Identification of an LPS-Induced Chemo-Attractive Peptide from Ciona robusta

 , , and
, , and
Abstract
:1. Introduction
2. Results
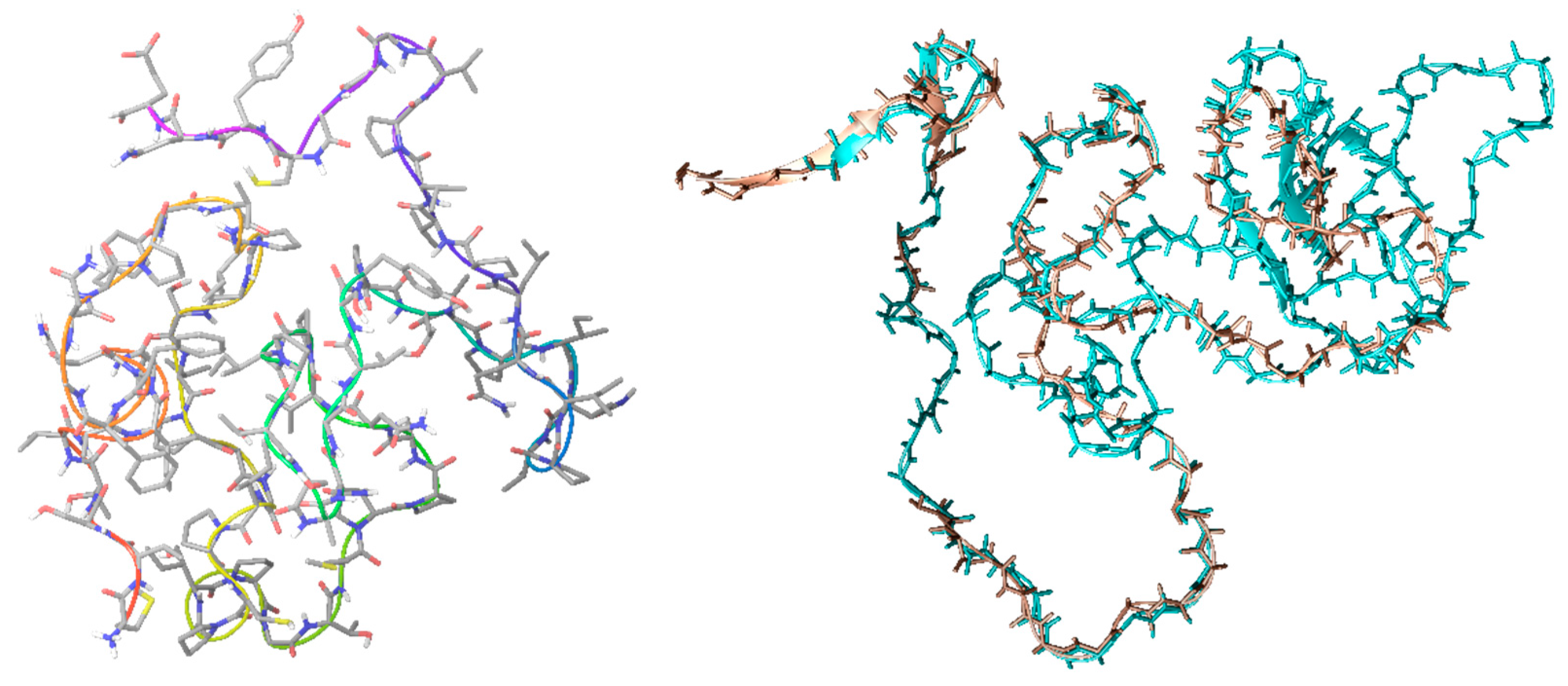
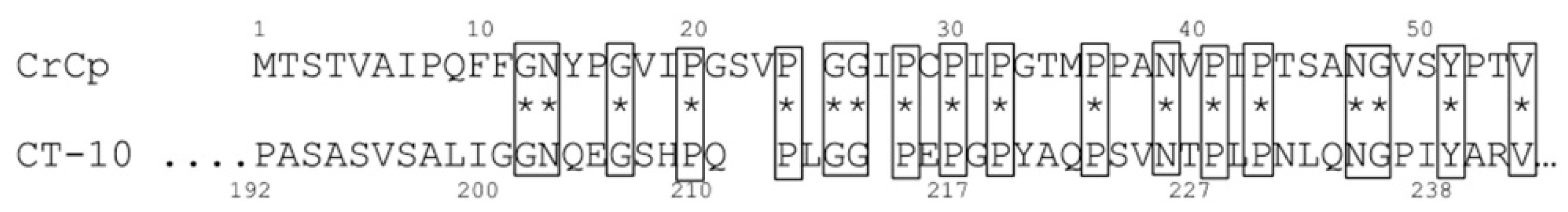
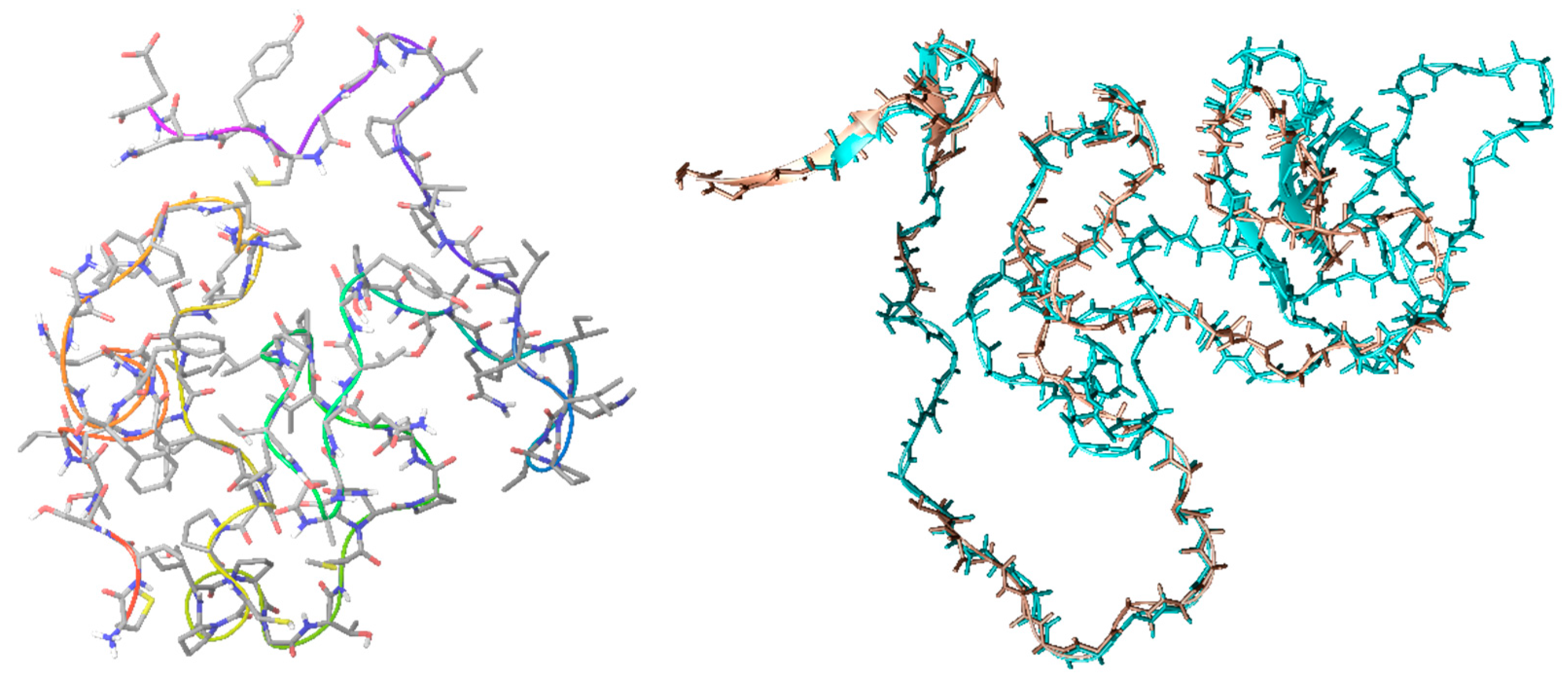
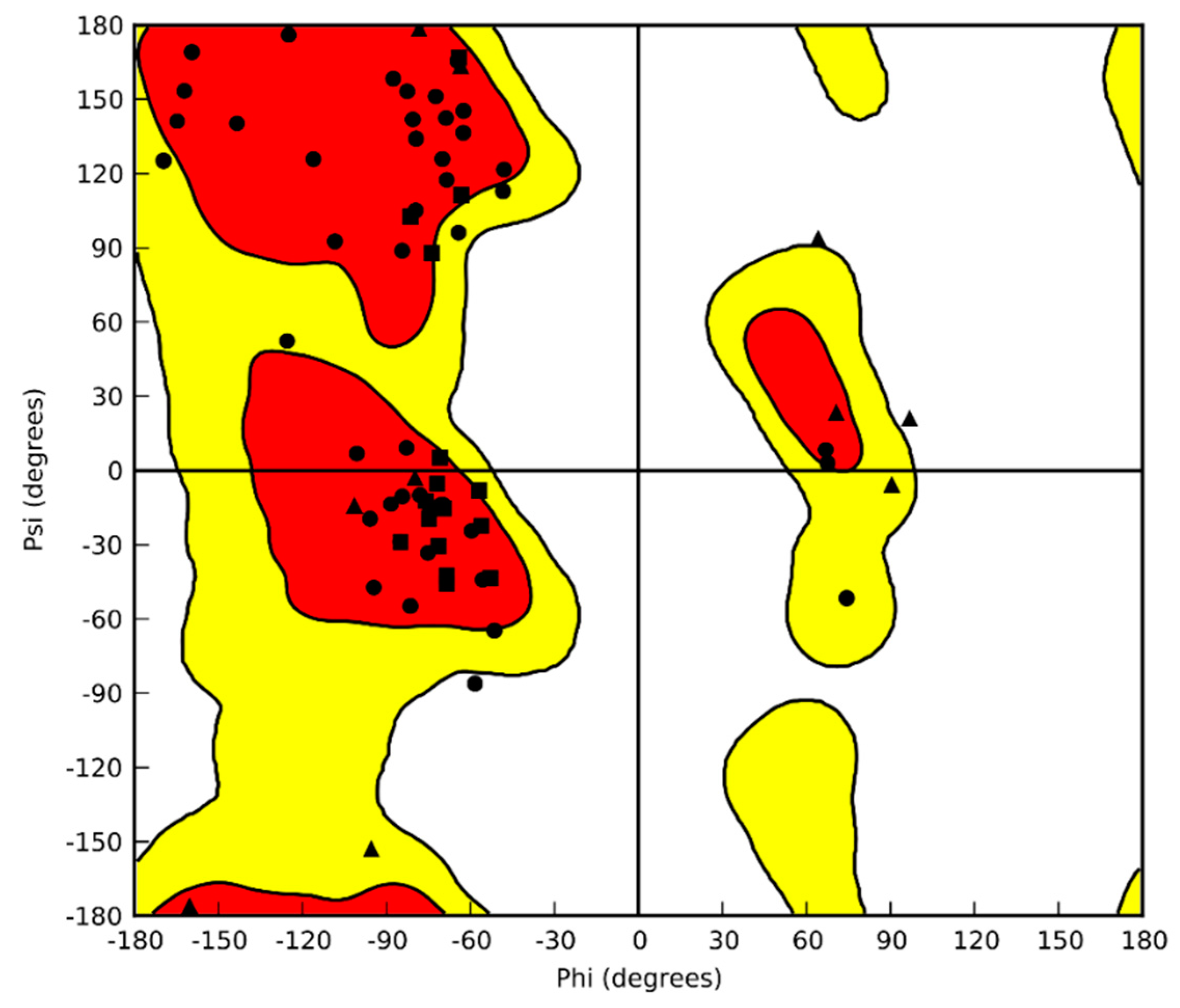
2.1. CrCP Structure and Homology
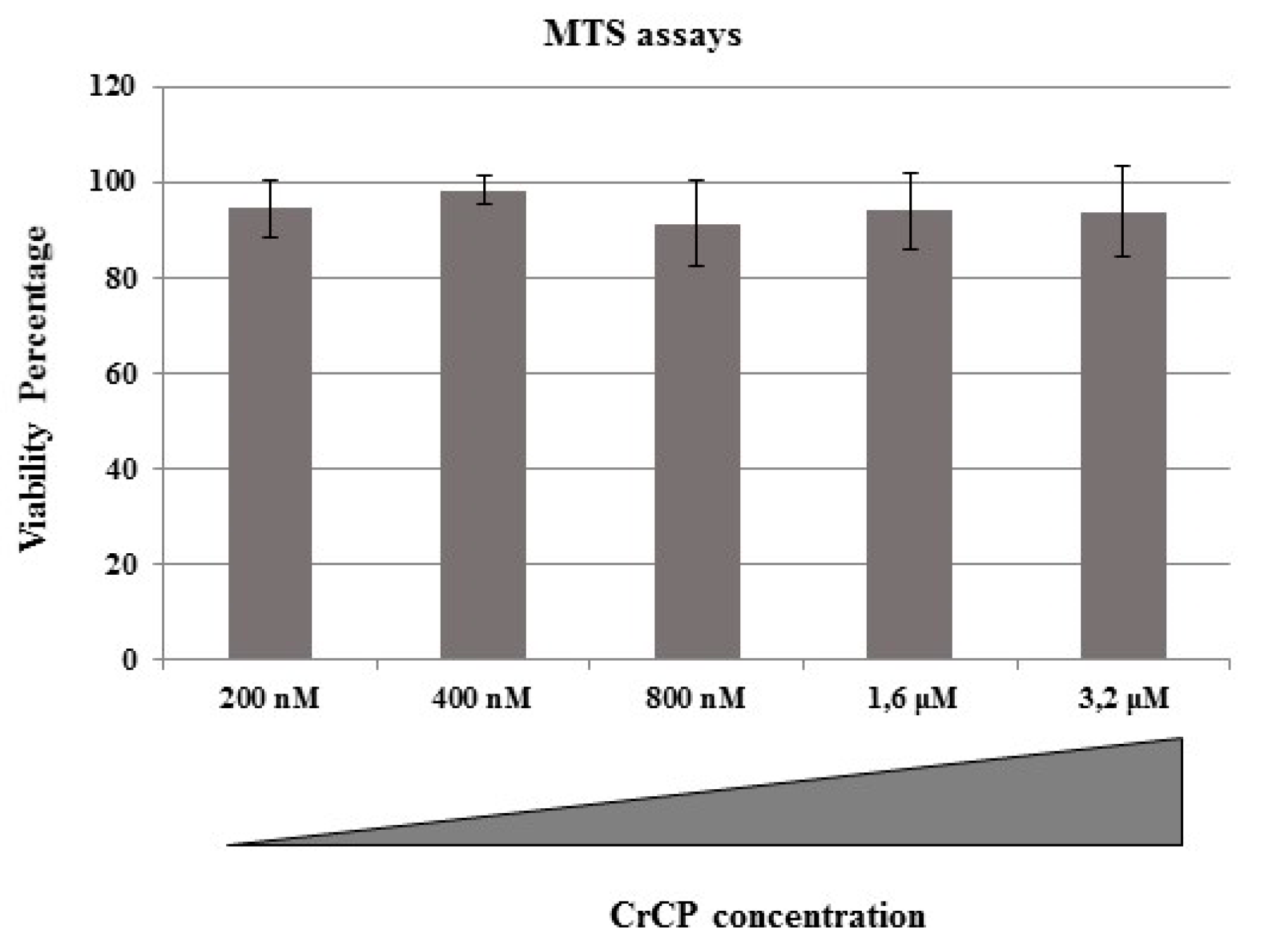
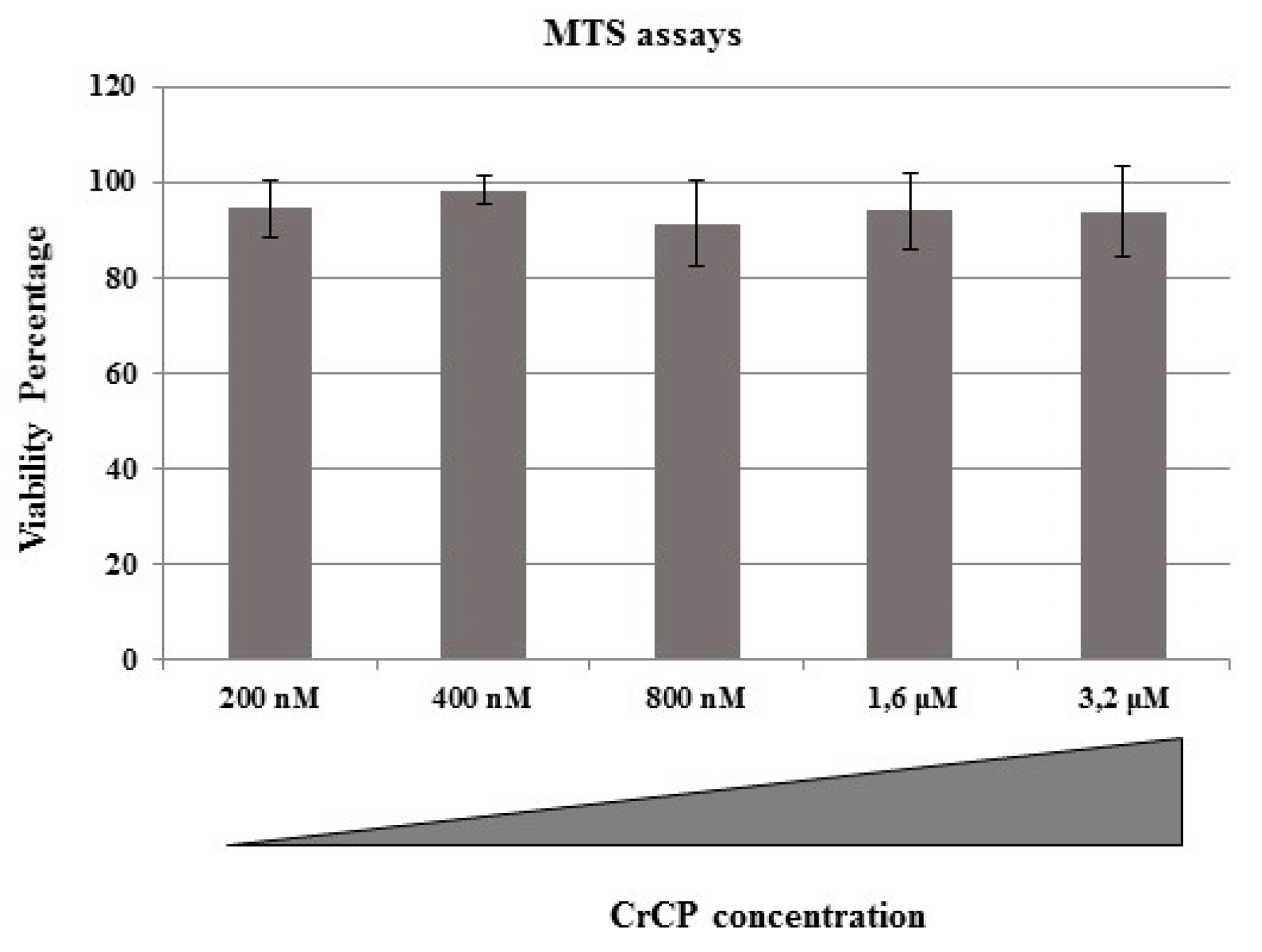
2.2. Cytotoxic Effects of CrCP on HuDe Cell Line Viability
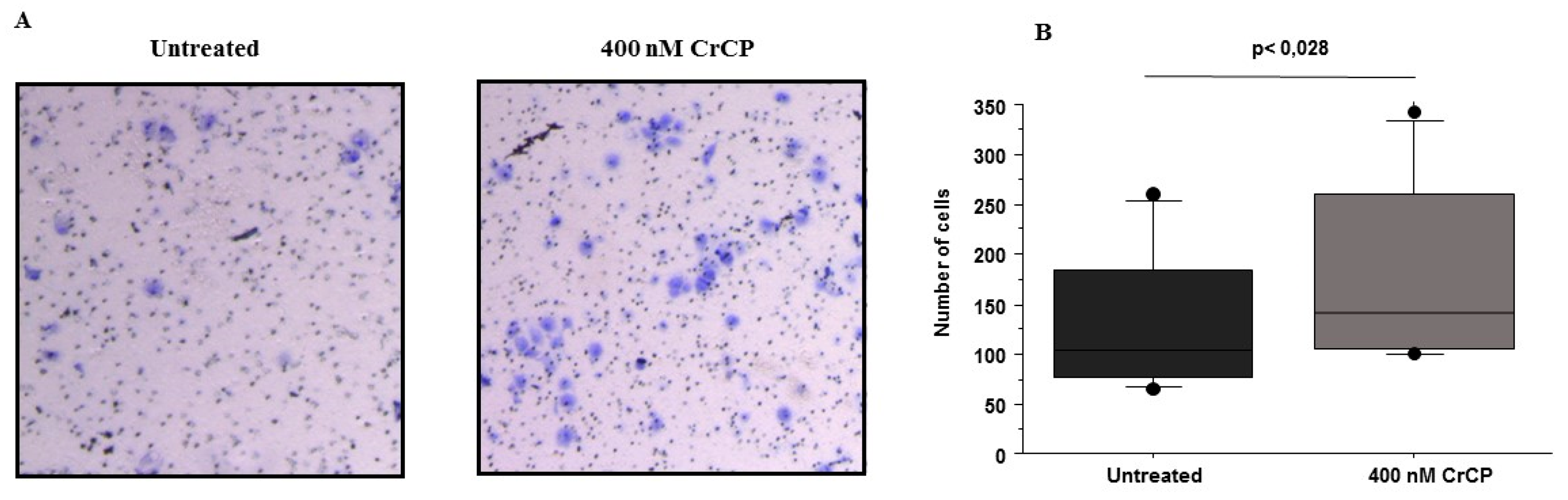
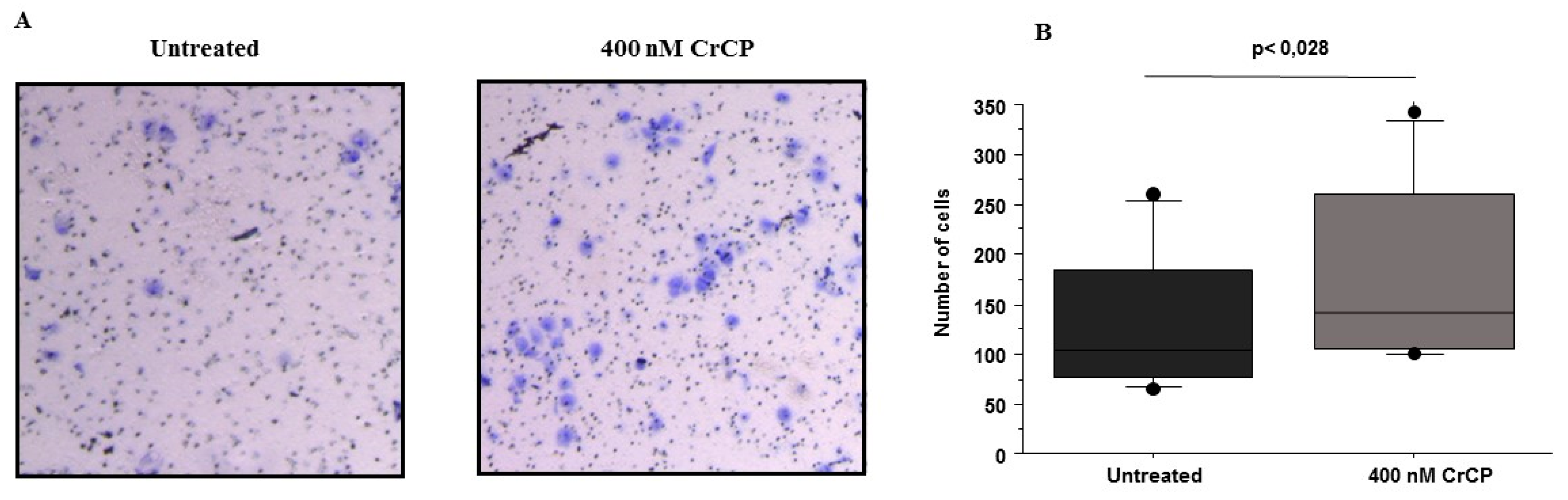
2.3. Effects of CrCP on HuDe Cell Motility
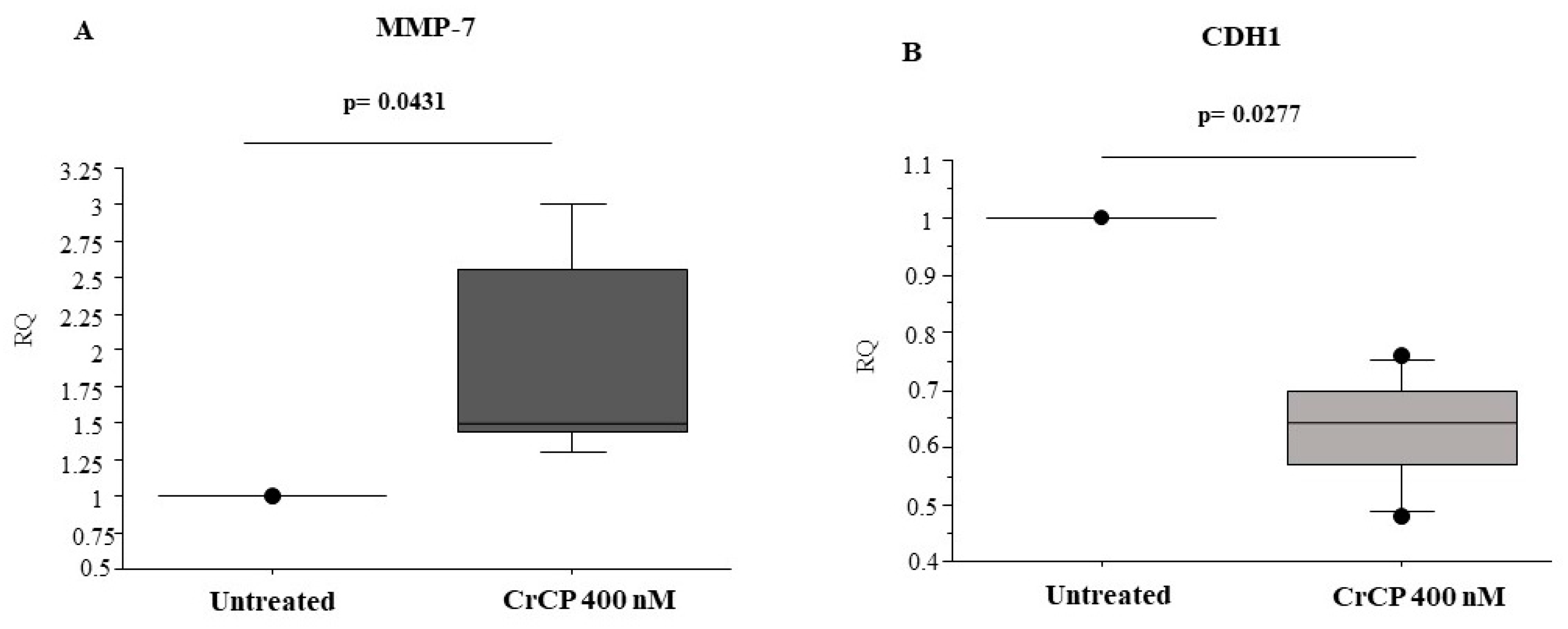
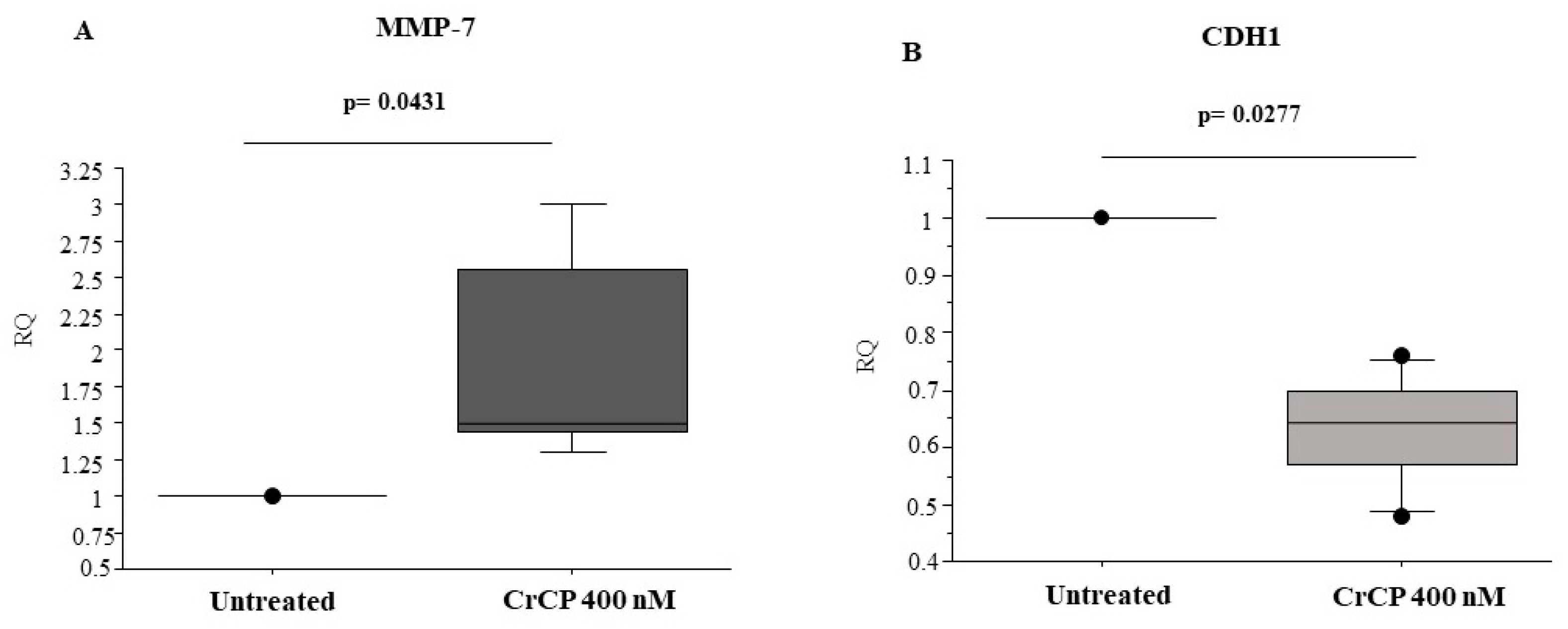
2.4. CrCP Modulates the Expression of Cell Motility Genes
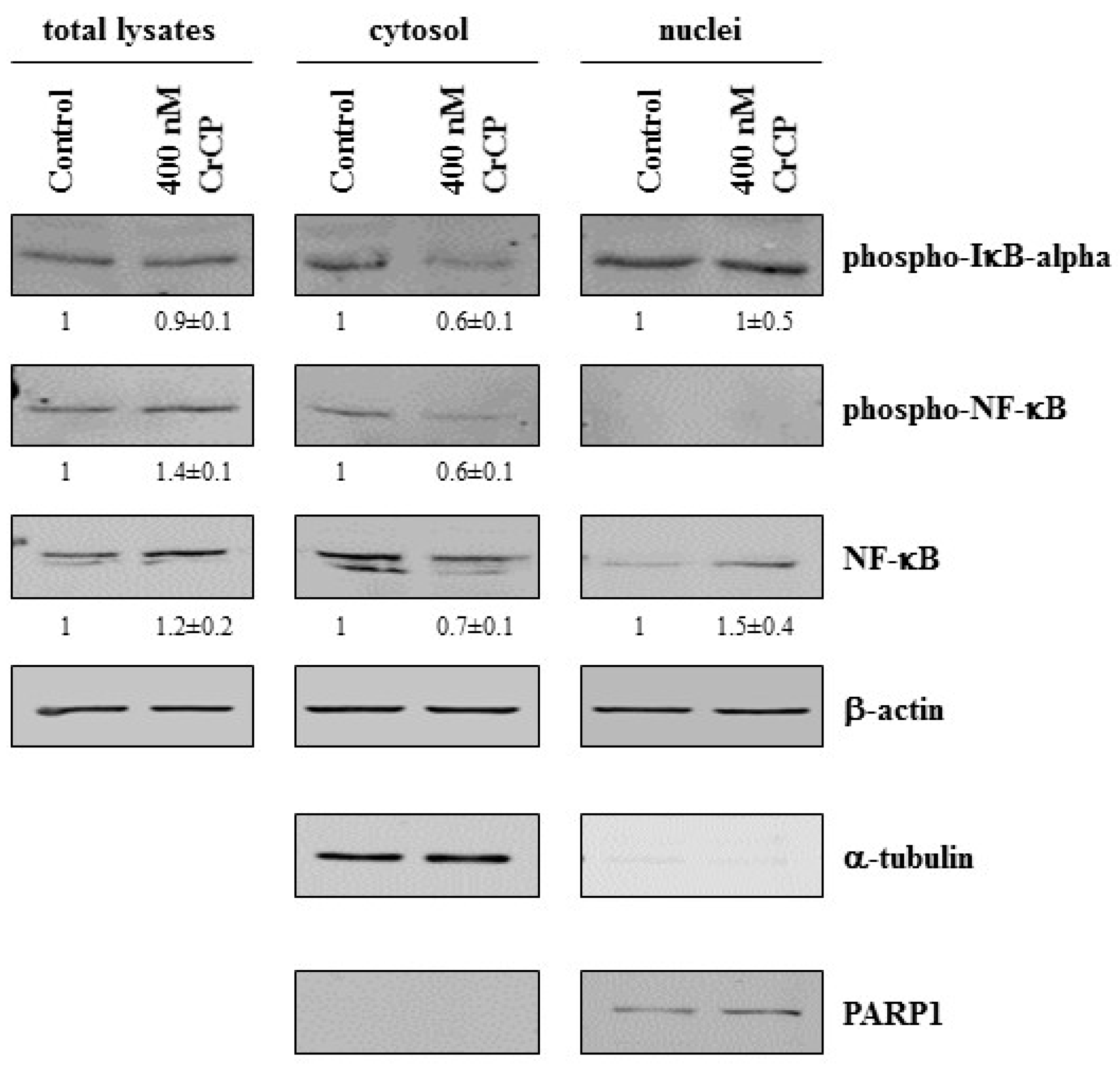
2.5. Western Blot Analysis
3. Discussion
4. Material and Methods
4.1. Homology Modeling
4.2. Molecular Dynamic Simulation
4.3. Cell Culture and Materials
4.4. MTS Assay
4.5. Wound-Healing Assay
4.6. Migration Assay
4.7. Western Blotting Analyses
4.8. Total RNA Preparation and qPCR Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bourlat, S.J.; Juliusdottir, T.; Lowe, C.J.; Freeman, R.; Aronowicz, J.; Kirschner, M.; Lander, E.S.; Thorndyke, M.; Nakano, H.; Kohn, A.B.; et al. Deuterostome phylogeny reveals monophyletic chordates and the new phylum Xenoturbellida. Nature 2006, 444, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Delsuc, F.; Brinkmann, H.; Chourrout, D.; Philippe, H. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 2006, 439, 965–968. [Google Scholar] [CrossRef] [PubMed]
- Franchi, N.; Ballarin, L. Immunity in Protochordates: The Tunicate Perspective. Front. Immunol. 2017, 8, 674. [Google Scholar] [CrossRef] [PubMed]
- Pennati, R.; Ficetola, G.F.; Brunetti, R.; Caicci, F.; Gasparini, F.; Griggio, F.; Sato, A.; Stach, T.; Kaul-Strehlow, S.; Gissi, C.; et al. Morphological Differences between Larvae of the Ciona intestinalis Species Complex: Hints for a Valid Taxonomic Definition of Distinct Species. PLoS ONE 2015, 10, e0122879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, C.; Lemaire, L.A.; Wang, W.; Yoon, P.H.; Choi, Y.A.; Parsons, L.R.; Matese, J.C.; Wang, W.; Levine, M.; Chen, K. Comprehensive single-cell transcriptome lineages of a proto-vertebrate. Nature 2019, 571, 349–354. [Google Scholar] [CrossRef]
- Horie, T.; Horie, R.; Chen, K.; Cao, C.; Nakagawa, M.; Kusakabe, T.G.; Satoh, N.; Sasakura, Y.; Levine, M. Regulatory cocktail for dopaminergic neurons in a protovertebrate identified by whole-embryo single-cell transcriptomics. Genes Dev. 2018, 32, 1297–1302. [Google Scholar] [CrossRef] [Green Version]
- Imai, K.S.; Levine, M.; Satoh, N.; Satou, Y. Regulatory blueprint for a chordate embryo. Science 2006, 312, 1183–1187. [Google Scholar] [CrossRef]
- Madgwick, A.; Magri, M.S.; Dantec, C.; Gailly, D.; Fiuza, U.M.; Guignard, L.; Hettinger, S.; Gomez-Skarmeta, J.L.; Lemaire, P. Evolution of embryonic cis-regulatory landscapes between divergent Phallusia and Ciona ascidians. Dev. Biol. 2019, 448, 71–87. [Google Scholar] [CrossRef]
- Parrinello, N.; Patricolo, E.; Canicatii, C. Inflammatory-like reaction in the tunic of Ciona intestinalis (Tunicata). Encapsulation tissue injury II. Biol. Bull. 1984, 167, 238–250. [Google Scholar] [CrossRef]
- Parrinello, N.; Patricolo, E.; Canicattì, C. Inflammatory-like reaction in the tunic of Ciona intestinalis (Tunicata). Encapsulation and tissue injury I. Biol. Bull. 1984, 167, 229–237. [Google Scholar] [CrossRef]
- Parrinello, N.; Rindone, D. Studies on the natural hemolytic system of the annelid worm Spirographis spallanzanii viviani (Polychaeta). Dev. Comp. Immunol. 1981, 5, 33–42. [Google Scholar] [CrossRef]
- Parrinello, N.; Arizza, V.; Cammarata, M.; Giaramita, F.T.; Pergolizzi, M.; Vazzana, M.; Vizzini, A.; Parrinello, D. Inducible lectins with galectin properties and human IL1alpha epitopes opsonize yeast during the inflammatory response of the ascidian Ciona intestinalis. Cell Tissue Res. 2007, 329, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Giacomelli, S.; Melillo, D.; Lambris, J.D.; Pinto, M.R. Immune competence of the Ciona intestinalis pharynx: Complement system-mediated activity. Fish Shellfish Immunol. 2012, 33, 946–952. [Google Scholar] [CrossRef] [PubMed]
- Parrinello, N.; Cammarata, M.; Parrinello, D. The Inflammatory Response of Urochordata: The Basic Process of the Ascidians’ Innate Immunity. In Advances in Comparative Immunology; Springer: Cham, Switzerland, 2018; p. 69. [Google Scholar]
- Pinto, M.R.; Chinnici, C.M.; Kimura, Y.; Melillo, D.; Marino, R.; Spruce, L.A.; De Santis, R.; Parrinello, N.; Lambris, J.D. CiC3-1a-mediated chemotaxis in the deuterostome invertebrate Ciona intestinalis (Urochordata). J. Immunol. 2003, 171, 5521–5528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cammarata, M.; Arizza, V.; Cianciolo, C.; Parrinello, D.; Vazzana, M.; Vizzini, A.; Salerno, G.; Parrinello, N. The prophenoloxidase system is activated during the tunic inflammatory reaction of Ciona intestinalis. Cell Tissue Res. 2008, 333, 481–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrinello, N.; Arizza, V.; Chinnici, C.; Parrinello, D.; Cammarata, M. Phenoloxidases in ascidian hemocytes: Characterization of the pro-phenoloxidase activating system. Comp. Biochem. Physiol. 2003, 135, 583–591. [Google Scholar] [CrossRef]
- Bonura, A.; Vizzini, A.; Salerno, G.; Parrinello, N.; Longo, V.; Colombo, P. Isolation and expression of a novel MBL-like collectin cDNA enhanced by LPS injection in the body wall of the ascidian Ciona intestinalis. Mol. Immunol. 2009, 46, 2389–2394. [Google Scholar] [CrossRef] [Green Version]
- Parrinello, D.; Sanfratello, M.A.; Vizzini, A.; Testasecca, L.; Parrinello, N.; Cammarata, M. The Ciona intestinalis immune-related galectin genes (CiLgals-a and CiLgals-b) are expressed by the gastric epithelium. Fish Shellfish Immunol. 2017, 62, 24–30. [Google Scholar] [CrossRef] [Green Version]
- Vizzini, A.; Parrinello, D.; Sanfratello, M.A.; Salerno, G.; Cammarata, M.; Parrinello, N. Inducible galectins are expressed in the inflamed pharynx of the ascidian Ciona intestinalis. Fish Shellfish Immunol. 2012, 32, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Parrinello, N.; Vizzini, A.; Arizza, V.; Salerno, G.; Parrinello, D.; Cammarata, M.; Giaramita, F.T.; Vazzana, M. Enhanced expression of a cloned and sequenced Ciona intestinalis TNFalpha-like (CiTNF alpha) gene during the LPS-induced inflammatory response. Cell Tissue Res. 2008, 334, 305–317. [Google Scholar] [CrossRef] [Green Version]
- Vizzini, A.; Di Falco, F.; Parrinello, D.; Sanfratello, M.A.; Mazzarella, C.; Parrinello, N.; Cammarata, M. Ciona intestinalis interleukin 17-like genes expression is upregulated by LPS challenge. Dev. Comp. Immunol. 2015, 48, 129–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vizzini, A.; Parrinello, D.; Sanfratello, M.A.; Trapani, M.R.; Mangano, V.; Parrinello, N.; Cammarata, M. Upregulated transcription of phenoloxidase genes in the pharynx and endostyle of Ciona intestinalis in response to LPS. J. Invertebr. Pathol. 2015, 126C, 6–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vizzini, A.; Pergolizzi, M.; Vazzana, M.; Salerno, G.; Di Sano, C.; Macaluso, P.; Arizza, V.; Parrinello, D.; Cammarata, M.; Parrinello, N. FACIT collagen (1alpha-chain) is expressed by hemocytes and epidermis during the inflammatory response of the ascidian Ciona intestinalis. Dev. Comp. Immunol. 2008, 32, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Bonura, A.; Vizzini, A.; Salerno, G.; Parrinello, D.; Parrinello, N.; Longo, V.; Montana, G.; Colombo, P. Cloning and expression of a novel component of the CAP superfamily enhanced in the inflammatory response to LPS of the ascidian Ciona intestinalis. Cell Tissue Res. 2010, 342, 411–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liberti, A.; Leigh, B.; De Santis, R.; Pinto, M.R.; Cannon, J.P.; Dishaw, L.J.; Litman, G.W. An Immune Effector System in the Protochordate Gut Sheds Light on Fundamental Aspects of Vertebrate Immunity. Results Probl. Cell Differ. 2015, 57, 159–173. [Google Scholar] [PubMed]
- Parrinello, D.; Parisi, M.; Parrinello, N.; Cammarata, M. Ciona robusta hemocyte populational dynamics and PO-dependent cytotoxic activity. Dev. Comp. Immunol. 2020, 103, 103519. [Google Scholar] [CrossRef]
- Jeffery, W.R. Closing the wounds: One hundred and twenty five years of regenerative biology in the ascidian Ciona intestinalis. Genesis 2015, 53, 48–65. [Google Scholar] [CrossRef] [Green Version]
- Vizzini, A.; Bonura, A.; Parrinello, D.; Sanfratello, M.A.; Longo, V.; Colombo, P. LPS challenge regulates gene expression and tissue localization of a Ciona intestinalis gene through an alternative polyadenylation mechanism. PLoS ONE 2013, 8, e63235. [Google Scholar] [CrossRef]
- Yu, T.; Su, X.; Pan, Y.; Zhuang, H. Receptor-transporting protein (RTP) family members play divergent roles in the functional expression of odorant receptors. PLoS ONE 2017, 12, e0179067. [Google Scholar] [CrossRef] [Green Version]
- Kobashigawa, Y.; Sakai, M.; Naito, M.; Yokochi, M.; Kumeta, H.; Makino, Y.; Ogura, K.; Tanaka, S.; Inagaki, F. Structural basis for the transforming activity of human cancer-related signaling adaptor protein CRK. Nat. Struct. Mol. Biol. 2007, 14, 503–510. [Google Scholar] [CrossRef]
- Mateos, B.; Conrad-Billroth, C.; Schiavina, M.; Beier, A.; Kontaxis, G.; Konrat, R.; Felli, I.C.; Pierattelli, R. The ambivalent role of proline residues in an intrinsically disordered protein: From disorder promoters to compaction facilitators. J. Mol. Biol. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonura, A.; Vizzini, A.; Vlah, S.; Gervasi, F.; Longo, A.; Melis, M.R.; Schildberg, F.A.; Colombo, P. Ci8 short, a novel LPS-induced peptide from the ascidian Ciona intestinalis, modulates responses of the human immune system. Immunobiology 2018, 223, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.R.; Sultan, M.T.; Park, H.J.; Lee, J.M.; Ju, H.W.; Lee, O.J.; Lee, D.J.; Kaplan, D.L.; Park, C.H. NF-kappaB signaling is key in the wound healing processes of silk fibroin. Acta Biomater. 2018, 67, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound repair and regeneration. Nature 2008, 453, 314–321. [Google Scholar] [CrossRef]
- Srinivasan, M.; Dunker, A.K. Proline rich motifs as drug targets in immune mediated disorders. Int. J. Pept. 2012, 2012, 634769. [Google Scholar] [CrossRef] [Green Version]
- Antoku, S.; Mayer, B.J. Distinct roles for Crk adaptor isoforms in actin reorganization induced by extracellular signals. J. Cell Sci. 2009, 122 Pt 22, 4228–4238. [Google Scholar] [CrossRef] [Green Version]
- Feller, S.M. Crk family adaptors-signalling complex formation and biological roles. Oncogene 2001, 20, 6348–6371. [Google Scholar] [CrossRef] [Green Version]
- Wright, P.E.; Dyson, H.J. Intrinsically disordered proteins in cellular signalling and regulation. Nature reviews. Mol. Cell Biol. 2015, 16, 18–29. [Google Scholar]
- Burighel, P.C. Urochordata: Ascidiacea. Microsc. Anat. Invertebr. 1997, 15, 126. [Google Scholar]
- Martin, P. Wound healing—Aiming for perfect skin regeneration. Science 1997, 276, 75–81. [Google Scholar] [CrossRef]
- Singer, A.J.; Clark, R.A. Cutaneous wound healing. N. Engl. J. Med. 1999, 341, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Shin, S.J.; Kim, K.O.; Kim, M.K.; Hyun, M.S.; Kim, T.N.; Jang, B.I.; Kim, S.W.; Song, S.K.; Kim, H.S.; et al. Relationship between E-cadherin, matrix metalloproteinase-7 gene expression and clinicopathological features in gastric carcinoma. Oncol. Rep. 2006, 16, 823–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caley, M.P.; Martins, V.L.; O’Toole, E.A. Metalloproteinases and Wound Healing. Adv. Wound Care 2015, 4, 225–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Gugel, E.; Saxena, M.; Bhardwaj, N. Modulation of innate immunity in the tumor microenvironment. Cancer Immunol. Immunother. 2016, 65, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Fingleton, B. Matrix metalloproteinases as regulators of inflammatory processes. Biochimica et biophysica acta. Mol. Cell Res. 2017, 1864, 2036–2042. [Google Scholar]
- Dunsmore, S.E.; Saarialho-Kere, U.K.; Roby, J.D.; Wilson, C.L.; Matrisian, L.M.; Welgus, H.G.; Parks, W.C. Matrilysin expression and function in airway epithelium. J. Clin. Investig. 1998, 102, 1321–1331. [Google Scholar] [CrossRef] [Green Version]
- McGuire, J.K.; Li, Q.; Parks, W.C. Matrilysin (matrix metalloproteinase-7) mediates E-cadherin ectodomain shedding in injured lung epithelium. Am. J. Pathol. 2003, 162, 1831–1843. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.E.; Parks, W.C. Metalloproteinases and their inhibitors: Regulators of wound healing. Int. J. Biochem. Cell Biol. 2008, 40, 1334–1347. [Google Scholar] [CrossRef] [Green Version]
- Hulpiau, P.; van Roy, F. Molecular evolution of the cadherin superfamily. Int. J. Biochem. Cell Biol. 2009, 41, 349–369. [Google Scholar] [CrossRef]
- Chen, L.-F.; Greene, W.C. Shaping the nuclear action of NF-κB. Nat. Rev. Mol. Cell Biol. 2004, 5, 392–401. [Google Scholar] [CrossRef]
- Na, J.; Lee, K.; Na, W.; Shin, J.Y.; Lee, M.J.; Yune, T.Y.; Lee, H.K.; Jung, H.S.; Kim, W.S.; Ju, B.G. Histone H3K27 Demethylase JMJD3 in Cooperation with NF-kappaB Regulates Keratinocyte Wound Healing. J. Investig. Dermatol. 2016, 136, 847–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banks, J.L.; Beard, H.S.; Cao, Y.; Cho, A.E.; Damm, W.; Farid, R.; Felts, A.K.; Halgren, T.A.; Mainz, D.T.; Maple, J.R.; et al. Integrated Modeling Program, Applied Chemical Theory (IMPACT). J. Comput. Chem. 2005, 26, 1752–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cusimano, A.; Puleio, R.; D’Alessandro, N.; Loria, G.R.; McCubrey, J.A.; Montalto, G.; Cervello, M. Cytotoxic activity of the novel small molecule AKT inhibitor SC66 in hepatocellular carcinoma cells. Oncotarget 2015, 6, 1707–1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Human Gene | Acronym | Genome Sequence | Code Number |
|---|---|---|---|
| β-actin | ACT | NM_001101 | QT00095431 |
| E-cadherin | CDH1 | NM_004360 | QT00080143 |
| Matrix metalloprotease-7 | MMP-7 | NM_002423 | QT00001456 |
| Matrix metalloprotease-9 | MMP-9 | NM_004994 | QT00040040 |
| Matrix metalloprotease-12 | MMP-12 | NM_002426 | QT01004472 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Longo, V.; Longo, A.; Martorana, A.; Lauria, A.; Augello, G.; Azzolina, A.; Cervello, M.; Colombo, P. Identification of an LPS-Induced Chemo-Attractive Peptide from Ciona robusta. Mar. Drugs 2020, 18, 209. https://doi.org/10.3390/md18040209
Longo V, Longo A, Martorana A, Lauria A, Augello G, Azzolina A, Cervello M, Colombo P. Identification of an LPS-Induced Chemo-Attractive Peptide from Ciona robusta. Marine Drugs. 2020; 18(4):209. https://doi.org/10.3390/md18040209
Chicago/Turabian StyleLongo, Valeria, Alessandra Longo, Annamaria Martorana, Antonino Lauria, Giuseppa Augello, Antonina Azzolina, Melchiorre Cervello, and Paolo Colombo. 2020. "Identification of an LPS-Induced Chemo-Attractive Peptide from Ciona robusta" Marine Drugs 18, no. 4: 209. https://doi.org/10.3390/md18040209