Protective Effect of Meretrix meretrix Oligopeptides on High-Fat-Diet-Induced Non-Alcoholic Fatty Liver Disease in Mice

 ,
,
Abstract
:1. Introduction
2. Results
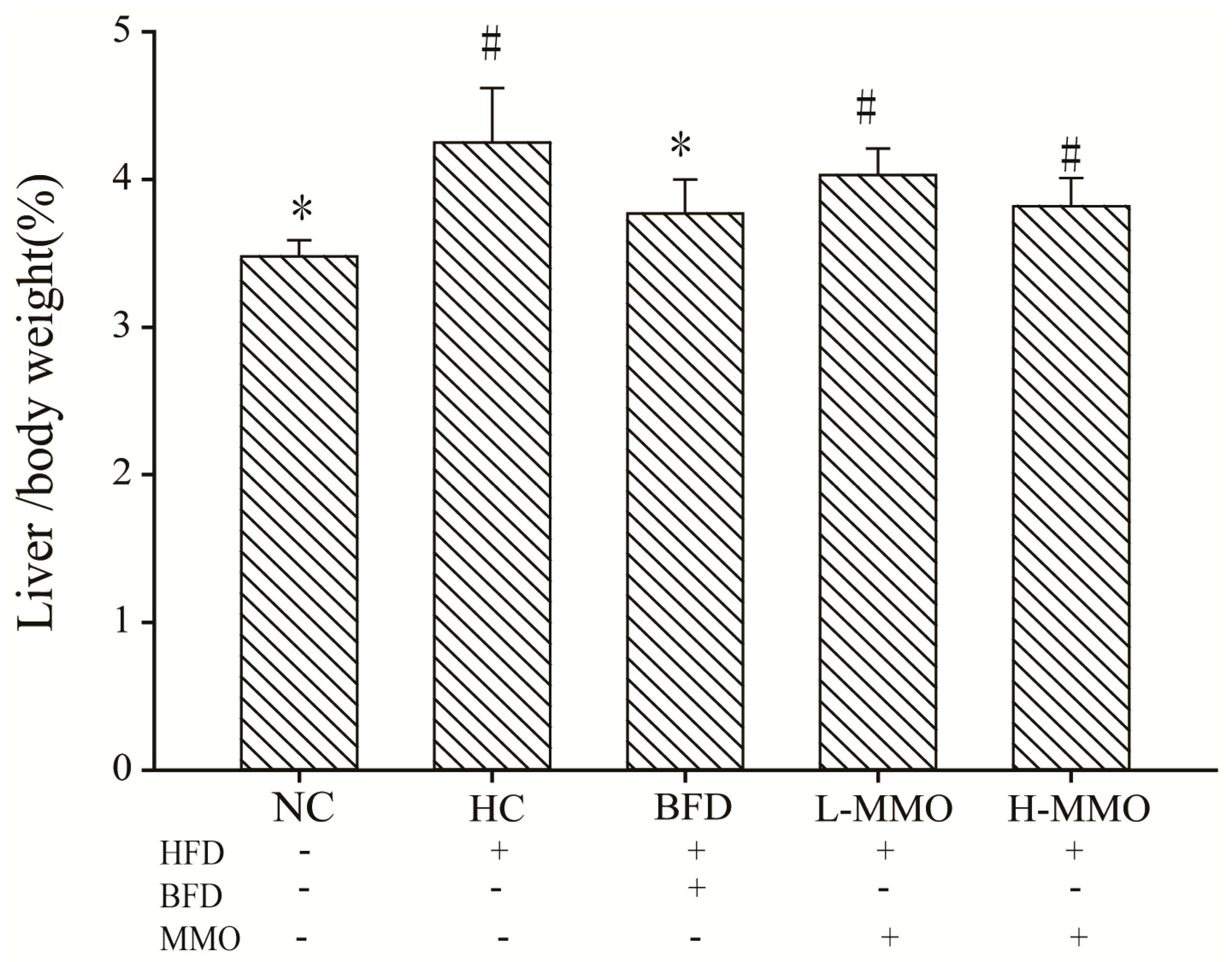
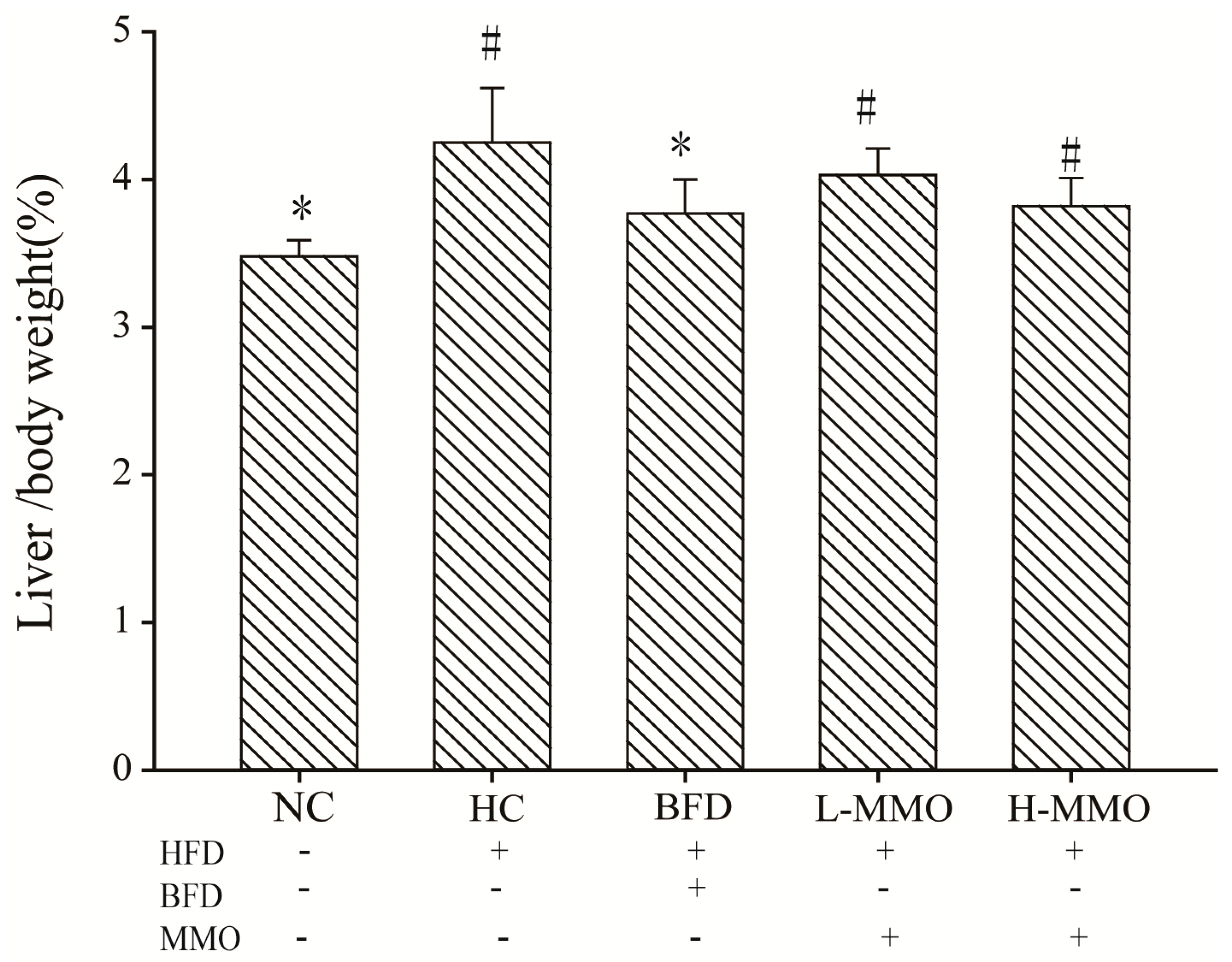
2.1. Effects of MMO on the Body Weight and Relative Liver Weight
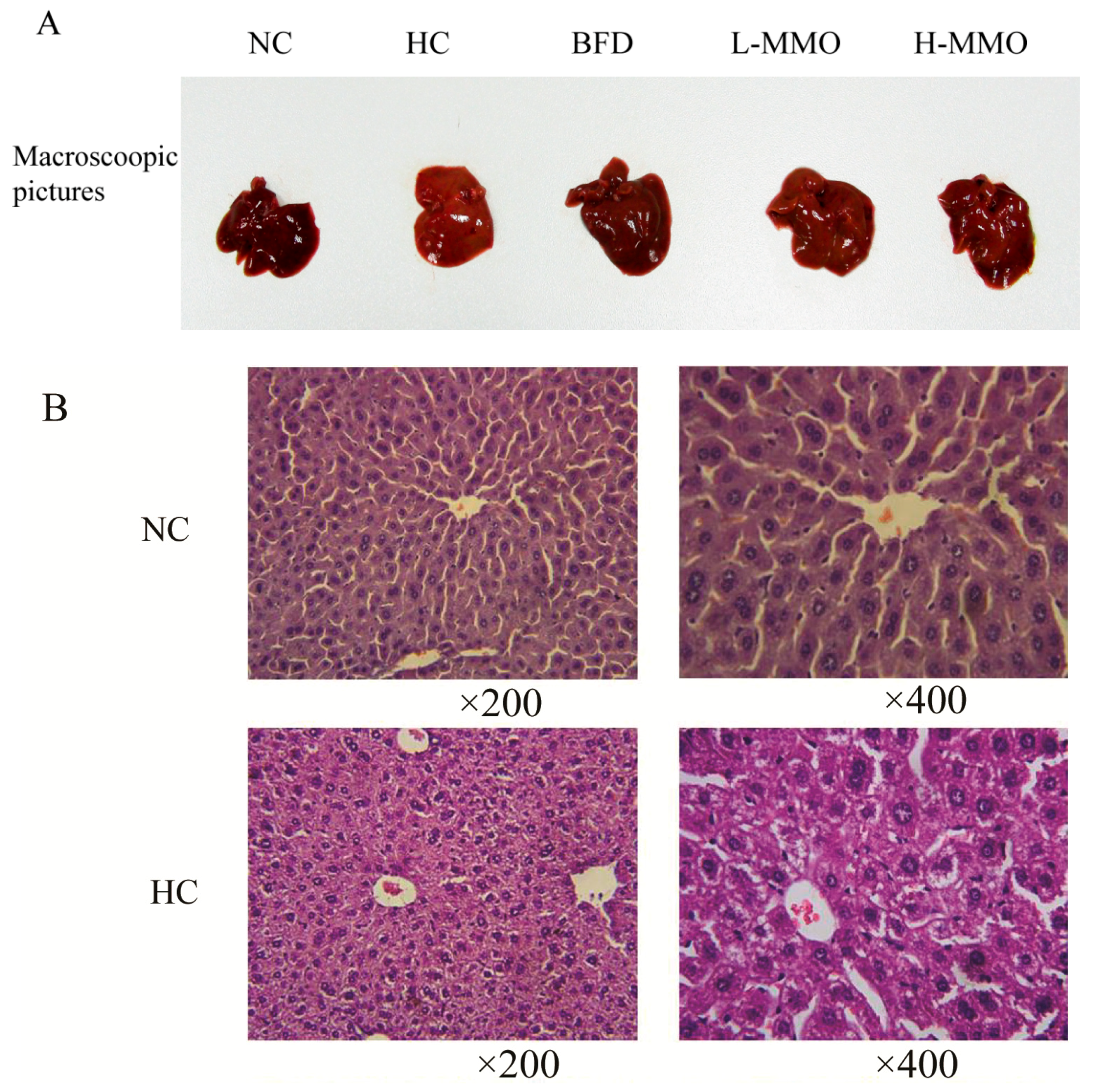
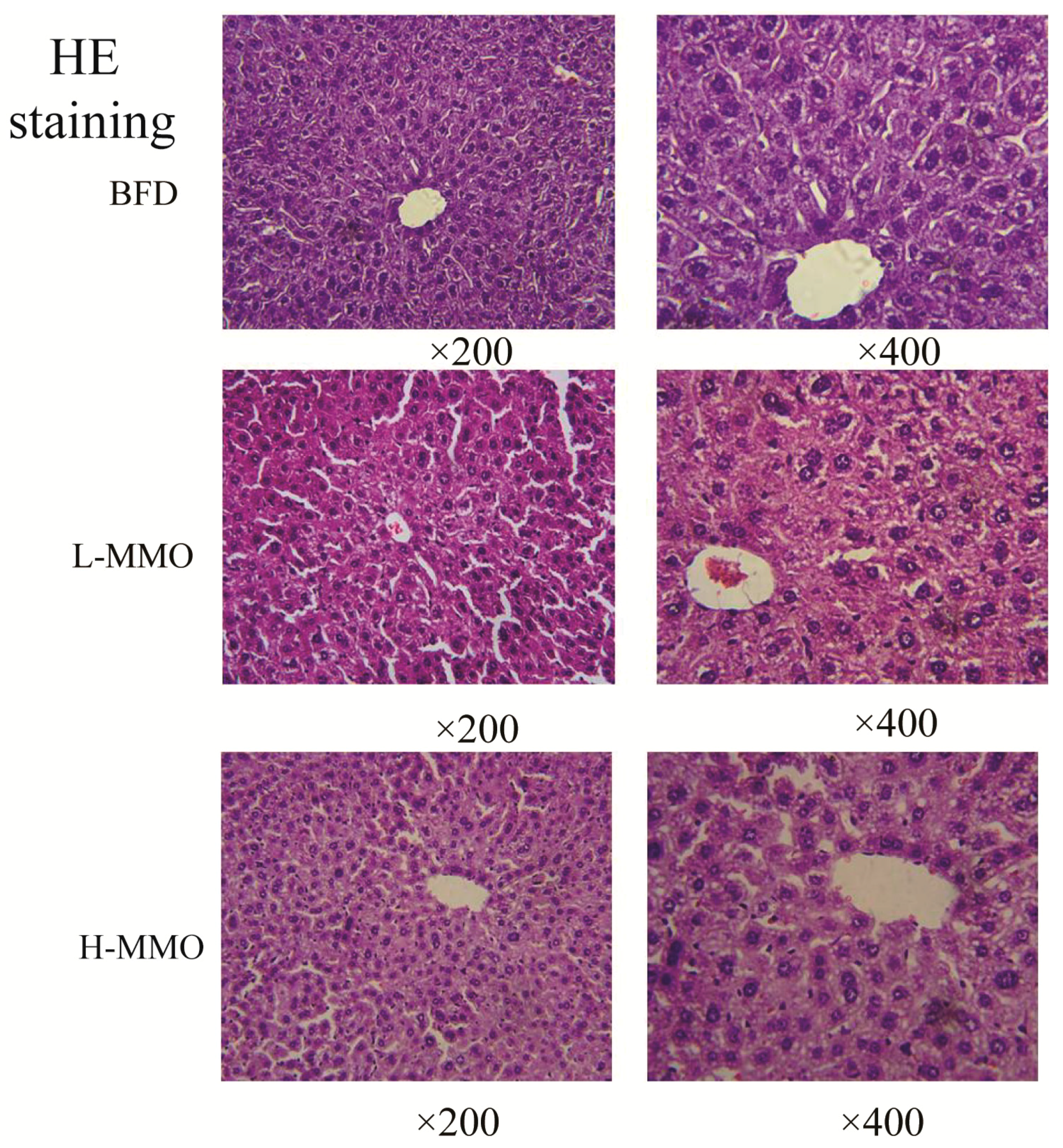
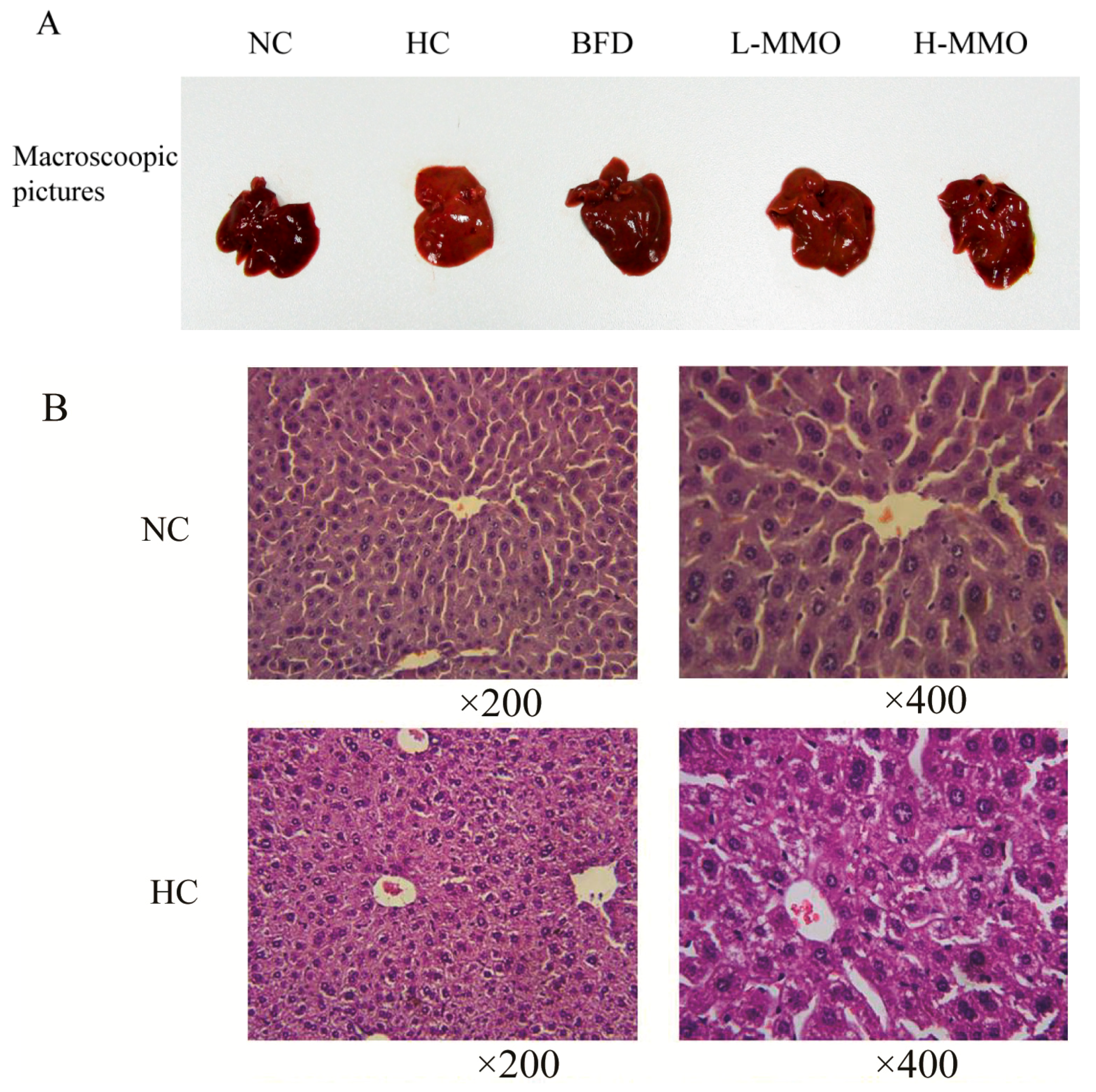
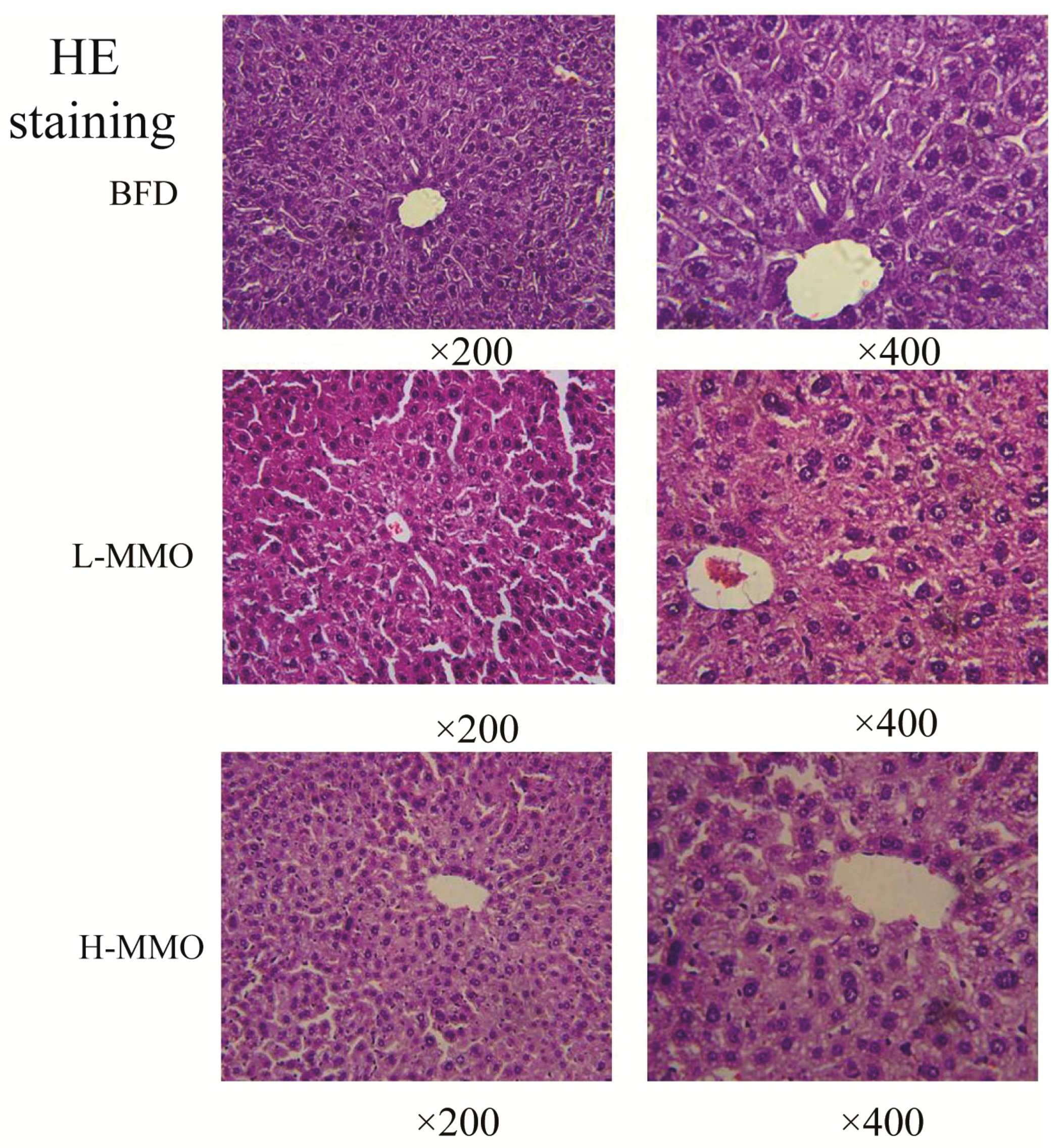
2.2. Effects of MMO on Liver Histopathology in Different Groups
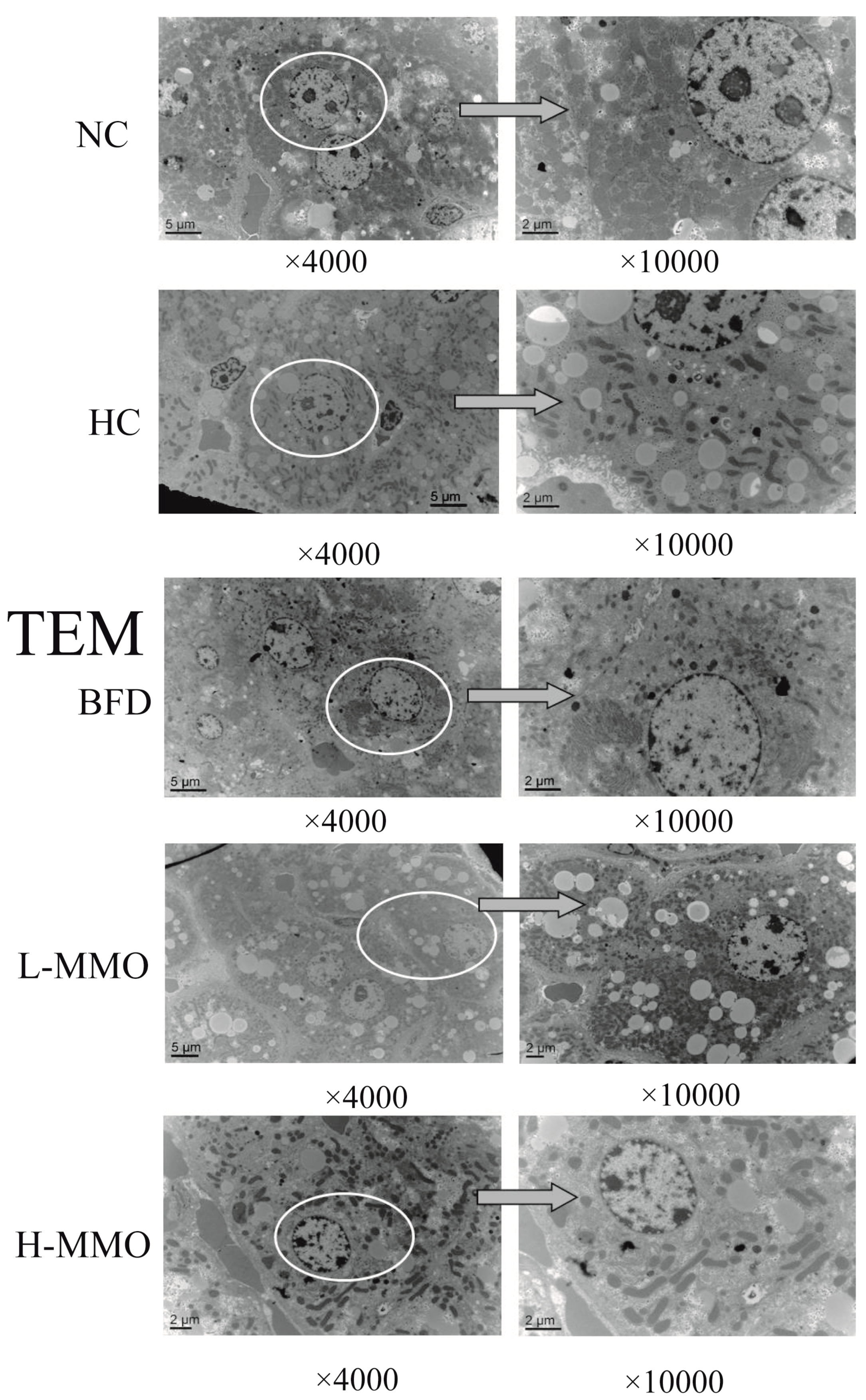
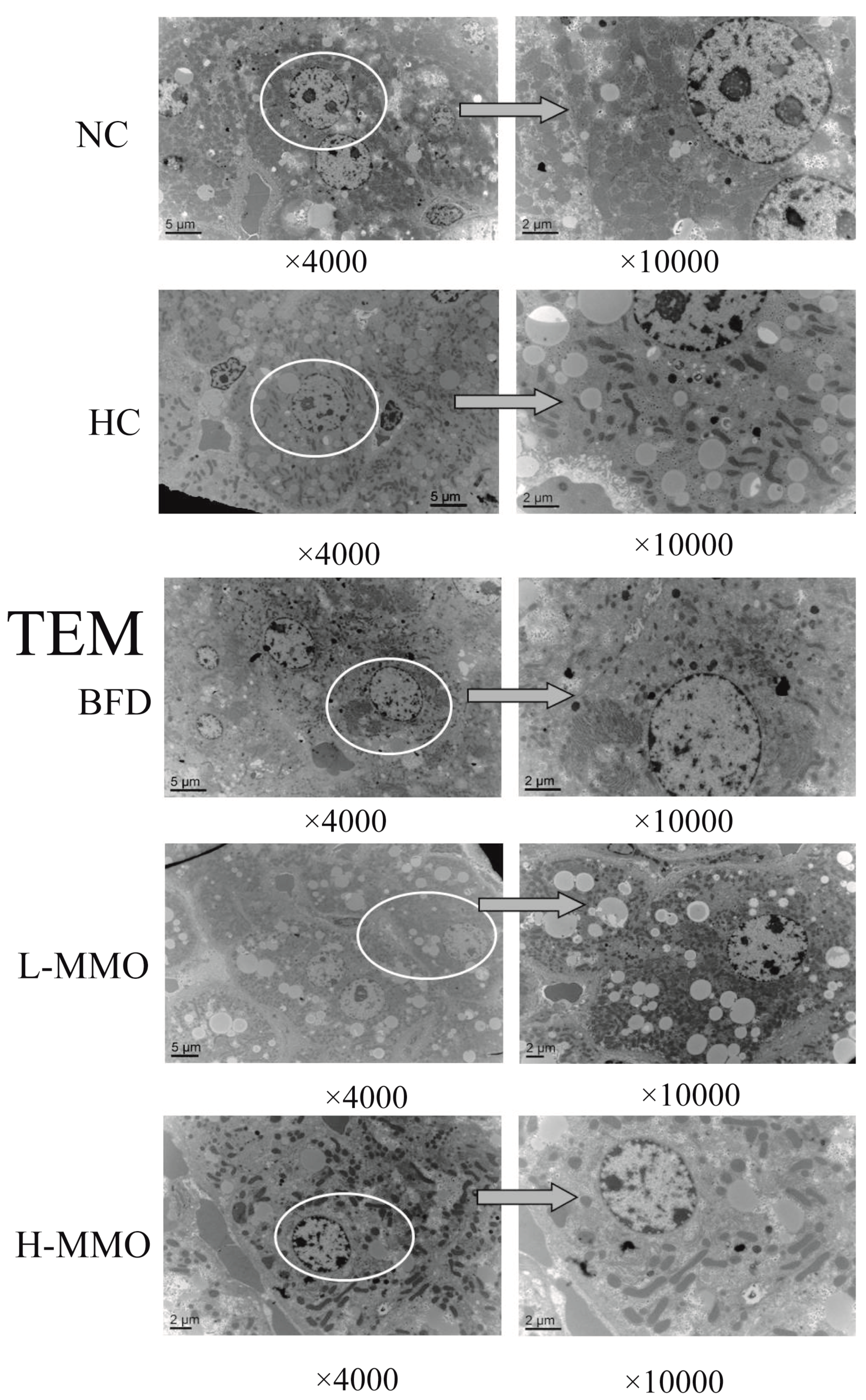
2.3. Effects of MMO on the Ultrastructural Changes in the Liver in Different Groups
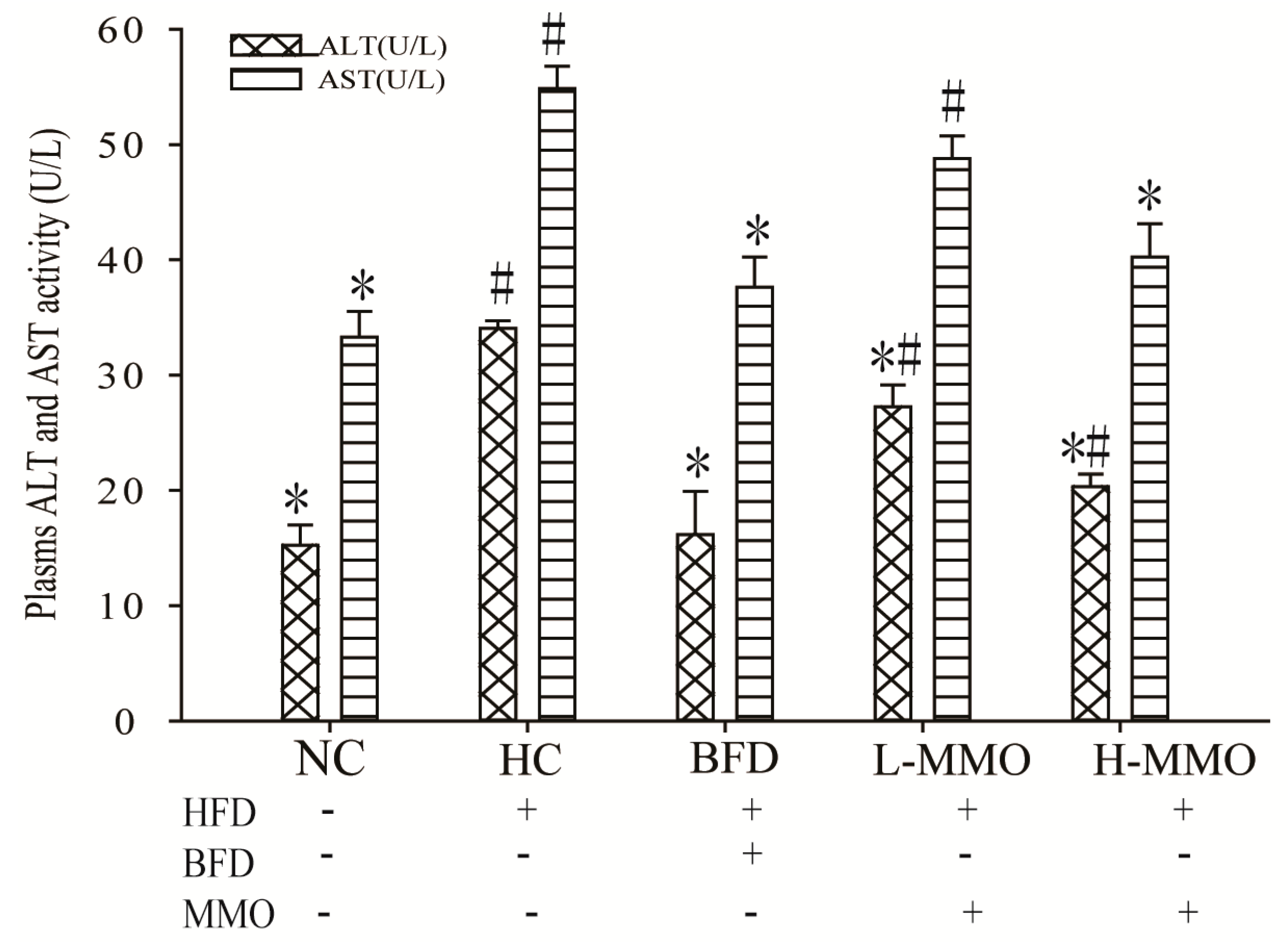
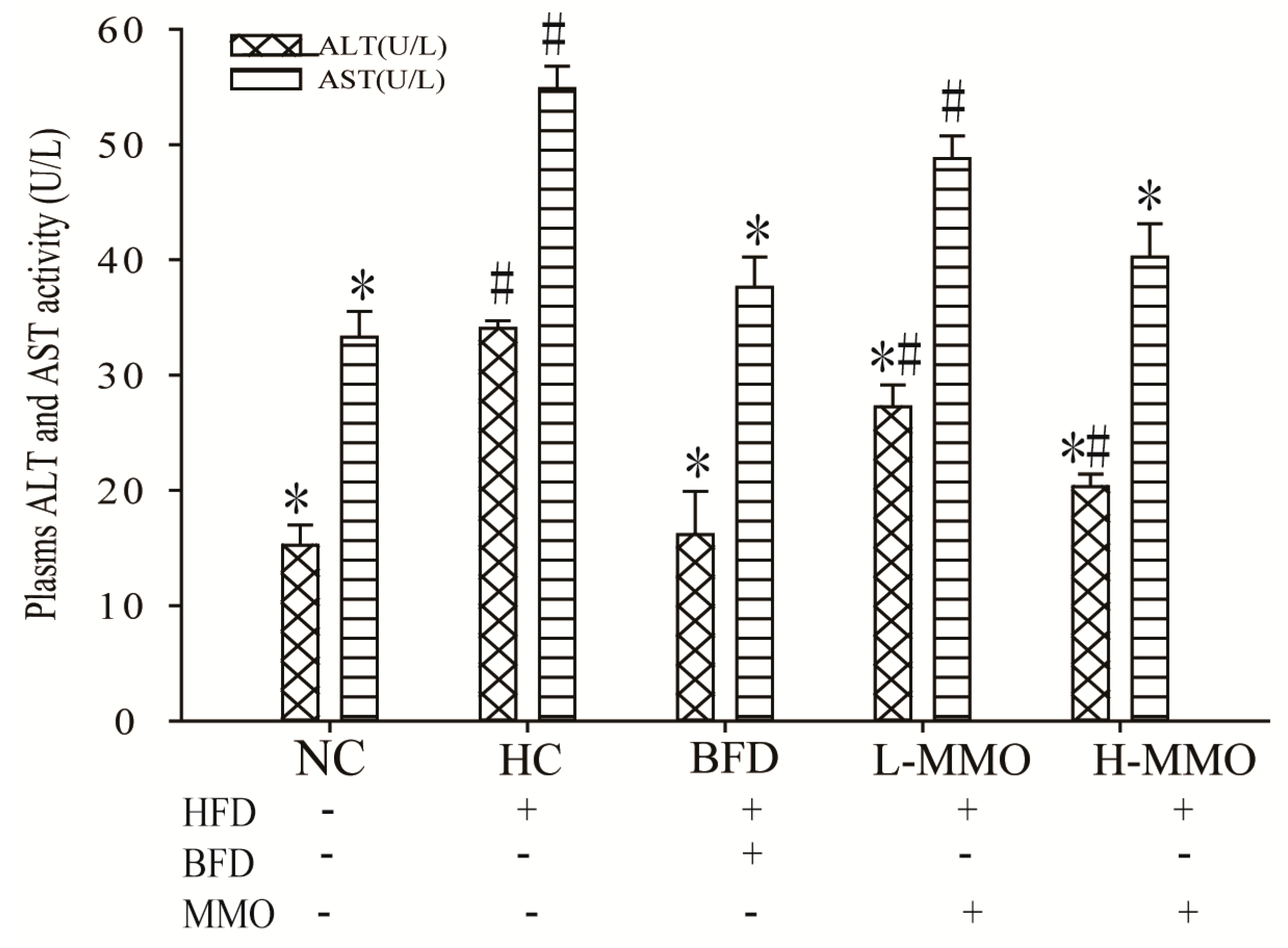
2.4. Effects of MMO on Serum Aspartate Transaminase (AST) and Alanine Transferase (ALT) Activities in Different Groups
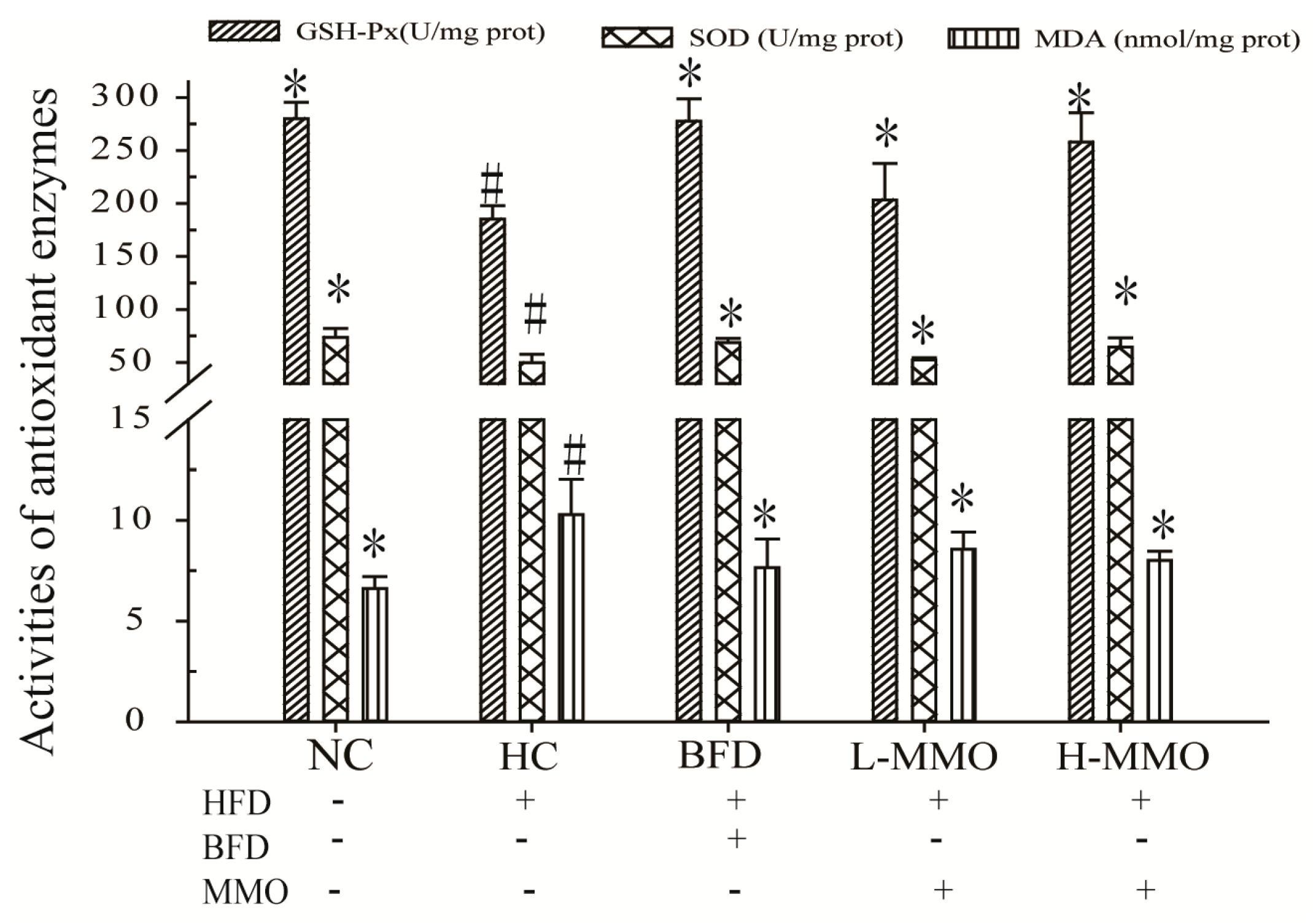
2.5. Effects of MMO on Antioxidant Capacities in Liver Tissues in Different Groups
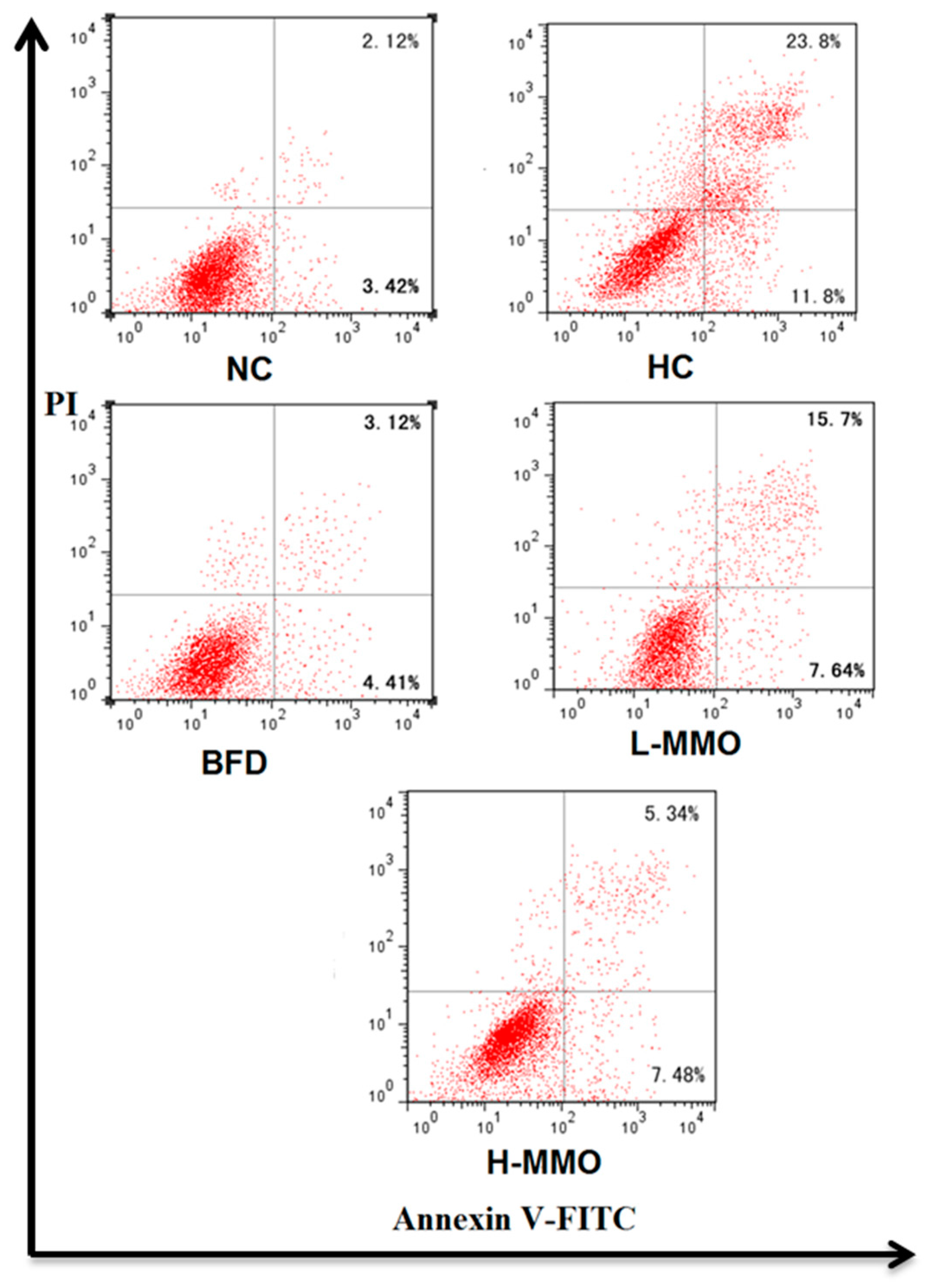
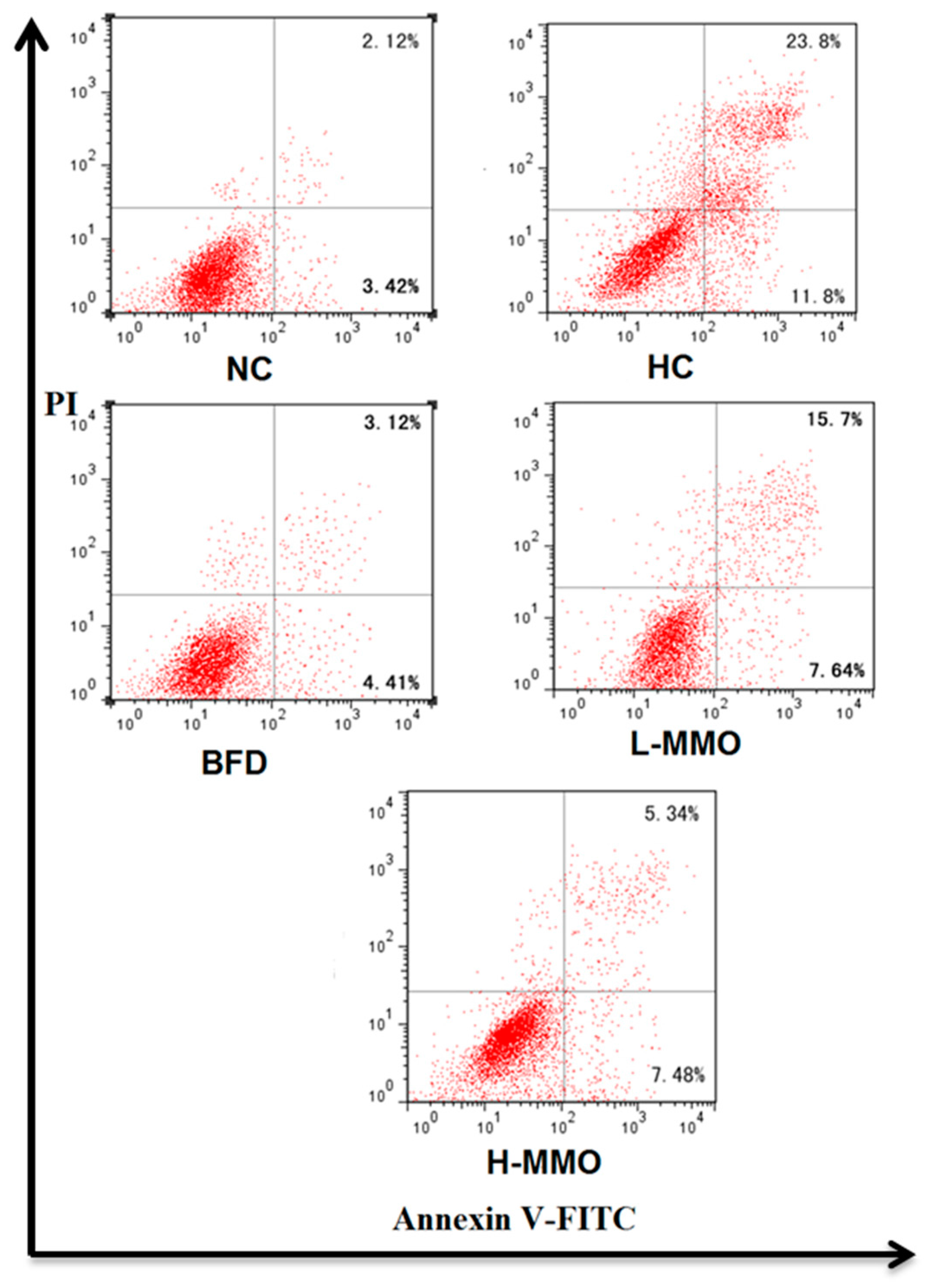
2.6. Effect of MMO on Apoptosis in NAFLD Mice
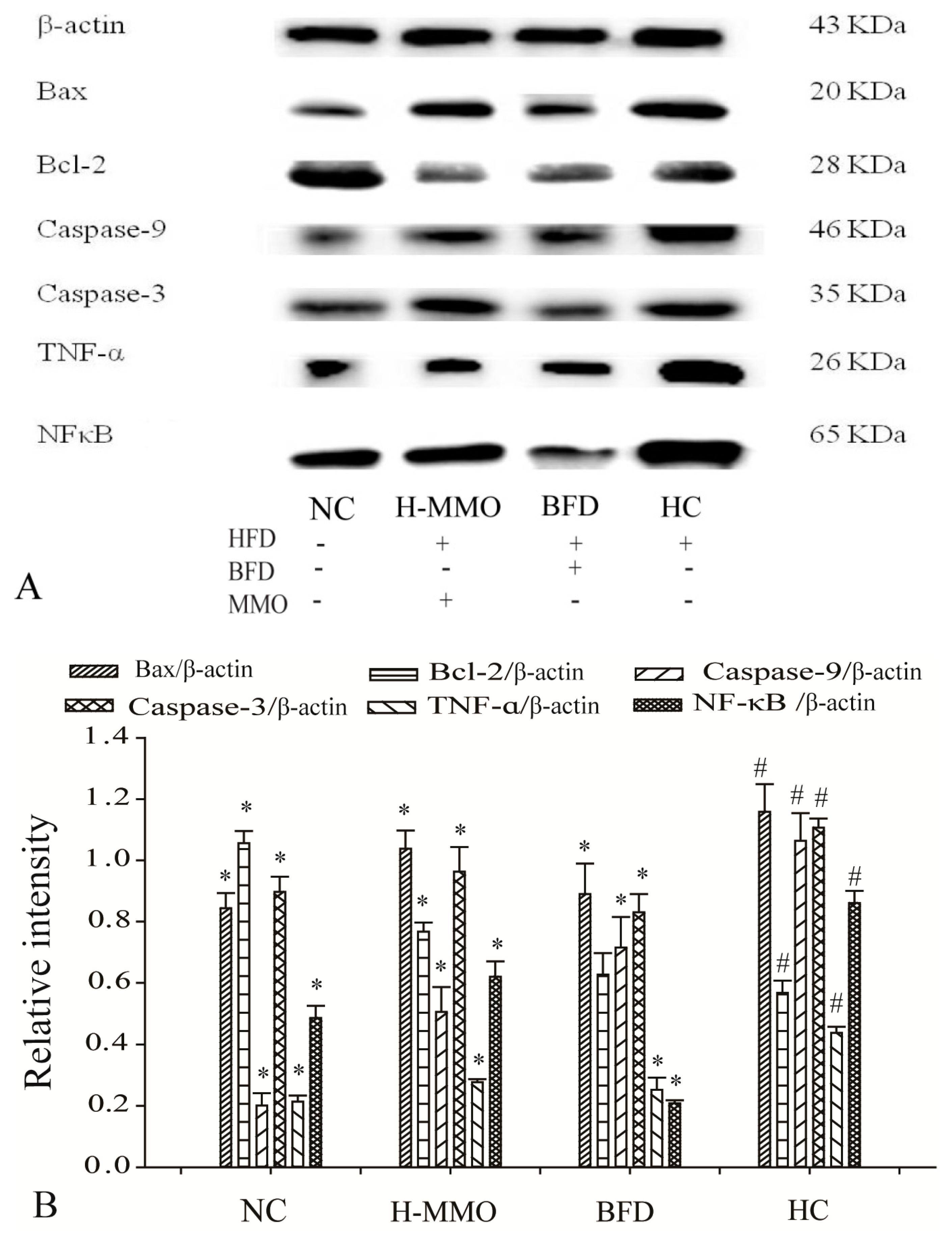
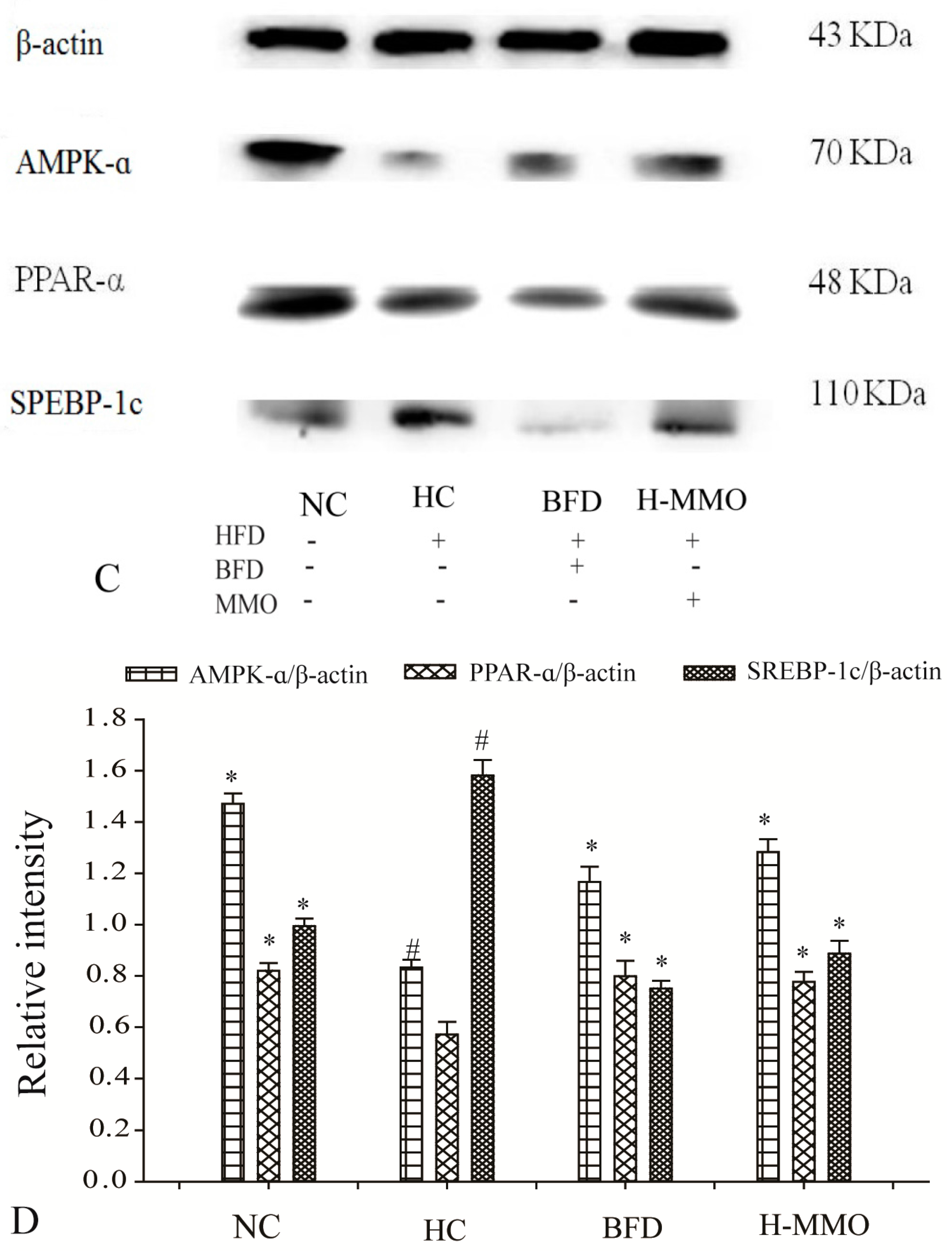
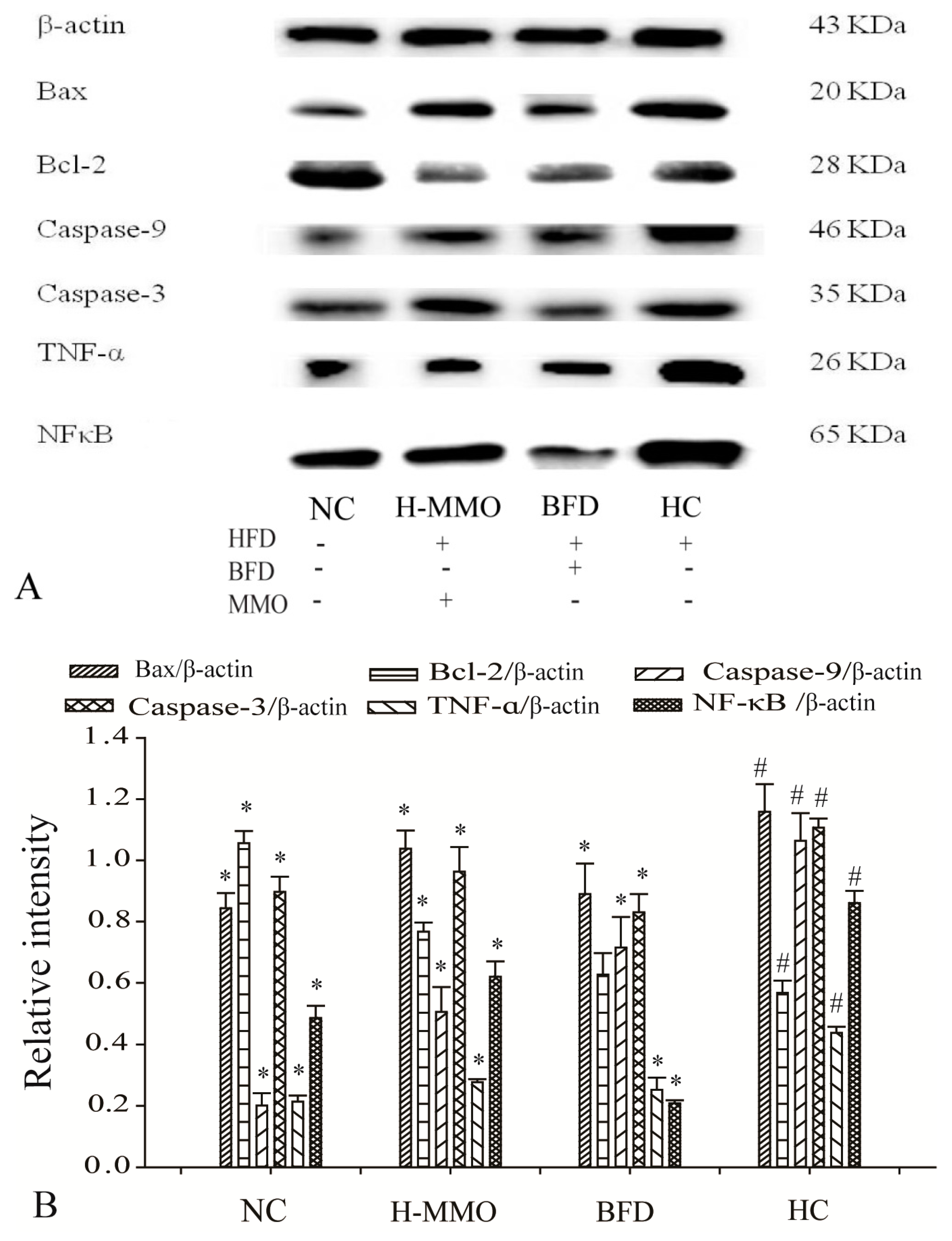
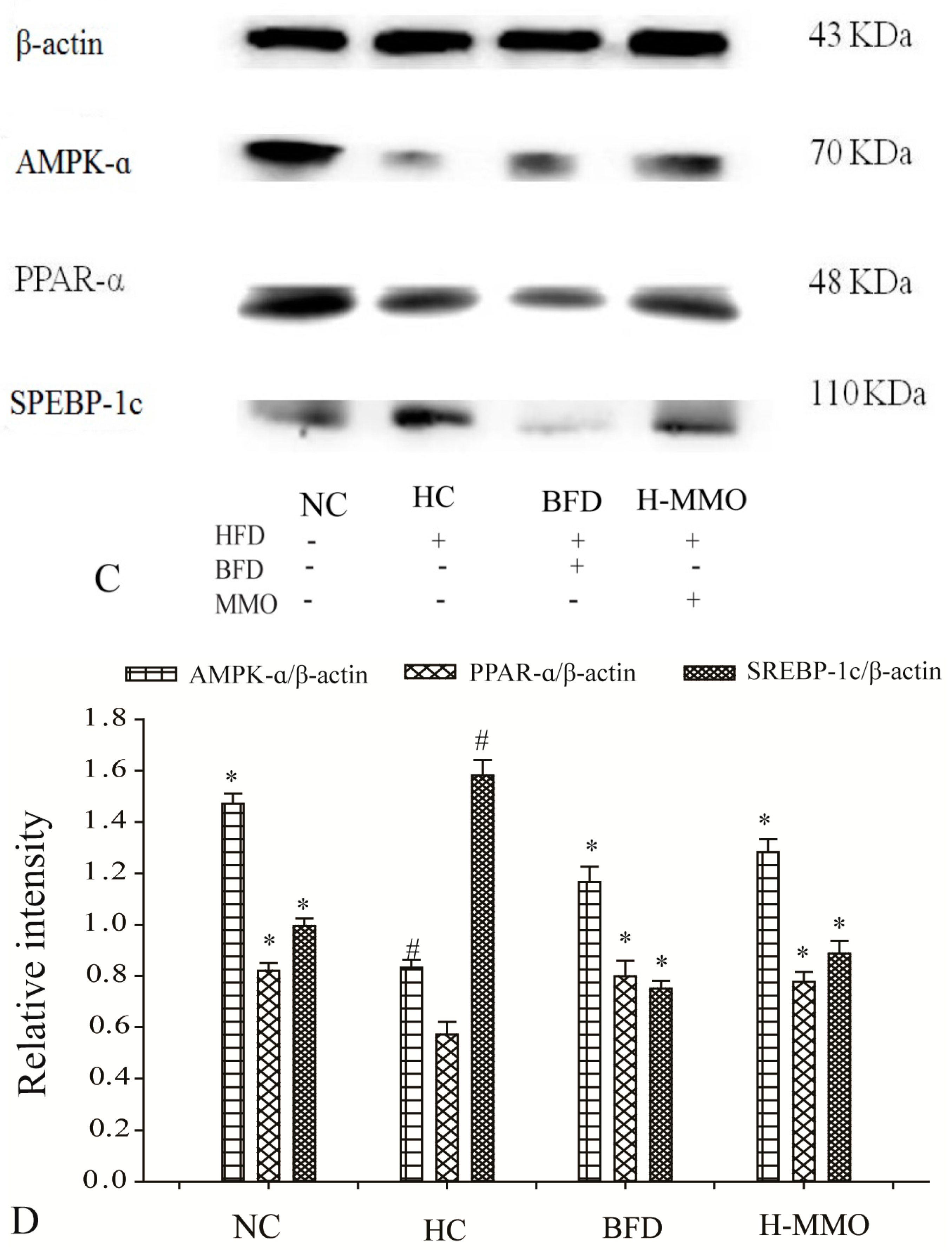
2.7. Effects of MMO on Hepatic Protein Expression Levels
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Preparation of Meretrix meretrix Oligopeptides (MMO)
4.3. Animals, Diet and Study Design
4.4. Biochemical Analysis of Serum Enzymes
4.5. Histopathological Examination
4.6. Hepatocytes Apoptosis of Liver in NAFLD Mice
4.7. Measurement of Glutathione Peroxidase (GSH-Px), Superoxide Dismutase (SOD) and Malondialdehyde (MDA) in Liver Tissue
4.8. Transmission Electron Microscopy (TEM)
4.9. Western Blot Analysis
4.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| NAFLD | non-alcoholic fatty liver disease |
| MMO | Meretrix meretrix oligopeptides |
| ALT | alanine aminotransferase |
| AST | aspartate transaminase |
| MDA | malonic dialdehyde |
| SOD | superoxide dismutase |
| GSH-Px | glutathione peroxidase |
| SDS | sodium dodecyl sulfate |
| TNF-α | tumor necrosis factor-α |
| NF-κB | nuclear factor kappa B |
| SREBP-1c | sterol regulatory element-binding protein 1c |
| PPAR-α | peroxisome proliferator-activated receptor α |
| Bax | Bcl-2 associated X protein |
| Bcl-2 | B-cell lymphoma-2 |
| Caspase-3 | cysteinyl aspartate specific proteinase -3 |
| NC | normal control diet |
| HFD | high-fat diet |
| H&E | hematoxylin and eosin |
| TEM | transmission electron microscopy |
References
- Petts, G.; Lloyd, K.; Goldin, R. Fatty liver disease. Diagn. Histopathol. 2014, 20, 102–108. [Google Scholar] [CrossRef]
- Berlanga, A.; Guiu-Jurado, E.; Porras, J.A.; Auguet, T. Molecular pathways in non-alcoholic fatty liver disease. Clin. Exp. Gastroenterol. 2014, 7, 221–239. [Google Scholar] [PubMed]
- Cave, M.; Deaciuc, I.; Mendez, C.; Song, Z.; Joshi-Barve, S.; Barve, S.; McClain, C. Nonalcoholic fatty liver disease: Predisposing factors and the role of nutrition. J. Nutr. Biochem. 2007, 18, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Almobarak, A.O.; Barakat, S.; Khalifa, M.H.; Elhoweris, M.H.; Elhassan, T.M.; Ahmed, M.H. Non alcoholic fatty liver disease (NAFLD) in a Sudanese population: What is the prevalence and risk factors? Arab J. Gastroenterol. 2014, 15, 12–15. [Google Scholar] [CrossRef] [PubMed]
- Fazel, Y.; Koenig, A.B.; Sayiner, M.; Goodman, Z.D.; Younossi, Z.M. Epidemiology and natural history of non-alcoholic fatty liver disease. Metab. Clin. Exp. 2016, 65, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zheng, L.; Dong, D.; Xu, L.; Yin, L.; Qi, Y.; Han, X.; Lin, Y.; Liu, K.; Peng, J. Effects of flavonoids from Rosa laevigata Michx fruit against high-fat diet-induced non-alcoholic fatty liver disease in rats. Food Chem. 2013, 141, 2108–2116. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Wu, Z.F.; Jiang, C.H.; Zhang, Q.W.; Ouyang, S.; Che, C.T.; Zhang, J.; Yin, Z.Q. The chloroform extract of Cyclocarya paliurus attenuates high-fat diet induced non-alcoholic hepatic steatosis in Sprague Dawley rats. Phytomedicine 2016, 23, 1475–1483. [Google Scholar] [CrossRef] [PubMed]
- Rolo, A.P.; Teodoro, J.S.; Palmeira, C.M. Role of oxidative stress in the pathogenesis of nonalcoholic steatohepatitis. Free Radic. Biol. Med. 2012, 52, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Qiang, X.; Xu, L.; Zhang, M.; Zhang, P.; Wang, Y.; Wang, Y.; Zhao, Z.; Chen, H.; Liu, X.; Zhang, Y. Demethyleneberberine attenuates non-alcoholic fatty liver disease with activation of AMPK and inhibition of oxidative stress. Biochem. Biophys. Res. Commun. 2016, 472, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Dowman, J.K.; Armstrong, M.J.; Tomlinson, J.W.; Newsome, P.N. Current therapeutic strategies in non-alcoholic fatty liver disease. Diabetes Obes. Metab. 2011, 13, 692–702. [Google Scholar] [CrossRef] [PubMed]
- Suleria, H.A.R.; Gobe, G.; Masci, P.; Osborne, S.A. Marine bioactive compounds and health promoting perspectives; innovation pathways for drug discovery. Trends Food Sci. Technol. 2016, 50, 44–55. [Google Scholar] [CrossRef]
- Mendis, E.; Rajapakse, N.; Byun, H.G.; Kim, S.K. Investigation of jumbo squid (Dosidicus gigas) skin gelatin peptides for their in vitro antioxidant effects. Life Sci. 2005, 77, 2166–2178. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.-J.; Jung, W.-K.; Byun, H.-G.; Kim, S.-K. Protective effect of an antioxidative peptide purified from gastrointestinal digests of oyster, Crassostrea gigas against free radical induced DNA damage. Bioresour. Technol. 2008, 99, 3365–3371. [Google Scholar] [CrossRef] [PubMed]
- Bai Renao, M.Y.; He, C.; Wei, H.; Yang, Q.; Wang, Y.; Xiong, H. Optimization of Enzymatic Hydrolysis of Meretrix meretrix Linnaeus. Protein and Analysis of the Antioxidant Activity of the Hydrolysates in vitro. Chin. Agric. Sci. Bull. 2013, 29, 44–49. [Google Scholar]
- Wu, J.; Xu, S.; Luo, S.; Fang, H. Antitumor activity of Meretrix meretrix glycopeptides MGP0501 in vivo. J. Nanchang Univ. (Nat. Sci.) 2014, 38, 492–497. [Google Scholar]
- Huang, F.; Zhao, S.; Yu, F.; Yang, Z.; Ding, G. Protective Effects and Mechanism of Meretrix meretrix Oligopeptides against Nonalcoholic Fatty Liver Disease. Mar. Drugs 2017, 15, 31. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhao, Y.; Zhang, Y.; Yang, Z.; Huang, F.; Ding, G. Preparation of NAFLD Cell Model Repair Polypeptide from Meretrix meretrix. J. Chin. Inst. Food Sci. Technol. 2017, 17, 33–39. [Google Scholar]
- Braud, L.; Battault, S.; Meyer, G.; Nascimento, A.; Gaillard, S.; de Sousa, G.; Rahmani, R.; Riva, C.; Armand, M.; Maixent, J.M.; et al. Antioxidant properties of tea blunt ROS-dependent lipogenesis: Beneficial effect on hepatic steatosis in a high fat-high sucrose diet NAFLD obese rat model. J. Nutr. Biochem. 2017, 40, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Hong, J.; Jeon, R.; Kim, H.S. Adzuki bean ameliorates hepatic lipogenesis and proinflammatory mediator expression in mice fed a high-cholesterol and high-fat diet to induce nonalcoholic fatty liver disease. Nutr. Res. 2016, 36, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Riordan, J.D.; Nadeau, J.H. Modeling progressive non-alcoholic fatty liver disease in the laboratory mouse. Mamm. Genome 2014, 25, 473–486. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.; Terauchi, Y. Lessons from Mouse Models of High-Fat Diet-Induced NAFLD. Int. J. Mol. Sci. 2013, 14, 21240–21257. [Google Scholar] [CrossRef] [PubMed]
- Preethi, J.; Vennila, K.; Penislusshiyan, S.; Velvizhi, S. Hepatoprotective and Antioxidant Role of Ziziphus jujuba Leaves on Paracetamol Induced Hepatic Damage in Rats. J. Dis. Med. Plants 2016, 2, 1–10. [Google Scholar]
- Yan, Z.; Fan, R.; Yin, S.; Zhao, X.; Liu, J.; Li, L.; Zhang, W.; Ge, L. Protective effects of Ginkgo biloba leaf polysaccharide on nonalcoholic fatty liver disease and its mechanisms. Int. J. Biol. Macromol. 2015, 80, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Mota, M.; Banini, B.A.; Cazanave, S.C.; Sanyal, A.J. Molecular mechanisms of lipotoxicity and glucotoxicity in nonalcoholic fatty liver disease. Metab. Clin. Exp. 2016, 65, 1049–1061. [Google Scholar] [CrossRef] [PubMed]
- Hlavinka, K. The Role of Reactive Oxygen Species in Insulin Resistance. Master’s Thesis, University of North Carolina at Greensboro, Greensboro, NC, USA, 2013. [Google Scholar]
- Ahmad, S.S.; Najmi, A.K.; Kaundal, M.; Akhtar, M. Gastroprotective Effect of Thymoquinone on Water Immersion Restraint Stress Induced Ulceration in Rats. Drug Res. 2017, 67, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Sha, F.; Chang, Y.Q.; Ding, J. Effects of two cooling modes of low temperature stress on antioxidant enzyme activities and malondiadehyde level in sea cucumber Apostichopus japonicus. J. Dalian Ocean Univ. 2015, 1, 25–29. [Google Scholar]
- Mukherjee, S.; Ghosh, S.; Choudhury, S.; Adhikary, A.; Manna, K.; Dey, S.; Sa, G.; Das, T.; Chattopadhyay, S. Pomegranate reverses methotrexate-induced oxidative stress and apoptosis in hepatocytes by modulating Nrf2-NF-κB pathways. J. Nutr. Biochem. 2013, 24, 2040–2050. [Google Scholar] [CrossRef] [PubMed]
- Perluigi, M.; Coccia, R.; Butterfield, D.A. 4-Hydroxy-2-Nonenal, a Reactive Product of Lipid Peroxidation, and Neurodegenerative Diseases: A Toxic Combination Illuminated by Redox Proteomics Studies. Antioxid. Redox Signal. 2012, 17, 1590–1609. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Gupta, S.C.; Kim, J.H. Historical perspectives on tumor necrosis factor and its superfamily: 25 years later, a golden journey. Blood 2012, 119, 651–665. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhang, H.; Wang, Y.; Song, F.; Yuan, Y. Inhibitory effects of quercetin on the progression of liver fibrosis through the regulation of NF-small ka, CyrillicB/Ismall ka, CyrillicBalpha, p38 MAPK, and Bcl-2/Bax signaling. Int. Immunopharmacol. 2017, 47, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.M.; Mann, D.A. Role of nuclear factor κB in liver health and disease. Clin. Sci. 2010, 118, 691–705. [Google Scholar] [CrossRef] [PubMed]
- Levy, M.; Claxton, D. Therapeutic Inhibition of BCL-2 and Related Family Members. Expert Opin. Investig. Drugs 2017, 26, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Wang, S.; Han, S.; Xie, K.; Wang, Y.; Li, J.; Liu, Y. Plant Bax Inhibitor-1 Interacts with ATG6 to Regulate Autophagy and Programmed Cell Death. Autophagy 2017, 13, 1161–1175. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Bi, X.W.; Li, P.F.; Xia, Z.J.; Huang, H.Q.; Jiang, W.Q.; Zhang, Y.J.; Wang, L. Overexpression of MYC and BCL2 Predicts Poor Prognosis in Patients with Extranodal NK/T-cell Lymphoma, Nasal Type. J. Cancer 2017, 8, 793–800. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Dose/(mg/kg) | Initial Weight/g | Final Weight/g | Body Weight Gain/g |
|---|---|---|---|---|
| NC | - | 22.86 ± 1.71 | 32.41 ± 2.68 * | 9.55 ± 2.02 * |
| HC (HFD) | - | 22.43 ± 1.76 | 37.69 ± 2.27 # | 15.27 ± 3.11 # |
| BFD (HFD + Bifendate) | 200 | 21.81 ± 2.20 | 31.83 ± 3.35 * | 10.03 ± 2.29 * |
| L-MMO (HFD + MMO) | 50 | 20.48 ± 2.31 | 32.89 ± 1.81 * | 12.41 ± 2.31 # |
| H-MMO (HFD MMO) | 250 | 22.26 ± 1.83 | 33.10 ± 1.15 * | 10.85 ± 1.83 * |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, F.; Wang, J.; Yu, F.; Tang, Y.; Ding, G.; Yang, Z.; Sun, Y. Protective Effect of Meretrix meretrix Oligopeptides on High-Fat-Diet-Induced Non-Alcoholic Fatty Liver Disease in Mice. Mar. Drugs 2018, 16, 39. https://doi.org/10.3390/md16020039
Huang F, Wang J, Yu F, Tang Y, Ding G, Yang Z, Sun Y. Protective Effect of Meretrix meretrix Oligopeptides on High-Fat-Diet-Induced Non-Alcoholic Fatty Liver Disease in Mice. Marine Drugs. 2018; 16(2):39. https://doi.org/10.3390/md16020039
Chicago/Turabian StyleHuang, Fangfang, Jiajia Wang, Fangmiao Yu, Yunping Tang, Guofang Ding, Zuisu Yang, and Yu Sun. 2018. "Protective Effect of Meretrix meretrix Oligopeptides on High-Fat-Diet-Induced Non-Alcoholic Fatty Liver Disease in Mice" Marine Drugs 16, no. 2: 39. https://doi.org/10.3390/md16020039