Insights into Red Sea Brine Pool Specialized Metabolism Gene Clusters Encoding Potential Metabolites for Biotechnological Applications and Extremophile Survival


Abstract
:1. Introduction
2. Results
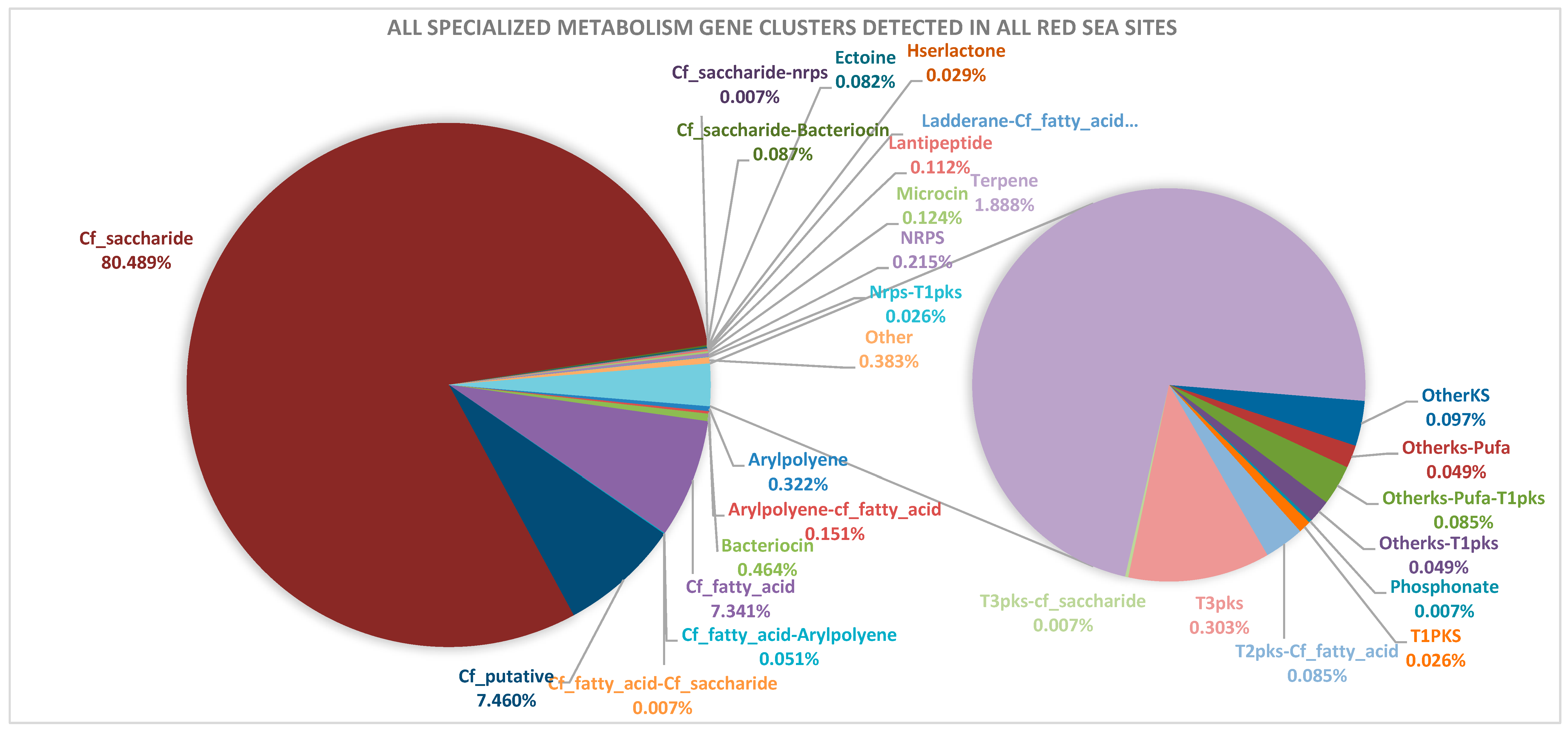
2.1. Abundance and Diversity of Specialized Metabolism Gene Clusters (SMGCs) in Red Sea Brine Pools
2.2. Red Sea Brine Pool SMGCs Code for Diverse Potential Functions
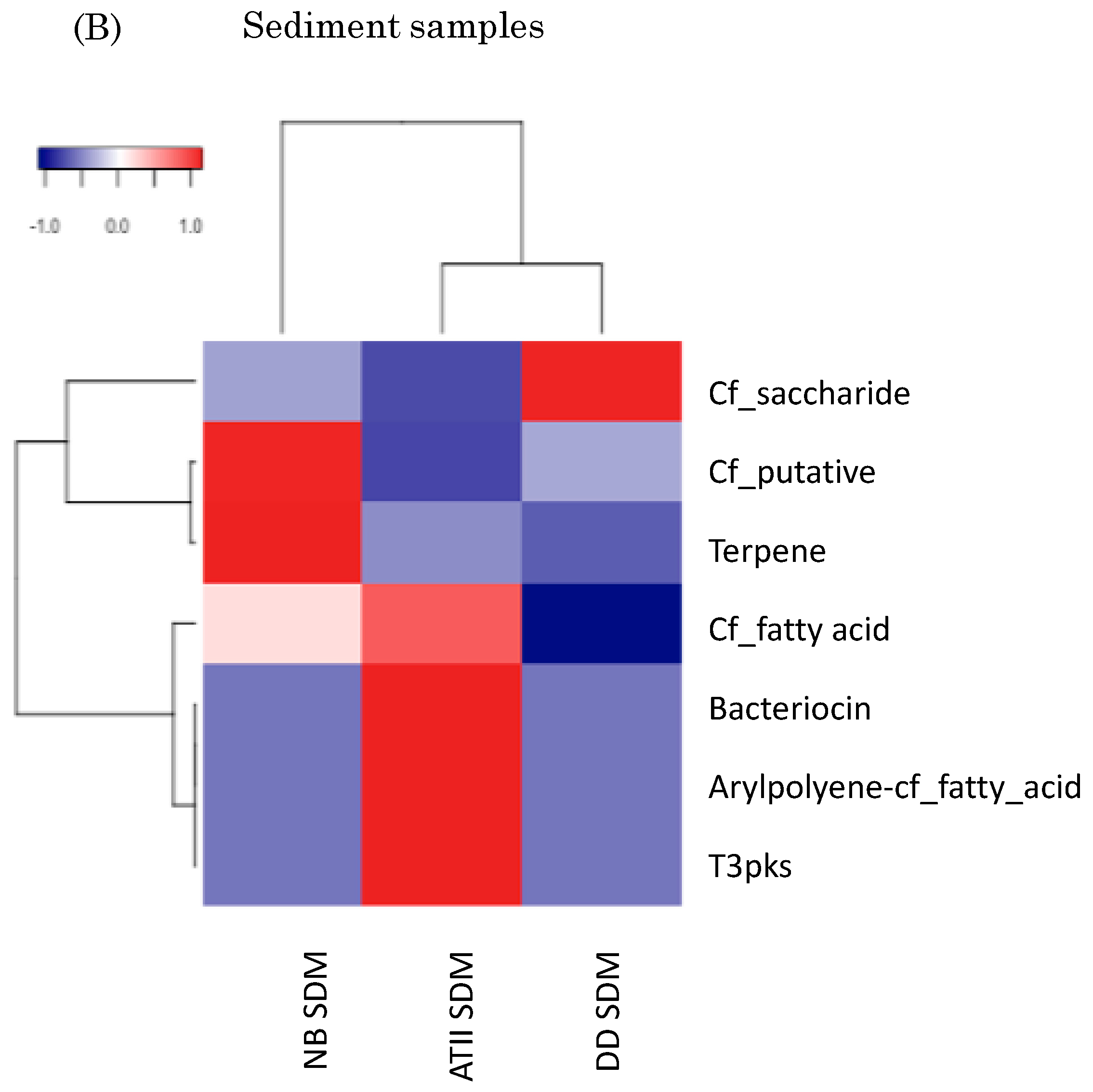
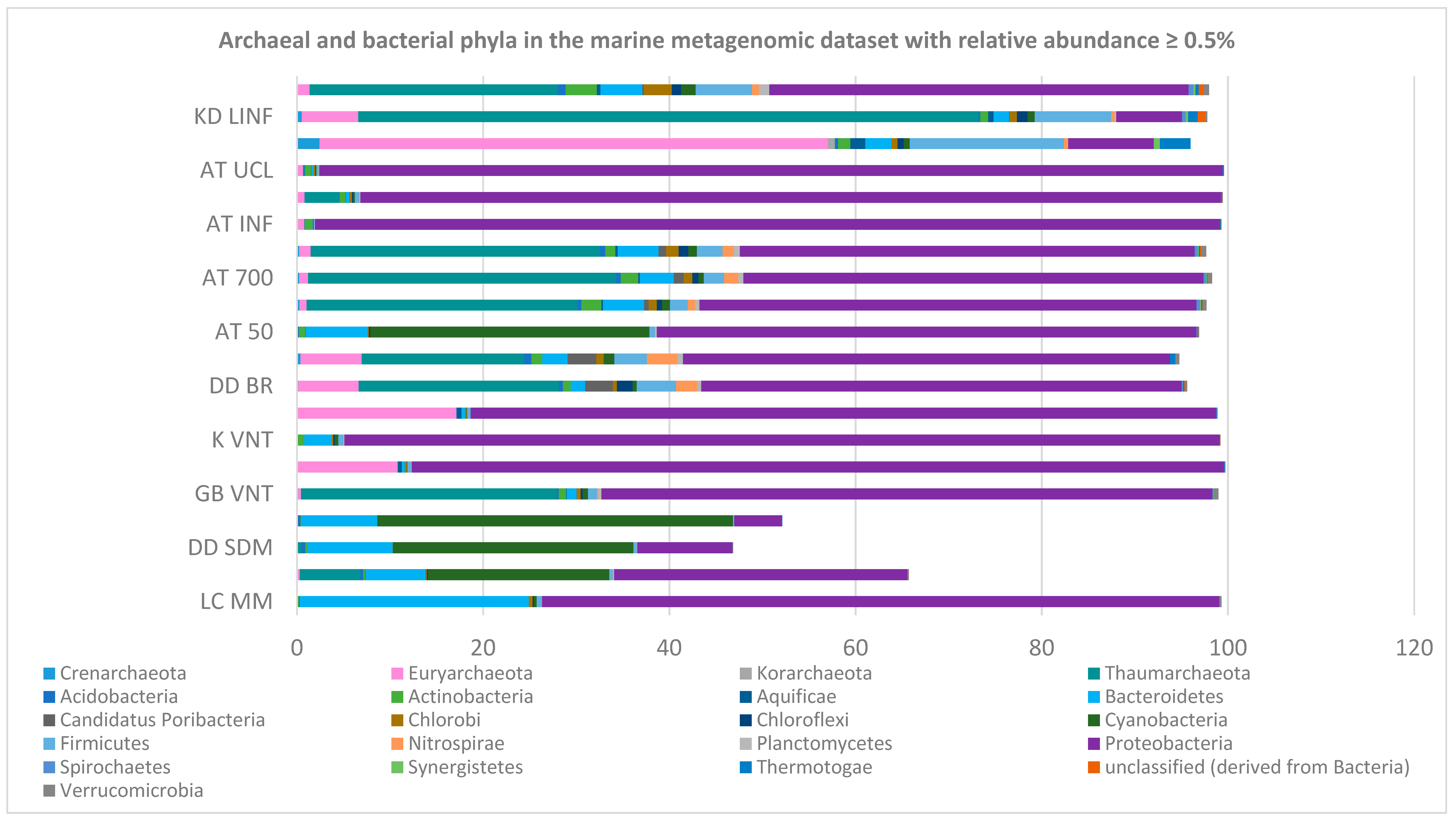
2.3. Red Sea Brine Prokaryotic Diversity in Relation to Specialized Metabolism Genes
2.4. Rare Leucine Codons within Red Sea Brine SMGCs and Low Similarity to Known Clusters with Characterized Products
3. Discussion
3.1. Saccharide and Putative SMGCs Are the Most Abundant Groups in the Red Sea Brine Dataset
3.2. Preliminary Evidence of Potential Products with Pharmaceutical Applications
3.3. Red Sea Brine SMGCs form a Unique Cluster
3.4. Environment–Microbe Interaction, Antagonistic Stressors and Extremophile Survival Implicated by Red Sea Brine SMGCs
3.5. Prolific Specialized-Metabolite-Producing Phyla Detection and Red Sea Brine Pool SMGC Dark Matter Analysis
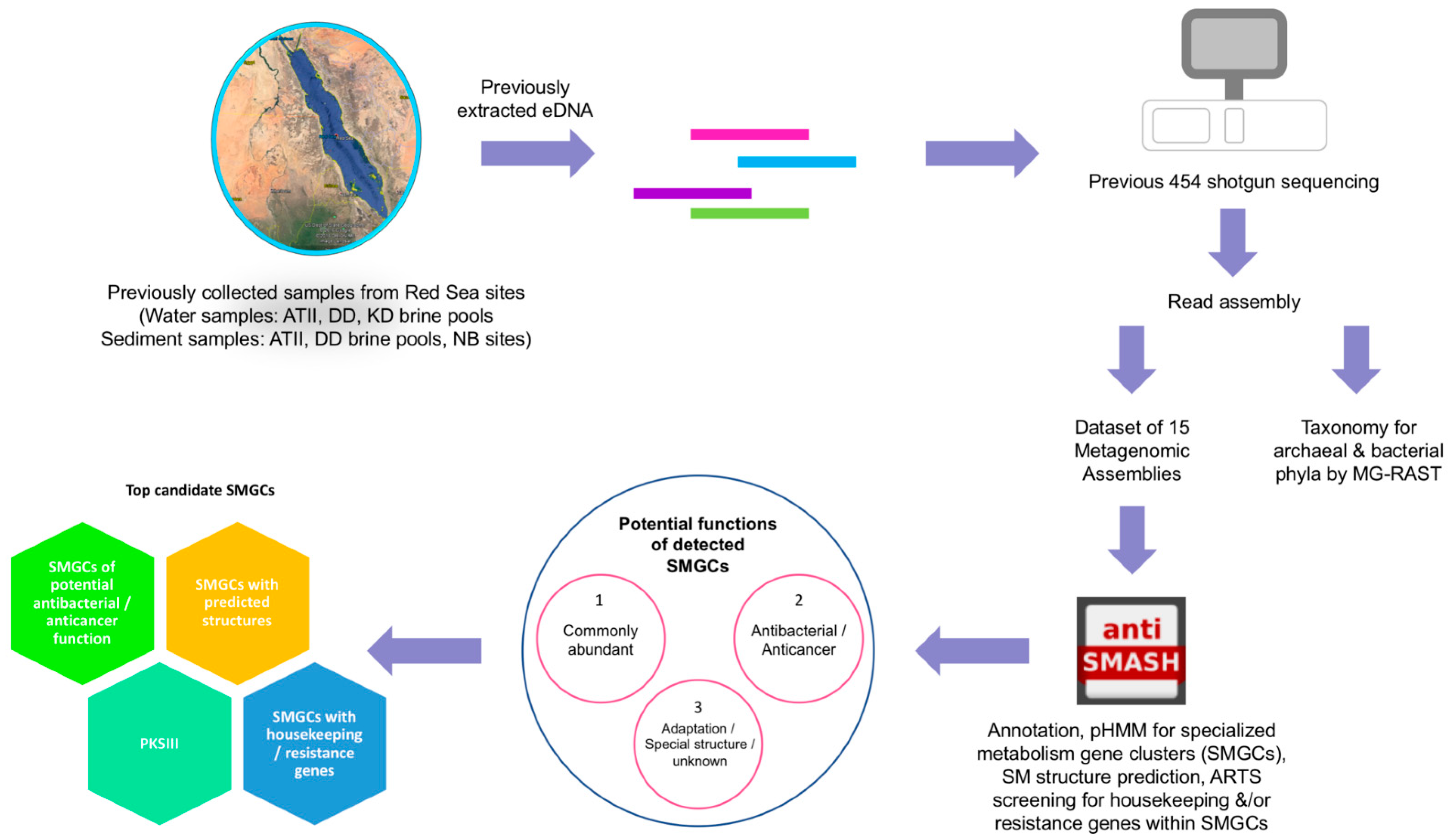
4. Materials and Methods
4.1. Sampling, DNA Extraction and Sequencing
4.2. Bioinformatics Assembly
4.3. Annotation, SMGCs Analyses and Hierarchical Classification
4.4. Taxonomic Classification
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability
References
- Williams, D.N. Antimicrobial resistance: Are we at the dawn of the post-antibiotic era? J. R. Coll. Physicians Edinb. 2016, 46, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, M.; Moore, R.T.; Rajamani, S.; Panchal, R.G. Bacterial genome engineering and synthetic biology: combating pathogens. BMC Microbiol. 2016, 16, 258. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.D.; Wright, G.D. Antibacterial drug discovery in the resistance era. Nature 2016, 529, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Martens, E.; Demain, A.L. The antibiotic resistance crisis, with a focus on the United States. J. Antibiot. (Tokyo) 2017, 70, 520–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holohan, C.; Van Schaeybroeck, S.; Longley, D.B.; Johnston, P.G. Cancer drug resistance: an evolving paradigm. Nat. Rev. Cancer 2013, 13, 714–726. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef]
- Demain, A.; Fang, A. The natural functions of secondary metabolites. Adv. Biochem. Eng. Biotechnol. 2000, 69, 222. [Google Scholar]
- Milshteyn, A.; Schneider, J.S.S.; Brady, S.F.F. Mining the metabiome: Identifying novel natural products from microbial communities. Chem. Biol. 2014, 21, 1211–1223. [Google Scholar] [CrossRef]
- Weber, T.; Blin, K.; Duddela, S.; Krug, D.; Kim, H.U.; Bruccoleri, R.; Lee, S.Y.; Fischbach, M.A.; Muller, R.; Wohlleben, W.; et al. antiSMASH 3.0—A comprehensive resource for the genome mining of biosynthetic gene clusters. Nucleic Acids Res. 2015, 1–7. [Google Scholar] [CrossRef]
- Ziemert, N.; Alanjary, M.; Weber, T. The evolution of genome mining in microbes—A review. Nat. Prod. Rep. 2016, 33, 988–1005. [Google Scholar] [CrossRef] [PubMed]
- Udwary, D.W.; Zeigler, L.; Asolkar, R.N.; Singan, V.; Lapidus, A.; Fenical, W.; Jensen, P.R.; Moore, B.S.; Walsh, C.T. Genome sequencing reveals complex secondary metabolome in the marine actinomycete Salinispora tropica. PNAS 2007, 104, 10376–10381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donia, M.S.S.; Cimermancic, P.; Schulze, C.J.J.; Wieland Brown, L.C.; Martin, J.; Mitreva, M.; Clardy, J.; Linington, R.G.G.; Fischbach, M.A.A.; Wieland Brown, L.C.; et al. A systematic analysis of biosynthetic gene clusters in the human microbiome reveals a common family of antibiotics. Cell 2014, 158, 1402–1414. [Google Scholar] [CrossRef] [Green Version]
- Schofield, M.M.; Sherman, D.H. Meta-omic characterization of prokaryotic gene clusters for natural product biosynthesis. Curr. Opin. Biotechnol. 2013, 24, 1151–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boddy, C.N. Bioinformatics tools for genome mining of polyketide and non-ribosomal peptides. J. Ind. Microbiol. Biotechnol. 2014, 41, 443–450. [Google Scholar] [CrossRef]
- Trindade, M.; van Zyl, L.J.; Navarro-Fernández, J.; Abd Elrazak, A. Targeted metagenomics as a tool to tap into marine natural product diversity for the discovery and production of drug candidates. Front. Microbiol. 2015, 6, 1–14. [Google Scholar] [CrossRef]
- Ul Hassan, S.S.; Shaikh, A.L. Marine actinobacteria as a drug treasure house. Biomed. Pharmacother. 2017, 87, 46–57. [Google Scholar] [CrossRef]
- Pettit, R.K. Culturability and Secondary Metabolite Diversity of Extreme Microbes: Expanding Contribution of Deep Sea and Deep-Sea Vent Microbes to Natural Product Discovery. Mar. Biotechnol. 2011, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Giddings, L.-A.; Newman, D.J. Bioactive Compounds from Extremophiles; Springer: Cham, Switzerland, 2015; ISBN 3319148354. [Google Scholar]
- Ferreira, A.J.S.; Siam, R.; Setubal, J.C.; Moustafa, A.; Sayed, A.; Chambergo, F.S.; Dawe, A.S.; Ghazy, M.A.; Sharaf, H.; Ouf, A.; et al. Core microbial functional activities in ocean environments revealed by global metagenomic profiling analyses. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Abdallah, R.Z.; Adel, M.; Ouf, A.; Sayed, A.; Ghazy, M.A.; Alam, I.; Essack, M.; Lafi, F.F.; Bajic, V.B.; El-Dorry, H.; et al. Aerobic methanotrophic communities at the Red Sea brine-seawater interface. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Antunes, A.; Ngugi, D.K.; Stingl, U. Microbiology of the Red Sea (and other) deep-sea anoxic brine lakes. Environ. Microbiol. Rep. 2011, 3, 416–433. [Google Scholar] [CrossRef] [PubMed]
- Elbehery, A.H.A.; Aziz, R.K.; Siam, R. Insertion sequences enrichment in extreme Red sea brine pool vent. Extremophiles 2017, 21, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Backer, H.; Schoell, M. New Deeps with Brines and Metalliferous Sediments in the Red Sea. Nat. Phys. Sci. 1972, 240, 153. [Google Scholar] [CrossRef]
- Hartmann, M.; Scholten, J.C.C.; Stoffers, P.; Wehner, F. Hydrographic structure of brine-filled deeps in the Red Sea - New results from the Shaban, Kebrit, Atlantis II, and discovery deep. Mar. Geol. 1998, 144, 311–330. [Google Scholar] [CrossRef]
- Anschutz, P.; Blanc, G.; Monnin, C.; Boulègue, J. Geochemical dynamics of the Atlantis II Deep (Red Sea): II. Composition of metalliferous sediment pore waters. Geochim. Cosmochim. Acta 2000, 64, 3995–4006. [Google Scholar] [CrossRef]
- Faber, E.; Botz, R.; Poggenburg, J.; Schmidt, M.; Stoffers, P.; Hartmann, M. Methane in Red Sea brines. Org. Geochem. 1998, 29, 363–379. [Google Scholar] [CrossRef]
- Eder, W.; Jahnke, L.L.; Schmidt, M.; Huber, R. Microbial Diversity of the Brine-Seawater Interface of the Kebrit Deep, Red Sea, Studied via 16S rRNA Gene Sequences and Cultivation Methods. Appl. Environ. Microbiol. 2001, 67, 3077–3085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siam, R.; Mustafa, G.A.; Sharaf, H.; Moustafa, A.; Ramadan, A.R.; Antunes, A.; Bajic, V.B.; Stingl, U.; Marsis, N.G.R.; Coolen, M.J.L.; et al. Unique prokaryotic consortia in geochemically distinct sediments from red sea Atlantis II and discovery deep brine pools. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Zierenberg, R.A. Deposition of Metalliferous Sediment Beneath a Brine Pool in the Atlantis II Deep, Red Sea. In Gorda Ridge; Springer: New York, NY, USA, 1990; pp. 131–142. [Google Scholar]
- Eustáquio, A.S.; Nam, S.J.; Penn, K.; Lechner, A.; Wilson, M.C.; Fenical, W.; Jensen, P.R.; Moore, B.S. The discovery of salinosporamide K from the marine bacterium “Salinispora pacifica” by Genome mining gives insight into pathway evolution. ChemBioChem 2011, 12, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Othoum, G.; Bougouffa, S.; Razali, R.; Bokhari, A.; Alamoudi, S.; Antunes, A.; Gao, X.; Hoehndorf, R.; Arold, S.T.; Gojobori, T.; et al. In silico exploration of Red Sea Bacillus genomes for natural product biosynthetic gene clusters. BMC Genomics 2018, 19, 382. [Google Scholar] [CrossRef]
- Al-Amoudi, S.; Essack, M.; Simões, M.F.; Bougouffa, S.; Soloviev, I.; Archer, J.A.C.; Lafi, F.F.; Bajic, V.B. Bioprospecting Red Sea Coastal Ecosystems for Culturable Microorganisms and Their Antimicrobial Potential. Mar. Drugs 2016, 14, 165. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, Y.M.; Ghazy, M.A.; Sayed, A.; Ouf, A.; El-Dorry, H.; Siam, R. Isolation and characterization of a heavy metal-resistant, thermophilic esterase from a Red Sea Brine Pool. Sci. Rep. 2013, 3, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sonbol, S.A.; Ferreira, A.J.S.; Siam, R. Red Sea Atlantis II brine pool nitrilase with unique thermostability profile and heavy metal tolerance. BMC Biotechnol. 2016, 16, 14. [Google Scholar] [CrossRef]
- Elbehery, A.H.A.; Leak, D.J.; Siam, R. Novel thermostable antibiotic resistance enzymes from the Atlantis II Deep Red Sea brine pool. Microb. Biotechnol. 2017, 10, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Adel, M.; Elbehery, A.H.A.; Aziz, S.K.; Aziz, R.K.; Grossart, H.P.; Siam, R. Viruses-to-mobile genetic elements skew in the deep Atlantis II brine pool sediments. Sci. Rep. 2016, 6, 32704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, F.; Paarmann, D.; D’Souza, M.; Olson, R.; Glass, E.; Kubal, M.; Paczian, T.; Rodriguez, A.; Stevens, R.; Wilke, A.; et al. The metagenomics RAST server—A public resource for the automatic phylogenetic and functional analysis of metagenomes. BMC Bioinform. 2008, 9, 386. [Google Scholar] [CrossRef] [PubMed]
- Alanjary, M.; Kronmiller, B.; Adamek, M.; Blin, K.; Weber, T.; Huson, D.; Philmus, B.; Ziemert, N. The Antibiotic Resistant Target Seeker (ARTS), an exploration engine for antibiotic cluster prioritization and novel drug target discovery. Nucleic Acids Res. 2017, 45, W42–W48. [Google Scholar] [CrossRef]
- Sheik, C.S.; Jain, S.; Dick, G.J. Metabolic flexibility of enigmatic SAR324 revealed through metagenomics and metatranscriptomics. Environ. Microbiol. 2014, 16, 304–317. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.J.; Lesniewski, R.A.; Dick, G.J. Genome-enabled transcriptomics reveals archaeal populations that drive nitrification in a deep-sea hydrothermal plume. ISME J. 2012, 6, 2269–2279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesniewski, R.A.; Jain, S.; Anantharaman, K.; Schloss, P.D.; Dick, G.J. The metatranscriptome of a deep-sea hydrothermal plume is dominated by water column methanotrophs and lithotrophs. ISME J. 2012, 6, 2257–2268. [Google Scholar] [CrossRef] [Green Version]
- Tang, K.; Liu, K.; Jiao, N.; Zhang, Y.; Chen, C.T.A. Functional Metagenomic Investigations of Microbial Communities in a Shallow-Sea Hydrothermal System. PLoS ONE 2013, 8, e72958. [Google Scholar] [CrossRef]
- Anderson, R.E.; Beltrán, M.T.; Hallam, S.J.; Baross, J.A. Microbial community structure across fluid gradients in the Juan de Fuca Ridge hydrothermal system. FEMS Microbiol. Ecol. 2013, 83, 324–339. [Google Scholar] [CrossRef] [PubMed]
- Stokke, R.; Dahle, H.; Roalkvam, I.; Wissuwa, J.; Daae, F.L.; Tooming-Klunderud, A.; Thorseth, I.H.; Pedersen, R.B.; Steen, I.H. Functional interactions among filamentous Epsilonproteobacteria and Bacteroidetes in a deep-sea hydrothermal vent biofilm. Environ. Microbiol. 2015, 17, 4063–4077. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Wolf, T.; Chevrette, M.G.; Lu, X.; Schwalen, C.J.; Kautsar, S.A.; Suarez Duran, H.G.; de los Santos, E.L.C.; Kim, H.U.; Nave, M.; et al. antiSMASH 4.0—improvements in chemistry prediction and gene cluster boundary identification. Nucleic Acids Res. 2017, 1854, 1019–1037. [Google Scholar] [CrossRef] [PubMed]
- Cimermancic, P.; Medema, M.H.H.; Claesen, J.; Kurita, K.; Wieland Brown, L.C.; Mavrommatis, K.; Pati, A.; Godfrey, P.A.A.; Koehrsen, M.; Clardy, J.; et al. Insights into secondary metabolism from a global analysis of prokaryotic biosynthetic gene clusters. Cell 2014, 158, 412–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weitnauer, G.; Mühlenweg, A.; Trefzer, A.; Hoffmeister, D.; Süßmuth, R.D.; Jung, G.; Welzel, K.; Vente, A.; Girreser, U.; Bechthold, A. Biosynthesis of the orthosomycin antibiotic avilamycin A: Deductions from the molecular analysis of the avi biosynthetic gene cluster of Streptomyces viridochromogenes Tü57 and production of new antibiotics. Chem. Biol. 2001, 8, 569–581. [Google Scholar] [CrossRef]
- Wang, F.; Xiao, X.; Ou, H.-Y.; Gai, Y.; Wang, F. Role and regulation of fatty acid biosynthesis in the response of Shewanella piezotolerans WP3 to different temperatures and pressures. J. Bacteriol. 2009, 191, 2574–2584. [Google Scholar] [CrossRef] [PubMed]
- Schöner, T.A.; Gassel, S.; Osawa, A.; Tobias, N.J.; Okuno, Y.; Sakakibara, Y.; Shindo, K.; Sandmann, G.; Bode, H.B. Aryl Polyenes, a Highly Abundant Class of Bacterial Natural Products, Are Functionally Related to Antioxidative Carotenoids. ChemBioChem 2016, 17, 247–253. [Google Scholar] [CrossRef]
- Churchill, M.E.A.; Sibhatu, H.M.; Uhlson, C.L.; Charis, L.U.; Uhlson, C.L. Defining the structure and function of acyl-homoserine lactone autoinducers. Methods Mol. Biol. 2011, 692, 159–171. [Google Scholar]
- Brahmkshatriya, P.; Brahmkshatriya, P. Terpenes: Chemistry, Biological Role, and Therapeutic Applications. In Natural Products; Ramawat, K., Mérillon, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 2665–2691. [Google Scholar]
- Kaur, S.; Kaur, S. Bacteriocins as potential anticancer agents. Front. Pharmacol. 2015, 6, 1–11. [Google Scholar] [CrossRef]
- Felnagle, E.A.; Jackson, E.E.; Chan, Y.A.; Podevels, A.M.; Berti, A.D.; McMahon, M.D.; Thomas, M.G.; Berti, D.; McMahon, M.D.; Thomas, M.G. Nonribosomal Peptide Synthetases Involved in the Production of Medically Relevant Natural Products. Mol. Pharm. 2008, 5, 191–211. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Xu, F.; Zeng, J.; Zhan, J. Type III polyketide synthases in natural product biosynthesis. IUBMB Life 2012, 64, 285–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, Y.P.; Go, M.K.; Yew, W.S. Exploiting the biosynthetic potential of type III polyketide synthases. Molecules 2016, 21, 806. [Google Scholar] [CrossRef]
- Metcalf, W.W.; van der Donk, W.A. Biosynthesis of phosphonic and phosphinic acid natural products. Annu. Rev. Biochem. 2009, 78, 65–94. [Google Scholar] [CrossRef]
- Abd Elrazak, A.; Ward, A.C.; Glassey, J. Polyunsaturated fatty acid production by marine bacteria. Bioprocess Biosyst. Eng. 2013, 36, 1641–1652. [Google Scholar] [CrossRef] [PubMed]
- Capece, M.C.; Clark, E.; Saleh, J.K.; Halford, D.; Heinl, N.; Hoskins, S.; Rothschild, L.J. Polyextremophiles and the constraints for terrestrial habitability. In Polyextremophiles: Life Under Multiple Forms of Stress; Seckbach, J., Oren, A., Stan-Lotter, H., Eds.; Springer: Dordrecht, The Netherlands, 2013; Volume 27, pp. 3–59. ISBN 978-94-007-6487-3. [Google Scholar]
- Jorge, C.D.; Borges, N.; Bagyan, I.; Bilstein, A.; Santos, H. Potential applications of stress solutes from extremophiles in protein folding diseases and healthcare. Extremophiles 2016, 20, 251–259. [Google Scholar] [CrossRef]
- Javidpour, P.; Deutsch, S.; Mutalik, V.K.; Hillson, N.J.; Petzold, C.J.; Keasling, J.D.; Beller, H.R. Investigation of proposed ladderane biosynthetic genes from anammox bacteria by heterologous expression in E. coli. PLoS ONE 2016, 11, e0151087. [Google Scholar] [CrossRef] [PubMed]
- Medema, M.H.; Fischbach, M.A. Computational approaches to natural product discovery. Nat. Chem. Biol. 2015, 11, 639–648. [Google Scholar] [CrossRef] [Green Version]
- Crits-Christoph, A.; Diamond, S.; Butterfield, C.N.; Thomas, B.C.; Banfield, J.F. Novel soil bacteria possess diverse genes for secondary metabolite biosynthesis. Nature 2018, 558, 440–444. [Google Scholar] [CrossRef]
- Naughton, L.M.; Romano, S.; O’Gara, F.; Dobson, A.D.W. Identification of secondary metabolite gene clusters in the Pseudovibrio Genus reveals encouraging biosynthetic potential toward the production of novel bioactive compounds. Front. Microbiol. 2017, 8, 1–15. [Google Scholar] [CrossRef]
- Blin, K.; Kim, H.U.; Medema, M.H.; Weber, T. Recent development of antiSMASH and other computational approaches to mine secondary metabolite biosynthetic gene clusters. Brief Bioinform. 2017, 1–11. [Google Scholar] [CrossRef]
- Ziko, L.; Saqr, A.H.A.; Ouf, A.; Gimpel, M.; Aziz, R.K.; Neubauer, P. Antibacterial and anticancer activities of orphan biosynthetic gene clusters from Atlantis II Red Sea brine pool. Microb. Cell Fact. 2019, 18, 1–16. [Google Scholar] [CrossRef]
- Van Niftrik, L.; Jetten, M.S.M. Anaerobic Ammonium-Oxidizing Bacteria: Unique Microorganisms with Exceptional Properties. Microbiol. Mol. Biol. Rev. 2012, 76, 585–596. [Google Scholar] [CrossRef] [Green Version]
- Russ, L.; Speth, D.R.; Jetten, M.S.M.; Op den Camp, H.J.M.; Kartal, B. Interactions between anaerobic ammonium and sulfur-oxidizing bacteria in a laboratory scale model system. Environ. Microbiol. 2014, 16, 3487–3498. [Google Scholar] [CrossRef] [PubMed]
- Atanasova, N.S.; Pietilä, M.K.; Oksanen, H.M. Diverse antimicrobial interactions of halophilic archaea and bacteria extend over geographical distances and cross the domain barrier. Microbiologyopen 2013, 2, 811–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Galinski, E.A.; Grant, W.D.; Oren, A.; Ventosa, A. Halophiles 2010: Life in saline environments. Appl. Environ. Microbiol. 2010, 76, 6971–6981. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, C.C.C.R.C.R.; Fernandes, P. Production of metabolites as bacterial responses to the marine environment. Mar. Drugs 2010, 8, 705–727. [Google Scholar] [CrossRef]
- Du, L.; Li, D.; Zhu, T.; Cai, S.; Wang, F.; Xiao, X.; Gu, Q. New alkaloids and diterpenes from a deep ocean sediment derived fungus Penicillium sp. Tetrahedron 2009, 65, 1033–1039. [Google Scholar] [CrossRef]
- Manivasagan, P.; Kang, K.-H.H.; Sivakumar, K.; Li-Chan, E.C.Y.Y.; Oh, H.-M.M.; Kim, S.-K.K. Marine actinobacteria: An important source of bioactive natural products. Environ. Toxicol. Pharmacol. 2014, 38, 172–188. [Google Scholar] [CrossRef] [PubMed]
- Baltz, R.H. Gifted microbes for genome mining and natural product discovery. J. Ind. Microbiol. Biotechnol. 2016, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Thornburg, C.C.; Zabriskie, T.M.; McPhail, K.L.; Mark Zabriskie, T.; McPhail, K.L. Deep-sea hydrothermal vents: Potential hot spots for natural products discovery. J. Nat. Prod. 2010, 73, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.B.; de la Torre, J.R.; Klotz, M.G.; Urakawa, H.; Pinel, N.; Arp, D.J.; Brochier-Armanet, C.; Chain, P.S.G.; Chan, P.P.; Gollabgir, A.; et al. Nitrosopumilus maritimus genome reveals unique mechanisms for nitrification and autotrophy in globally distributed marine crenarchaea. Proc. Natl. Acad. Sci. USA 2010, 107, 8818–8823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Sher, D.; Kelly, L.; Shi, Y.; Huang, K.; Knerr, P.J.; Joewono, I.; Rusch, D.; Chisholm, S.W.; van der Donk, W.A. Catalytic promiscuity in the biosynthesis of cyclic peptide secondary metabolites in planktonic marine cyanobacteria. Proc. Natl. Acad. Sci. USA 2010, 107, 10430–10435. [Google Scholar] [CrossRef] [Green Version]
- Kojima, H.; Tokizawa, R.; Fukui, M. Mizugakiibacter sediminis gen. nov., sp. nov., isolated from a freshwater lake. Int. J. Syst. Evol. Microbiol. 2014, 64, 3983–3987. [Google Scholar] [CrossRef]
- Romanenko, L.A.; Tanaka, N.; Kurilenko, V. V; Svetashev, V.I.; Lyudmila, C.; Romanenko, A. Tamlana sedimentorum sp. nov., isolated from shallow sand sediments of the Sea of Japan. Int. J. Syst. Evol. Microbiol. 2014, 64, 2891–2896. [Google Scholar] [CrossRef]
- Nadeem, F.; Oves, M.; Qari, H.; Ismail, I. Red Sea Microbial Diversity for Antimicrobial and Anticancer Agents. J. Mol. Biomark. Diagn. 2015, 7, 1–14. [Google Scholar]
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27. [Google Scholar] [CrossRef] [PubMed]
- Niu, B.; Fu, L.; Sun, S.; Li, W. Artificial and natural duplicates in pyrosequencing reads of metagenomic data. BMC Bioinform. 2010, 11, 187. [Google Scholar] [CrossRef] [PubMed]
- Margulies, M.; Egholm, M.; Altman, W.E.; Attiya, S.; Bader, J.S.; Bemben, L.A.; Berka, J.; Braverman, M.S.; Chen, Y.-J.; Chen, Z.; et al. Genome sequencing in microfabricated high-density picolitre reactors. Nature 2005, 437, 376–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilke, A.; Bischof, J.; Harrison, T.; Brettin, T.; D’Souza, M.; Gerlach, W.; Matthews, H.; Paczian, T.; Wilkening, J.; Glass, E.M.; et al. A RESTful API for Accessing Microbial Community Data for MG-RAST. PLoS Comput. Biol. 2015, 11, e1004008. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.T.; Fischbach, M.A. Natural products version 2.0: Connecting genes to molecules. J. Am. Chem. Soc. 2010, 132, 2469–2493. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Detailed Description | Sites | Reference Sampling & Read Sequences/Assembly | Number of Reads | Number of Reads after Trimming | Number of Assembled Reads | Number of Contigs > 500 bp | Average Contig Size (bp) | Largest Contig Size (bp) |
|---|---|---|---|---|---|---|---|---|---|
| Atlantis II water column (Water above brine pool) | Atlantis II 50 m water column | ATII 50 | [19]/This study | 1,461,910 | 1,461,904 | 582,768 | 36,262 | 1149 | 21,887 |
| Atlantis II 200 m water column | ATII 200 | 1,260,578 | 1,260,561 | 530,441 | 34,640 | 1131 | 25,392 | ||
| Atlantis II 700 m water column | ATII 700 | 1,128,514 | 1,128,507 | 554,335 | 32,860 | 1285 | 33,783 | ||
| Atlantis II 1500 m water column | ATII 1500 | 833,739 | 833,730 | 316,101 | 20,374 | 1331 | 51,927 | ||
| Atlantis II brine water | Atantis II brine-seawater interface | ATII INF | [20]/This study | 832,138 | 832,128 | 743,064 | 9933 | 1214 | 25,150 |
| Atantis II brine Upper Convective Layer | ATII UCL | 886,030 | 886,019 | 794,715 | 11,994 | 1454 | 103,389 | ||
| Atantis II brine Lower Convective Layer | ATII LCL | 4,104,966 | 4,104,994 | 3,901,967 | 19,165 | 2084 | 350,936 | ||
| Discovery Deep brine water | Discovery Deep brine-seawater interface | DD INF | [20]/This study | 1,095,181 | 1,095,157 | 752,025 | 14,144 | 1201 | 28,080 |
| Discovery Deep brine water | DD BR | 1,111,044 | 1,111,032 | 763,387 | 15,306 | 1216 | 22,118 | ||
| Kebrit Deep brine water | Kebrit Deep Upper brine-seawater interface | KD UINF | [20]/This study | 1,562,521 | 1,562,512 | 1,020,749 | 24,517 | 1495 | 58,542 |
| Kebrit Deep Lower brine-seawater interface | KD LINF | 1,510,272 | 1,510,262 | 926,337 | 31,983 | 1241 | 38,825 | ||
| Kebrit Deep brine water | KD BR | 1,379,832 | 1,379,814 | 913,803 | 22,280 | 945 | 14,864 | ||
| Other water metagenomes | Guaymas Basin deep-sea hydrothermal vent plume water | GB VNT | [39,40,41]/This study | 628,619 | 628,569 | 155,841 | 7654 | 1082 | 17,353 |
| Kueishantao shallow-sea hydrothermal vent (water above vent) | KSW VNT | [42]/This study | 261,446 | 261,399 | 199,237 | 411 | 4685 | 179,360 | |
| Kueishantao shallow-sea hydrothermal vent (water) | K VNT | [42]/This study | 444,655 | 444,597 | 338,480 | 2194 | 1843 | 88,498 | |
| Juan de Fuca Ridge hydrothermal vent diffuse flow seawater | JDF VNT | [43]/This study | 226,981 | 226,916 | 35,357 | 9366 | 1135 | 9366 | |
| Sediments | Atlantis II sediments | ATII SDM | [28,36]/This study | 1,138,406 | 1,138,381 | 478,453 | 30,352 | 1194 | 33,674 |
| Discovery Deep sediments | DD SDM | 1,258,290 | 1,258,273 | 597,552 | 38,529 | 1233 | 38,081 | ||
| Non-brine sediments | NB SDM | 253,568 | 253,564 | 92,530 | 7292 | 1177 | 1315 | ||
| Other metagenome (biofilm) | Loki’s Castle deep-sea vent biofilm (microbial mat) | LC MM | [44]/This study | 717,550 | 717,135 | 525,719 | 7897 | 1546 | 42,387 |
| Total | 22,096,240 | 22,095,454 | 14,222,861 | 377,153 | - | - |
| Detailed Description | Assembly | Number of SMGCs | Normalized Number of SMGCs * | Types of SMGCs | Number of Phyla | SMGCs Detected Uniquely Once at a Particular Site |
|---|---|---|---|---|---|---|
| Red Sea metagenomic samples: | ||||||
| Atlantis II 1500 m water column | ATII 1500 | 168 | 531.48 | 9 | 33 | Otherks-Pufa-T1pks, T2pks-Cf_fatty_acid |
| Atlantis II 700 m water column | ATII 700 | 269 | 485.27 | 8 | 32 | Otherks-Pufa, Otherks-T1pks |
| Kebrit Deep Lower brine-seawater interface | KD LINF | 334 | 360.56 | 9 | 33 | Cf_saccharide-Bacteriocin, Hserlactone |
| Atlantis II 200 m water column | ATII 200 | 170 | 320.49 | 8 | 30 | Cf_fatty_acid-Arylpolyene |
| Atantis II brine Upper Convective Layer | ATII UCL | 210 | 264.25 | 7 | 21 | |
| Atlantis II 50 m water column | ATII 50 | 146 | 250.53 | 8 | 30 | |
| Kebrit Deep Upper brine-seawater interface | KD UINF | 252 | 246.88 | 13 | 32 | Ladderane-Cf_fatty_acid, Nrps-T1pks, T1PKS |
| Atantis II brine-seawater interface | ATII INF | 162 | 218.02 | 6 | 19 | |
| Discovery Deep sediments | DD SDM | 114 | 190.78 | 4 | 23 | |
| Non-brine sediments | NB SDM | 16 | 172.92 | 4 | 9 | |
| Kebrit Deep brine water | KD BR | 149 | 163.05 | 4 | 23 | |
| Atlantis II sediments | ATII SDM | 70 | 146.30 | 7 | 26 | |
| Atantis II brine Lower Convective Layer | ATII LCL | 524 | 134.29 | 13 | 25 | Cf_fatty_acid-Cf_saccharide, Cf_saccharide-nrps, Phosphonate, T3pks-cf_saccharide |
| Discovery Deep brine-seawater interface | DD INF | 94 | 125.00 | 1 | 20 | |
| Discovery Deep brine water | DD BR | 73 | 95.63 | 2 | 20 | |
| Other metagenomic samples: | ||||||
| Guaymas Basin deep-sea hydrothermal vent plume water | GB VNT | 11 | 70.58 | 3 | 30 | Pufa |
| Kueishantao shallow-sea hydrothermal vent (water above vent) | KSW VNT | 11 | 55.21 | 4 | 12 | Thiopeptide |
| Kueishantao shallow-sea hydrothermal vent (water) | K VNT | 10 | 29.54 | 5 | 13 | |
| Juan de Fuca Ridge hydrothermal vent diffuse flow seawater | JDF VNT | 3 | 84.85 | 3 | 14 | |
| Loki’s Castle deep-sea vent biofilm (microbial mat) | LC MM | 26 | 49.46 | 5 | 23 | Acyl_amino_acids |
| General Functional Classification: | Product (Enzyme) | Gene Cluster Names | Representative Basic Structure | Potential Function/Application of Product | Percentage of Total SMGCs | |
|---|---|---|---|---|---|---|
| 1. Products of predicted functions commonly abundant in microbes | Saccharide | Cf_saccharide Cf_saccharide-Bacteriocin Cf_saccharide-nrps Cf_fatty_acid-Cf_saccharide T3pks-cf_saccharide | - | Microbe-host interactions e.g. lipopolysaccharides. Some saccharides that are diffusible were reported to have antibacterial activities [46,47]. | 80.61% | |
| Fatty Acid | Cf_fatty_acid Arylpolyene-cf_fatty_acid Cf_fatty_acid-Arylpolyene Cf_fatty_acid-Cf_saccharide Ladderane-Cf_fatty_acid T2pks-Cf_fatty_acid | - | Structural functions and reported that composition can change as an adaptation to temperature and pressure in the environment [48]. | 7.69% | ||
| Aryl polyenes | Arylpolyene Arylpolyene-cf_fatty_acid Cf_fatty_acid-Arylpolyene |  | Aryl polyene SMGCs found in abundance in Gram negative bacteria. Previously reported to have a protective role from damage caused by reactive oxygen species [46,49]. | 0.52% | ||
| Acyl-homoserine lactones | Hserlactone |  | Quorum sensing [50]. | 0.03% | ||
| 2. Subset of products with potential antibacterial and/or anticancer effects: | Terpenes | Terpene |  | A subset of the terpenes possesses antibacterial effect [51]. | 1.89% | |
| Peptides | Ribosomal peptides | Bacteriocin Cf_saccharide-Bacteriocin Microcin Lantipeptide |  | Some have antibacterial activity, and some have selective cancer cytotoxic activity [52]. | 0.78% | |
| Non-ribosomal peptides | Cf_saccharide-nrps NRPS Nrps-T1pks | - | Many non-ribosomal peptides have antibacterial (e.g., β-lactams) and anticancer (e.g. bleomycin) effects [53]. | 0.25% | ||
| Polyketides | (Type I Polyketide synthase) | Nrps-T1pks Otherks-Pufa-T1pks Otherks-T1pks T1PKS | - | A subset are responsible for antibiotic synthesis e.g. the type I polyketide synthase (PKSI) producing erythromycin [54]. | 0.2% | |
| (Type II Polyketide synthase) | T2pks-Cf_fatty_acid | - | Some type II polyketide synthase (PKSII) enzymes produce aromatic polyketide antibiotics e.g. oxytetracycline [54]. | 0.09% | ||
| (Type III Polyketide synthase) | T3pks T3pks-cf_saccharide | - | Type III Polyketide synthase (PKSIII) enzymes are capable of producing an array of compounds including pyrones—a subset of pyrones were previously reported to have antibacterial and anticancer effects [55]. | 0.31% | ||
| Phosphonates | Phosphonate |  | Some natural phosphonates are antibacterials e.g. fosfomycin. Also have structural functions [56]. | 0.01% | ||
| 3. Miscellaneous: products are predicted to confer adaptation to the environment/special structure/unknown function: | Others | Cf_putative Other OtherKS Otherks-Pufa Otherks-Pufa-T1pks Otherks-T1pks | - | Some code for biosynthetic gene clusters of types that are still unknown [9]. | 8.13% | |
| Polyunsaturated fatty acids | Otherks-Pufa Otherks-Pufa-T1pks |  | Polyunsaturated fatty acids (PUFAs) are membrane adaptations to piezophiles, thermophiles and psychrophiles to prevent membrane crystallization [57,58]. | 0.14% | ||
| Ectoine | Ectoine |  | Halophilic adaptation & effective in vitro in preventing protein misfolding characteristic in diseases e.g. Alzheimer’s [59]. | 0.08% | ||
| Ladderane | Ladderane-Cf_fatty_acid |  | Unique component of anammoxosome membrane in anammox (anaerobic ammonium oxidizing) bacteria and potential biofuel [60]. | 0.05% | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ziko, L.; Adel, M.; Malash, M.N.; Siam, R. Insights into Red Sea Brine Pool Specialized Metabolism Gene Clusters Encoding Potential Metabolites for Biotechnological Applications and Extremophile Survival. Mar. Drugs 2019, 17, 273. https://doi.org/10.3390/md17050273
Ziko L, Adel M, Malash MN, Siam R. Insights into Red Sea Brine Pool Specialized Metabolism Gene Clusters Encoding Potential Metabolites for Biotechnological Applications and Extremophile Survival. Marine Drugs. 2019; 17(5):273. https://doi.org/10.3390/md17050273
Chicago/Turabian StyleZiko, Laila, Mustafa Adel, Mohamed N. Malash, and Rania Siam. 2019. "Insights into Red Sea Brine Pool Specialized Metabolism Gene Clusters Encoding Potential Metabolites for Biotechnological Applications and Extremophile Survival" Marine Drugs 17, no. 5: 273. https://doi.org/10.3390/md17050273