
Natural Marine and Synthetic Xenobiotics Get on Nematode’s Nerves: Neuro-Stimulating and Neurotoxic Findings in Caenorhabditis elegans

Abstract
:1. Introduction
2. Results
2.1. Acute Exposure
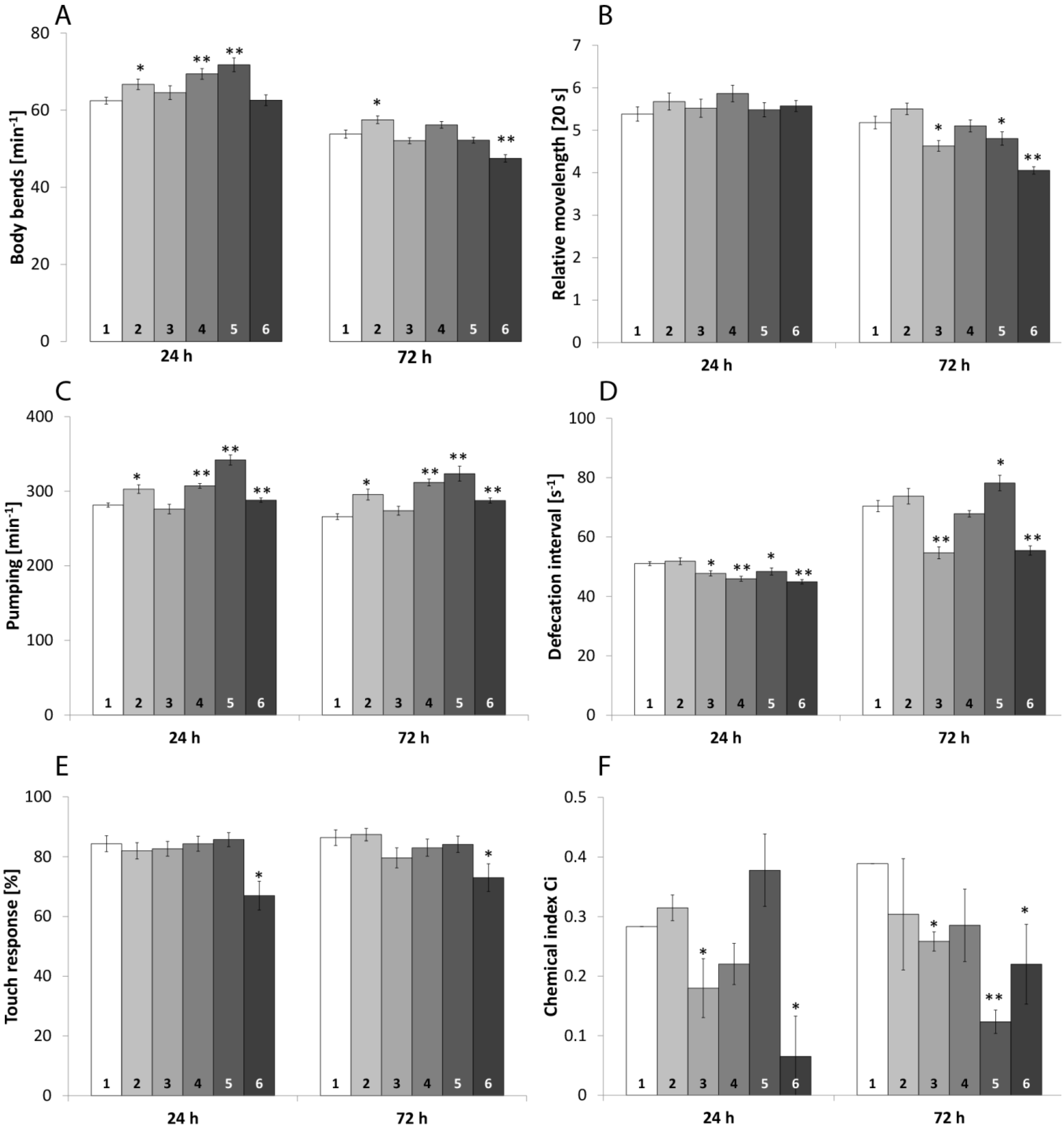
2.1.1. Locomotive Behavior
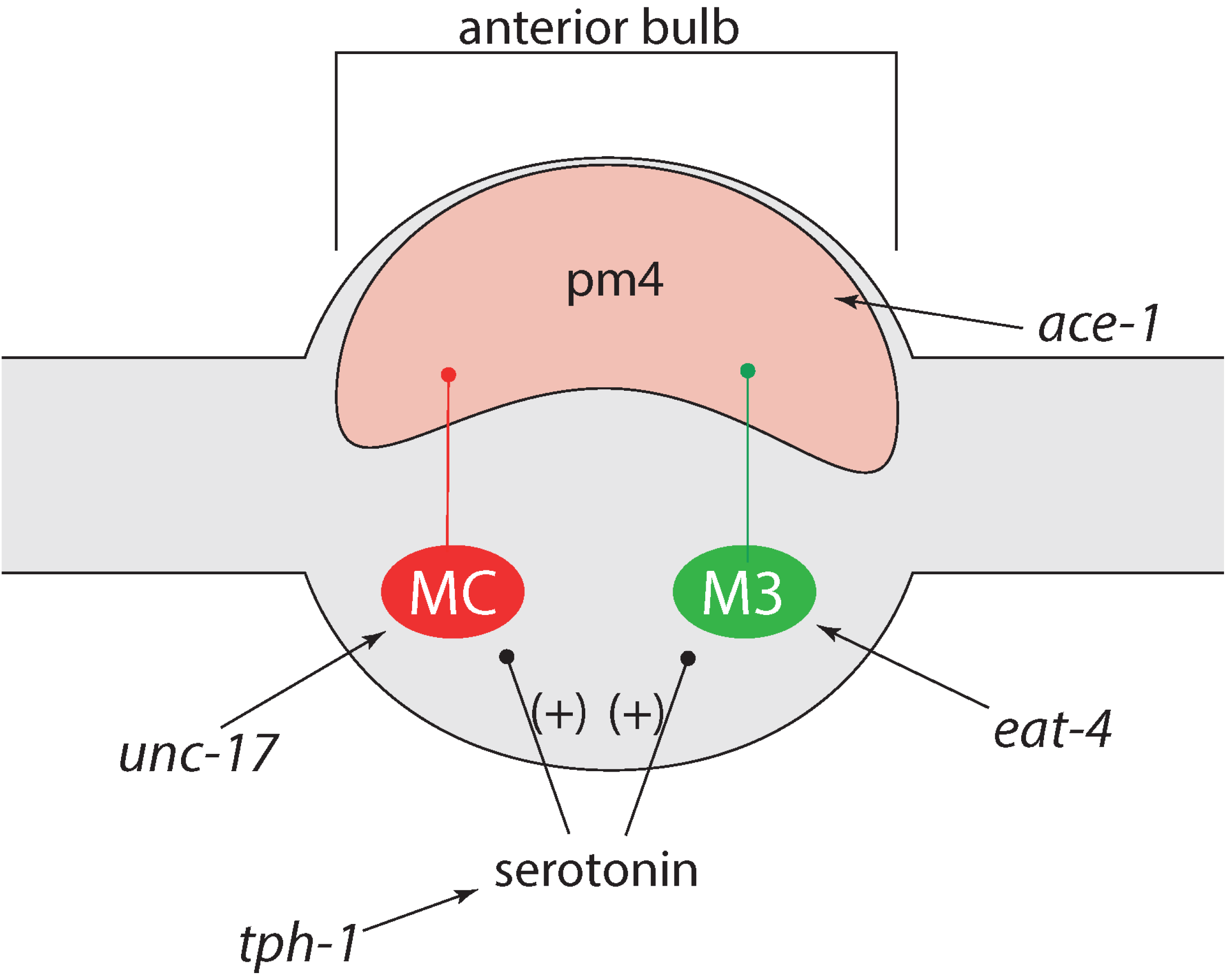
2.1.2. Pharynx Pumping


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
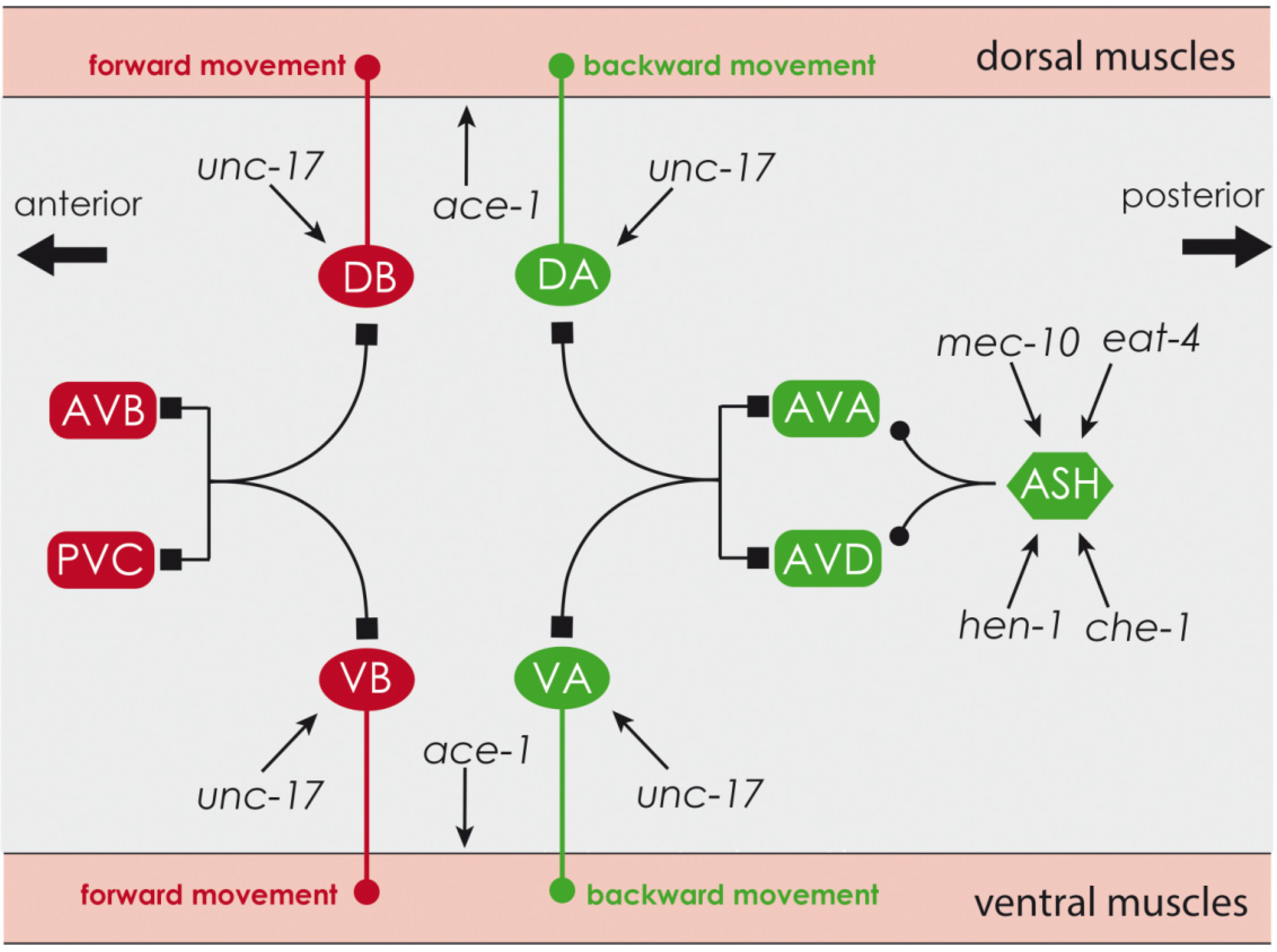{kind=link}
| Neurophysiological Behavior | tex | 0.1 T + 50 D Relative to | 50 T + 50 D Relative to | ||
|---|---|---|---|---|---|
| 0.1 T | 50 D | 50 T | 50 D | ||
| Body bends | 24 h | * | - | - | * |
| 72 h | ** | * | ** | ** | |
| Relative movelength | 24 h | - | - | - | - |
| 72 h | * | - | ** | ** | |
| Pumping | 24 h | * | ** | * | * |
| 72 h | * | - | - | ** | |
| Defecation | 24 h | * | - | * | - |
| 72 h | - | * | - | ** | |
| Touch response | 24 h | - | - | * | * |
| 72 h | - | - | - | - | |
| Chemical index | 24 h | - | - | - | - |
| 72 h | - | * | - | - | |
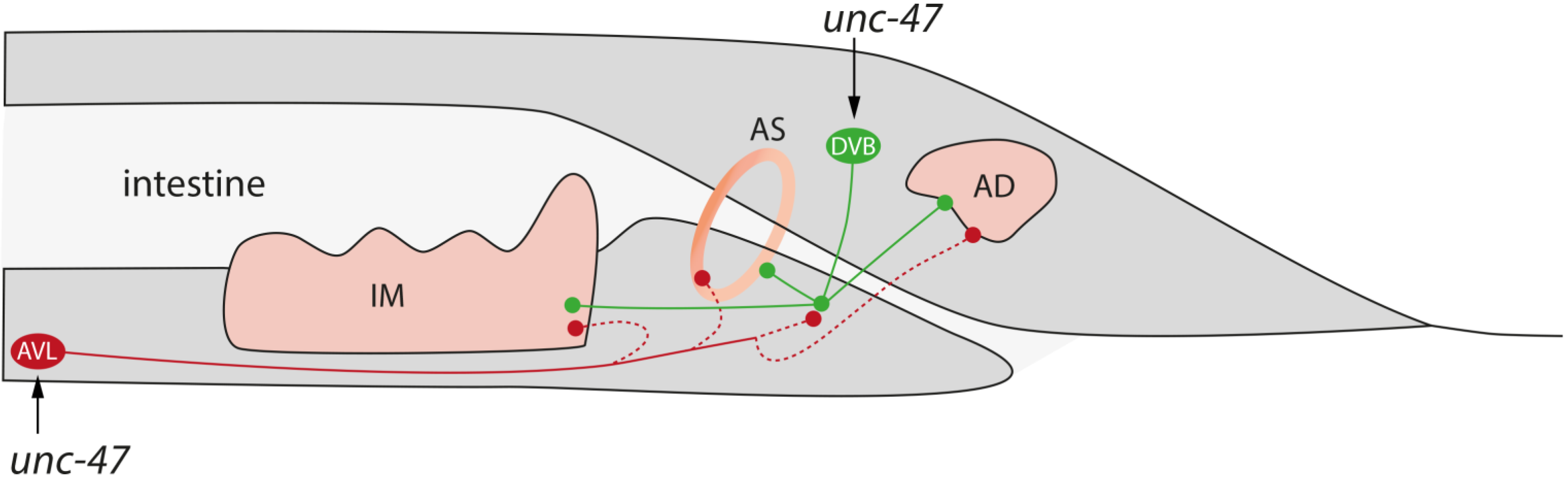
2.1.3. Defecation
2.1.4. Mechanical Sensory Perception
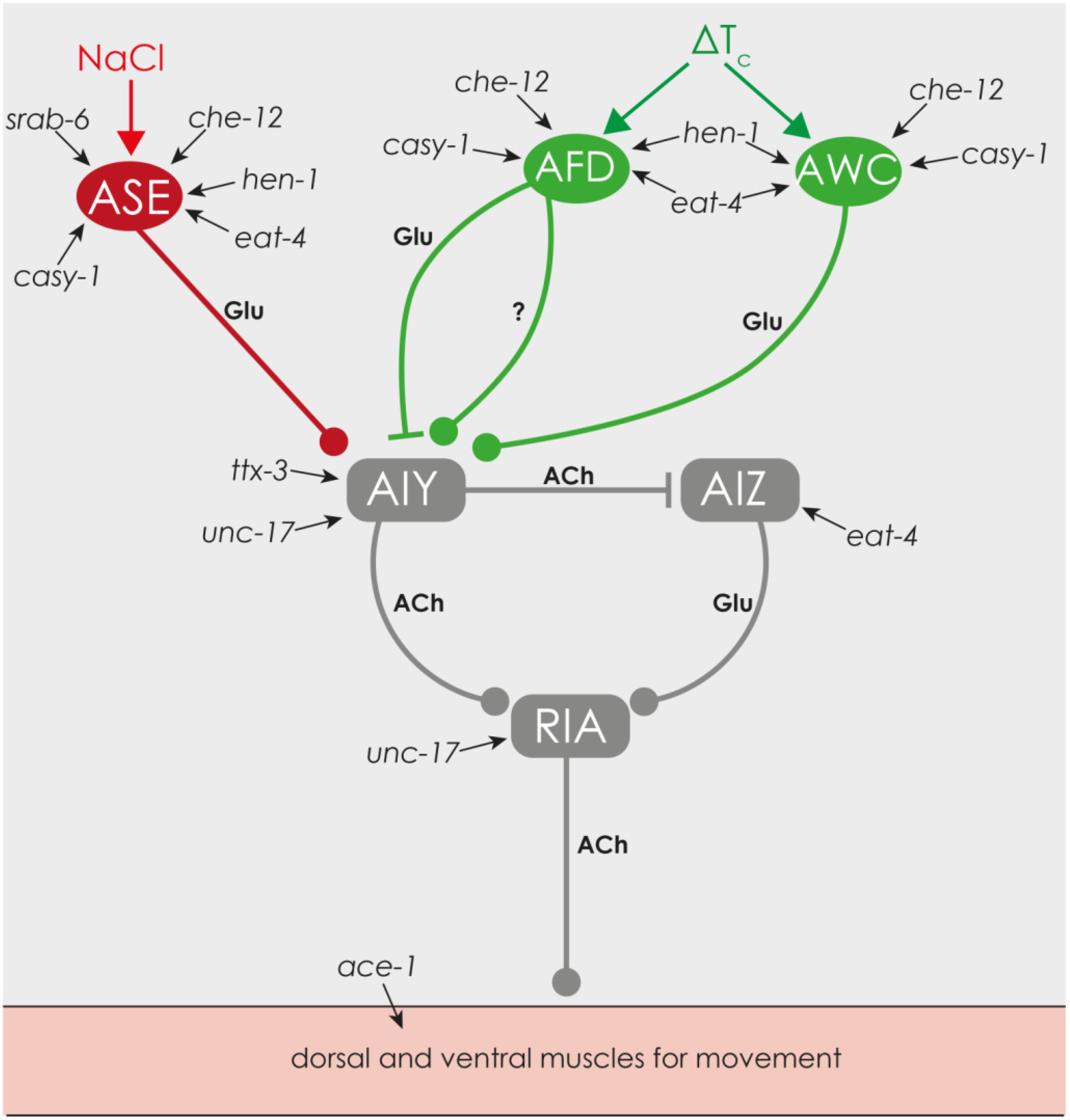
2.1.5. Chemical and Thermal Sensory Perception
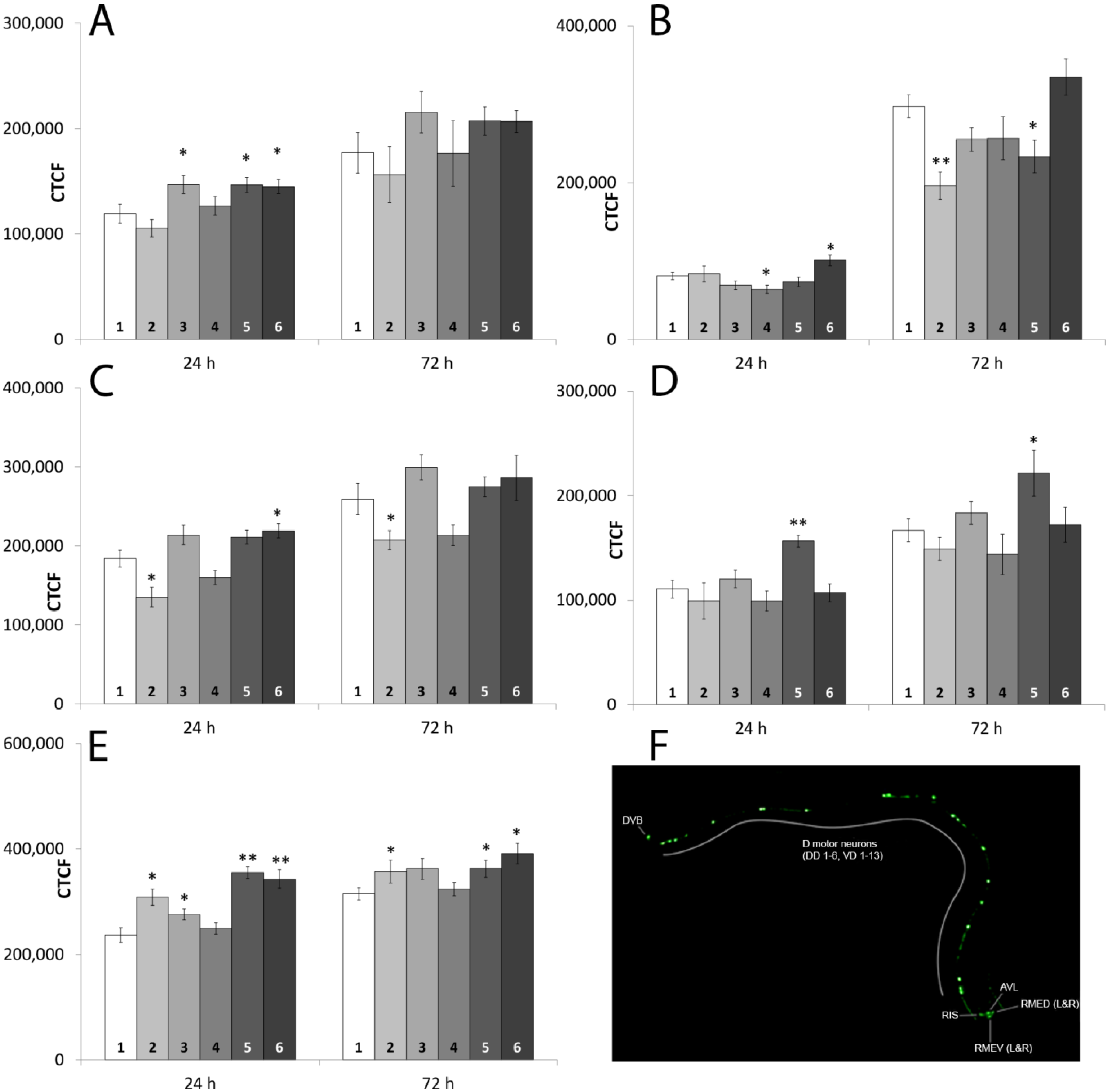
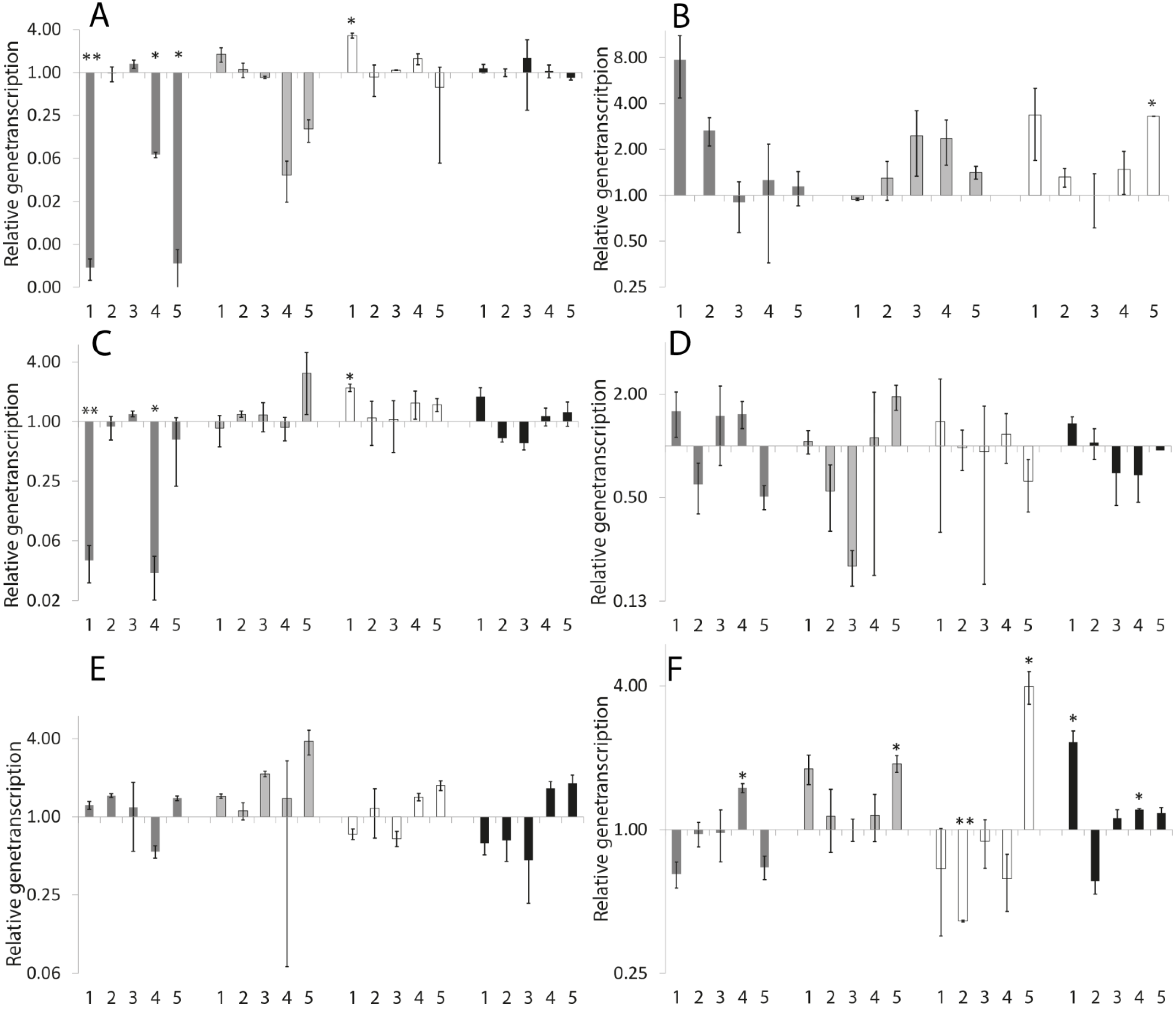
2.1.6. Neuromolecular Experiments
| Gene | Function |
|---|---|
| ace-1 (abnormal AcetylCholinEsterase) | Acetylcholinesterase |
| casy-1 (CAlSYntenin/Alcadein homolog) | Learning |
| che-12 (abnormal CHEmotaxis) | Chemorezeptor |
| dat-1 (DopAmine Transporter) | Dopamine-transporter |
| dop-1 (DOPamine receptor) | D1-dopamine-receptor |
| dop-3 (DOPamine receptor) | D2- dopamine-receptor |
| eat-4 (EATing: abnormal pharyngeal pumping) | Vesicular glutamate transporter |
| gbb-1 (GABAB receptor subunit) | GABA-receptor |
| grd-12 (GRounDhog (hedgehog-like family)) | Growth and movement |
| hda-4 (Histone DeAcetylase) | Histone deacetylase |
| hen-1 (HEsitatioN behavior) | Integration of sensory stimuli |
| mec-10 (MEChanosensory abnormality) | Mechanical stimulation |
| srab-6 (Serpentine Receptor, class AB (class A-like)) | Needed for chemotaxis |
| tph-1 (TryPtophan Hydroxylase) | Serotonin synthesis |
| ttx-3 (abnormal ThermoTaXis) | For AIY interneurons |
| unc-17 (UNCoordinated) | Vesicular acetylcholine transporter |
| unc-47 (UNCoordinated) | Vesicular GABA transporter |
| Gene | 0.1 µM TBBP-A | 50 µM TBBP-A | 50 µM DBAA | 0.1 µM TBBP-A + 50 µM DBAA | 50 µM TBBP-A + 50 µM DBAA |
|---|---|---|---|---|---|
| mean ± SEM | mean ± SEM | mean ± SEM | mean ± SEM | mean ± SEM | |
| ace-1 | 1.52 ± 0.66 | 4.02 ± 1.67 | 2.97 ± 1.34 | 1.28 ± 0.82 | 4.02 ± 2.21 |
| casy-1 | 0.41 ** ± 0.13 | 1.75 * ± 0.32 | 0.73 ± 0.23 | 4.22 * ± 1.24 | 2.14 * ± 0.35 |
| che-12 | 0.82 ± 0.23 | 1.33 ± 0.22 | 0.49 * ± 0.12 | 2.12 ** ± 0.21 | 0.99 ± 0.21 |
| dat-1 | 0.59 ± 0.20 | 2.37 ± 0.73 | 1.20 ± 0.22 | 2.32 * ± 0.31 | 2.09 ** ± 0.14 |
| dop-1 | 1.31 ± 0.46 | 3.06 ± 1.02 | 0.56 * ± 0.23 | 1.08 ± 0.33 | 1.11 ± 0.24 |
| dop-3 | 1.09 ± 0.24 | 2.70 * ± 0.73 | 0.49 * ± 0.18 | 2.09 ** ± 0.17 | 1.62 * ± 0.20 |
| eat-4 | 0.86 ± 0.29 | 0.64 * ± 0.18 | 0.40 ** ± 0.08 | 0.63 * ± 0.13 | 4.51 * ± 1.42 |
| gbb-1 | 0.61 * ± 0.12 | 1.27 ± 0.21 | 0.49 * ± 0.11 | 1.48 * ± 0.22 | 1.20 ± 0.16 |
| grd-12 | 0.47 * ± 0.16 | 0.75 ± 0.20 | 0.65 * ± 0.15 | 0.68 ** ± 0.07 | 0.70 ± 0.34 |
| hda-4 | 1.15 ± 0.13 | 1.14 * ± 0.07 | 1.23 ± 0.15 | 1.53 * ± 0.19 | 1.26* ± 0.09 |
| hen-1 | 2.01 ± 0.69 | 2.56 * ± 0.57 | 3.59 * ± 1.20 | 3.02 * ± 0.89 | 0.50 * ± 0.13 |
| mec-10 | 0.95 ± 0.09 | 1.25 * ± 0.07 | 1.05 ± 0.10 | 2.29 ** ± 0.14 | 0.86 * ± 0.07 |
| srab-6 | 0.78 *± 0.08 | 1.07 ± 0.11 | 1.83 * ± 0.28 | 2.17 ** ± 0.24 | 1.88 * ± 0.23 |
| tph-1 | 0.85 ± 0.10 | 1.08 ± 0.07 | 0.80 ± 0.17 | 1.74 ** ± 0.14 | 1.05 ± 0.15 |
| ttx-3 | 0.59 * ± 0.15 | 1.48 ± 0.33 | 1.43 ± 0.28 | 1.17 ± 0.17 | 2.26 * ± 0.56 |
| unc-17 | 2.52 * ± 0.73 | 1.51 * ± 0.23 | 1.03 ± 0.10 | 2.37 * ± 0.34 | 1.68 * ± 0.25 |
| unc-47 | 0.47 ** ± 0.06 | 3.76 * ± 1.01 | 0.75 * ± 0.12 | 5.85 * ± 2.02 | 2.53 ** ± 0.21 |
| Gene | 0.1 μM TBBP-A + 50 μM DBAA Relative to | 50 μM TBBP-A + 50 μM DBAA Relative to | ||
|---|---|---|---|---|
| 0.1 μM TBBP-A | 50 μM DBAA | 50 μM TBBP-A | 50 μM DBAA | |
| ace-1 | n.s. | n.s. | n.s. | n.s. |
| casy-1 | * | * | n.s. | * |
| che-12 | * | ** | n.s. | n.s. |
| dat-1 | * | * | n.s. | * |
| dop-1 | n.s. | n.s. | n.s. | n.s. |
| dop-3 | * | ** | n.s. | * |
| eat-4 | n.s. | n.s. | n.s. | * |
| gbb-1 | * | * | n.s. | * |
| grd-12 | n.s. | n.s. | n.s. | n.s. |
| hda-4 | n.s. | n.s. | n.s. | n.s. |
| hen-1 | n.s. | * | * | n.s. |
| mec-10 | ** | ** | * | n.s. |
| srab-6 | * | n.s. | * | n.s. |
| tph-1 | * | * | n.s. | n.s. |
| ttx-3 | n.s. | n.s. | n.s. | n.s. |
| unc-17 | n.s. | * | n.s. | n.s. |
| unc-47 | n.s. | n.s. | n.s. | ** |
2.1.7. Fluorescence-Microscopic Observations of GFP Fusion-Proteins

| Strain | tex | 0.1 T + 50 D Relative to | 50 T + 50 D Relative to | ||
|---|---|---|---|---|---|
| 0.1 T | 50 D | 50 T | 50 D | ||
| BZ555 | 24 h | * | - | - | - |
| 72 h | - | - | - | - | |
| DA1240 | 24 h | - | - | * | ** |
| 72 h | - | - | * | * | |
| EG1285 | 24 h | ** | ** | - | ** |
| 72 h | ** | * | - | * | |
| GR1366 | 24 h | * | ** | - | - |
| 72 h | * | * | - | - | |
| LX929 | 24 h | * | ** | - | ** |
| 72 h | - | - | - | * | |
2.2. Chronic Exposure
2.2.1. Neuromolecular Experiments

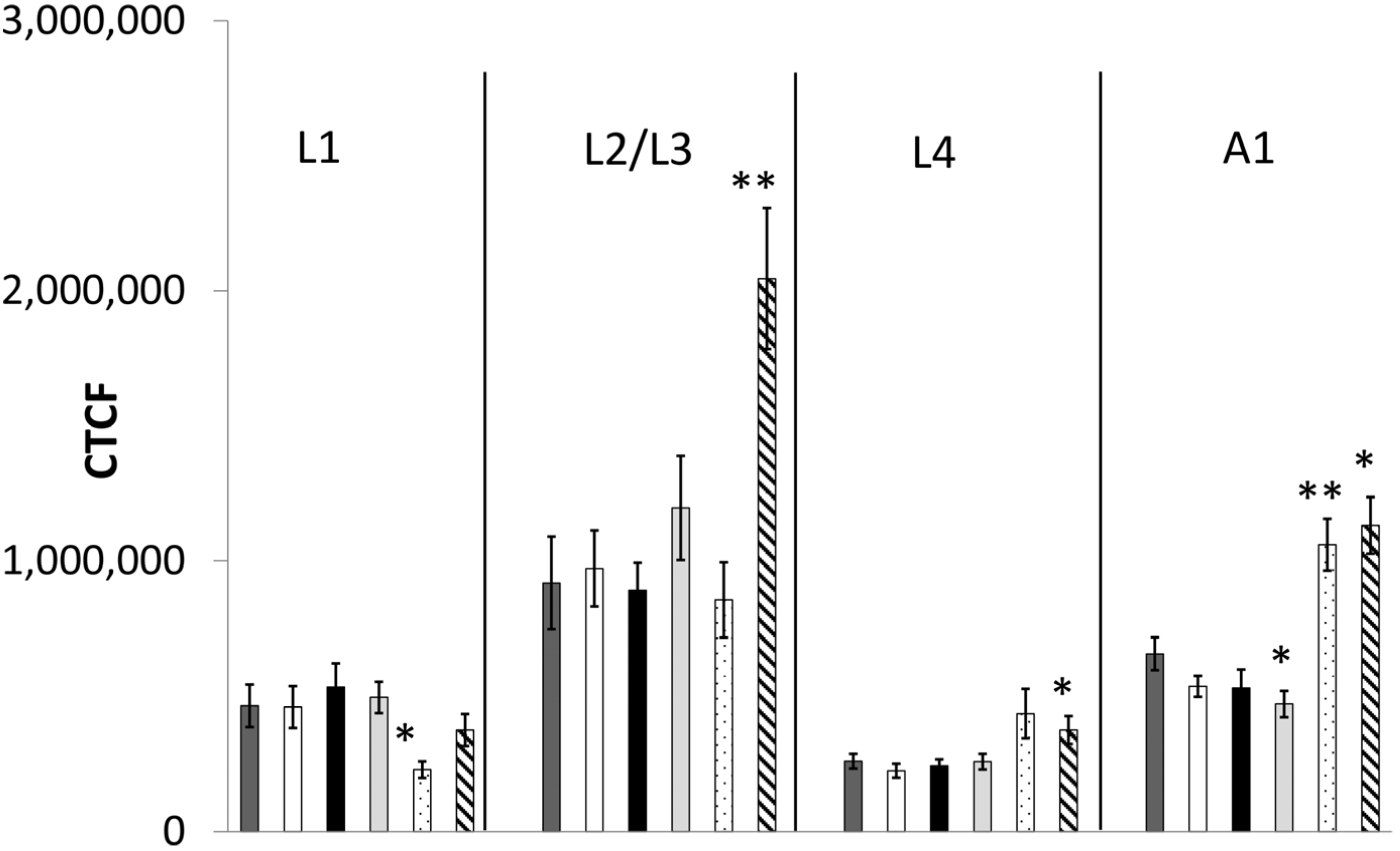
2.2.2. Fluorescence-Microscopic Observations of GFP Fusion-Proteins

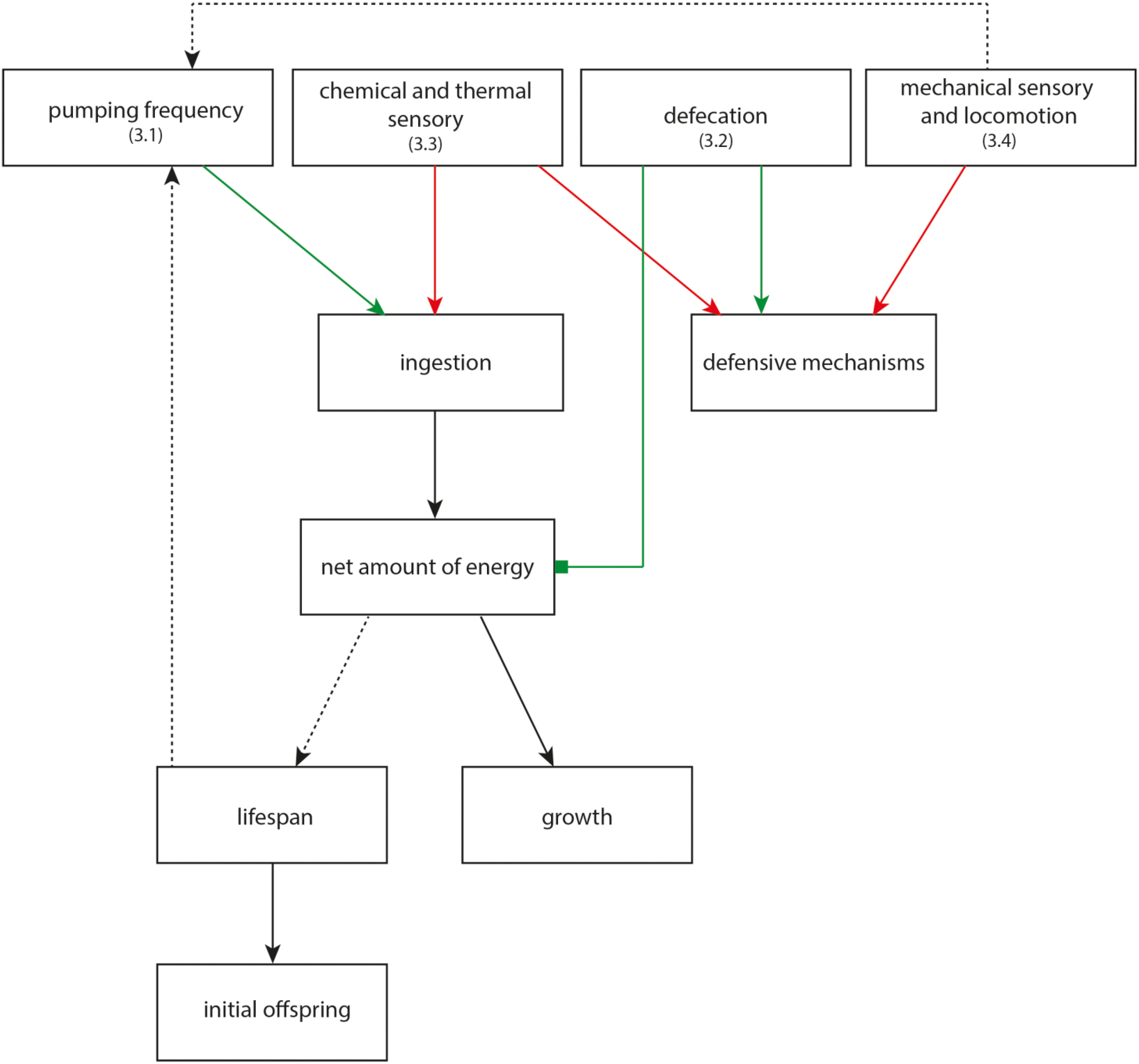
3. Discussion
3.1. Increased Pumping Frequency Leads to Increased Growth


3.2. Reduced Defecation Interval Correlates with Short Lifespan

3.3. Reduced Chemical and Thermal Perception Reduces Net-Energy

3.4. Reduction of Mechanical Sensory and Locomotive Behavior Decrease Chances of Survival in Nature

3.5. Neuro-Ontogenesis
4. Material and Methods
4.1. Strains
4.2. Organobromines and Exposure
4.3. Neurophysiologic Experiments
4.3.1. Autonomic Behavior
4.3.2. Mechanical Sensory Stimulus
4.3.3. Chemical Sensory
4.4. Neuromolecular Experiments
4.4.1. Selected Genes
4.4.2. RNA Preparation
4.4.3. qRT-PCR
4.5. Specific Gene Expression in Neurons
4.6. Statistical Significance
5. Concluding Remarks
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Asplund, B.; Grimvall, A. Organohalogens in nature. Environ. Sci. Technol. 1991, 25, 1346–1350. [Google Scholar] [CrossRef]
- Hütteroth, A.; Putschew, A.; Jekel, M. Natural production of organic bromine compounds in berlin lakes. Environ. Sci. Technol. 2007, 41, 3607–3612. [Google Scholar] [CrossRef] [PubMed]
- Kümmerer, K. Drugs in the environment: Emission of drugs, diagnostic aids and disinfectants into wastewater by hospitals in relation to other sources—A review. Chemosphere 2001, 45, 957–969. [Google Scholar] [CrossRef] [PubMed]
- Fan, A.; Howd, R.; Davis, B. Risk assessment of environmental chemicals. Annu. Rev. Pharmacol. Toxicol. 1995, 35, 341–368. [Google Scholar] [CrossRef] [PubMed]
- Saul, N.; Chakrabarti, S.; Sturzenbaum, S.R.; Menzel, R.; Steinberg, C.E. Neurotoxic action of microcystin-LR is reflected in the transcriptional stress response of Caenorhabditis elegans. Chem. Biol. Interact. 2014, 223C, 51–57. [Google Scholar] [CrossRef]
- Ju, J.; Ruan, Q.; Li, X.; Liu, R.; Li, Y.; Pu, Y.; Yin, L.; Wang, D. Neurotoxicological evaluation of microcystin-LR exposure at environmental relevant concentrations on nematode Caenorhabditis elegans. Environ. Sci. Pollut. Res. Int. 2013, 20, 1823–1830. [Google Scholar] [CrossRef] [PubMed]
- Saul, N.; Baberschke, N.; Chakrabarti, S.; Sturzenbaum, S.R.; Lieke, T.; Menzel, R.; Jonas, A.; Steinberg, C.E. Two organobromines trigger lifespan, growth, reproductive and transcriptional changes in Caenorhabditis elegans. Environ. Sci. Pollut. Res. 2014, 21, 10419–10431. [Google Scholar] [CrossRef]
- Watanabe, I.; Kashimoto, T. The flame retardant tetrabromobisphenol-A and its metabolite found in river and marine sediments in Japan. Chemosphere 1983, 12, 1533–1539. [Google Scholar] [CrossRef]
- Shaw, S.D.; Blum, A.; Weber, R.; Kannan, K.; Rich, D.; Lucas, D.; Koshland, C.P.; Dobraca, D.; Hanson, S.; Birnbaum, L.S. Halogenated flame retardants: Do the fire safety benefits justify the risks? Rev. Environ. Health 2010, 25, 261–305. [Google Scholar] [CrossRef] [PubMed]
- Pullen, S.; Boecker, R.; Tiegs, G. The flame retardants tetrabromobisphenol A and tetrabromobisphenol A-bisallylether suppress the induction of interleukin-2 receptor alpha chain (CD25) in murine splenocytes. Toxicology 2003, 184, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Kibakaya, E.C.; Stephen, K.; Whalen, M.M. Tetrabromobisphenol A has immunosuppressive effects on human natural killer cells. J. Immunotoxicol. 2009, 6, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, A.; Saigusa, D.; Tetsu, N.; Yamakuni, T.; Tomioka, Y.; Hishinuma, T. Neurobehavioral effects of tetrabromobisphenol A, a brominated flame retardant, in mice. Toxicol. Lett. 2009, 189, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Paul, N.A.; Nys, R.D.; Steinberg, P.D. Chemical defence against bacteria in the red alga Asparagopsis armata: Linking structure with function. Mar. Ecol. Prog. Ser. 2006, 306, 87–101. [Google Scholar] [CrossRef]
- Richardson, S.D.; Thruston, A.D.; Rav-Acha, C.; Groisman, L.; Popilevsky, I.; Juraev, O.; Glezer, V.; McKague, A.B.; Plewa, M.J.; Wagner, E.D. Tribromopyrrole, brominated acids, and other disinfection byproducts produced by disinfection of drinking water rich in bromide. Environ. Sci. Technol. 2003, 37, 3782–3793. [Google Scholar] [CrossRef] [PubMed]
- Putschew, A.; Mania, M.; Jekel, M. Occurrence and source of brominated organic compounds in surface waters. Chemosphere 2003, 52, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Melnick, R.L.; Nyska, A.; Foster, P.M.; Roycroft, J.H.; Kissling, G.E. Toxicity and carcinogenicity of the water disinfection byproduct, dibromoacetic acid, in rats and mice. Toxicology 2007, 230, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Moser, V.C.; Phillips, P.M.; Levine, A.B.; McDaniel, K.L.; Sills, R.C.; Jortner, B.S.; Butt, M.T. Neurotoxicity produced by dibromoacetic acid in drinking water of rats. Toxicol. Sci. 2004, 79, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Harris, T.W.; Baran, J.; Bieri, T.; Cabunoc, A.; Chan, J.; Chen, W.J.; Davis, P.; Done, J.; Grove, C.; Howe, K.; et al. Wormbase 2014: New views of curated biology. Nucleic Acids Res. 2014, 42, D789–D793. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.W.; Thomas, J.H. Regulation of a periodic motor program in C. elegans. J. Neurosci. 1994, 14, 1953–1962. [Google Scholar]
- Doncaster, C.C. Nematode feeding mechanisms. I. Observations on Rhabditis and Pelodera. Nematologica 1962, 8, 313–320. [Google Scholar] [CrossRef]
- Lehane, M.J. Peritrophic matrix structure and function. Annu. Rev. Entomol. 1997, 42, 525–550. [Google Scholar] [CrossRef] [PubMed]
- Lenaerts, I.; Walker, G.A.; van Hoorebeke, L.; Gems, D.; vanfleteren, J.R. Dietary restriction of Caenorhabditis elegans by axenic culture reflects nutritional requirement for constituents provided by metabolically active microbes. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2008, 63, 242–252. [Google Scholar] [CrossRef]
- Sulston, J.E.; Horvitz, H.R. Post-embryonic cell lineages of the nematode Caenorhabditis elegans. Dev. Boil. 1977, 56, 110–156. [Google Scholar] [CrossRef]
- Hirose, T.; Nakano, Y.; Nagamatsu, Y.; Misumi, T.; Ohta, H.; Ohshima, Y. Cyclic GMP-dependent protein kinase EGL-4 controls body size and lifespan in C. elegans. Development 2003, 130, 1089–1099. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, P.; Nishikawa, J.L.; Breitkreutz, B.J.; Tyers, M. Systematic identification of pathways that couple cell growth and division in yeast. Science 2002, 297, 395–400. [Google Scholar] [CrossRef] [PubMed]
- So, S.; Miyahara, K.; Ohshima, Y. Control of body size in C. elegans dependent on food and insulin/IGF-1 signal. Genes Cells 2011, 16, 639–651. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.A.; Shukitt-Hale, B.; Kalt, W.; Ingram, D.K.; Joseph, J.A.; Wolkow, C.A. Blueberry polyphenols increase lifespan and thermotolerance in Caenorhabditis elegans. Aging Cell 2006, 5, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Saul, N.; Pan, B.; Menzel, R.; Steinberg, C.E. The non-target organism Caenorhabditis elegans withstands the impact of sulfamethoxazole. Chemosphere 2013, 93, 2373–2380. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.M.; Shanmuganayagam, D.; Weindruch, R. Caloric restriction and aging: Studies in mice and monkeys. Toxicol. Pathol. 2009, 37, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Colman, R.J.; Anderson, R.M.; Johnson, S.C.; Kastman, E.K.; Kosmatka, K.J.; Beasley, T.M.; Allison, D.B.; Cruzen, C.; Simmons, H.A.; Kemnitz, J.W.; et al. Caloric restriction delays disease onset and mortality in rhesus monkeys. Science 2009, 325, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Voland, E. Altern und Lebenslauf—ein Evolutionsbiologischer Aufriss. In Generationen: Multidisziplinäre Perspektiven; Künemund, H., Szydlik, M., Eds.; VS Verlag für Sozialwissenschaften: Wiesbaden, Germany, 2009. [Google Scholar]
- Huang, C.; Xiong, C.; Kornfeld, K. Measurements of age-related changes of physiological processes that predict lifespan of Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2004, 101, 8084–8089. [Google Scholar] [CrossRef] [PubMed]
- Avery, L. Motor neuron m3 controls pharyngeal muscle relaxation timing in Caenorhabditis elegans. J. Exp. Biol. 1993, 175, 283–297. [Google Scholar] [PubMed]
- Riddle, D.L.; Blumenthal, T.; Meyer, B.J.; Avery, L.; Thomas, J.H. Feeding. In C. elegans II, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1997. [Google Scholar]
- Raizen, D.M.; Avery, L. Electrical activity and behavior in the pharynx of Caenorhabditis elegans. Neuron 1994, 12, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Niacaris, T.; Avery, L. Serotonin regulates repolarization of the C. elegans pharyngeal muscle. J. Exp. Biol. 2003, 206, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Sanger, G.J.; Yoshida, M.; Yahyah, M.; Kitazumi, K. Increased defecation during stress or after 5-hydroxytryptophan: Selective inhibition by the 5-HT(4) receptor antagonist, SB-207266. Br. J. Pharmacol. 2000, 130, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Martinez, V.; Wang, L.; Rivier, J.; Grigoriadis, D.; Tache, Y. Central CRF, urocortins and stress increase colonic transit via CRF1 receptors while activation of CRF2 receptors delays gastric transit in mice. J. Physiol. 2004, 556, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Branicky, R.; Hekimi, S. What keeps C. elegans regular: The genetics of defecation. Trends Genet. 2006, 22, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Eastman, C.; Horvitz, H.R.; Jin, Y. Coordinated transcriptional regulation of the unc-25 glutamic acid decarboxylase and the unc-47 GABA vesicular transporter by the Caenorhabditis elegans UNC-30 homeodomain protein. J. Neurosci. 1999, 19, 6225–6234. [Google Scholar] [PubMed]
- Riddle, D.L.B.T.; Meyer, B.J.; Priess, J.R. Defecation motor program. In C. elegans II, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1997. [Google Scholar]
- De Bono, M.; Maricq, A.V. Neuronal substrates of complex behaviors in C. elegans. Annu. Rev. Neurosci. 2005, 28, 451–501. [Google Scholar] [CrossRef] [PubMed]
- Wehner, R.; Gehring, W.J. Verhaltensökologie. In Zoologie; Thieme Verlag: Stuttgart, Germany, 2007. [Google Scholar]
- Grewal, P.S.; Wright, D.J. Migration of Caenorhabditis elegans larvae towards bacteria and the nature of the bacterial stimulus. Fund. Appl. Nematol. 1992, 15, 159–166. [Google Scholar]
- Bargmann, C.I.; Hartwieg, E.; Horvitz, H.R. Odorant-selective genes and neurons mediate olfaction in C. elegans. Cell 1993, 74, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Irazoqui, J.E.; Urbach, J.M.; Ausubel, F.M. Evolution of host innate defence: Insights from Caenorhabditis elegans and primitive invertebrates. Nat. Rev. Immunol. 2010, 10, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Felix, M.A.; Braendle, C. The natural history of Caenorhabditis elegans. Curr. Biol. 2010, 20, R965–R969. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.E.; Artan, M.; Seo, K.; Lee, S.J. Regulation of lifespan by chemosensory and thermosensory systems: Findings in invertebrates and their implications in mammalian aging. Front. Genet. 2012, 3, 218. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Shen, Y. Structural basis for degeneracy among thermosensory neurons in Caenorhabditis elegans. J. Neurosci. 2012, 32, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Bargmann, C.I. Chemosensation in C. elegans. Available online: http://www.wormbook.org (accessed on 5 April 2014).
- Ishihara, T.; Iino, Y.; Mohri, A.; Mori, I.; Gengyo-Ando, K.; Mitani, S.; Katsura, I. HEN-1, a secretory protein with an LDL receptor motif, regulates sensory integration and learning in Caenorhabditis elegans. Cell 2002, 109, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, N.; Kuhara, A.; Nakamura, F.; Okochi, Y.; Mori, I. Bidirectional regulation of thermotaxis by glutamate transmissions in Caenorhabditis elegans. EMBO J. 2011, 30, 1376–1388. [Google Scholar] [CrossRef] [PubMed]
- Hoerndli, F.J.; Walser, M.; Frohli Hoier, E.; de Quervain, D.; Papassotiropoulos, A.; Hajnal, A. A conserved function of C. elegans CASY-1 calsyntenin in associative learning. PLoS ONE 2009, 4, e4880. [Google Scholar] [CrossRef] [PubMed]
- Bacaj, T.; Lu, Y.; Shaham, S. The conserved proteins CHE-12 and DYF-11 are required for sensory cilium function in Caenorhabditis elegans. Genetics 2008, 178, 989–1002. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Pai, S.; Zhao, Z.; Mah, A.; Newbury, R.; Johnsen, R.C.; Altun, Z.; Moerman, D.G.; Baillie, D.L.; Stein, L.D. Identification of a nematode chemosensory gene family. Proc. Natl. Acad. Sci. USA 2005, 102, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Biron, D.; Wasserman, S.; Thomas, J.H.; Samuel, A.D.; Sengupta, P. An olfactory neuron responds stochastically to temperature and modulates Caenorhabditis elegans thermotactic behavior. Proc. Natl. Acad. Sci. USA 2008, 105, 11002–11007. [Google Scholar] [CrossRef] [PubMed]
- Kimata, T.; Sasakura, H.; Ohnishi, N.; Nishio, N.; Mori, I. Thermotaxis of C. elegans as a model for temperature perception, neural information processing and neural plasticity. Worm 2012, 1, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Chase, D.L.; Pepper, J.S.; Koelle, M.R. Mechanism of extrasynaptic dopamine signaling in Caenorhabditis elegans. Nat. Neurosci. 2004, 7, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Sawin, E.R.; Ranganathan, R.; Horvitz, H.R. C. elegans locomotory rate is modulated by the environment through a dopaminergic pathway and by experience through a serotonergic pathway. Neuron 2000, 26, 619–631. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, S.A.; Durand, S.; Dasgupta, A.; Radonovich, M.; Cimarelli, A.; Brady, J.N.; Mahieux, R.; Pise-Masison, C.A. The transcription profile of TAX-3 is more similar to TAX-1 than TAX-2: Insights into HTLV-3 potential leukemogenic properties. PLoS ONE 2012, 7, e41003. [Google Scholar] [CrossRef] [PubMed]
- Zane, L.; Jeang, K.T. The importance of ubiquitination and sumoylation on the transforming activity of HTLV TAX-1 and TAX-2. Retrovirology 2012, 9, 103. [Google Scholar] [CrossRef] [PubMed]
- Hohnberg, K.; Traunspurger, W. Foraging theory and partial consumption in a tardigrade-nematode system. Behav. Ecol. 2009, 20, 884–890. [Google Scholar] [CrossRef]
- Bento, G.; Ogawa, A.; Sommer, R.J. Co-option of the hormone-signalling module dafachronic acid-DAF-12 in nematode evolution. Nature 2010, 466, 494–497. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, J.M.; Horvitz, H.R. A dual mechanosensory and chemosensory neuron in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 1993, 90, 2227–2231. [Google Scholar] [CrossRef] [PubMed]
- Rankin, C.H.; Broster, B.S. Factors affecting habituation and recovery from habituation in the nematode Caenorhabditis elegans. Behav. Neurosci. 1992, 106, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, S.; Wintle, R.F.; Kindt, K.S.; Nuttley, W.M.; Arvan, R.; Fitzmaurice, P.; Bigras, E.; Merz, D.C.; Hebert, T.E.; van der Kooy, D.; et al. Dopamine modulates the plasticity of mechanosensory responses in Caenorhabditis elegans. EMBO J. 2004, 23, 473–482. [Google Scholar] [CrossRef] [PubMed]
- White, J.G.; Southgate, E.; Thomson, J.N.; Brenner, S. The structure of the nervous system of the nematode Caenorhabditis elegans. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1986, 314, 1–340. [Google Scholar] [CrossRef]
- Steinberg, P.L.; Merguerian, P.A.; Bihrle, W., III; Seigne, J.D. The cost of learning robotic-assisted prostatectomy. Urology 2008, 72, 1068–1072. [Google Scholar] [CrossRef] [PubMed]
- McIntire, S.L.; Jorgensen, E.; Kaplan, J.; Horvitz, H.R. The gabaergic nervous system of Caenorhabditis elegans. Nature 1993, 364, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Goodman, M.B. Mechanosensation. Available online: http://www.wormbook.org (accessed on 5 April 2014).
- Riddle, D.L.; Blumenthal, T.; Meyer, B.J.; Avery, L.; Thomas, J.H. Introduction: The neural circuit for locomotion. In C. elegans II, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1997. [Google Scholar]
- Connon, R.E.; Geist, J.; Werner, I. Effect-based tools for monitoring and predicting the ecotoxicological effects of chemicals in the aquatic environment. Sensors 2012, 12, 12741–12771. [Google Scholar] [CrossRef] [PubMed]
- De Nadal, E.; Ammerer, G.; Posas, F. Controlling gene expression in response to stress. Nat. Rev. Genet. 2011, 12, 833–845. [Google Scholar] [PubMed]
- Wu, S.; Lu, J.; Rui, Q.; Yu, S.; Cai, T.; Wang, D. Aluminum nanoparticle exposure in L1 larvae results in more severe lethality toxicity than in L4 larvae or young adults by strengthening the formation of stress response and intestinal lipofuscin accumulation in nematodes. Environ. Toxicol. Pharmacol. 2011, 31, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Sahara, T.; Goda, T.; Ohgiya, S. Comprehensive expression analysis of time-dependent genetic responses in yeast cells to low temperature. J. Biol. Chem. 2002, 277, 50015–50021. [Google Scholar] [CrossRef] [PubMed]
- Kilian, J.; Whitehead, D.; Horak, J.; Wanke, D.; Weinl, S.; Batistic, O.; D’Angelo, C.; Bornberg-Bauer, E.; Kudla, J.; Harter, K. The atgenexpress global stress expression data set: Protocols, evaluation and model data analysis of UV-B light, drought and cold stress responses. Plant J. 2007, 50, 347–363. [Google Scholar] [CrossRef] [PubMed]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [PubMed]
- Lewis, J.A.; Fleming, J.T. Basic culture methods. Methods Cell Biol. 1995, 48, 3–29. [Google Scholar] [PubMed]
- Ju, J.; Lieke, T.; Saul, N.; Pu, Y.; Yin, L.; Kochan, C.; Putschew, A.; Baberschke, N.; Steinberg, C.E.W. Neurotoxic evaluation of two organobromine model compounds and natural AOBr-containing surface water samples by a Caenorhabditis elegans test. Ecotoxicol. Environ. Saf. 2014, 104, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Hart, A.C. Behavior in WormBook. Available online: http://www.wormbook.org (accessed on 5 April 2014).
- Liedtke, W.; Tobin, D.M.; Bargmann, C.I.; Friedman, J.M. Mammalian TRPV4 (VR-OAC) directs behavioral responses to osmotic and mechanical stimuli in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2003, 100 (Suppl. 2), 14531–14536. [Google Scholar] [CrossRef] [PubMed]
- Ward, S. Chemotaxis by the nematode Caenorhabditis elegans: Identification of attractants and analysis of the response by use of mutants. Proc. Natl. Acad. Sci. USA 1973, 70, 817–821. [Google Scholar] [CrossRef] [PubMed]
- Heugens, E.H.; Hendriks, A.J.; Dekker, T.; van Straalen, N.M.; Admiraal, W. A review of the effects of multiple stressors on aquatic organisms and analysis of uncertainty factors for use in risk assessment. Crit. Rev. Toxicol. 2001, 31, 247–284. [Google Scholar] [CrossRef] [PubMed]
- Van der Linden, A.M.; Beverly, M.; Kadener, S.; Rodriguez, J.; Wasserman, S.; Rosbash, M.; Sengupta, P. Genome-wide analysis of light- and temperature-entrained circadian transcripts in Caenorhabditis elegans. PLoS Biol. 2010, 8, e1000503. [Google Scholar] [CrossRef] [PubMed]
- Simonetta, S.H.; Romanowski, A.; Minniti, A.N.; Inestrosa, N.C.; Golombek, D.A. Circadian stress tolerance in adult Caenorhabditis elegans. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2008, 194, 821–828. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lieke, T.; Steinberg, C.E.W.; Ju, J.; Saul, N. Natural Marine and Synthetic Xenobiotics Get on Nematode’s Nerves: Neuro-Stimulating and Neurotoxic Findings in Caenorhabditis elegans. Mar. Drugs 2015, 13, 2785-2812. https://doi.org/10.3390/md13052785
Lieke T, Steinberg CEW, Ju J, Saul N. Natural Marine and Synthetic Xenobiotics Get on Nematode’s Nerves: Neuro-Stimulating and Neurotoxic Findings in Caenorhabditis elegans. Marine Drugs. 2015; 13(5):2785-2812. https://doi.org/10.3390/md13052785
Chicago/Turabian StyleLieke, Thora, Christian E. W. Steinberg, Jingjuan Ju, and Nadine Saul. 2015. "Natural Marine and Synthetic Xenobiotics Get on Nematode’s Nerves: Neuro-Stimulating and Neurotoxic Findings in Caenorhabditis elegans" Marine Drugs 13, no. 5: 2785-2812. https://doi.org/10.3390/md13052785