UPLC–MS/MS Identification of Sterol Sulfates in Marine Diatoms

 , ,
, ,  ,
,
Abstract
:1. Introduction
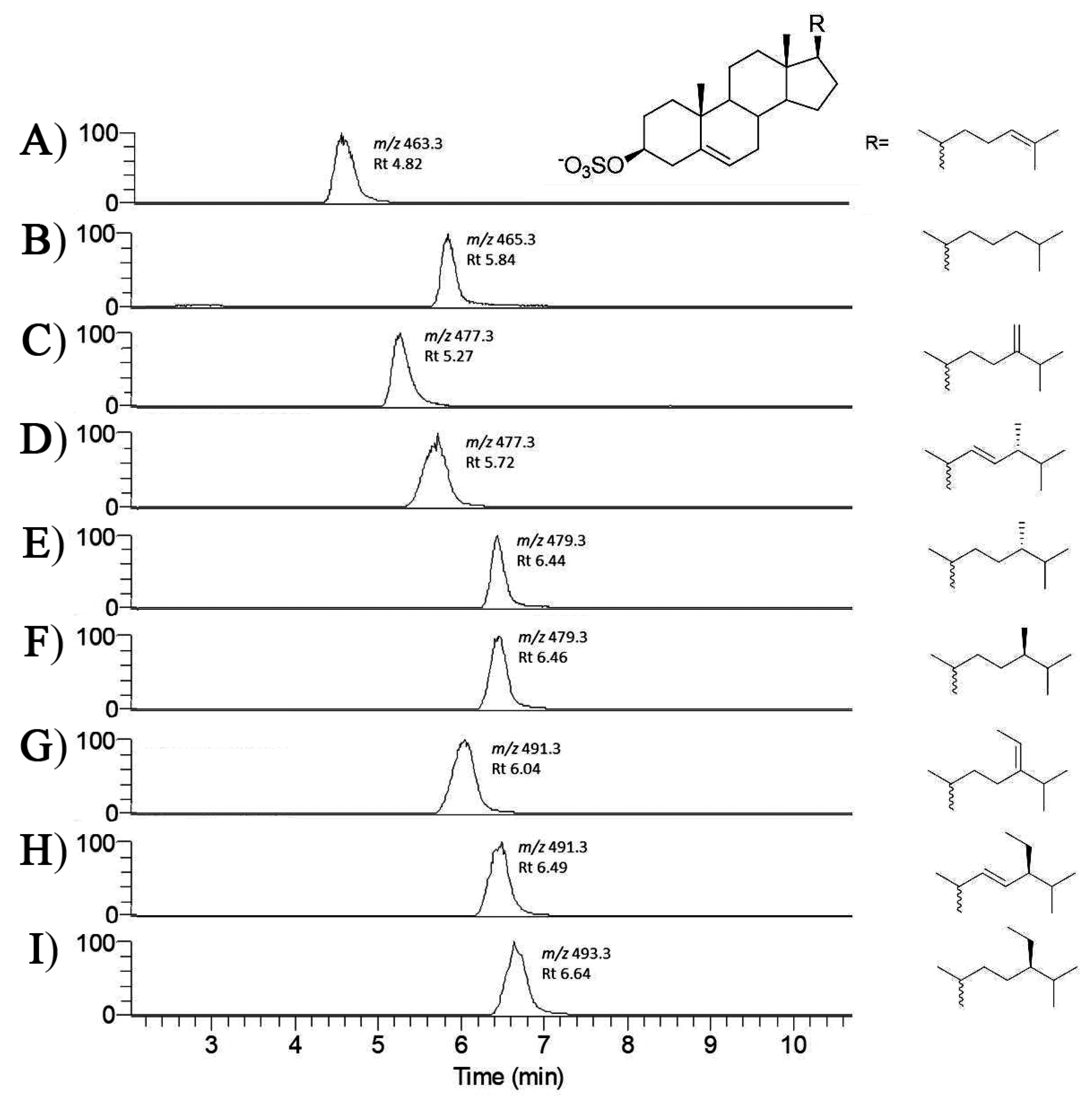
2. Results
3. Discussion
4. Materials and Methods
4.1. General
4.2. Chemical Derivatization of Sterols to Sterol Sulfates
4.3. Microalgal Material
4.4. Sample Preparation for LC/MS Analysis
4.5. Ultra Performance Liquid Chromatography/High Resolution Mass Spectrometry (UPLC/HRMS)
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nelson, D.M.; Treguer, P.; Brzezinski, M.A.; Leynaert, A.; Queguiner, B. Production and dissolution of biogenic silica in the ocean—Revised global estimates, comparison with regional data and relationship to biogenic sedimentation. Glob. Biogeochem. Cycles 1995, 9, 359–372. [Google Scholar] [CrossRef]
- Bidle, K.D.; Azam, F. Accelerated dissolution of diatom silica by marine bacterial assemblages. Nature 1999, 397, 508–512. [Google Scholar] [CrossRef]
- Benoiston, A.S.; Ibarbalz, F.M.; Bittner, L.; Guidi, L.; Jahn, O.; Dutkiewicz, S.; Bowler, C. The evolution of diatoms and their biogeochemical functions. Phil. Trans. R. Soc. B 2017, 372, 20160397. [Google Scholar] [CrossRef] [PubMed]
- Bozarth, A.; Maier, U.G.; Zauner, S. Diatoms in biotechnology: Modern tools and applications. Appl. Microbiol. Biotechnol. 2009, 82, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Vardi, A. Cell signaling in marine diatoms. Commun. Integr. Biol. 2008, 2, 134–136. [Google Scholar] [CrossRef]
- Miralto, A.; Barone, G.; Romano, G.; Poulet, S.A.; Ianora, A.; Russo, G.L.; Buttino, I.; Mazzarella, G.; Laabir, M.; Cabrini, M.; et al. The insidious effect of diatoms on copepod reproduction. Nature 1999, 402, 173–176. [Google Scholar] [CrossRef]
- Ribalet, F.; Wichard, T.; Pohnert, G.; Ianora, A.; Miralto, A.; Casotti, R. Age and nutrient limitation enhance polyunsaturated aldehyde production in marine diatoms. Phytochemistry 2007, 68, 2059–2067. [Google Scholar] [CrossRef]
- Barreiro, A.; Carotenuto, Y.; Lamari, N.; Esposito, F.; d’Ippolito, G.; Fontana, A.; Romano, G.; Ianora, A.; Miralto, A.; Guisande, C. Diatom induction of reproductive failure in copepods: The effect of PUAs versus non volatile oxylipins. J. Exp. Mar. Biol. Ecol. 2011, 401, 13–19. [Google Scholar] [CrossRef]
- Ianora, A.; Bastianini, M.; Carotenuto, Y.; Casotti, R.; Roncalli, V.; Miralto, A.; Romano, G.; Gerecht, A.; Fontana, A.; Turner, J.T. Non-volatile oxylipins render some diatom blooms more toxic for copepod reproduction. Harmful Alga 2015, 44, 1–7. [Google Scholar] [CrossRef]
- Fontana, A.; D’Ippolito, G.; Cutignano, A.; Romano, G.; Lamari, N.; Gallucci, A.M.; Ianora, A. LOX-induced lipid peroxidation mechanism responsible for the detrimental effect of marine diatoms on zooplankton grazers. Chembiochem 2007, 8, 1810–1818. [Google Scholar] [CrossRef]
- Fontana, A.; d’Ippolito, G.; Cutignano, A.; Miralto, A.; Ianora, A.; Romano, G.; Cimino, G. Chemistry of oxylipin pathways in marine diatoms. Pure Appl. Chem. 2007, 79, 481–490. [Google Scholar] [CrossRef]
- d’Ippolito, G.; Nuzzo, G.; Sardo, A.; Manzo, E.; Gallo, C.; Fontana, A. Lipoxygenases and lipoxygenase products in marine diatoms. Methods Enzymol. 2018, 605, 69–100. [Google Scholar] [PubMed]
- Cimino, G.; Ciavatta, M.L.; Fontana, A.; Gavagnin, M. Metabolites of marine opisthobranchs: Chemistry and biological activity. In Bioactive Compounds from Natural Sources; Tringali, C., Ed.; Taylor & Francis: London, UK, 2001; pp. 579–637. [Google Scholar]
- Marthi, R.; Fontana, A.; Uriz, M.J.; Cimino, G. Quantitative assessment of natural toxicity in sponge: Toxicity bioassay versus compound quantification. J. Chem. Ecol. 2003, 29, 1307–1318. [Google Scholar] [CrossRef]
- Mollo, E.; Fontana, A.; Roussis, V.; Polese, G.; Amodeo, P.; Ghiselin, M.T. Sensing marine biomolecules: Smell, taste, and the evolutionary transition from aquatic to terrestrial life. Front. Chem. 2014, 2, 92. [Google Scholar] [CrossRef] [PubMed]
- Fontana, A.; Ciavatta, M.L.; D’Souza, L.; Mollo, E.; Naik, C.G.; Parameswaran, P.S.; Cimino, G. Selected chemo-ecological studies of marine opisthobranchs from Indian coasts. J. Indian Inst. Sci. 2001, 81, 403–415. [Google Scholar]
- Gallo, C.; d’Ippolito, G.; Nuzzo, G.; Sardo, A.; Fontana, A. Autoinhibitory sterol sulfates mediate programmed cell death in a bloom-forming marine diatom. Nat. Commun. 2018, 8, 1292–1303. [Google Scholar] [CrossRef] [PubMed]
- Gallo, C.; Nuzzo, G.; d’Ippolito, G.; Manzo, E.; Sardo, A.; Fontana, A. Sterol sulfates and sulfotransferases in marine diatoms. Methods Enzymol. 2018, 605, 101–138. [Google Scholar]
- Rampen, S.W.; Abbas, B.A.; Schouten, S.; Sinninghe Dams, J.S. A comprehensive study of sterols in marine diatoms (Bacillariophyta): Implications for their use as tracers for diatom productivity. Limnol. Oceanogr. 2010, 55, 91–105. [Google Scholar] [CrossRef]
- McKee, T.C.; Cardellina, J.H.; Riccio, R.; D’Auria, M.V.; Iorizzi, M.; Minale, L.; Buckheit, R.W. HIV-inhibitory natural products. 11. Comparative studies of sulfated sterols from marine invertebrates. J. Med. Chem. 1994, 37, 793–797. [Google Scholar] [CrossRef]
- Patil, A.D.; Freyer, A.J.; Breen, A.; Carte, B.; Johnson, R.K. Halistanol disulfate B, a novel sulfated sterol from the sponge Pachastrella sp.: Inhibitor of endothelin converting enzyme. J. Nat. Prod. 1996, 59, 606–608. [Google Scholar] [CrossRef]
- Rudi, A.; Yosief, T.; Loya, S.; Hizi, A.; Schleyer, M.; Kashman, Y. Clathsterol, a novel anti-HIV-1 RT sulfated sterol from the sponge clathria species. J. Nat. Prod. 2001, 64, 1451–1453. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Shen, X.R.; Kong, J.L. Arenicolsterol A, a novel cytotoxic enolic sulfated sterol from a marine annelid Arenicola cristata. Chin. J. Chem. 2005, 23, 599–602. [Google Scholar] [CrossRef]
- Yang, S.W.; Buivich, A.; Chan, T.M.; Smith, M.; Lachowicz, J.; Pomponi, S.A.; Chu, M. A new sterol sulfate, Sch 572423, from a marine sponge, Topsentia sp. Bioorg. Med. Chem. Lett. 2003, 13, 1791–1794. [Google Scholar] [CrossRef]
- DiGirolamo, J.A.; Li, X.-C.; Jacob, M.R.; Clark, A.M.; Ferreira, D. Reversal of fluconazole resistance by sulfated sterols from the marine sponge Topsentia sp. J. Nat. Prod. 2009, 72, 1524–1528. [Google Scholar] [CrossRef] [PubMed]
- Morinaka, B.I.; Pawlik, J.R.; Molinski, T.F. Amaroxocanes A and B: Sulfated dimeric sterols defend the caribbean coral reef sponge Phorbas amaranthus from fish predators. J. Nat. Prod. 2009, 72, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Festa, C.; De Marino, S.; Dauria, M.V.; Bifulco, G.; Renga, B.; Fiorucci, S.; Zampella, A. Solomonsterols A and B from Theonella swinhoei. the first example of C-24 and C-23 sulfated sterols from a marine source endowed with a PXR agonistic activity. J. Med. Chem. 2011, 54, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Sepe, V.; Bifulco, G.; Renga, B.; D’Amore, C.; Fiorucci, S.; Zampella, A. Discovery of sulfated sterols from marine invertebrates as a new class of marine natural antagonists of farnesoid-x-receptor. J. Med. Chem. 2011, 54, 1314–1320. [Google Scholar] [CrossRef]
- Casanovas, A.; Hannibal-Bach, H.K.; Jensen, O.N.; Ejsing, C.S. Shotgun lipidomic analysis of chemically sulfated sterols compromises analytical sensitivity: Recommendation for large-scale global lipidome analysis. Eur. J. Lipid Sci. Technol. 2014, 116, 1618–1620. [Google Scholar] [CrossRef] [Green Version]
- Gouveia, M.J.; Brindley, P.-J.; Santos, L.L.; Correia da Costa, J.M.; Gomes, P.; Vale, N. Mass spectrometry techniques in the survey of steroid metabolites as potential disease biomarkers: A review. Metabolism 2013, 62, 1206–1217. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Guijo, A.; Oji, V.; Hartmann, M.F.; Schuppe, H.C.; Traupe, H.; Wudy, S.A. High levels of oxysterol sulfates in serum of patients with steroid sulfatase deficiency. J. Lipid Res. 2015, 56, 403–412. [Google Scholar] [CrossRef]
- Sánchez-Guijo, A.; Oji, V.; Hartmann, M.F.; Traupe, H.; Wudy, S.A. Simultaneous quantification of cholesterol sulfate, androgen sulfates, and progestagen sulfates in human serum by LC–MS/MS. J. Lipid Res. 2015, 56, 1843–1851. [Google Scholar] [CrossRef]
- Strott, C.A.; Higashi, Y. Cholesterol sulfate in human physiology: What’s it all about? J. Lipid Res. 2003, 44, 1268–1278. [Google Scholar] [CrossRef] [PubMed]
- Guillard, R.R.L.; Ryther, J.H. Studies of marine planktonic diatoms. I. Cyclotella nana Hustedt and Detonula confervacea Cleve. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Raniello, R.; Iannicelli, M.M.; Nappo, M.; Avila, C.; Zupo, V. Production of Cocconeis neothumensis (Bacillariophyceae) biomass in batch cultures and bioreactors for biotechnological applications: Light and nutrient requirements. J. Appl. Phycol. 2007, 19, 383–391. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
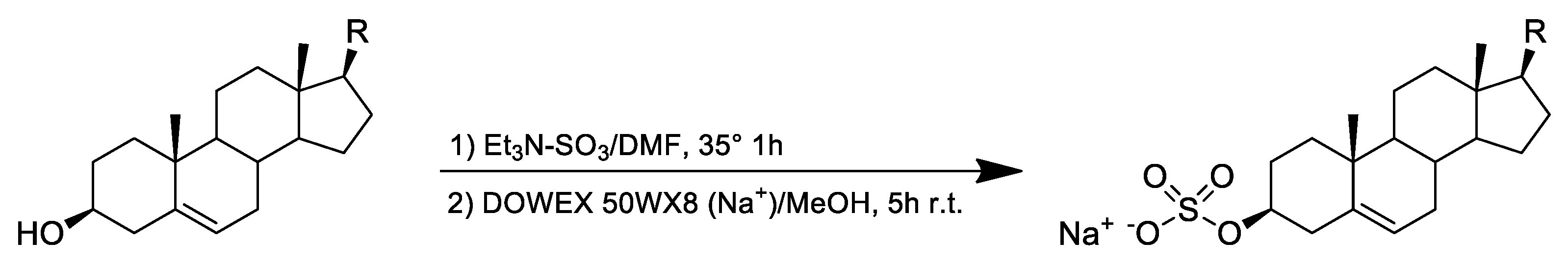{kind=link}
| Compounds | Molecular Formula | Calculated MS | Measured m/z | Species Distribution |
|---|---|---|---|---|
| Desmosterol sulfate | C27H43O4S− | 463.2887 | 463.2896 | C. closterium P. tricornutum |
| Cholesterol sulfate | C27H45O4S− | 465.3044 | 465.3047 | S. marinoi |
| 24-methylene cholesterol sulfate | C28H45O4S− | 477.3044 | 477.3043 | S. marinoi C. cryptica C. scutellum P. tricornutum T. pseudonana T. rotula |
| Brassicasterol sulfate (24β-methyl-22-dehydrocholesterol) | C28H45O4S− | 477.3044 | 477.3052 | N. shiloi C. scutellum Diploneis sp. |
| 24-methyl cholesterol sulfate | C28H47O4S− | 479.3200 | 479.3205 | S. marinoi 1 C. Cryptica 1 T. pseudonana T. rotula |
| Fucosterol sulfate | C29H47O4S− | 491.3200 | 491.3201 | S. marinoi N. shiloi |
| β-Sitosterol sulfate (24β-ethyl cholesterol) | C29H49O4S− | 493.3357 | 493.3361 | S. marinoi |
| Diatom Species | Compounds | µg StS/mg C |
|---|---|---|
| C. closterium | Desmosterol sulfate | 26.21 ± 2.6 |
| Total StS | 26.21 ± 2.6 | |
| N. shiloi | Brassicasterol sulfate | 5.94 ± 1.0 |
| Fucosterol sulfate | 60.69 ± 7.3 | |
| Total StS | 66.63 ± 5.2 | |
| C. scutellum | 24-methylene cholesterol sulfate | 10.09 ± 1.3 |
| Brassicasterol sulfate | 3.91 ± 0.1 | |
| Total StS | 14.00 ± 1.45 | |
| Diploneis sp. | Brassicasterol sulfate | 11.03 ± 2.9 |
| Total StS | 11.03 ± 2.9 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nuzzo, G.; Gallo, C.; D’Ippolito, G.; Manzo, E.; Ruocco, N.; Russo, E.; Carotenuto, Y.; Costantini, M.; Zupo, V.; Sardo, A.; et al. UPLC–MS/MS Identification of Sterol Sulfates in Marine Diatoms. Mar. Drugs 2019, 17, 10. https://doi.org/10.3390/md17010010
Nuzzo G, Gallo C, D’Ippolito G, Manzo E, Ruocco N, Russo E, Carotenuto Y, Costantini M, Zupo V, Sardo A, et al. UPLC–MS/MS Identification of Sterol Sulfates in Marine Diatoms. Marine Drugs. 2019; 17(1):10. https://doi.org/10.3390/md17010010
Chicago/Turabian StyleNuzzo, Genoveffa, Carmela Gallo, Giuliana D’Ippolito, Emiliano Manzo, Nadia Ruocco, Ennio Russo, Ylenia Carotenuto, Maria Costantini, Valerio Zupo, Angela Sardo, and et al. 2019. "UPLC–MS/MS Identification of Sterol Sulfates in Marine Diatoms" Marine Drugs 17, no. 1: 10. https://doi.org/10.3390/md17010010