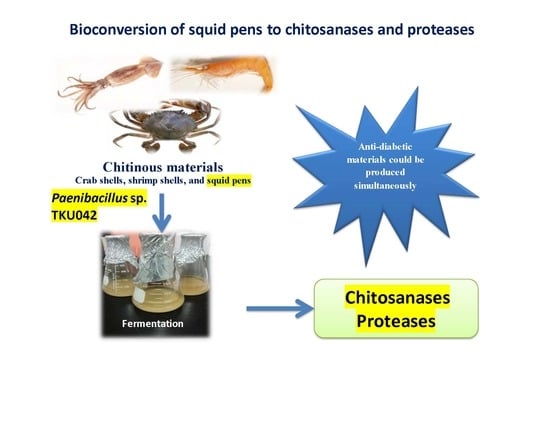
Conversion of Squid Pens to Chitosanases and Proteases via Paenibacillus sp. TKU042

 ,
, 
 ,
,  and
and 
Abstract
:
1. Introduction
2. Results and Discussion
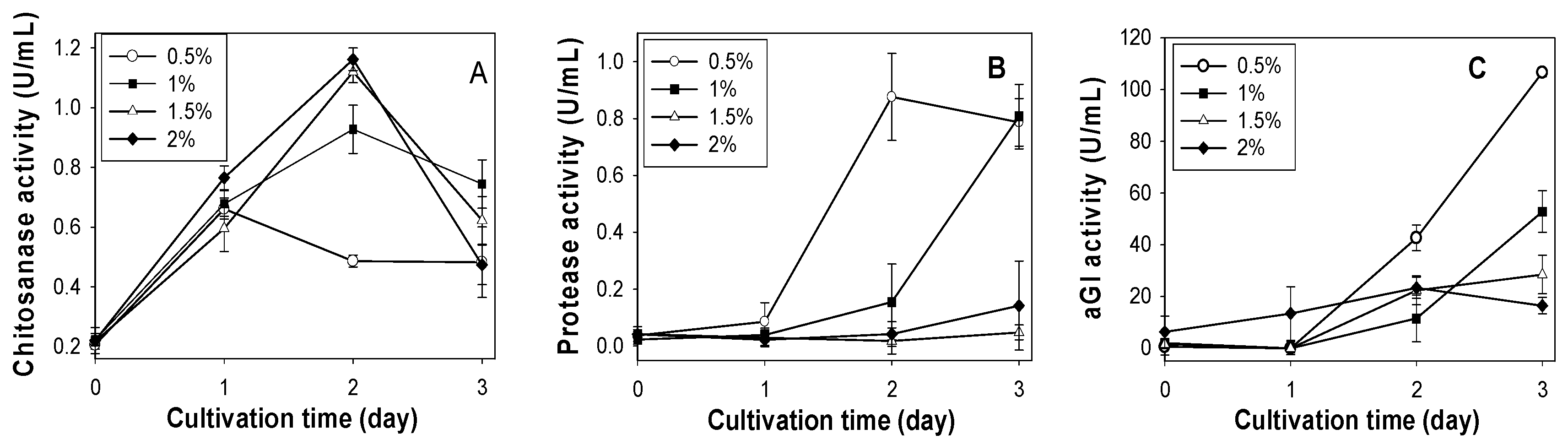
2.1. Screening of Chitinous Materials as Sole C/N for Chitosanase Production
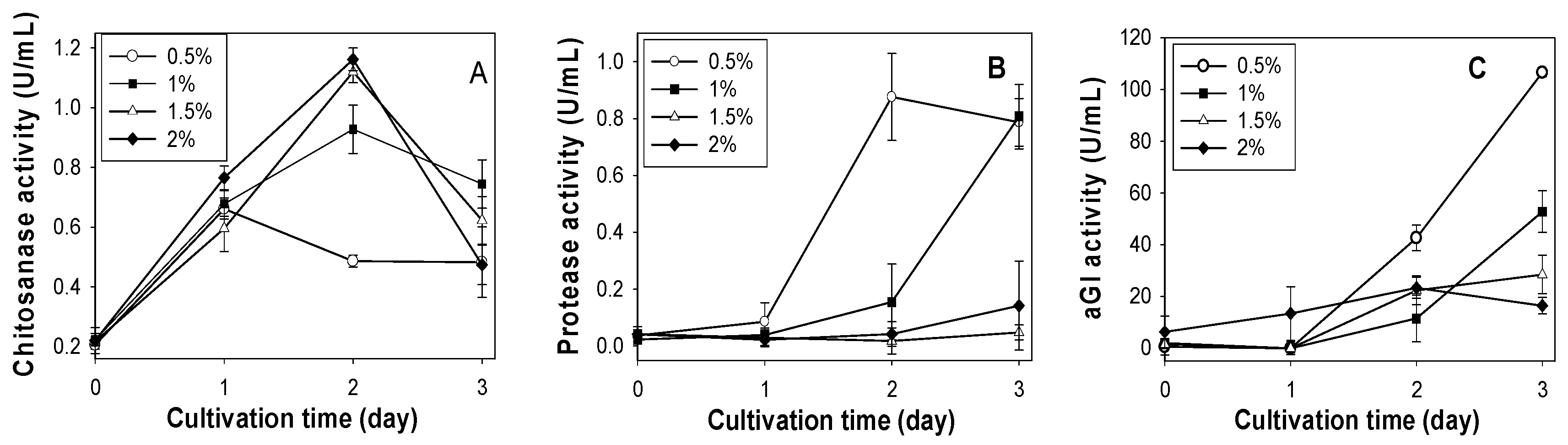
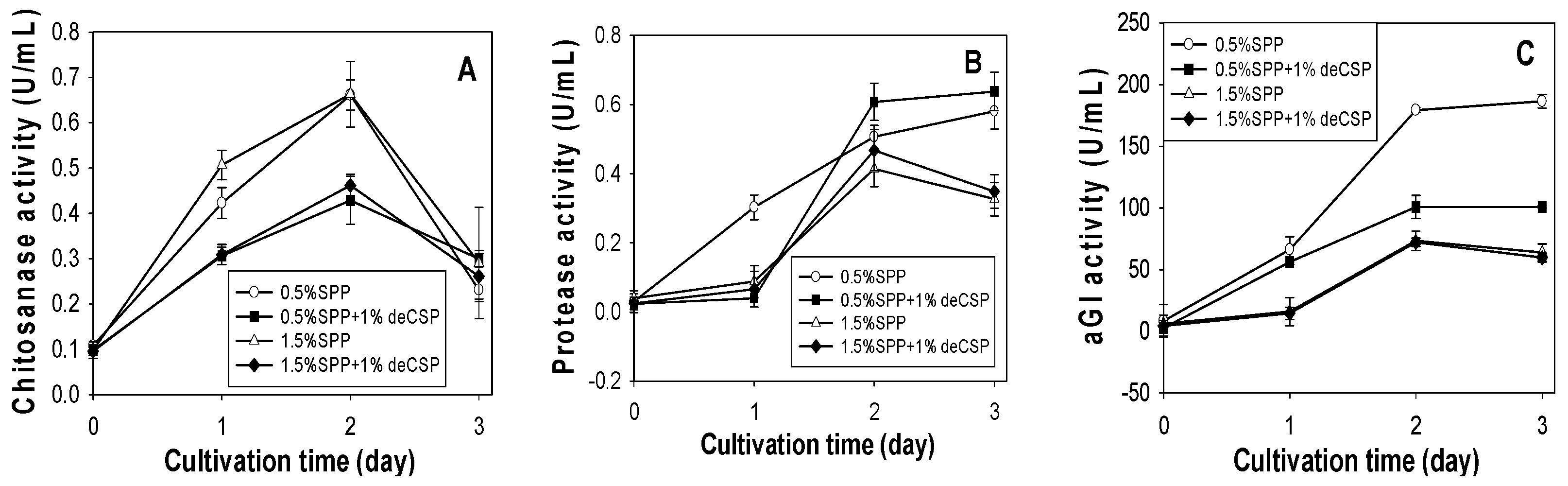
2.2. Effect of SPP Concentration on Chitosanase, Protease and αGI Production
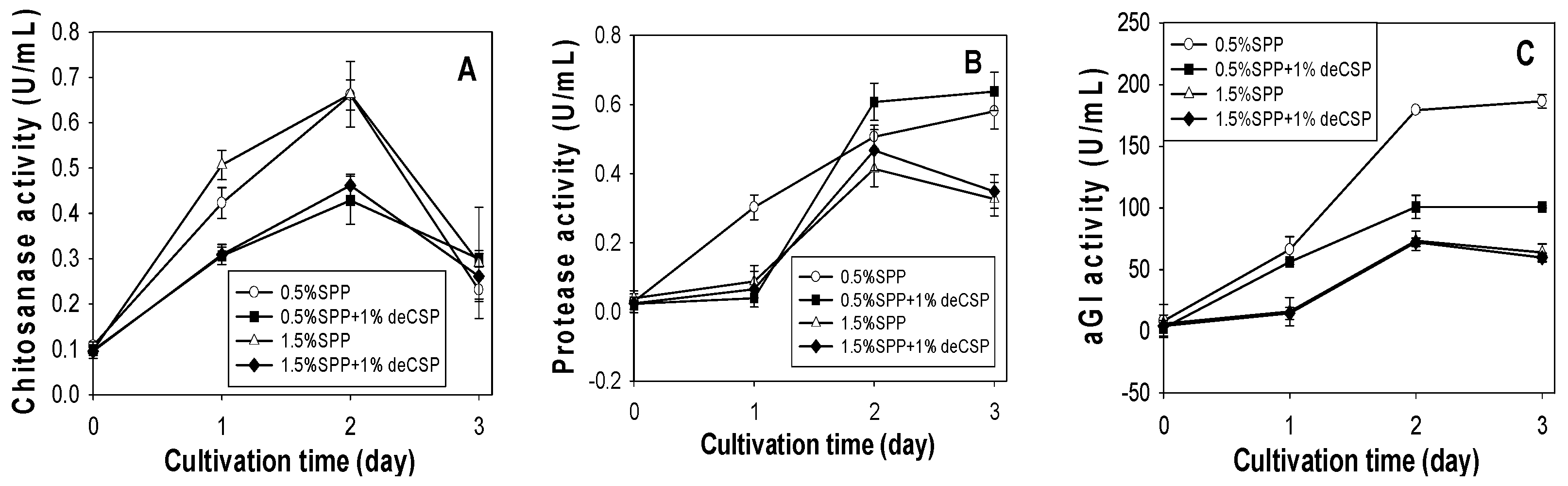
2.3. Production of Chitosanase, Protease and αGI from SPP and deCSP by Different Bacteria
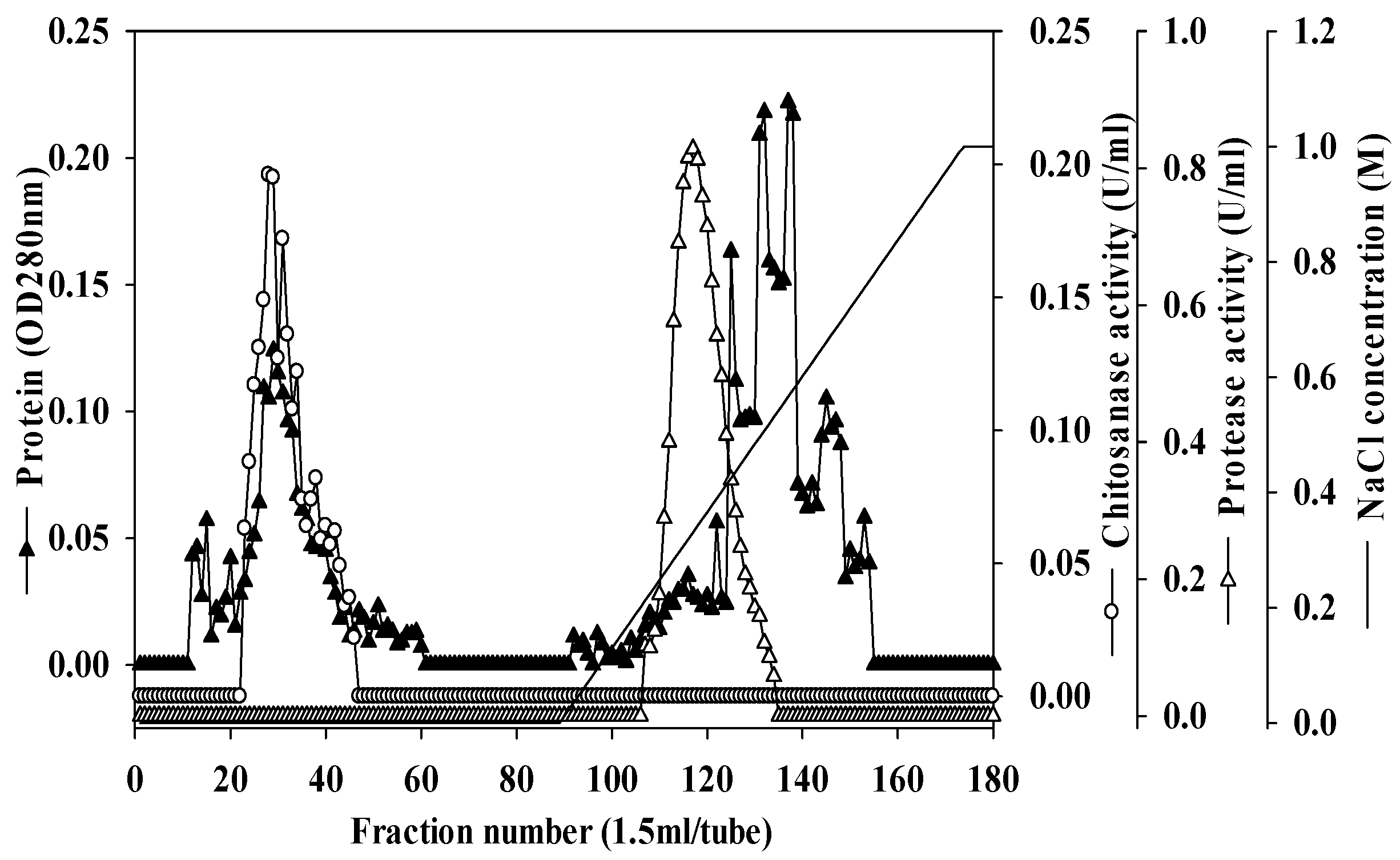
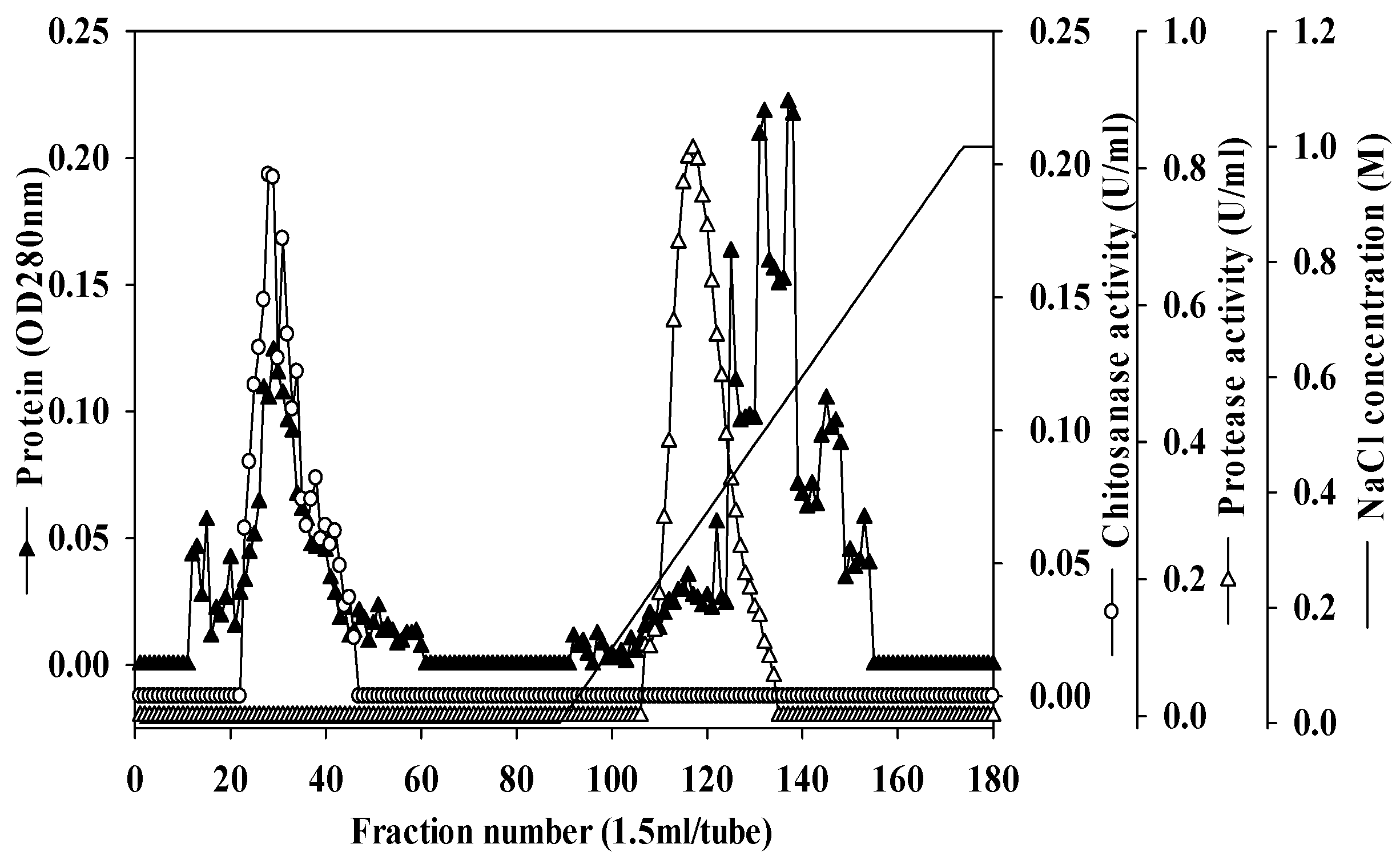
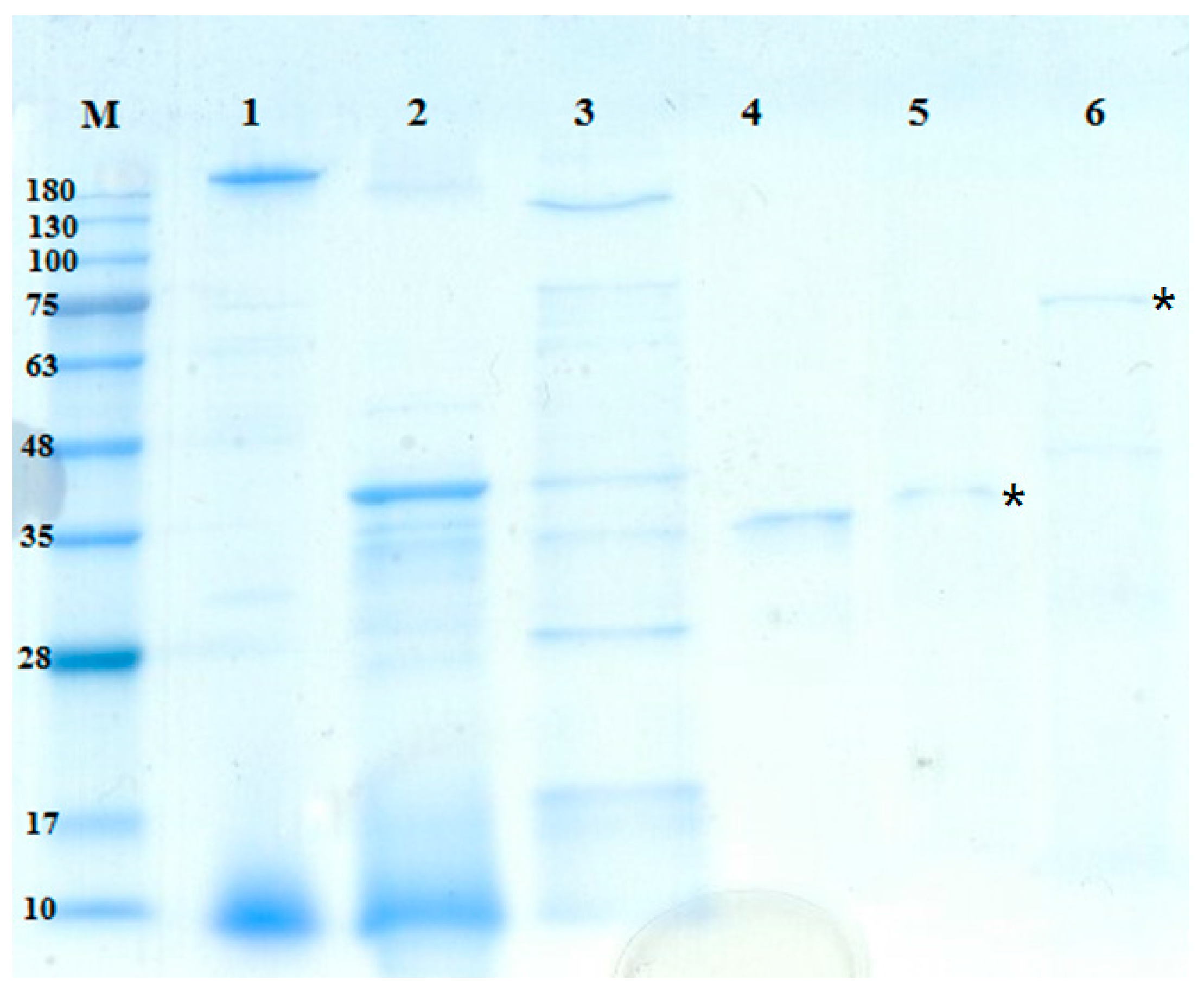
2.4. Purification and Characterization of Chitosanase and Protease
3. Materials and Methods
3.1. Materials
3.2. Measurement of Enzyme Activities
3.2.1. Chitosanase Activity
3.2.2. Protease Activity
3.2.3. Chitinase Activity
3.2.4. Exochitinase Activity
3.3. Measurement of Alpha Glucosidase Inhibitor
3.4. Screening of Chitinous Materials as Sole C/N for Enzyme Activity
3.5. Effect of SPP Concentration on Enzymes and αGI Activity
3.6. Production of Enzymes and αGI from SPP and deCSP Using Different Bacteria
3.7. Purification of Chitosanase and Protease
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wang, S.L.; Liang, T. Microbial reclamation of squid pens and shrimp shells. Res. Chem. Intermed. 2017, 43, 3445–3462. [Google Scholar] [CrossRef]
- Wang, S.L. Microbial reclamation of squid pen. Biocatal. Agric. Biotechnol. 2012, 1, 177–180. [Google Scholar] [CrossRef]
- Kaur, S.; Dhillon, G.S. Recent trends in biological extraction of chitin from marine shell wastes: A review. Crit. Rev. Biotechnol. 2015, 35, 44–61. [Google Scholar] [CrossRef] [PubMed]
- Younes, I.; Rinaudo, M. Chitin and chitosan preparation from marine sources. Structure, properties and applications. Mar. Drugs 2015, 13, 1133–1174. [Google Scholar] [CrossRef] [PubMed]
- Younes, I.; Hajji, S.; Rinaudo, M.; Chaabouni, M.; Jellouli, K.; Nasri, M. Optimization of proteins and minerals removal from shrimp shells to produce highly acetylated chitin. Int. J. Biol. Macromol. 2016, 84, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Ghorbel-Bellaaj, O.; Jellouli, K.; Younes, I.; Manni, L.; Ouled Salem, M.; Nasri, M. A solvent-stable metalloprotease produced by Pseudomonas aeruginosa A2 grown on shrimp shell waste and its application in chitin extraction. Appl. Biochem. Biotechnol. 2011, 164, 410–425. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.W.; Chen, Y.Y.; Pan, P.S.; Wang, S.L. Purification of chitinase/chitosanase from Bacillus cereus and discovery of an enzyme inhibitor. Int. J. Biol. Macromol. 2014, 63, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.W.; Lo, B.C.; Wang, S.L. Chitinolytic bacteria-assisted conversion of squid pen and its effect on dyes and adsorption. Mar. Drugs 2015, 13, 4576–4593. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.L.; Su, J.W.; Liang, T.W.; Nguyen, A.D.; Wang, S.L. Production, purification and characterization of a chitosanase from Bacillus cereus. Res. Chem. Intermed. 2014, 40, 2237–2248. [Google Scholar] [CrossRef]
- Liang, T.W.; Chen, W.T.; Lin, Z.H.; Kuo, Y.H.; Nguyen, A.D.; Pan, P.S.; Wang, S.L. An amphiprotic novel chitosanase from Bacillus mycoides and its application in the production of chitooligomers with their antioxidant and anti-inflammatory evaluation. Int. J. Mol. Sci. 2016, 17, 1302. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.W.; Jen, S.N.; Nguyen, A.D.; Wang, S.L. Application of chitinous materials in production and purification of a poly (L-lactic acid) depolymerase from Pseudomonas tamsuii TKU015. Polymers 2016, 8, 98. [Google Scholar] [CrossRef]
- Wang, S.L.; Chen, S.J.; Liang, T.W.; Lin, Y.D. A novel nattokinase produced by Pseudomonas sp. TKU015 using shrimp shells as substrate. Process Biochem. 2009, 44, 70–76. [Google Scholar] [CrossRef]
- Kuo, Y.H.; Liang, T.W.; Liu, K.C.; Hsu, Y.W.; Hsu, H.; Wang, S.L. Isolation and identification of a novel antioxidant with antitumor activity from Serratia ureilytica using squid pen as fermentation substrate. Mar. Biotechnol. 2011, 13, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Li, H.T.; Zhang, L.J.; Lin, Z.H.; Kuo, Y.H. Conversion of squid pen to homogentisic acid via Paenibacillus sp. TKU036 and the antioxidant and anti-inflammatory activities of homogentisic acid. Mar. Drugs 2016, 14, 183. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Wu, P.C.; Liang, T.W. Utilization of squid pen for the efficient production of chitosanase and antioxidants through prolonged autoclave treatment. Carbohydr. Res. 2009, 244, 979–984. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.B.; Nguyen, A.D.; Wang, S.L. Utilization of fishery processing by product squid pens for Paenibacillus sp. fermentation on producing potent α-glucosidase inhibitors. Mar. Drugs 2017, 15, 274. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.B.; Wang, S.L. Reclamation of marine chitinous materials for the production of α-glucosidase inhibitors via microbial conversion. Mar. Drugs 2017, 15, 350. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.B.; Wang, S.L. New novel α-glucosidase inhibitors produced by microbial conversion. Process Biochem. 2017, 65, 228–232. [Google Scholar] [CrossRef]
- Wang, S.L.; Huang, T.Y.; Wang, C.Y.; Liang, T.W.; Yen, Y.H.; Sakata, Y. Bioconversion of squid pen by Lactobacillus paracasei subsp paracasei TKU010 for the production of proteases and lettuce enhancing biofertilizers. Bioresour. Technol. 2008, 99, 5436–5443. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Hsu, W.H.; Liang, T.W. Conversion of squid pen by Pseudomonas aeruginosa K-187 fermentation for the production of N-acetyl chitooligosaccharides and biofertilizers. Carbohydr. Res. 2010, 345, 880–885. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.W.; Wu, C.C.; Cheng, W.T.; Chen, Y.C.; Wang, C.L.; Wang, I.L.; Wang, S.L. Exopolysaccharides and antimicrobial biosurfactants produced by Paenibacillus macerans TKU029. Appl. Biochem. Biotechnol. 2014, 172, 933–950. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.W.; Tseng, S.C.; Wang, S.L. Production and characterization of antioxidant properties of exopolysaccharides from Paenibacillus mucilaginosus TKU032. Mar. Drugs 2016, 14, 40. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.W.; Wang, S.L. Recent advances in exopolysaccharides from Paenibacillus spp.: Production, isolation, structure, and bioactivities. Mar. Drugs 2015, 13, 1847–1863. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Wang, C.Y.; Yen, Y.H.; Liang, T.W.; Chen, S.Y.; Chen, C.H. Enhanced production of insecticidal prodigiosin from Serratia marcescens TKU011 in media containing squid pen. Process Biochem. 2012, 47, 1684–1690. [Google Scholar] [CrossRef]
- Hsu, C.H.; Nguyen, A.D.; Chen, Y.W.; Wang, S.L. Tyrosinase inhibitors and insecticidal materials produced by Burkholderia cepacia using squid pen as the sole carbon and nitrogen source. Res. Chem. Intermed. 2014, 40, 2249–2258. [Google Scholar] [CrossRef]
- Grady, E.N.; MacDonald, J.; Liu, L.; Richman, A.; Yuan, Z.C. Current knowledge and perspectives of Paenibacillus: A review. Microb. Cell Fact. 2016, 15, 203. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.P.; Yamaki, K.; Yoshihashi, T.; Ohnishi, K.M.; Li, X.T.; Cheng, Y.Q.; Mori, Y.; Li, L.T. Purification and identification of 1-deoxynojirimycin (DNJ) in okara fermented by Bacillus subtilis B2 from Chinese traditional food (meitaoza). J. Agric. Food Chem. 2010, 58, 4097–4103. [Google Scholar] [CrossRef] [PubMed]
- Nam, H.; Jung, H.; Karuppasamy, S.; Park, Y.S.; Cho, Y.S.; Lee, J.Y.; Seong, S.; Suh, J.G. Anti-diabetic effect of the soybean extract fermented by Bacillus subtilis MORI in db/db mice. Food Sci. Biotechnol. 2012, 21, 1669–1676. [Google Scholar] [CrossRef]
- Wang, C.L.; Huang, T.H.; Liang, T.W.; Wang, S.L. Production and characterization of exopolysaccharides and antioxidant from Paenibacillus sp. TKU023. New Biotechnol. 2011, 28, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Luo, J.; Ye, H.; Sun, Y.; Lu, Z.; Zeng, X. Production, characterization and antioxidant activities in vitro of exopolysaccharides from endophytic bacterium Paenibacillus polymyxa EJS-3. Carbohydr. Polym. 2009, 78, 275–281. [Google Scholar] [CrossRef]
- Fang, Y.; Liu, S.; Lu, M.; Jiao, Y.; Wang, S. A novel method for promoting antioxidant exopolysaccharides production of Bacillus licheniformis. Carbohydr. Polym. 2013, 92, 1172–1176. [Google Scholar] [CrossRef] [PubMed]
- Ash, C.; Priest, F.G.; Collins, M.D. Molecular identification of rRNA group 3 bacilli (Ash, Farrow, Wallbanks and Collins) using a PCR probe test. Proposal for the creation of a new genus Paenibacillus. Antonie Van Leewenhoek 1993, 64, 253–260. [Google Scholar] [CrossRef]
- Pagnoncelli, M.G.B.; de Araújo1, N.K.; da Silva, N.M.P.; de Assis, C.F.; Rodrigues, S.; de Macedo, G.B. Chitosanase production by Paenibacillus ehimensis and its application for chitosan hydrolysis. Braz. Arch. Biol. Technol. 2010, 53, 1461–1468. [Google Scholar] [CrossRef]
- Omumasaba, C.A.; Yoshida, N.; Sekiguchi, Y.; Kariya, K.; Ogawa, K. Purification and some properties of a novel chitosanase from Bacillus subtilis KH1. J. Gen. Appl. Microbiol. 2000, 46, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, K.; Fujita, T.; Kuroshima, K.; Sakane, T.; Yokota, A.; Takata, R. Purification and gene cloning of a chitosanase from Bacillus ehimensis EAG1. J. Biosci. Bioeng. 1999, 87, 383–385. [Google Scholar] [CrossRef]
- Chiang, C.L.; Chang, C.T.; Sung, H.Y. Purification and properties of chitosanase from a mutant of Bacillus subtilis IMR-NK1. Enzyme Microb. Technol. 2003, 32, 260–267. [Google Scholar] [CrossRef]
- Yilmaz, B.; Baltaci, M.O.; Sisecioglu, M.; Adiguzel, A. Thermotolerant alkaline protease enzyme from Bacillus licheniformis A10: Purification, characterization, effects of surfactants and organic solvents. J. Enzyme Inhib. Med. Chem. 2016, 31, 1241–1247. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, A.; Sygusch, J. Purification and characterization of three chitosanase activities from Bacillus megaterium P1. Appl. Environ. Microbiol. 1990, 56, 844–848. [Google Scholar] [PubMed]
- Kurakake, M.; You, S.; Nakagawa, K.; Sugihara, M.; Komaki, T. Properties of chitosanase from Bacillus cereus S1. Curr. Microbiol. 2000, 40, 6–9. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Kim, E.J.; Piao, Z.; Yun, Y.C.; Shin, Y.C. Purification and characterization of chitosanase from Bacillus sp. strain KCTC 0377BP and its application for the production of chitosan oligosaccharides. Appl. Environ. Microbiol. 2004, 70, 4522–4531. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.Y.; Jo, K.J.; Jin, Y.L.; Kim, K.Y.; Shin, J.H.; Kim, Y.W.; Park, R.D. Characterization and Kinetics of 45 kDa Chitosanase from Bacillus sp. P16. Biosci. Biotechnol. Biochem. 2003, 67, 1875–1882. [Google Scholar] [CrossRef] [PubMed]
- Kim, P.I.; Kang, T.H.; Chung, K.J.; Kim, I.S.; Chung, K.C. Purification of a constitutive chitosanase produced by Bacillus sp. MET 1299 with cloning and expression of the gene. FEMS Microbiol. Lett. 2004, 240, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Zitouni, M.; Fortin, M.; Scheerle, R.K.; Letzel, T.; Matteau, D.; Rodrigue, S.; Brzezinski, R. Biochemical and molecular characterization of a thermostable chitosanase produced by the strain Paenibacillus sp. 1794 newly isolated from compost. Appl. Microbiol. Biotechnol. 2013, 97, 5801–5813. [Google Scholar] [CrossRef] [PubMed]
- Kimoto, H.; Kusaoke, H.; Yamamoto, I.; Fujii, Y. Biochemical and genetic properties of Paenibacillus glycosyl hydrolase having chitosanase activity and discoidin domain. J. Biol. Chem. 2002, 17, 14695–14702. [Google Scholar] [CrossRef] [PubMed]
- Rai, S.K.; Roy, J.K.; Mukherjee, A.K. Characterisation of a detergent-stable alkaline protease from a novel thermophilic strain Paenibacillus tezpurensis sp. nov. AS-S24-II. Appl. Microbiol. Biotechnol. 2010, 85, 1437–1450. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Pan, Y.; She, Q.; Chen, L. A novel carboxyl-terminal protease derived from Paenibacillus lautus CHN26 exhibiting high activities at multiple sites of substrates. BMC Biotechnol. 2013, 13, 89. [Google Scholar] [CrossRef] [PubMed]
- Mothe, T.; Sultanpuram, V.R. Production, purification and characterization of a thermotolerant alkaline serine protease from a novel species Bacillus caseinilyticus. 3 Biotech 2016, 6, 53. [Google Scholar] [CrossRef] [PubMed]
- Rahman, R.N.Z.A.; Razak, C.N.; Ampon, K.; Basri, M.; Yunus, W.M.Z.; Salleh, A.B. Purification and characterization of a heat-stable alkaline protease from Bacillus stearothermophilus Fl. Appl. Microbiol. Biotechnol. 1994, 40, 822–827. [Google Scholar] [CrossRef]
- Takami, H.; Akiba, T.; Horikoshi, K. Production of extremely thermostable alkaline protease from Bacillus sp. AH-101. Appl. Microbiol. Biotechnol. 1989, 30, 120–124. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Activity (U/mL) | ||||
|---|---|---|---|---|---|
| Exochitinase | Chitinase | Chitosanase | Protease | aGI | |
| Paenibacillus macerans TKU029 | - | 0.118 + 0.027 | 0.122 + 0.035 | 0.925 + 0.032 | 62.90 + 6.755 |
| P. mucilaginosus TKU032 | - | 0.072 + 0.016 | 0.086 + 0.021 | 1.366 + 0.045 | 178.38 + 10.312 |
| Paenibacillus sp. TKU037 | - | 0.135 + 0.018 | 0.069 + 0.019 | 1.203 + 0.041 | 180.77 + 11.620 |
| Paenibacillus sp. TKU042 | - | 0.182 + 0.036 | 0.088 + 0.015 | 1.257 + 0.038 | 185.45 + 3.085 |
| Bacillus licheniformis TKU004 | - | - | 0.089 + 0.022 | 0.121 + 0.023 | 91.29 + 8.692 |
| Bacillus subtilis TKU007 | - | 0.163 + 0.023 | - | 0.178 + 0.011 | 96.06 + 7.008 |
| Lactobacillus paracasei TKU010 | - | 0.216 + 0.033 | - | 0.113 + 0.009 | - |
| Serratia marcescens TKU011 | - | 0.201 + 0.029 | 0.149 + 0.021 | 0.115 + 0.012 | 73.63 + 4.090 |
| Serratia ureilytica TKU013 | 0.276 + 0.012 | 0.266 + 0.018 | 0.099 + 0.017 | 0.202 + 0.024 | 64.52 + 3.474 |
| Pseudomonas tamsuii TKU015 | - | 0.193 + 0.030 | 0.105 + 0.013 | 0.103 + 0.008 | 51.54 + 4.627 |
| Serratia sp. TKU016 | 0.152 + 0.021 | 0.104 + 0.012 | 0.110 + 0.022 | 0.101 + 0.007 | 68.962.292 |
| Serratia sp. TKU020 | 0.121 + 0.016 | 0.102 + 0.015 | 0.151 + 0.013 | 0.099 + 0.009 | 73.63 + 6.420 |
| Bacillus cereus TKU028 | - | 0.127 + 0.018 | 0.111 + 0.018 | 0.776 + 0.053 | 68.72 + 5.143 |
| Bacillus mycoildes TKU038 | - | - | 0.101 + 0.017 | 0.463 + 0.036 | 111.67 + 1.036 |
| Bacillus mycoildes TKU040 | - | 0.191 + 0.020 | - | - | - |
| Rhizobium sp.TKU041 | - | - | - | - | - |
| Strain | Activity (U/mL) | ||||
|---|---|---|---|---|---|
| Exochitinase | Chitinase | Chitosanase | Protease | aGI | |
| Paenibacillus macerans TKU029 | - | 0.376 + 0.026 | 0.857 + 0.015 | 0.135 ± 0.011 | 60.13 ± 6.844 |
| P. mucilaginosus TKU032 | - | 0.101 + 0.017 | 0.155 + 0.016 | 0.193 ± 0.005 | 175.40 ± 0.606 |
| Paenibacillus sp. TKU037 | - | 0.105 + 0.018 | 0.247 + 0.014 | 0.222 ± 0.004 | 175.80 ± 0.851 |
| Paenibacillus sp.TKU042 | - | 0.185 + 0.019 | 0.928 + 0.014 | 0.199 + 0.005 | 88.30 ± 4.502 |
| Bacillus licheniformis TKU004 | 1.211 + 0.017 | 0.074 + 0.017 | 0.120 + 0.014 | 0.707 + 0.020 | 174.59 ± 4.295 |
| Bacillus subtilis TKU007 | - | 0.118 + 0.017 | 0.314 + 0.017 | 0.219 + 0.005 | 58.64 ± 7.368 |
| Lactobacillus paracasei TKU010 | - | - | 0.166 + 0.015 | 0.090 + 0.010 | - |
| Serratia marcescens TKU011 | 8.039 + 0.296 | 0.177 + 0.018 | 0.150 + 0.013 | 0.477 + 0.010 | - |
| Serratia ureilytica TKU013 | 11.545 + 0.431 | 0.319 + 0.019 | 0.112 + 0.013 | 0.910 + 0.031 | 52.60 ± 8.418 |
| Pseudomonas tamsuii TKU015 | - | 0.105 + 0.015 | 0.056 + 0.013 | 0.113 + 0.009 | - |
| Serratia sp. TKU016 | - | 0.111 + 0.018 | 0.302 + 0.012 | 0.370 + 0.012 | - |
| Serratia sp. TKU020 | 9.327 + 0.325 | 0.201 + 0.018 | 0.118 + 0.014 | 0.478 + 0.021 | 52.49 ± 0.778 |
| Bacillus cereus TKU028 | - | 0.223 + 0.018 | 0.520 + 0.015 | 0.153 + 0.007 | 68.64 ± 3.972 |
| Bacillus mycoildes TKU038 | - | 0.332 + 0.019 | 0.611 + 0.016 | 0.240 + 0.004 | 71.46 ± 3.635 |
| Bacillus mycoildes TKU040 | - | 0.350 + 0.016 | 0.165 + 0.013 | 0.083 + 0.011 | - |
| Rhizobium sp.TKU041 | - | - | - | 0.198 + 0.012 | - |
| Step | Total Protein (mg) | Total Activity (U) | Specific Activity (U/mg) | Recovery (%) | Purification (Fold) |
|---|---|---|---|---|---|
| Cultural supernatant | 211.43 | 53.03 | 0.25 | 100.00% | 1.00 |
| Ethanol precipitation | 56.95 | 26.81 | 0.47 | 50.56% | 1.88 |
| Macro-Prep High S | 4.59 | 4.62 | 1.01 | 8.71% | 4.01 |
| Macro-Prep DEAE | 0.52 | 1.64 | 3.14 | 3.09% | 12.51 |
| Step | Total Protein (mg) | Total Activity (U) | Specific Activity (U/mg) | Recovery (%) | Purification (Fold) |
|---|---|---|---|---|---|
| Cultural supernatant | 211.43 | 86.90 | 0.41 | 100.00% | 1.00 |
| Ethanol precipitation | 56.95 | 60.35 | 1.06 | 69.45% | 2.58 |
| Macro-Prep High S | 2.72 | 17.10 | 6.29 | 19.68% | 15.30 |
| Sephacryl 200 | 0.20 | 7.56 | 38.19 | 8.70% | 92.91 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doan, C.T.; Tran, T.N.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.-L. Conversion of Squid Pens to Chitosanases and Proteases via Paenibacillus sp. TKU042. Mar. Drugs 2018, 16, 83. https://doi.org/10.3390/md16030083
Doan CT, Tran TN, Nguyen VB, Nguyen AD, Wang S-L. Conversion of Squid Pens to Chitosanases and Proteases via Paenibacillus sp. TKU042. Marine Drugs. 2018; 16(3):83. https://doi.org/10.3390/md16030083
Chicago/Turabian StyleDoan, Chien Thang, Thi Ngoc Tran, Van Bon Nguyen, Anh Dzung Nguyen, and San-Lang Wang. 2018. "Conversion of Squid Pens to Chitosanases and Proteases via Paenibacillus sp. TKU042" Marine Drugs 16, no. 3: 83. https://doi.org/10.3390/md16030083