Intraocular Penetration of a vNAR: In Vivo and In Vitro VEGF165 Neutralization

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
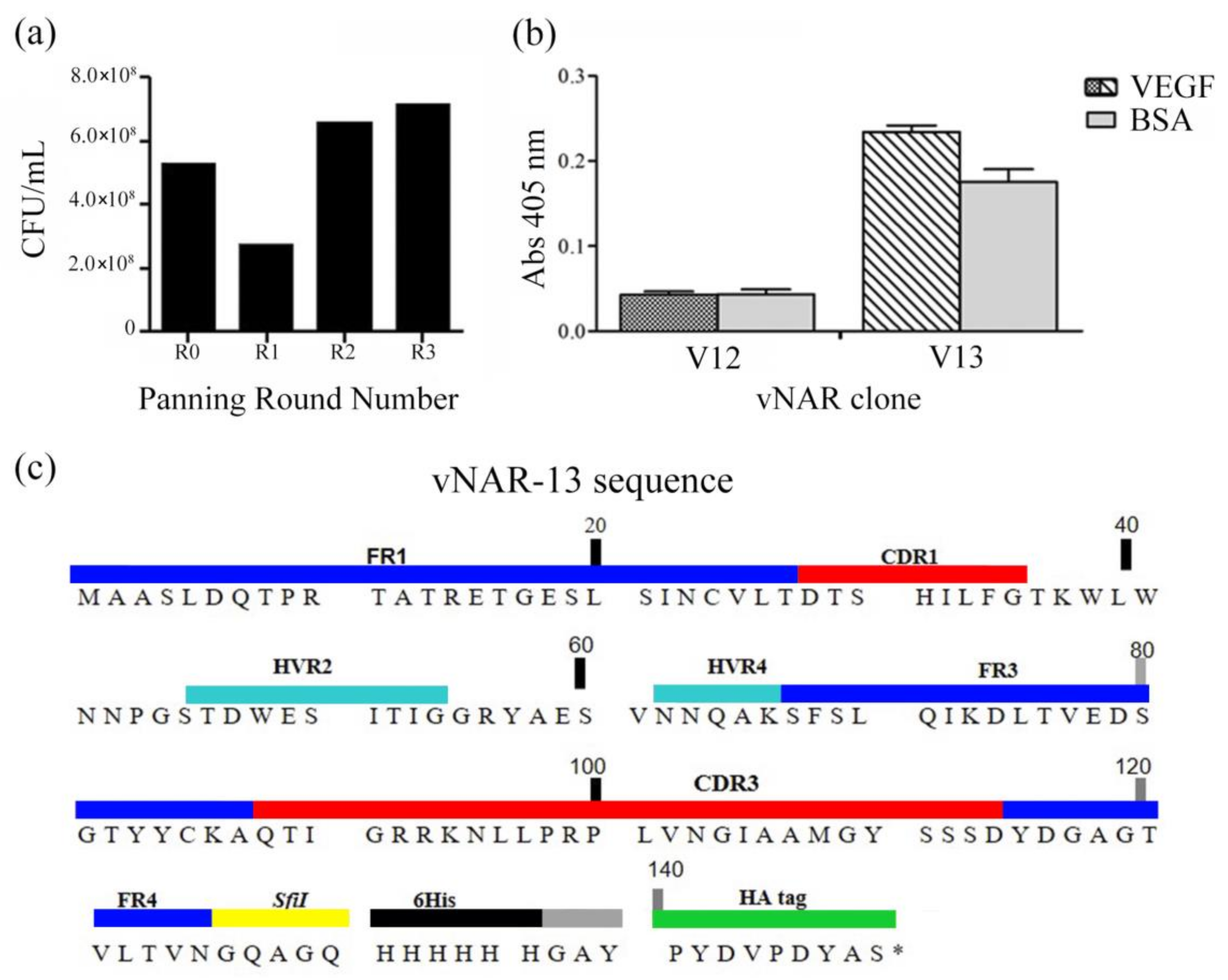
2.1. vNAR Phage Display and Sequence
2.2. Expression and Purification of vNAR from Inclusion Bodies
2.3. Angiogenesis Co-Culture Assay
2.4. Oxygen-Induced Retinopathy (OIR) Model
2.5. Intraocular Penetration
3. Discussion
4. Materials and Methods
4.1. vNAR Fragment Isolation from Immune Library
4.2. vNAR Expression and Purification
4.3. Refolding of V13 Protein and Western Blot
4.4. Co-Culture Angiogenesis Assay
4.5. Ethics Statements
4.6. Oxygen-Induced Retinopathy (OIR) Model
4.7. Intraocular Penetration of V13 by ELISA
5. Conclusions
6. Patents
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Boyer, D.; Hopkins, J.; Sorof, J.; Ehrlich, J. Anti-vascular endothelial growth factor therapy for diabetic macular edema. Ther. Adv. Endocrinol. Metab. 2013, 4, 151–169. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N.; Kerbel, R.S. Angiogenesis as a therapeutic target. Nature 2005, 438, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Miao, H.Q.; Hu, K.; Jimenez, X.; Navarro, E.; Zhang, H.; Lu, D.; Ludwig, D.L.; Balderes, P.; Zhu, Z. Potent neutralization of VEGF biological activities with a fully human antibody Fab fragment directed against VEGF receptor 2. Biochem. Biophys. Res. Commun. 2006, 345, 438–445. [Google Scholar] [CrossRef] [PubMed]
- Amadio, M.L.; Govoni, S.; Pascale, A. Targeting VEGF in eye neovascularization: what’s new?: a comprehensive review on current therapies and oligonucleotide-based interventions under development. Pharmacol. Res. 2016, 235–269. [Google Scholar] [CrossRef]
- Schmidt-Erfurth, U.; Chong, V.; Loewenstein, A.; Larsen, M.; Souied, E.; Schlingemann, R.; Eldem, B.; Monés, J.; Richard, G.; Bandello, F. Guidelines for the management of neovascular age-related macular degeneration by the European Society of Retina Specialists (EURETINA). Br. J. Ophthalmol. 2014, 98, 1144–1167. [Google Scholar] [CrossRef] [PubMed]
- Amadio, M.; Bucolo, C.; Leggio, G.M.; Drago, F.; Govoni, S.; Pascale, A. The PKCbeta/HuR/VEGF pathway in diabetic retinopathy. Biochem. Pharmacol. 2010, 80, 1230–1237. [Google Scholar] [CrossRef] [PubMed]
- Lupo, G.; Motta, C.; Giurdanella, G.; Anfuso, C.D.; Alberghina, M.; Drago, F.; Salomone, S.; Bucolo, C. Role of phospholipases A2 in diabetic retinopathy: In vitro and in vivo studies. Biochem. Pharmacol. 2013, 86, 1603–1613. [Google Scholar] [CrossRef] [PubMed]
- El-Asrar, A.M.A. Role of inflammation in the pathogenesis of diabetic retinopathy. Middle East Afr. J. Ophthalmol. 2012, 19, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Hanout, M.; Ferraz, D.; Ansari, M.; Maqsood, N.; Kherani, S.; Sepah, Y.J.; Rajagopalan, N.; Ibrahim, M.; Do, D.V.; Nguyen, Q.N. Therapies for neovascular age-related macular degeneration: Current approaches and pharmacologic agents in development. BioMed Res. Int. 2013, 2013, 830837. [Google Scholar] [CrossRef] [PubMed]
- Holz, F.G.; Schmitz-Valckenberg, S.; Fleckenstein, M. Recent developments in the treatment of age-related macular degeneration. J. Clin. Investig. 2014, 124, 1430–1438. [Google Scholar] [CrossRef] [PubMed]
- Bucolo, C.; Drago, F.; Salomone, S. Ocular drug delivery: A clue from nanotechnology. Front. Pharmacol. 2012, 3, 188. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, E.B.; Farah, M.E.; Maia, M.; Penha, F.M.; Regatieri, C.; Melo, G.B.; Pinheiro, M.M.; Zanetti, C.R. Therapeutic monoclonal antibodies in ophthalmology. Prog. Retin. Eye Res. 2009, 28, 117–144. [Google Scholar] [CrossRef] [PubMed]
- Ng, E.W.M.; Adamis, A.P. Targeting angiogenesis, the underlying disorder in neovascular age-related macular degeneration. Can. J. Ophthalmol. 2005, 40, 352–368. [Google Scholar] [CrossRef]
- Nguyen, D.; Luo, J.; Zhang, K.; Zhang, M. Current therapeutic approaches in neovascular age-related macular degeneration. Discov. Med. 2013, 15, 343–348. [Google Scholar] [PubMed]
- Michaelides, M.; Kaines, A.; Hamilton, R.D.; Fraser-Bell, S.; Rajendram, R.; Quhill, F.; Boos, C.J.; Xing, W.; Egan, C.; Peto, T.; et al. A prospective randomized trial of intravitreal bevacizumab or laser therapy in the management of diabetic macular edema (BOLT study) 12-month data: Report 2. Ophthalmology 2010, 117, 1078–1086.e2. [Google Scholar] [CrossRef] [PubMed]
- Thomas, B.J.; Shienbaum, G.; Boyer, D.S.; Flynn, H.W., Jr. Evolving strategies in the management of diabetic macular edema: Clinical trials and current management. Can. J. Ophthalmol. 2013, 48, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Georgopoulos, M.; Polak, K.; Prager, F.; Prünte, C.; Schmidt-Erfurth, U. Characteristics of severe intraocular inflammation following intravitreal injection of bevacizumab (Avastin). Br. J. Ophthalmol. 2009, 93, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.; Hollands, H.; Hollands, S.; Sharma, S. Incidence and characteristics of acute intraocular inflammation after intravitreal injection of bevacizumab: A retrospective cohort study. Can. J. Ophthalmol. 2010, 45, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Yu, S.; Liu, K.; Chen, F.E.; Song, Z.; Zhang, X.; Xu, X.; Sun, X. Acute intraocular inflammation caused by endotoxin after intravitreal injection of counterfeit bevacizumab in Shanghai, China. Ophthalmology 2013, 120, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C.H.; Holz, F.G. Preclinical aspects of anti-VEGF agents for the treatment of wet AMD: Ranibizumab and bevacizumab. Eye 2011, 25, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Semeraro, F.; Morescalchi, F.; Duse, S.; Gambicorti, E.; Romano, M.R.; Costagliola, C. Systemic thromboembolic adverse events in patients treated with intravitreal anti-VEGF drugs for neovascular age-related macular degeneration: An overview. Expert Opin. Drug Saf. 2014, 13, 785–802. [Google Scholar] [CrossRef] [PubMed]
- Kovaleva, M.; Ferguson, L.; Steven, J.; Porter, A.; Barelle, C. Shark variable new antigen receptor biologics a novel technology platform for therapeutic drug development. Expert Opin. Biol. Ther. 2014, 14, 1527–1539. [Google Scholar] [CrossRef] [PubMed]
- Zielonka, S.; Empting, M.; Grzeschik, J.; Könning, D.; Barelle, C.J.; Kolmar, H. Structural insights and biomedical potential of IgNAR scaffolds from sharks. mAbs 2015, 7, 15–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudson, P.; Souriau, C. Engineered antibodies. Nat. Med. 2003, 9, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Wesolowski, J.; Alzogaray, V.; Reyelt, J.; Unger, M.; Juarez, K.; Urrutia, M.; Cauerhff, A.; Danquah, W.; Rissiek, B.; Scheuplein, F.; et al. Single domain antibodies: Promising experimental and therapeutic tools in infection and immunity. Med. Microbiol. Immunol. 2009, 198, 157–174. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.R.; Saunders, K.; Grace, C.; Jin, M.; Piche-Nicholas, M.; Steven, J.; O’Dwyer, R.; Wu, L.; Khetemenee, L.; Vugmeyster, Y.; et al. Improving the pharmacokinetic properties of biologics by fusion to an anti-HSA shark vNAR domain. mAbs 2012, 4, 673–685. [Google Scholar] [CrossRef] [PubMed]
- Bojalil, R.; Mata-González, M.T.; Sánchez-Muñoz, F.; Yee, Y.; Argueta, I.; Bolaños, L.; Amezcua-Guerra, L.M.; Camacho-Villegas, T.A.; Sánchez-Castrejón, E.; García-Ubbelohde, W.J.; et al. Anti-tumor necrosis factor VNAR single domains reduce lethality and regulate underlying inflammatory response in a murine model of endotoxic shock. BMC Immunol. 2013, 14, 17. [Google Scholar] [CrossRef] [PubMed]
- Kopsidas, G.; Roberts, A.S.; Coia, G.; Streltsov, V.A.; Nuttall, S.D. In vitro improvement of a shark IgNAR antibody by Qb replicase mutation and ribosome display mimics in vivo affinity maturation. Immunol. Lett. 2006, 107, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, K.; Dolezal, O.; Parisi, K.; Angerosa, J.; Dogovski, C.; Barraclough, M.; Sanalla, M.; Casey, J.-L.; González, I.; Perugini, M.A.; et al. Shark variable new antigen receptor (vNAR) single domain antibody fragments: Stability and diagnostic applications. Antibodies 2013, 2, 66–81. [Google Scholar] [CrossRef]
- Liu, J.L.; Anderson, G.P.; Goldman, E.R. Isolation of antitoxin single domain antibodies from a semi-synthetic spiny dogfish shark display library. BMC Biotechnol. 2007, 7, 78. [Google Scholar] [CrossRef] [PubMed]
- Streltsov, V.A.; Varghese, J.N.; Carmichael, J.A.; Irving, R.A.; Hudson, P.J.; Nuttall, S.D. Structural evidence for evolution of shark Ig new antigen receptor variable domain antibodies from a cell-surface receptor. Proc. Natl. Acad. Sci. USA 2004, 101, 12444–12449. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.L.; Anderson, G.P.; Delehanty, J.B.; Baumann, R.; Hayhurst, A.; Goldman, E.R. Selection of cholera toxin specific IgNAR single-domain antibodies from a naive shark library. Mol. Immunol. 2007, 44, 1775–1783. [Google Scholar] [CrossRef] [PubMed]
- Zielonka, S.; Weber, N.; Becker, S.; Doerner, A.; Christmann, A.; Christmann, C.; Uth, C.; Fritz, J.; Schäfer, E.; Steinmann, B.; et al. Shark attack: High affinity binding proteins derived from shark vNAR domains by stepwise in vitro affinity maturation. J. Biotechnol. 2014, 191, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Geoghegan, K.F.; Dixon, H.B.F.; Rosner, P.J.; Hoth, L.R.; Lanzetti, A.J.; Borzilleri, K.A.; Marr, E.S.; Pezzullo, L.H.; Martin, L.B.; LeMotte, P.K.; et al. Spontaneous a-N-6-Phosphogluconoylation of a “His Tag” in Escherichia coli: The cause of extra mass of 258 or 178 Da in fusion proteins. Anal. Biochem. 1999, 267, 169–184. [Google Scholar] [CrossRef] [PubMed]
- Waltenberger, J.; Mayr, U.; Frank, H.; Hombach, V. Suramin is a potent inhibitor of vascular endothelial growth factor: A contribution to the molecular basis of its antiangiogenic action. J. Mol. Cell. Cardiol. 1996, 28, 1523–1529. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, N.; Martin, J.; Ruan, Q.; Rafique, A.; Rosconia, M.P.; Shi, E.; Pyles, E.A.; Yancopoulos, G.D.; Stahl, N.; Wiegard, S.J. Binding and neutralization of vascular endothelial growth factor (VEGF) and related ligands by VEGF Trap, ranibizumab and bevacizumab. Angiogenesis 2012, 15, 171–185. [Google Scholar] [CrossRef] [PubMed]
- Dastjerdi, M.H.; Sadrai, Z.; Saban, D.R.; Zhang, Q.; Dana, R. Corneal penetration of topical and subconjunctival bevacizumab. Investig. Ophthalmol. Vis. Sci. 2011, 52, 8718–8723. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.X.; Ma, J.X. Ocular neovascularization: Implication of endogenous angiogenesis inhibitor and potential therapy. Prog. Retin. Eye Res. 2007, 26, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Falavarjani, K.G.; Nguyen, Q.D. Adverse events and complications associated with intravitreal injection of anti-VEGF agents: A review of literature. Eye 2013, 27, 787–789. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N.; Adamis, A.P. Ten years of anti-vascular endothelial growth factor therapy. Nat. Rev. Drug Discov. 2016, 15, 385–403. [Google Scholar] [CrossRef] [PubMed]
- Agrahari, V.; Agrahari, V.; Mandal, A.; Pal, D.; Mitra, A.K. How are we improving the delivery to back of the eye? advances and challenges of novel therapeutic approaches. Expert Opin. Drug Deliv. 2016, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Pecen, P.E.; Kaiser, P.K. Current phase 1/2 research for neovascular age-related macular degeneration. Curr. Opin. Ophthalmol. 2015, 26, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Fleetwood, F.; Güler, R.; Gordon, E.; Ståhl, S.; Claesson-Welsh, L.; Löfblom, J. Novel affinity binders for neutralization of vascular endothelial growth factor (VEGF) signaling. Cell. Mol. Life Sci. 2015, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, W.; Cheng, S.-F.; Dastjerdi, M.H.; Ferrari, G.; Reza, D. Corneal neovascularization and the utility of topical VEGF inhibition: Ranibizumab (Lucentis) vs. bevacizumab (Avastin). Ocul. Surf. 2012, 10, 67–83. [Google Scholar] [CrossRef] [PubMed]
- Goel, M.; Picciani, R.G.; Lee, R.K.; Bhattacharya, S.K. Aqueous humor dynamics: A review. Open Ophthalmol. J. 2010, 4, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.J.; Ebmeier, S.E.; Sutherland, W.M.; Ghazi, N.G. Potential penetration of topical ranibizumab (Lucentis) in the rabbit eye. Eye 2011, 25, 1504–1511. [Google Scholar] [CrossRef] [PubMed]
- Kovaleva, M.; Johnson, K.; Steven, J.; Barelle, C.J.; Porter, A. Therapeutic potential of shark anti-ICOSL vNAR domains is exemplified in a murine model of autoimmune non-infectious uveitis. Front. Immunol. 2017, 8, 1121. [Google Scholar] [CrossRef] [PubMed]
- Camacho-Villegas, T.A.; Mata-Gonzalez, M.T.; Paniagua-Solis, J.; Sanchez, E.; Licea, A. Human TNF cytokine neutralization with a vNAR from Heterodontus francisci shark: A potential therapeutic use. mAbs 2013, 5, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Barbas, C.F.; Burton, D.R.; Scott, J.K. Phage Display: A Laboratory Manual; Cold Spring Harbor Laboratory Press, Cold Spring Harbor: New York, NY, USA, 2001; ISBN 0-87-969546-3. [Google Scholar]
- Stahl, A.; Connor, K.M.; Sapieha, P.; Chen, J.; Dennison, R.J.; Krah, N.M.; Seaward, M.R.; Willett, K.L.; Aderman, C.M.; Guerin, K.I.; et al. The mouse retina as an angiogenesis model. Investig. Ophthalmol. Vis. Sci. 2010, 51, 2813–2826. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.E.H.; Wesoloiuski, E.; McLellan, A.; Kostyk, S.K.; D’Amato, X.R.; Sullivan, R.; D’Amore, P.A. Oxygen-induced retinopathy in the mouse. Investig. Ophthalmol. Vis. Sci. 1994, 35, 101–111. [Google Scholar]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camacho-Villegas, T.A.; Mata-González, M.T.; García-Ubbelohd, W.; Núñez-García, L.; Elosua, C.; Paniagua-Solis, J.F.; Licea-Navarro, A.F. Intraocular Penetration of a vNAR: In Vivo and In Vitro VEGF165 Neutralization. Mar. Drugs 2018, 16, 113. https://doi.org/10.3390/md16040113
Camacho-Villegas TA, Mata-González MT, García-Ubbelohd W, Núñez-García L, Elosua C, Paniagua-Solis JF, Licea-Navarro AF. Intraocular Penetration of a vNAR: In Vivo and In Vitro VEGF165 Neutralization. Marine Drugs. 2018; 16(4):113. https://doi.org/10.3390/md16040113
Chicago/Turabian StyleCamacho-Villegas, Tanya A., María Teresa Mata-González, Walter García-Ubbelohd, Linda Núñez-García, Carolina Elosua, Jorge F. Paniagua-Solis, and Alexei F. Licea-Navarro. 2018. "Intraocular Penetration of a vNAR: In Vivo and In Vitro VEGF165 Neutralization" Marine Drugs 16, no. 4: 113. https://doi.org/10.3390/md16040113