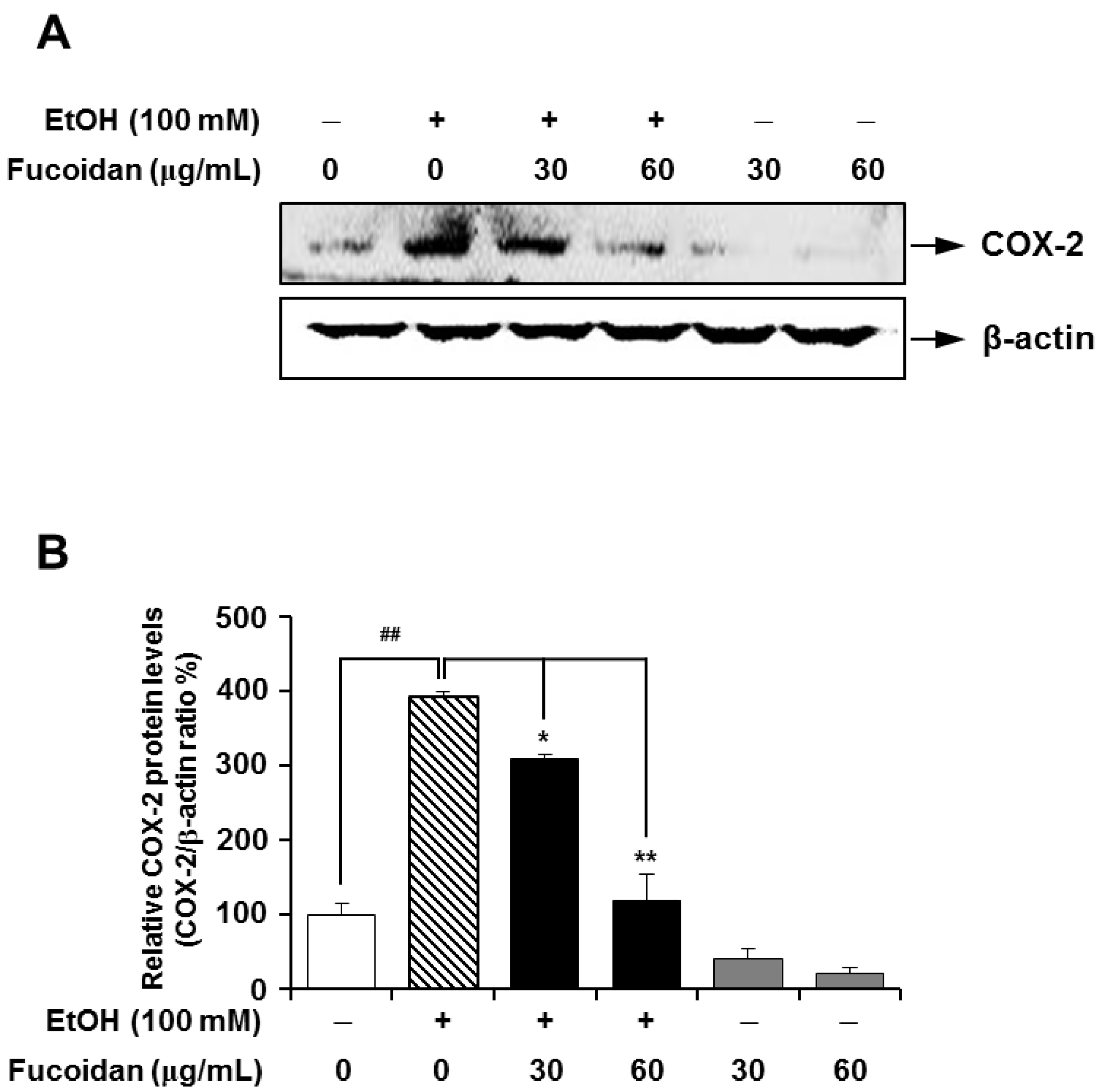
2.5. Fucoidan Suppresses the Expression Level of COX-2


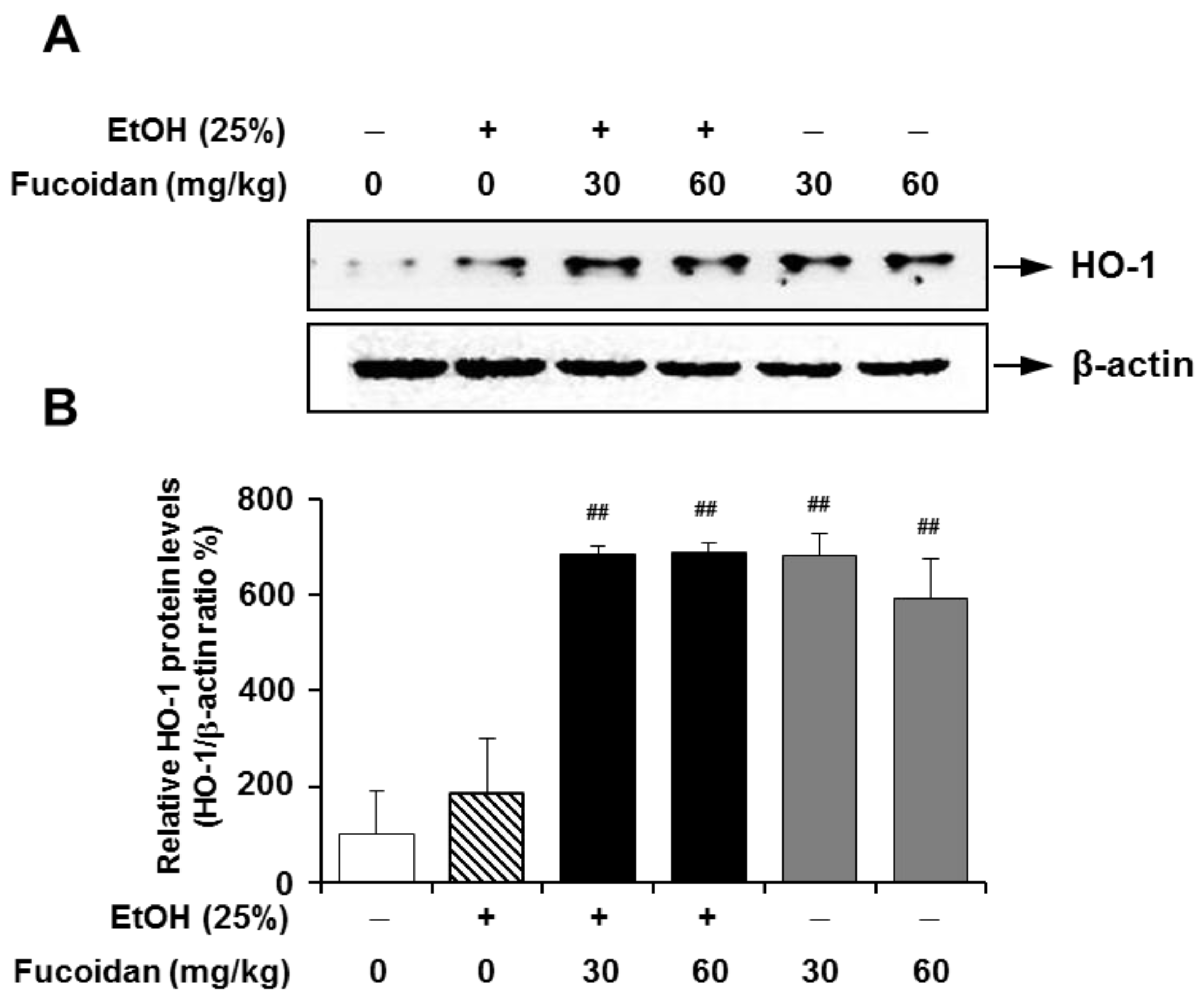
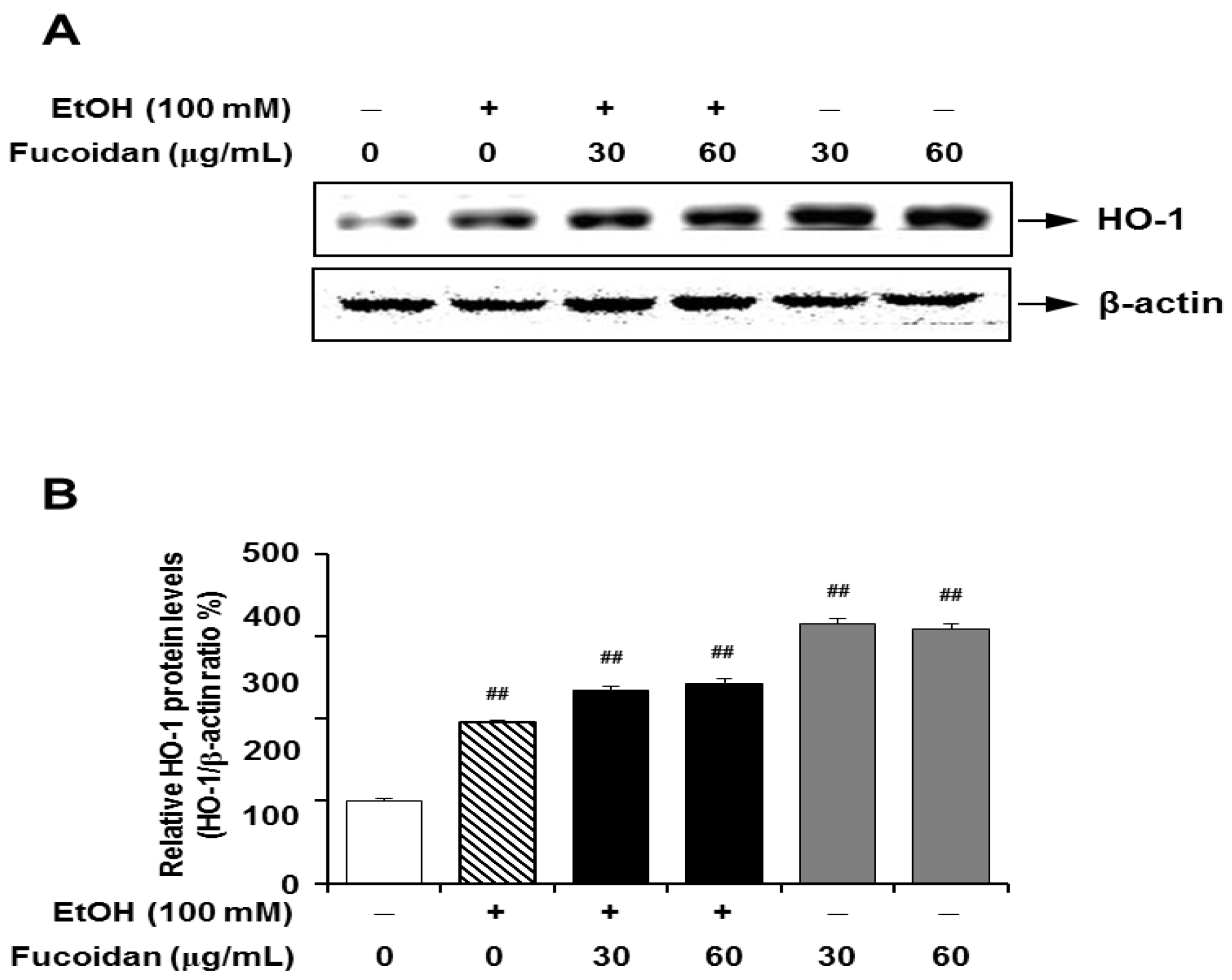
2.6. Fucoidan Increases the Expression Levels of HO-1


3. Experimental Section
3.1. Animals
3.2. Reagents
3.3. Fucoidan Preparation
3.4. Treatment with Alcohol and Fucoidan on Mice and Measurement of Serum ALT, AST and Organ Weight
3.5. HepG2 Cell Culture and Treatment with Alcohol and Fucoidan
3.6. Measurement of Nitric Oxide Production
3.7. Protein Extraction and Immunoblot
3.8. Statistics
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Room, R.; Babor, T.; Rehm, J. Alcohol and public health. Lancet 2005, 365, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Kim, C.H.; Kim, D.J.; Park, J.H.; Kim, T.O.; Yang, S.Y.; Moon, Y.S.; Kim, T.N.; Kim, H.K.; Park, H.Y.; et al. Prevalence of alcoholic liver disease among Korean adults: results from the fourth Korea National Health and Nutrition Examination Survey, 2009. Subst. Use Misuse 2011, 46, 1755–1762. [Google Scholar] [CrossRef] [PubMed]
- Zamora Nava, L.E.; Aguirre Valadez, J.; Chavez-Tapia, N.C.; Torre, A. Acute-on-chronic liver failure: A review. Ther. Clin. Risk Manag. 2014, 10, 295–303. [Google Scholar] [PubMed]
- Friedman, S.L. Liver fibrosis—From bench to bedside. J. Hepatol. 2003, 38 (Suppl. 1), S38–S53. [Google Scholar] [CrossRef] [PubMed]
- Kawaratani, H.; Tsujimoto, T.; Douhara, A.; Takaya, H.; Moriya, K.; Namisaki, T.; Noguchi, R.; Yoshiji, H.; Fujimoto, M.; Fukui, H. The effect of inflammatory cytokines in alcoholic liver disease. Mediators Inflamm. 2013, 2013, 495156. [Google Scholar] [CrossRef] [PubMed]
- Tirapelli, L.F.; Batalhao, M.E.; Jacob-Ferreira, A.L.; Tirapelli, D.P.; Carnio, E.C.; Tanus-Santos, J.E.; Queiroz, R.H.; Uyemura, S.A.; Padovan, C.M.; Tirapelli, C.R. Chronic ethanol consumption induces histopathological changes and increases nitric oxide generation in the rat liver. Tissue Cell 2011, 43, 384–391. [Google Scholar] [CrossRef] [PubMed]
- De Bleser, P.J.; Niki, T.; Rogiers, V.; Geerts, A. Transforming growth factor-beta gene expression in normal and fibrotic rat liver. J. Hepatol. 1997, 26, 886–893. [Google Scholar] [CrossRef] [PubMed]
- Baghy, K.; Iozzo, R.V.; Kovalszky, I. Decorin-TGFbeta axis in hepatic fibrosis and cirrhosis. J. Histochem. Cytochem. 2012, 60, 262–268. [Google Scholar] [PubMed]
- Kyung-Oh, D. Effects of TGF-beta1 Ribbon Antisense on CCl(4)-induced Liver Fibrosis. Korean J. Physiol. Pharmacol. 2008, 12, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.S.; Kim, S.J.; Kwon do, Y.; Ahn, C.W.; Kim, Y.S.; Choi, D.W.; Kim, Y.C. Alleviation of alcoholic liver injury by betaine involves an enhancement of antioxidant defense via regulation of sulfur amino acid metabolism. Food Chem. Toxicol. 2013, 62, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Ganey, P.E.; Barton, Y.W.; Kinser, S.; Sneed, R.A.; Barton, C.C.; Roth, R.A. Involvement of cyclooxygenase-2 in the potentiation of allyl alcohol-induced liver injury by bacterial lipopolysaccharide. Toxicol. Appl. Pharmacol. 2001, 174, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.W.; Jang, J.Y.; Lee, S.H.; Kim, S.G.; Cheon, Y.K.; Kim, Y.S.; Cho, Y.D.; Kim, H.S.; Lee, J.S.; Jin, S.Y.; et al. Increased expression of cyclooxygenase-2 is associated with the progression to cirrhosis. Korean J. Intern. Med. 2010, 25, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Baraona, E.; Zeballos, G.A.; Shoichet, L.; Mak, K.M.; Lieber, C.S. Ethanol consumption increases nitric oxide production in rats, and its peroxynitrite-mediated toxicity is attenuated by polyenylphosphatidylcholine. Alcohol. Clin. Exp. Res. 2002, 26, 883–889. [Google Scholar] [CrossRef] [PubMed]
- Morse, D.; Lin, L.; Choi, A.M.; Ryter, S.W. Heme oxygenase-1, a critical arbitrator of cell death pathways in lung injury and disease. Free Radic Biol. Med. 2009, 47, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Bae, J.U.; Hong, K.W.; Kim, C.D. HO-1 Induced by Cilostazol Protects Against TNF-alpha-associated Cytotoxicity via a PPAR-gamma-dependent Pathway in Human Endothelial Cells. Korean J. Physiol. Pharmacol. 2011, 15, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Clement, M.J.; Tissot, B.; Chevolot, L.; Adjadj, E.; Du, Y.; Curmi, P.A.; Daniel, R. NMR characterization and molecular modeling of fucoidan showing the importance of oligosaccharide branching in its anticomplementary activity. Glycobiology 2010, 20, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D’Incecco, A.; Piccoli, A.; Totani, L.; Tinari, N.; Morozevich, G.E.; Berman, A.E.; Bilan, M.I.; et al. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Fitton, J.H. Therapies from fucoidan; multifunctional marine polymers. Mar. Drugs 2011, 9, 1731–1760. [Google Scholar] [CrossRef] [PubMed]
- Nakazato, K.; Takada, H.; Iha, M.; Nagamine, T. Attenuation of N-nitrosodiethylamine-induced liver fibrosis by high-molecular-weight fucoidan derived from Cladosiphon okamuranus. J. Gastroenterol. Hepatol. 2010, 25, 1692–1701. [Google Scholar] [CrossRef] [PubMed]
- Senni, K.; Gueniche, F.; Foucault-Bertaud, A.; Igondjo-Tchen, S.; Fioretti, F.; Colliec-Jouault, S.; Durand, P.; Guezennec, J.; Godeau, G.; Letourneur, D. Fucoidan a sulfated polysaccharide from brown algae is a potent modulator of connective tissue proteolysis. Arch. Biochem. Biophys. 2006, 445, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Thring, T.S.; Hili, P.; Naughton, D.P. Anti-collagenase, anti-elastase and anti-oxidant activities of extracts from 21 plants. BMC Complement. Altern. Med. 2009, 9, 27. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, S.; Itoh, A.; Isoda, K.; Kondoh, M.; Kawase, M.; Yagi, K. Fucoidan partly prevents CCl4-induced liver fibrosis. Eur. J. Pharmacol. 2008, 580, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Saito, A.; Yoneda, M.; Yokohama, S.; Okada, M.; Haneda, M.; Nakamura, K. Fucoidan prevents concanavalin A-induced liver injury through induction of endogenous IL-10 in mice. Hepatol. Res. 2006, 35, 190–198. [Google Scholar] [PubMed]
- Meenakshi, S.; Umayaparvathi, S.; Saravanan, R.; Manivasagam, T.; Balasubramanian, T. Hepatoprotective effect of fucoidan isolated from the seaweed Turbinaria decurrens in ethanol intoxicated rats. Int. J. Biol. Macromol. 2014, 67, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Pruett, S.B.; Han, Y.C.; Wu, W.J. A brief review of immunomodulation caused by acute administration of ethanol: Involvement of neuroendocrine pathways. Alcohol. Alcohol. Suppl. 1994, 2, 431–437. [Google Scholar] [PubMed]
- Wang, J.F.; Spitzer, J.J. Alcohol-induced thymocyte apoptosis is accompanied by impaired mitochondrial function. Alcohol 1997, 14, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Mathews, S.; Xu, M.; Wang, H.; Bertola, A.; Gao, B. Animal models of alcohol-induced liver disease: Pathophysiology, translational relevance and challenges. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 306, G819–G823. [Google Scholar] [CrossRef] [PubMed]
- Castilla, A.; Prieto, J.; Fausto, N. Transforming growth factors beta 1 and alpha in chronic liver disease. Effects of interferon alfa therapy. N. Engl. J. Med. 1991, 324, 933–940. [Google Scholar] [CrossRef] [PubMed]
- Shek, F.W.; Benyon, R.C. How can transforming growth factor beta be targeted usefully to combat liver fibrosis? Eur. J. Gastroenterol. Hepatol. 2004, 16, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.J.; Xu, D.; Guo, Y.; Ping, J.; Chen, L.B.; Wang, H. Protection by and anti-oxidant mechanism of berberine against rat liver fibrosis induced by multiple hepatotoxic factors. Clin. Exp. Pharmacol. Physiol. 2008, 35, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.S.; Deitrich, R.A. Ethanol metabolism and effects: nitric oxide and its interaction. Curr. Clin. Pharmacol. 2007, 2, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Nanji, A.A.; Miao, L.; Thomas, P.; Rahemtulla, A.; Khwaja, S.; Zhao, S.; Peters, D.; Tahan, S.R.; Dannenberg, A.J. Enhanced cyclooxygenase-2 gene expression in alcoholic liver disease in the rat. Gastroenterology 1997, 112, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Reimers, E.; Santolaria-Fernandez, F.; Martin-Gonzalez, M.C.; Fernandez-Rodriguez, C.M.; Quintero-Platt, G. Alcoholism: A systemic proinflammatory condition. World J. Gastroenterol. 2014, 20, 14660–14671. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.S.; Kim, I.D.; Kwon, R.H.; Lee, J.Y.; Kang, J.S.; Ha, B.J. The effects of fucoidan extracts on CCl(4)-induced liver injury. Arch. Pharm. Res. 2008, 31, 622–627. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.W.; Jung, K.H.; Lee, H.S.; Zheng, H.M.; Choi, M.J.; Lee, C.; Hong, S.S. Suppression by fucoidan of liver fibrogenesis via the TGF-beta/Smad pathway in protecting against oxidative stress. Biosci. Biotechnol. Biochem. 2011, 75, 833–840. [Google Scholar] [CrossRef] [PubMed]
- Bakhautdin, B.; Das, D.; Mandal, P.; Roychowdhury, S.; Danner, J.; Bush, K.; Pollard, K.; Kaspar, J.W.; Li, W.; Salomon, R.G.; et al. Protective role of HO-1 and carbon monoxide in ethanol-induced hepatocyte cell death and liver injury in mice. J. Hepatol. 2014, 61, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Li, J.P.; Gao, Y.; Chu, S.F.; Zhang, Z.; Xia, C.Y.; Mou, Z.; Song, X.Y.; He, W.B.; Guo, X.F.; Chen, N.H. Nrf2 pathway activation contributes to anti-fibrosis effects of ginsenoside Rg1 in a rat model of alcohol- and CCl4-induced hepatic fibrosis. Acta Pharmacol. Sin. 2014, 35, 1031–1044. [Google Scholar] [CrossRef] [PubMed]
- Zeng, T.; Zhang, C.L.; Song, F.Y.; Zhao, X.L.; Yu, L.H.; Zhu, Z.P.; Xie, K.Q. The activation of HO-1/Nrf-2 contributes to the protective effects of diallyl disulfide (DADS) against ethanol-induced oxidative stress. Biochim. Biophys. Acta 2013, 1830, 4848–4859. [Google Scholar] [CrossRef] [PubMed]
- Tsoyi, K.; Kim, H.J.; Shin, J.S.; Kim, D.H.; Cho, H.J.; Lee, S.S.; Ahn, S.K.; Yun-Choi, H.S.; Lee, J.H.; Seo, H.G.; et al. HO-1 and JAK-2/STAT-1 signals are involved in preferential inhibition of iNOS over COX-2 gene expression by newly synthesized tetrahydroisoquinoline alkaloid, CKD712, in cells activated with lipopolysacchride. Cell Signal 2008, 20, 1839–1847. [Google Scholar] [CrossRef] [PubMed]
- Mandal, P.; Pritchard, M.T.; Nagy, L.E. Anti-inflammatory pathways and alcoholic liver disease: Role of an adiponectin/interleukin-10/heme oxygenase-1 pathway. World J. Gastroenterol. 2010, 16, 1330–1336. [Google Scholar] [CrossRef] [PubMed]
- Haschemi, A.; Wagner, O.; Marculescu, R.; Wegiel, B.; Robson, S.C.; Gagliani, N.; Gallo, D.; Chen, J.F.; Bach, F.H.; Otterbein, L.E. Cross-regulation of carbon monoxide and the adenosine A2a receptor in macrophages. J. Immunol. 2007, 178, 5921–5929. [Google Scholar] [CrossRef] [PubMed]
- Nishino, T.; Nishioka, C.; Ura, H.; Nagumo, T. Isolation and partial characterization of a novel amino sugar-containing fucan sulfate from commercial Fucus vesiculosus fucoidan. Carbohydr. Res. 1994, 255, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Do, H.; Kang, N.S.; Pyo, S.; Billiar, T.R.; Sohn, E.H. Differential regulation by fucoidan of IFN-gamma-induced NO production in glial cells and macrophages. J. Cell Biochem. 2010, 111, 1337–1345. [Google Scholar] [CrossRef] [PubMed]
- Harrison-Findik, D.D.; Schafer, D.; Klein, E.; Timchenko, N.A.; Kulaksiz, H.; Clemens, D.; Fein, E.; Andriopoulos, B.; Pantopoulos, K.; Gollan, J. Alcohol metabolism-mediated oxidative stress down-regulates hepcidin transcription and leads to increased duodenal iron transporter expression. J. Biol. Chem. 2006, 281, 22974–22982. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.R.; Kwak, J.H.; Noh, S.J.; Pronto, J.R.; Ko, K.S.; Rhee, B.D.; Xu, Z.; Kim, N.; Han, J. Kobophenol A Inhibits Sodium Nitroprusside-Induced Cardiac H9c2 Cell Death through Suppressing Activation of JNK and Preserving Mitochondrial Anti-apoptotic Bcl-2 and Mcl-1. Chem. Pharm. Bull. (Tokyo) 2014, 62, 713–718. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, J.D.; Lee, S.R.; Kim, T.; Jang, S.-A.; Kang, S.C.; Koo, H.J.; Sohn, E.; Bak, J.P.; Namkoong, S.; Kim, H.K.; et al. Fucoidan from Fucus vesiculosus Protects against Alcohol-Induced Liver Damage by Modulating Inflammatory Mediators in Mice and HepG2 Cells. Mar. Drugs 2015, 13, 1051-1067. https://doi.org/10.3390/md13021051
Lim JD, Lee SR, Kim T, Jang S-A, Kang SC, Koo HJ, Sohn E, Bak JP, Namkoong S, Kim HK, et al. Fucoidan from Fucus vesiculosus Protects against Alcohol-Induced Liver Damage by Modulating Inflammatory Mediators in Mice and HepG2 Cells. Marine Drugs. 2015; 13(2):1051-1067. https://doi.org/10.3390/md13021051
Chicago/Turabian StyleLim, Jung Dae, Sung Ryul Lee, Taeseong Kim, Seon-A Jang, Se Chan Kang, Hyun Jung Koo, Eunsoo Sohn, Jong Phil Bak, Seung Namkoong, Hyoung Kyu Kim, and et al. 2015. "Fucoidan from Fucus vesiculosus Protects against Alcohol-Induced Liver Damage by Modulating Inflammatory Mediators in Mice and HepG2 Cells" Marine Drugs 13, no. 2: 1051-1067. https://doi.org/10.3390/md13021051
APA StyleLim, J. D., Lee, S. R., Kim, T., Jang, S.-A., Kang, S. C., Koo, H. J., Sohn, E., Bak, J. P., Namkoong, S., Kim, H. K., Song, I. S., Kim, N., Sohn, E.-H., & Han, J. (2015). Fucoidan from Fucus vesiculosus Protects against Alcohol-Induced Liver Damage by Modulating Inflammatory Mediators in Mice and HepG2 Cells. Marine Drugs, 13(2), 1051-1067. https://doi.org/10.3390/md13021051