Secondary Metabolites of the Genus Didemnum: A Comprehensive Review of Chemical Diversity and Pharmacological Properties



Abstract
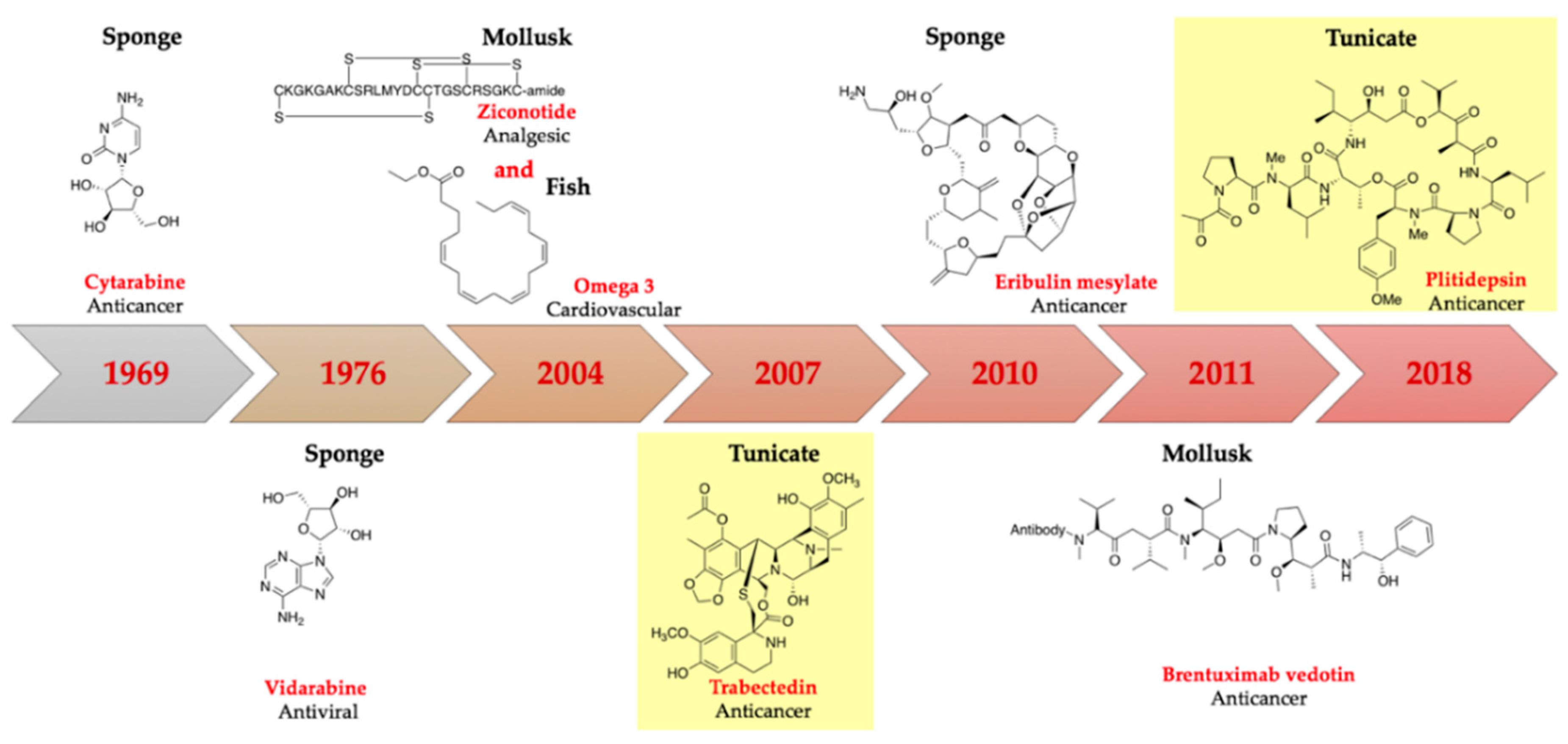
:1. Introduction
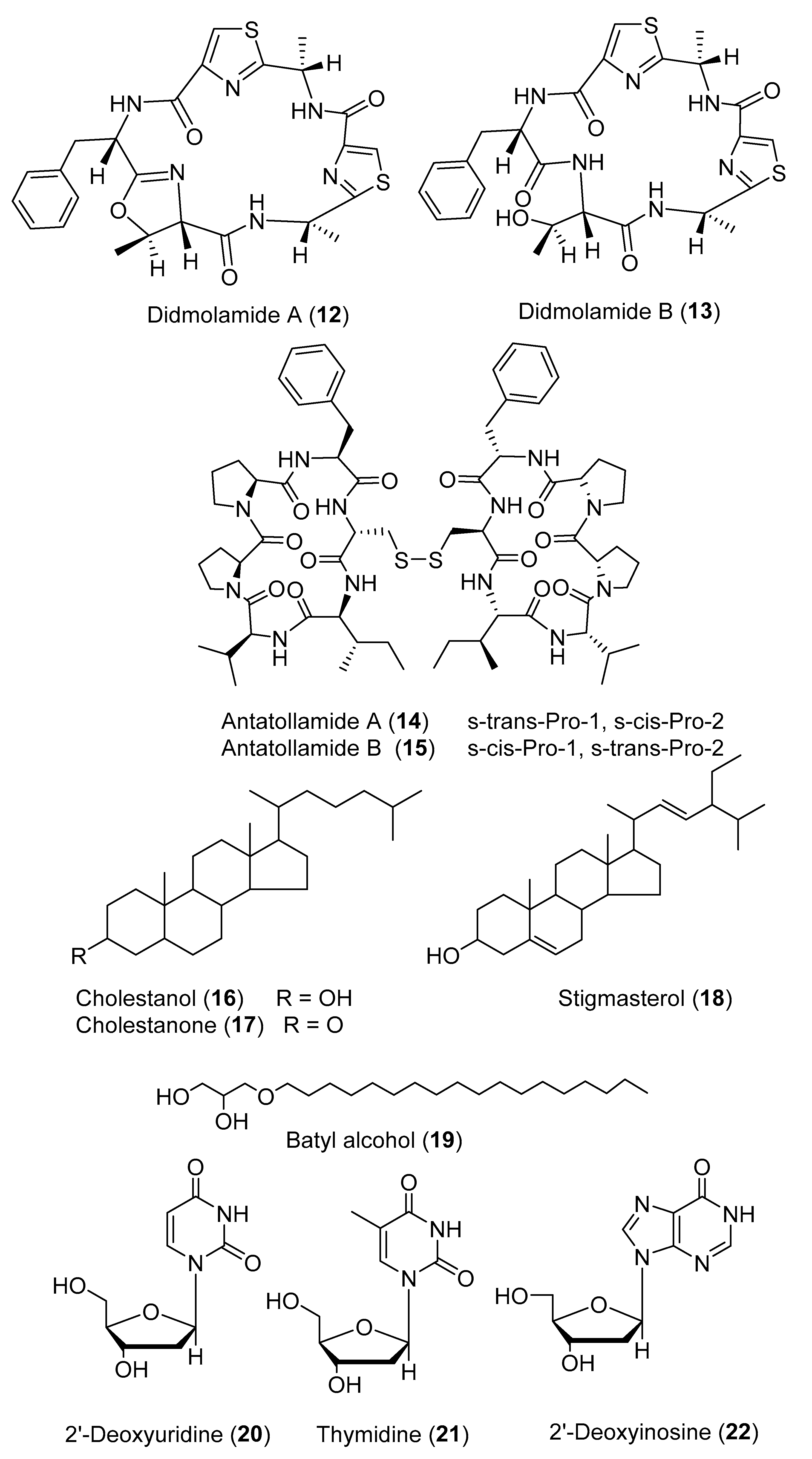
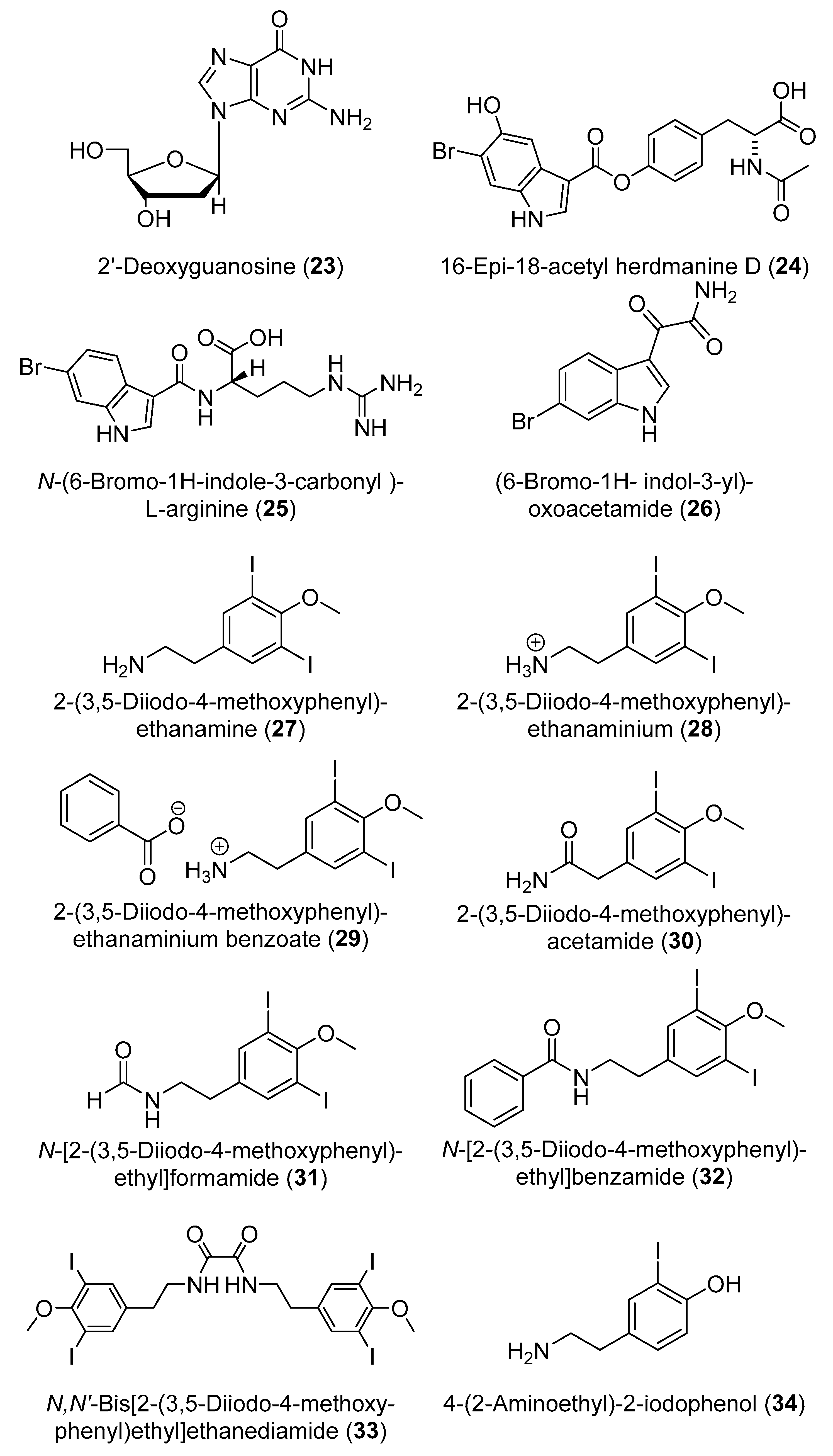
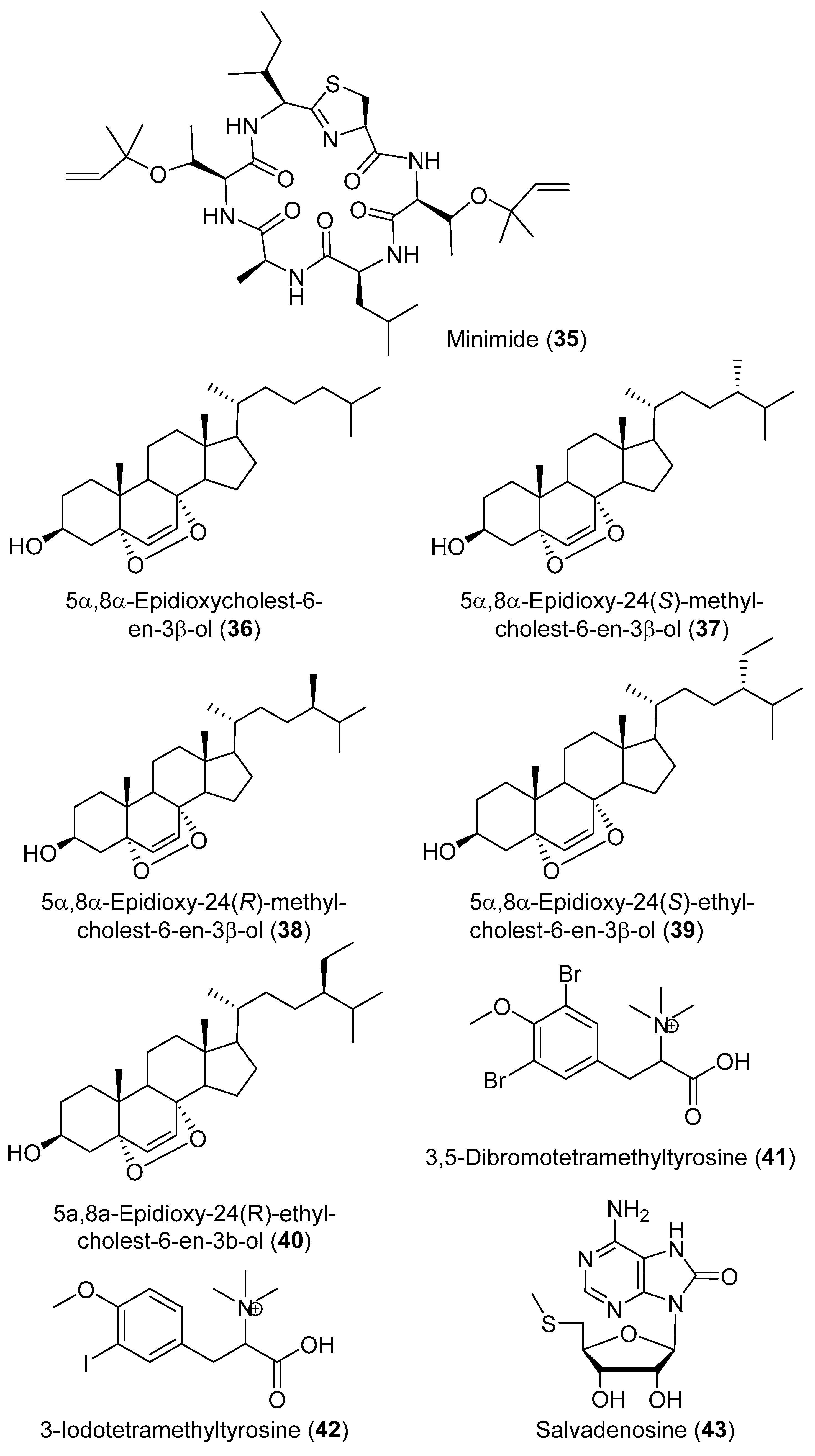
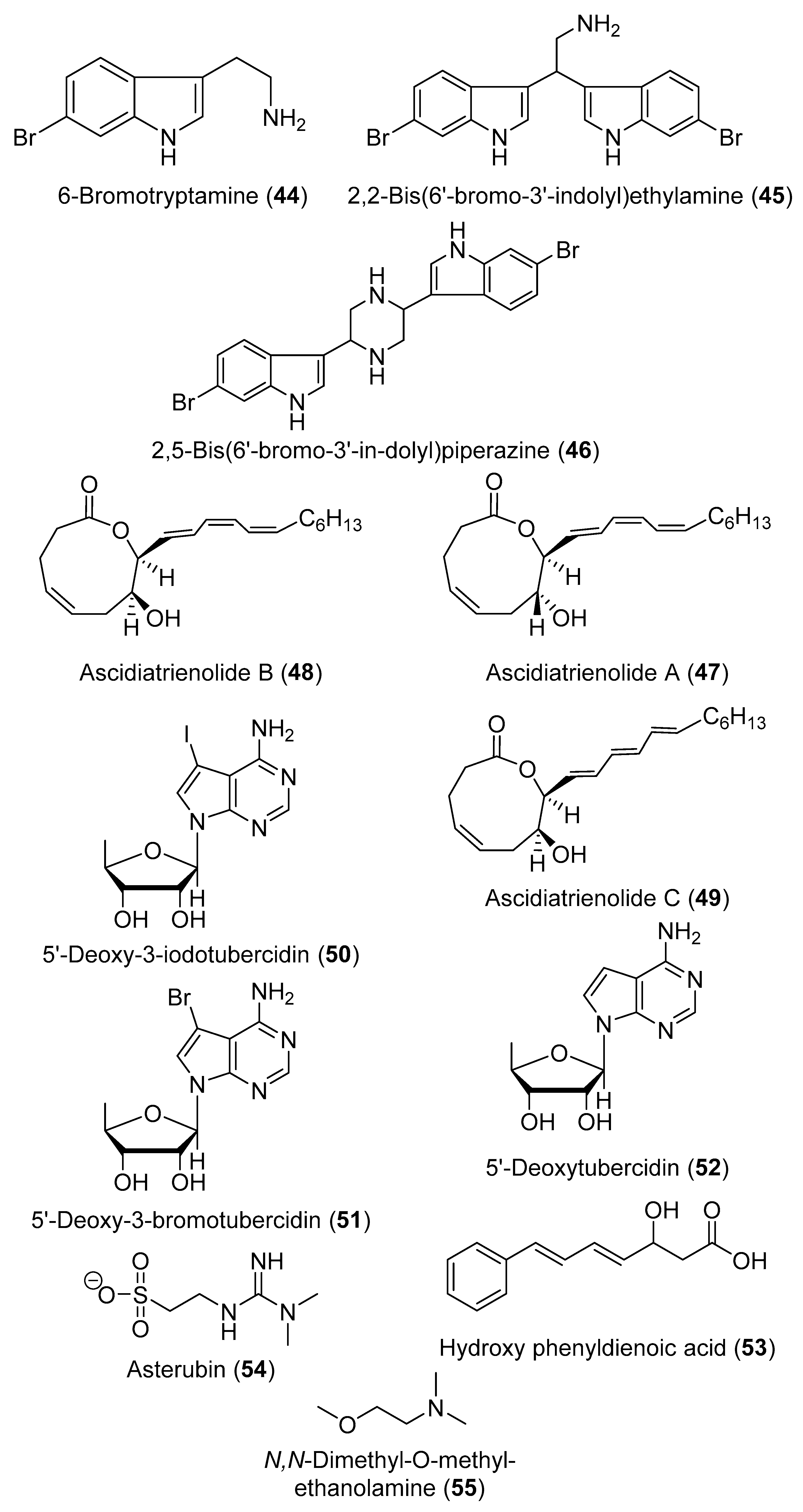
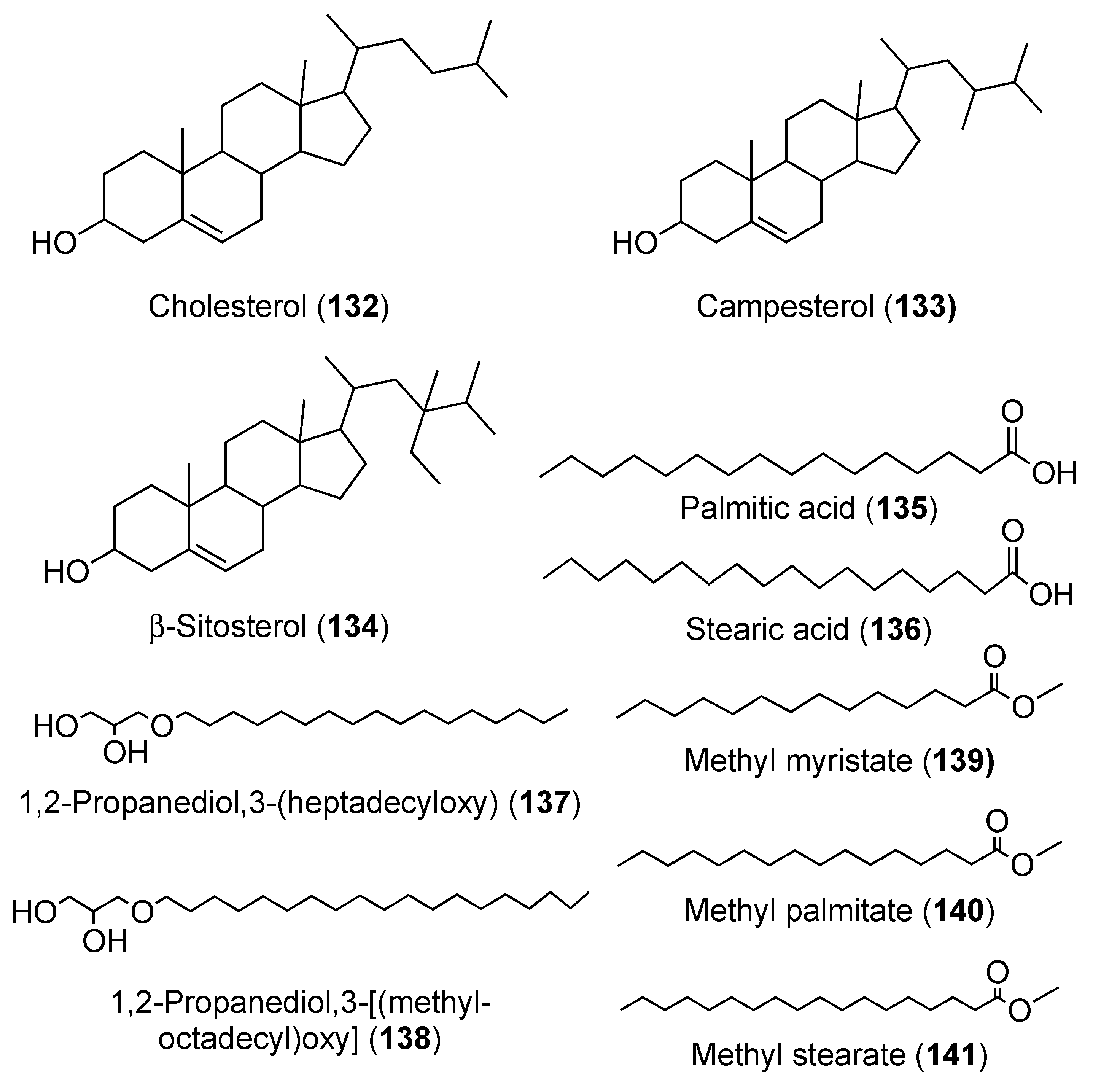
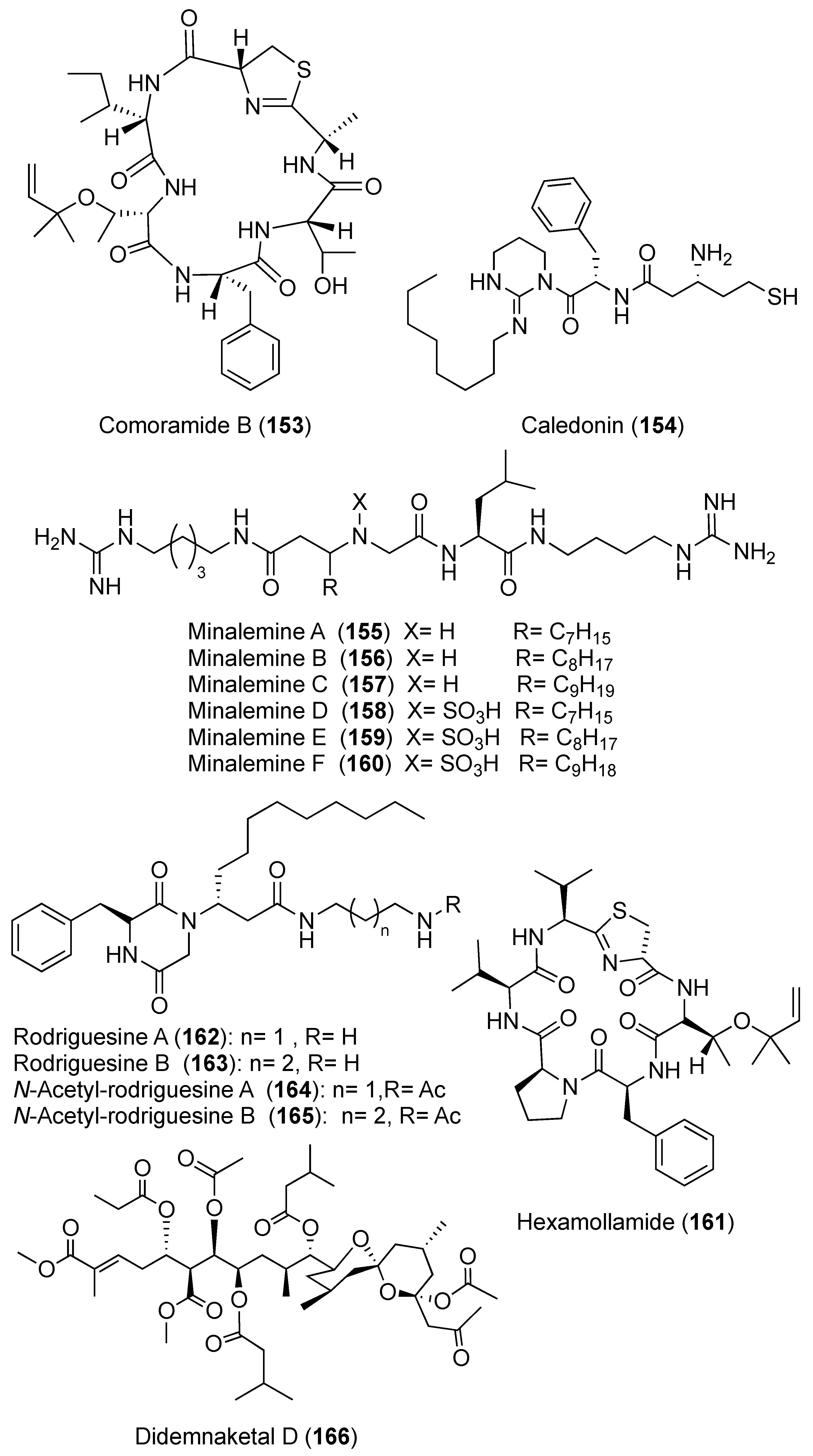
2. Secondary Metabolites with No Currently Reported Bioactivity from the Genus Didemnum (Table S6)
3. Secondary Metabolites with Reported Biological Activities
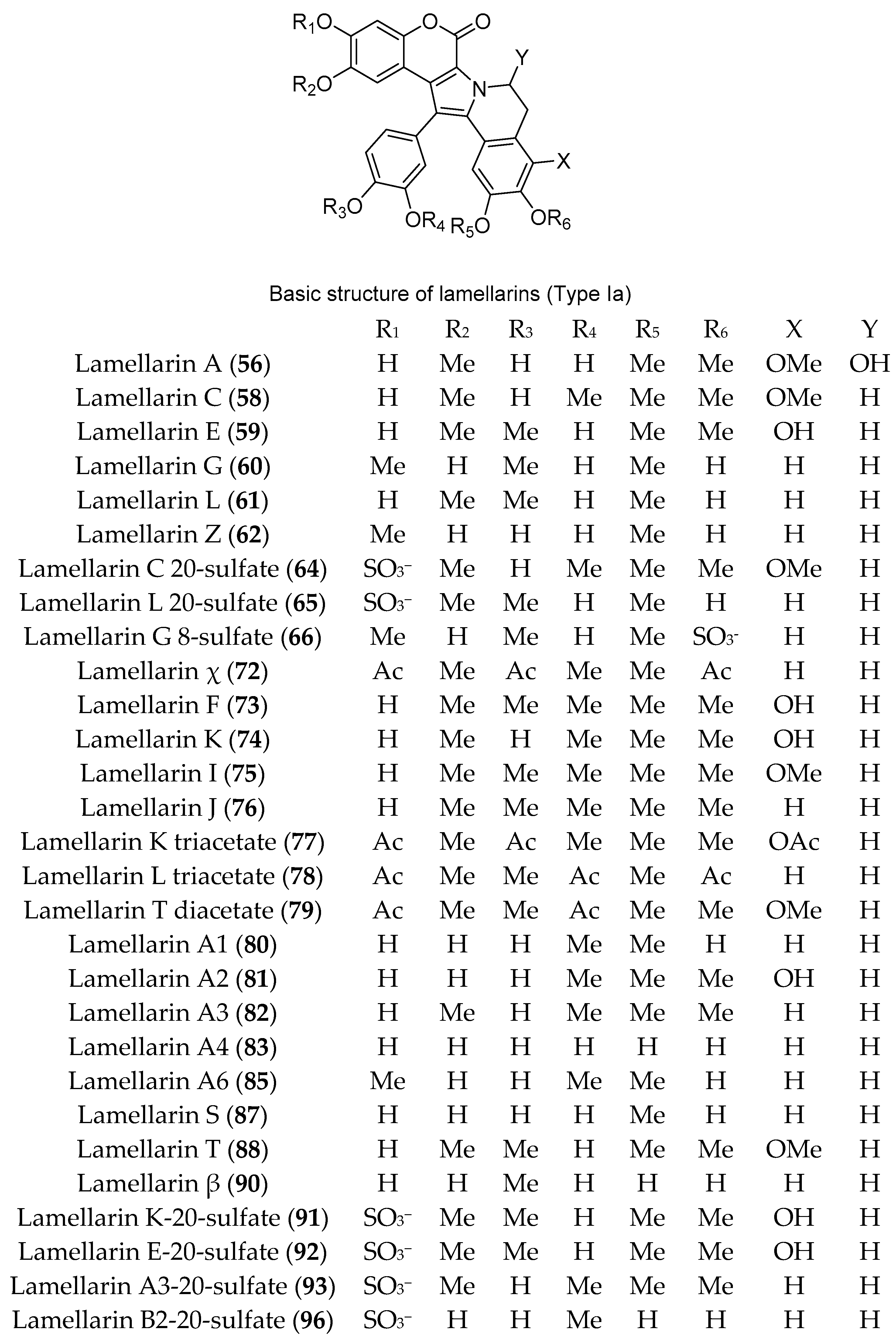
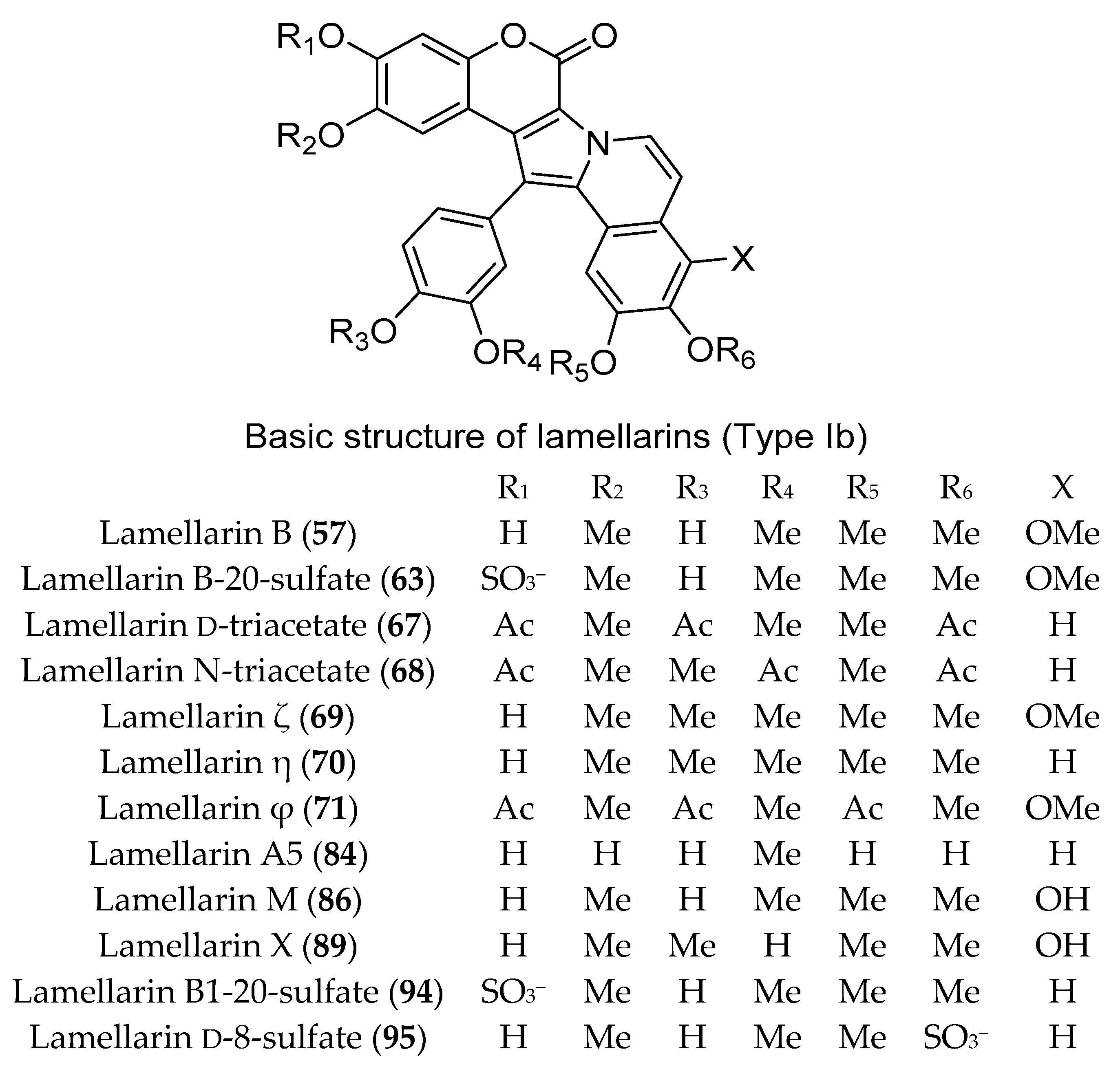
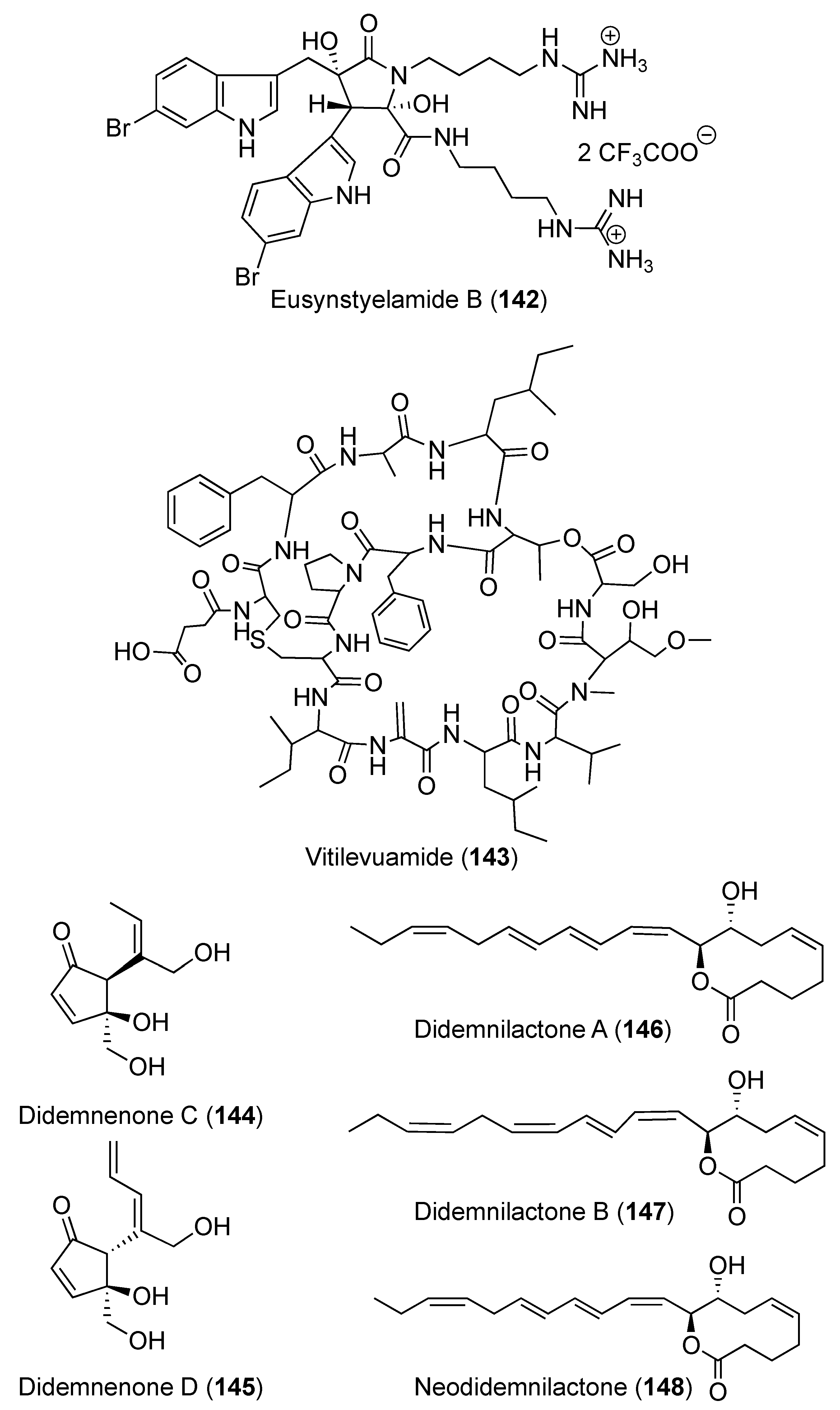
3.1. Compounds with Antitumor/Anticancer and Antiproliferative Activities (Table S1)
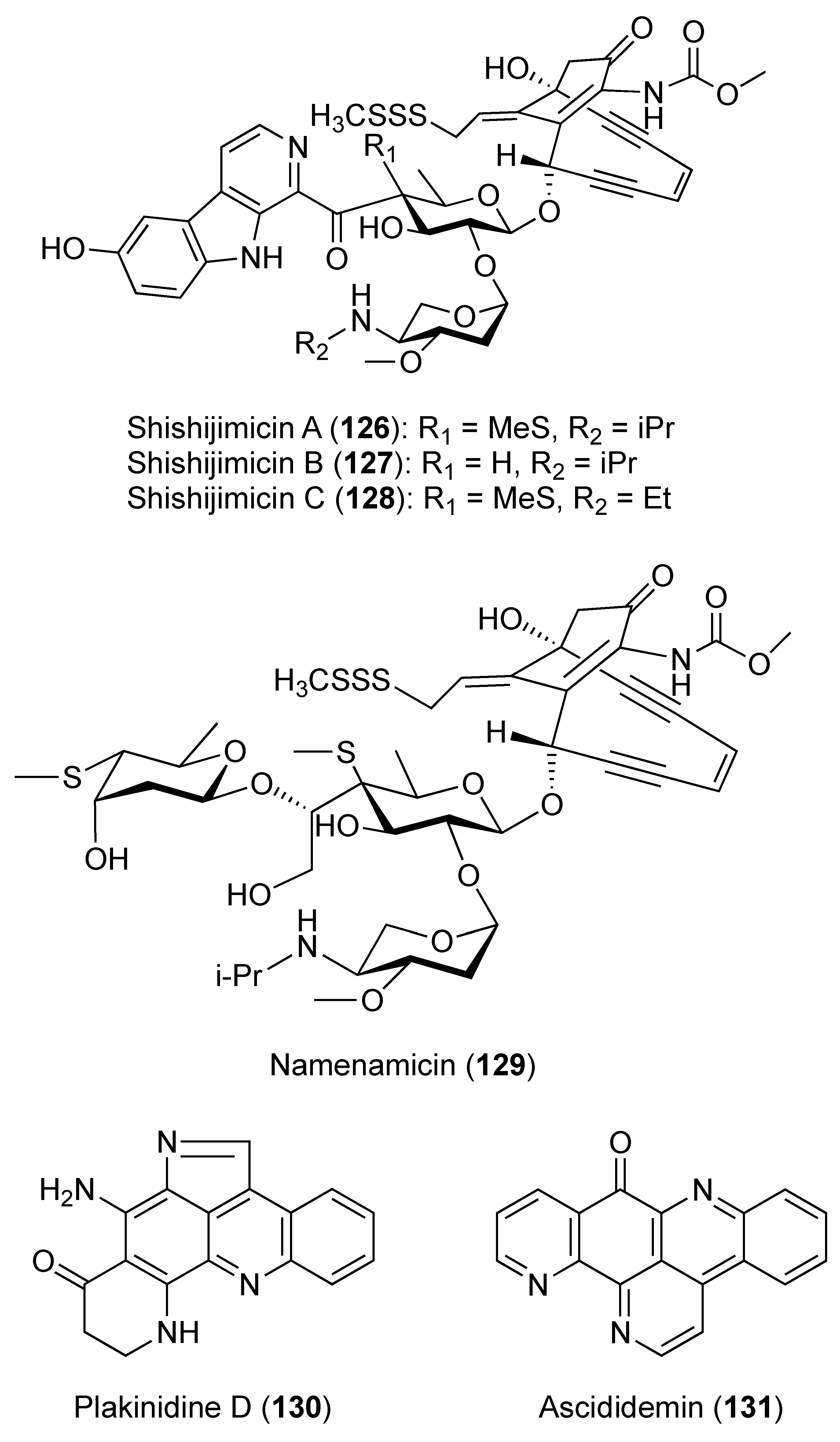
3.2. Compounds with Antimicrobial Activities (Table S2)
3.2.1. Compounds with Antibacterial Activities (Table S2)
3.2.2. Compounds with Antiviral Activities (Table S2)
3.2.3. Compounds with Antifungal Activities (Table S2)
3.3. Compounds with Antimalarial and Antitrypanosomal Activities (Table S3)
3.4. Compounds with Antidiabetic Activity (Table S4)
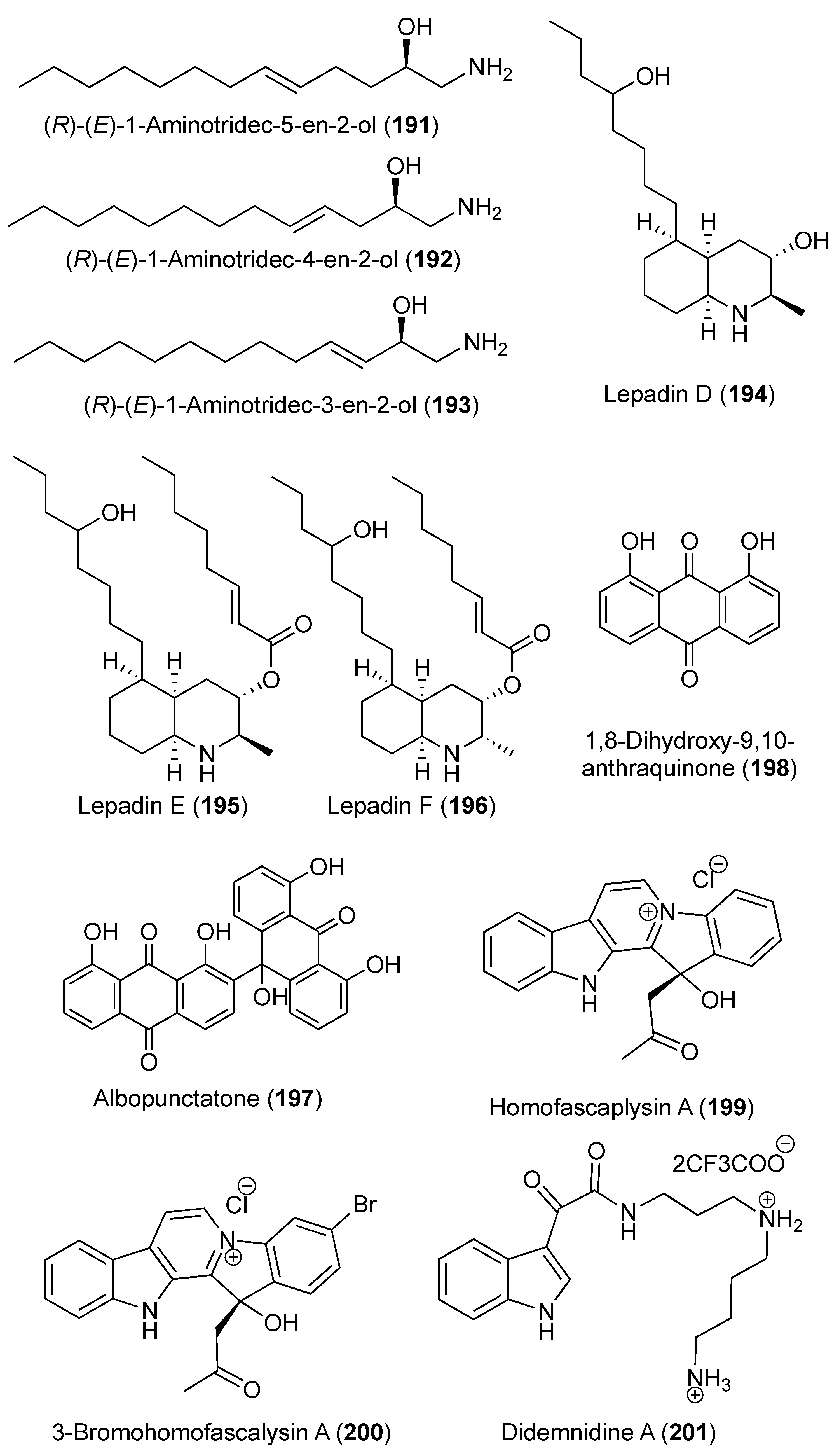
3.5. Compounds that Affect the Central Nervous System (Table S5)
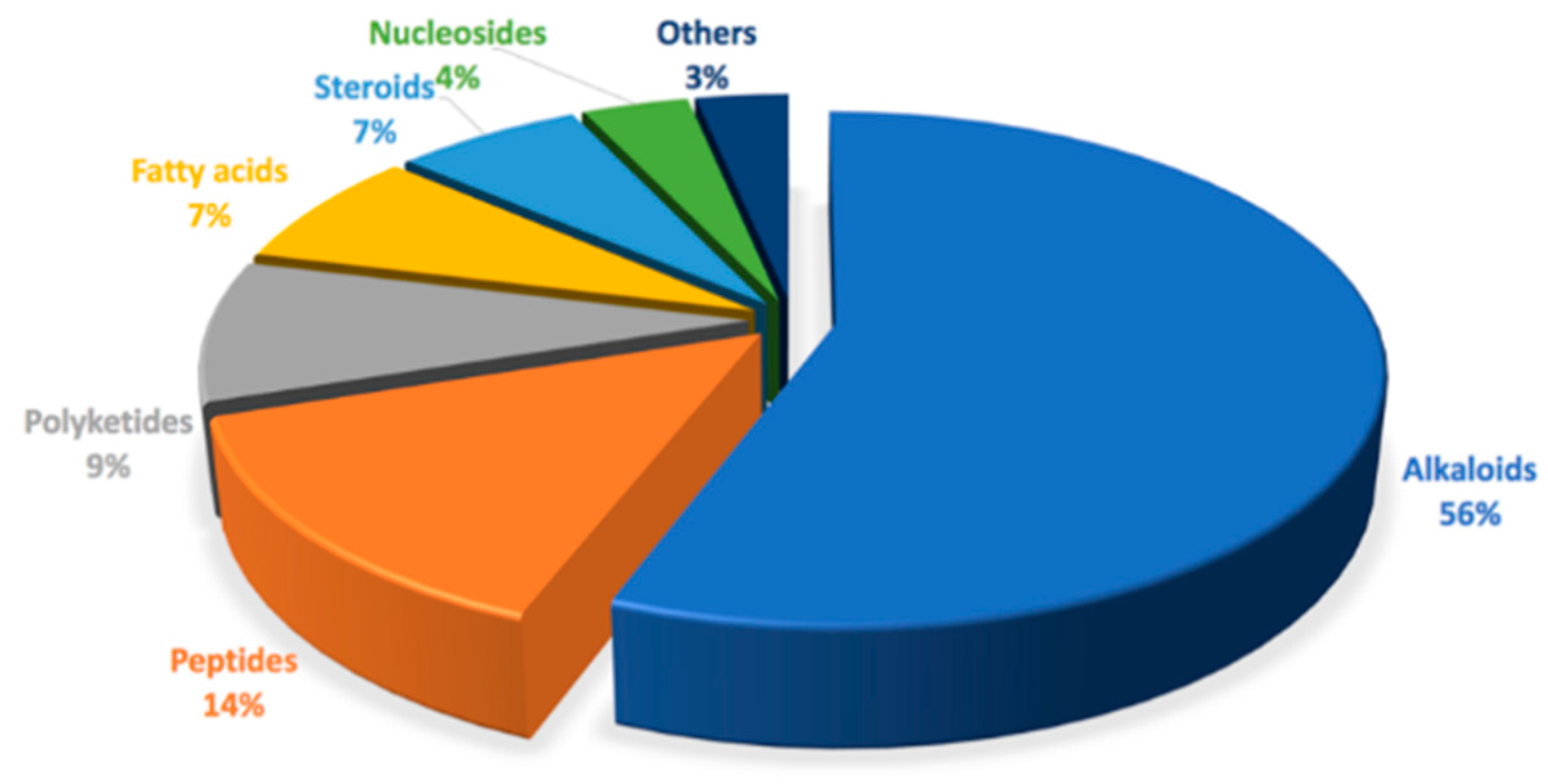
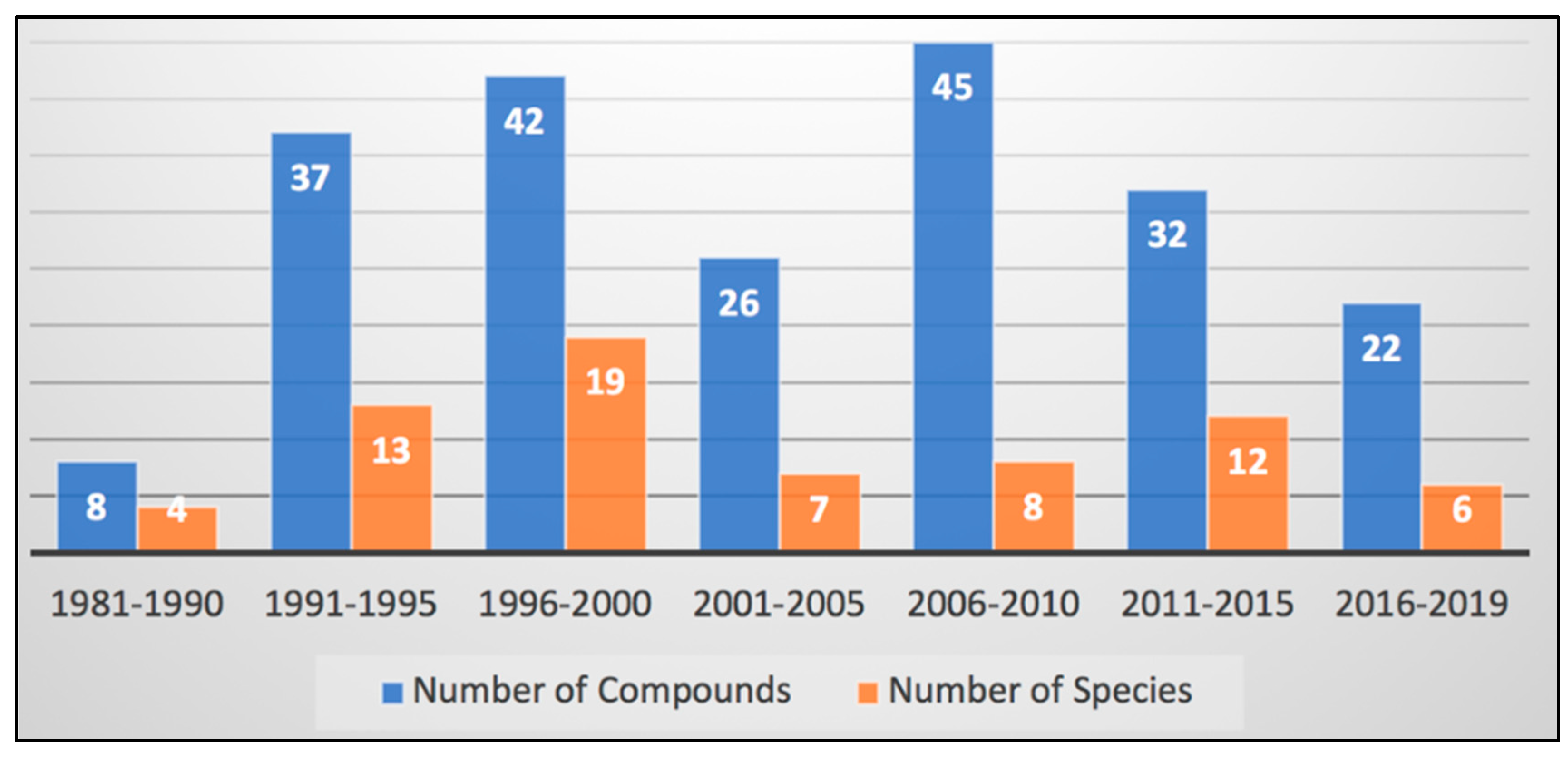
4. Discussion: The Chemistry and Chemical Potential of Didemnum
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Mayer, A.M.; Glaser, K.B.; Cuevas, C.; Jacobs, R.S.; Kem, W.; Little, R.D.; McIntosh, J.M.; Newman, D.J.; Potts, B.C.; Shuster, D.E. The odyssey of marine pharmaceuticals: A current pipeline perspective. Trends Pharmacol. Sci. 2010, 31, 255–265. [Google Scholar] [CrossRef]
- Rangel, M.; Falkenberg, M. An overview of the marine natural products in clinical trials and on the market. J. Coast. Life Med. 2015, 3, 421–428. [Google Scholar]
- Pereira, F. Have marine natural product drug discovery efforts been productive and how can we improve their efficiency? Expert Opin. Drug Dis. 2019, 14, 717–722. [Google Scholar] [CrossRef] [Green Version]
- Blunt, J.W.; Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2018, 35, 8–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2014, 31, 160–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2015, 32, 116–211. [Google Scholar] [CrossRef] [Green Version]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2017, 34, 235–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munroa, M.; Prinsepd, M.R. Marine natural products. Nat. Prod. Rep. 2016, 33, 382–431. [Google Scholar] [CrossRef] [Green Version]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2019, 36, 122–173. [Google Scholar] [CrossRef] [Green Version]
- Shenkar, N.; Swalla, B.J. Global diversity of Ascidiacea. PLoS ONE 2011, 6. [Google Scholar] [CrossRef]
- Da Silva Oliveira, F.A.; Michonneau, F.; da Cruz Lotufo, T.M. Molecular phylogeny of Didemnidae (Ascidiacea: Tunicata). Zool. J. Linn. Soc. 2017, 180, 603–612. [Google Scholar] [CrossRef]
- Hirose, M.; Nozawa, Y.; Hirose, E. Genetic isolation among morphotypes in the photosymbiotic didemnid Didemnum molle (Ascidiacea, Tunicata) from the Ryukyus and Taiwan. Zool. Sci. 2010, 27, 959–964. [Google Scholar] [CrossRef] [PubMed]
- Casso, M.; Tagliapietra, D.; Turon, X.; Pascual, M. High fusibility and chimera prevalence in an invasive colonial ascidian. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tianero, M.D.B.; Kwan, J.C.; Wyche, T.P.; Presson, A.P.; Koch, M.; Barrows, L.R.; Bugni, T.S.; Schmidt, E.W. Species specificity of symbiosis and secondary metabolism in ascidians. ISME J. 2015, 9, 615–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palanisamy, S.K.; Rajendran, N.; Marino, A. Natural products diversity of marine ascidians (tunicates; ascidiacea) and successful drugs in clinical development. Nat. Prod. Bioprospect 2017, 7, 1–111. [Google Scholar] [CrossRef] [Green Version]
- Ireland, C.M.; Durso, A.R., Jr.; Scheuer, P.J. N,N′-Diphenethylurea, a metabolite from the marine ascidian Didemnum ternatanum. J. Nat. Prod. 1981, 44, 360–361. [Google Scholar] [CrossRef]
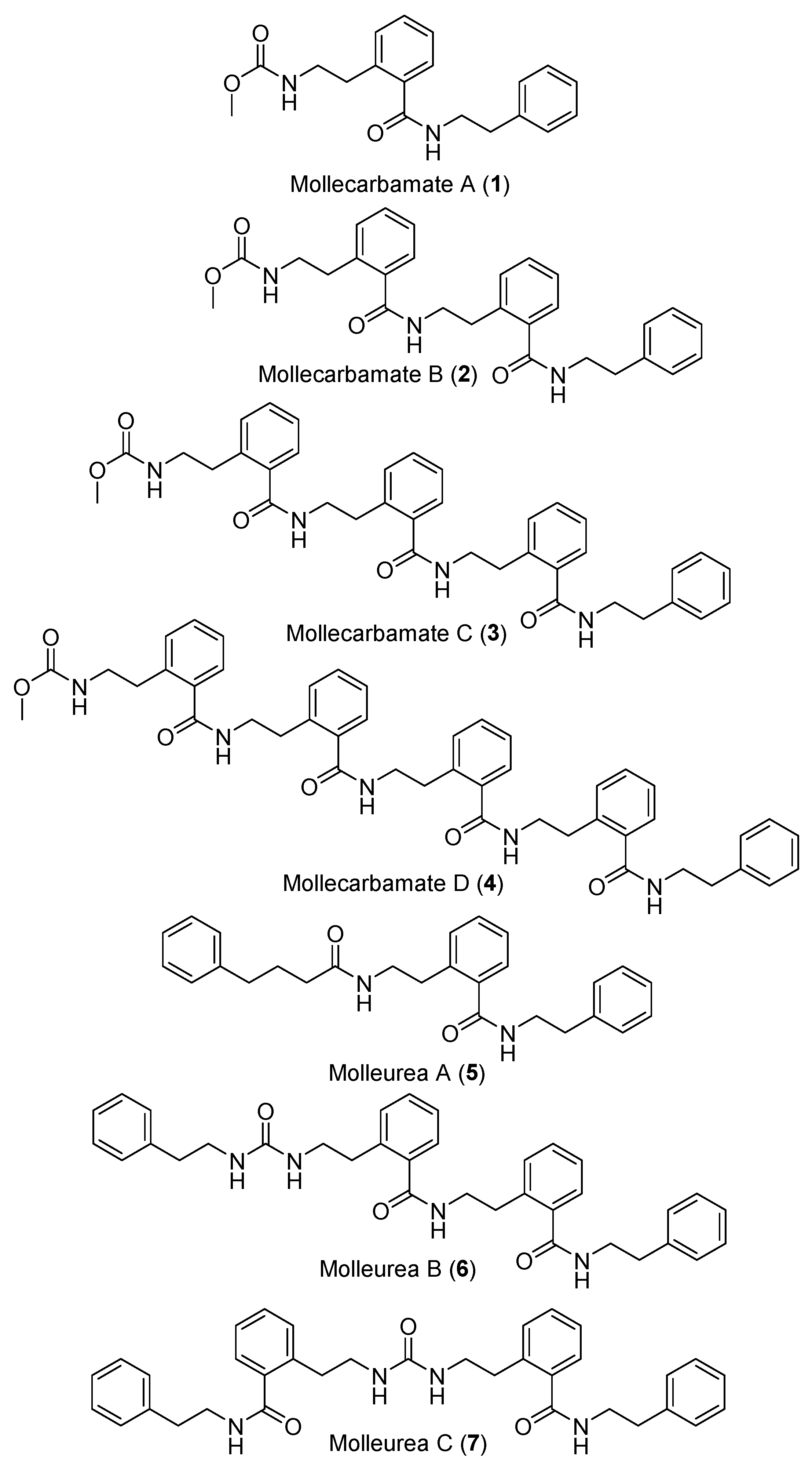
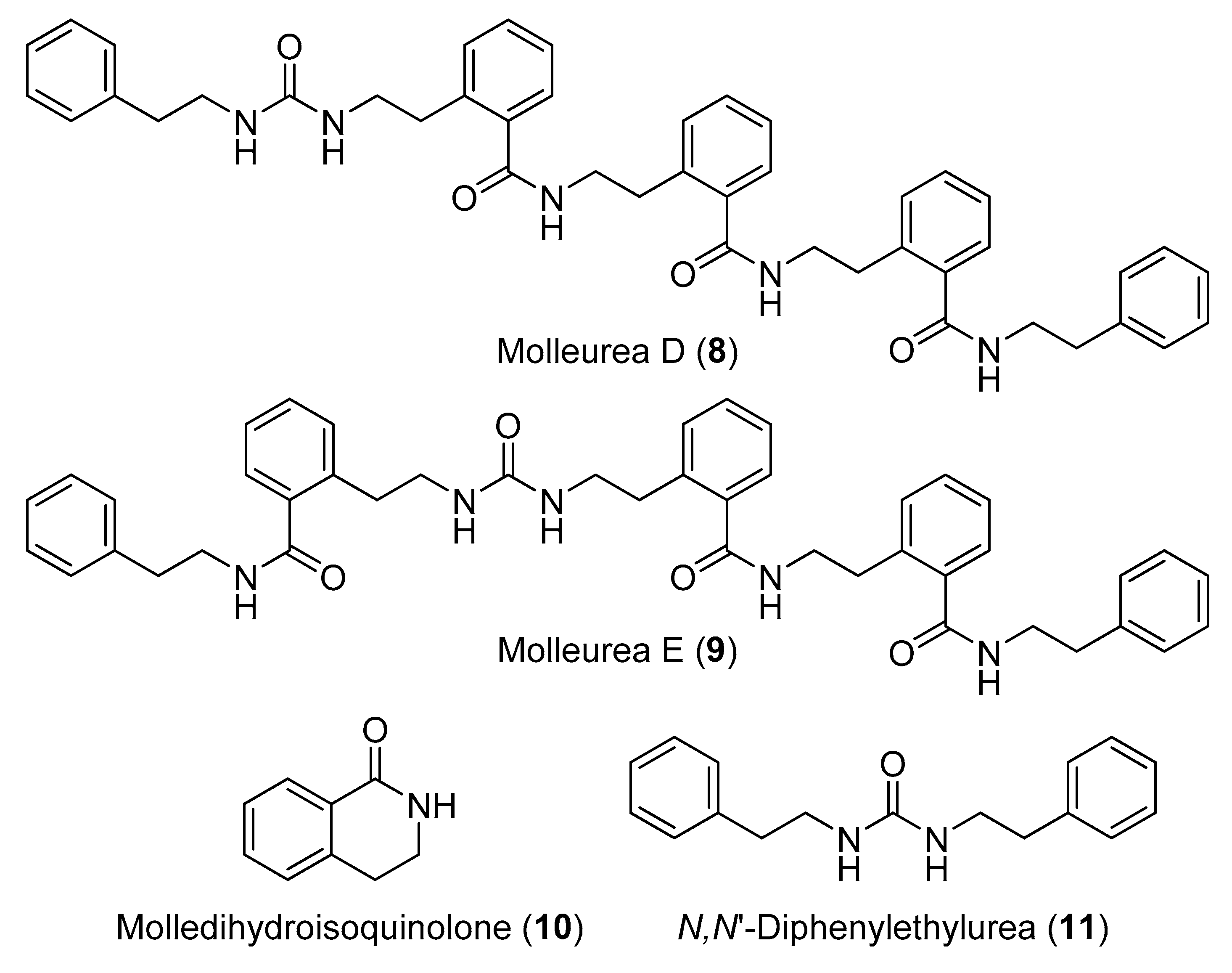
- Issac, M.; Aknin, M.; Gauvin-Bialecki, A.; Pond, C.D.; Barrows, L.R.; Kashman, Y.; Carmeli, S. Mollecarbamates, molleureas, and molledihydroisoquinolone, O-carboxyphenethylamide metabolites of the ascidian Didemnum molle collected in Madagascar. J. Nat. Prod. 2017, 80, 1844–1852. [Google Scholar] [CrossRef]
- Rudi, A.; Chill, L.; Aknin, M.; Kashman, Y. Didmolamide A and B, two new cyclic hexapeptides from the marine ascidian Didemnum molle. J. Nat. Prod. 2003, 66, 575–577. [Google Scholar] [CrossRef]
- Salib, M.N.; Molinski, T.F. Cyclic hexapeptide dimers, antatollamides A and B, from the ascidian Didemnum molle. A tryptophan-derived auxiliary for L-and D-amino acid assignments. J. Org. Chem. 2017, 82, 10181–10187. [Google Scholar] [CrossRef]
- Takeara, R.; Lopes, J.L.C.; Lopes, N.P.; Jimenez, P.C. Constituintes químicos da ascídia Didemnum psammatodes (Sluiter, 1895) coletada na costa cearense. Quím. Nova 2007, 30, 1179–1181. [Google Scholar] [CrossRef] [Green Version]
- Hahn, D.; Kim, G.J.; Choi, H.; Kang, H. A novel bromoindole alkaloid from a Korean colonial tunicate Didemnum sp. Nat. Prod. Sci. 2015, 21, 278–281. [Google Scholar] [CrossRef]
- Solano, G.; Motti, C.A.; Jaspars, M. New iodotyramine derivatives from Didemnum rubeum. Tetrahedron 2009, 65, 7482–7486. [Google Scholar] [CrossRef]
- Donia, M.S.; Ruffner, D.E.; Cao, S.; Schmidt, E.W. Accessing the hidden majority of marine natural products through metagenomics. ChemBioChem 2011, 12, 1230–1236. [Google Scholar] [CrossRef] [PubMed]
- Bensemhoun, J.; Bombarda, I.; Aknin, M.; Faure, R.; Gaydou, E.M. 5α, 8α-Epidioxysterols from the tunicate Didemnum salary. Biochem. Syst. Ecol. 2009, 36, 942–944. [Google Scholar] [CrossRef]
- Bromley, C.L.; Raab, A.; Parker-Nance, S.; Beukes, D.R.; Jaspars, M.; Davies-Coleman, M.T. Hyphenated LC-ICP-MS/ESI-MS identification of halogenated metabolites in South African marine ascidian extracts. S. Afr. J. Chem. 2018, 71, 111–117. [Google Scholar] [CrossRef]
- Jamison, M.T.; Boddy, C.N.; Molinski, T.F. Salvadenosine, a 5′-Deoxy-5′-(methylthio) nucleoside from the Bahamian tunicate Didemnum sp. J. Org. Chem. 2014, 79, 9992–9997. [Google Scholar] [CrossRef] [Green Version]
- Fahy, E.; Potts, B.C.; Faulkner, D.J.; Smith, K. 6-Bromotryptamine derivatives from the Gulf of California tunicate Didemnum candidum. J. Nat. Prod. 1991, 54, 564–569. [Google Scholar] [CrossRef]
- Lindquist, N.; Fenical, W. Ascidiatrienolids A–C, novel lactonized eicosanoids from the colonial marine ascidian Didemnum candidum. Tetrahedron Lett. 1989, 30, 2735–2738. [Google Scholar] [CrossRef]
- Mitchell, S.S.; Pomerantz, S.C.; Concepción, G.P.; Ireland, C.M. Tubercidin analogs from the ascidian Didemnum voeltzkowi. J. Nat. Prod. 1996, 59, 1000–1001. [Google Scholar] [CrossRef]
- Isaacs, S.; Kashman, Y.; Benayahu, Y. 3-Hydroxy-7-phenyl-4E,6E-heptadienoic acid from an ascidian Didemnum granulatum. J. Nat. Prod. 1994, 57, 648–649. [Google Scholar] [CrossRef]
- Kossuga, M.H.; de Lira, S.P.; Nascimento, A.M.; Gambardella, M.T.P.; Berlinck, R.G.; Torres, Y.R.; Nascimento, G.G.; Pimenta, E.F.; Silva, M.; Thiemann, O.H. Isolamento e atividades biológicas de produtos naturais das esponjas Monanchora arbuscula, Aplysina sp., Petromica ciocalyptoides e Topsentia ophiraphidites, da Ascídia Didemnum ligulum e do Octocoral Carijoa riisei. Quim. Nova 2007, 30, 1194–1202. [Google Scholar] [CrossRef]
- Davis, R.A.; Carroll, A.R.; Pierens, G.K.; Quinn, R.J. New lamellarin alkaloids from the Australian ascidian, Didemnum chartaceum. J. Nat. Prod. 1999, 62, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.M.; Srinivasulu, M.; Satyanarayana, N.; Kondapi, A.K.; Venkateswarlu, Y. New potent cytotoxic lamellarin alkaloids from Indian ascidian Didemnum obscurum. Tetrahedron 2005, 61, 9242–9247. [Google Scholar] [CrossRef]
- Plisson, F.; Huang, X.C.; Zhang, H.; Khalil, Z.; Capon, R.J. Lamellarins as inhibitors of p-glycoprotein-mediated multidrug resistance in a human colon cancer cell line. Chem. Asian J. 2012, 7, 1616–1623. [Google Scholar] [CrossRef] [PubMed]
- Carroll, A.; Bowden, B.; Coll, J. Studies of Australian ascidians. I. Six new lamellarin-class alkaloids from a colonial ascidian, Didemnum sp. Aust. J. Chem. 1993, 46, 489–501. [Google Scholar] [CrossRef]
- Urban, S.; Capon, R.J. Lamellarin-S: A new aromatic metabolite from an Australian tunicate, Didemnum sp. Aust. J. Chem. 1996, 49, 711–713. [Google Scholar] [CrossRef]
- Ham, J.-Y.; Kang, H.-J. A novel cytotoxic alkaloid of lamellarin class from a marine ascidian Didemnum sp. Bull. Korean Chem. Soc. 2002, 23, 163–166. [Google Scholar]
- Bracegirdle, J.; Robertson, L.P.; Hume, P.A.; Page, M.J.; Sharrock, A.V.; Ackerley, D.F.; Carroll, A.R.; Keyzers, R.A. Lamellarin sulfates from the Pacific tunicate Didemnum ternerratum. J. Nat. Prod. 2019, 82, 2000–2008. [Google Scholar] [CrossRef]
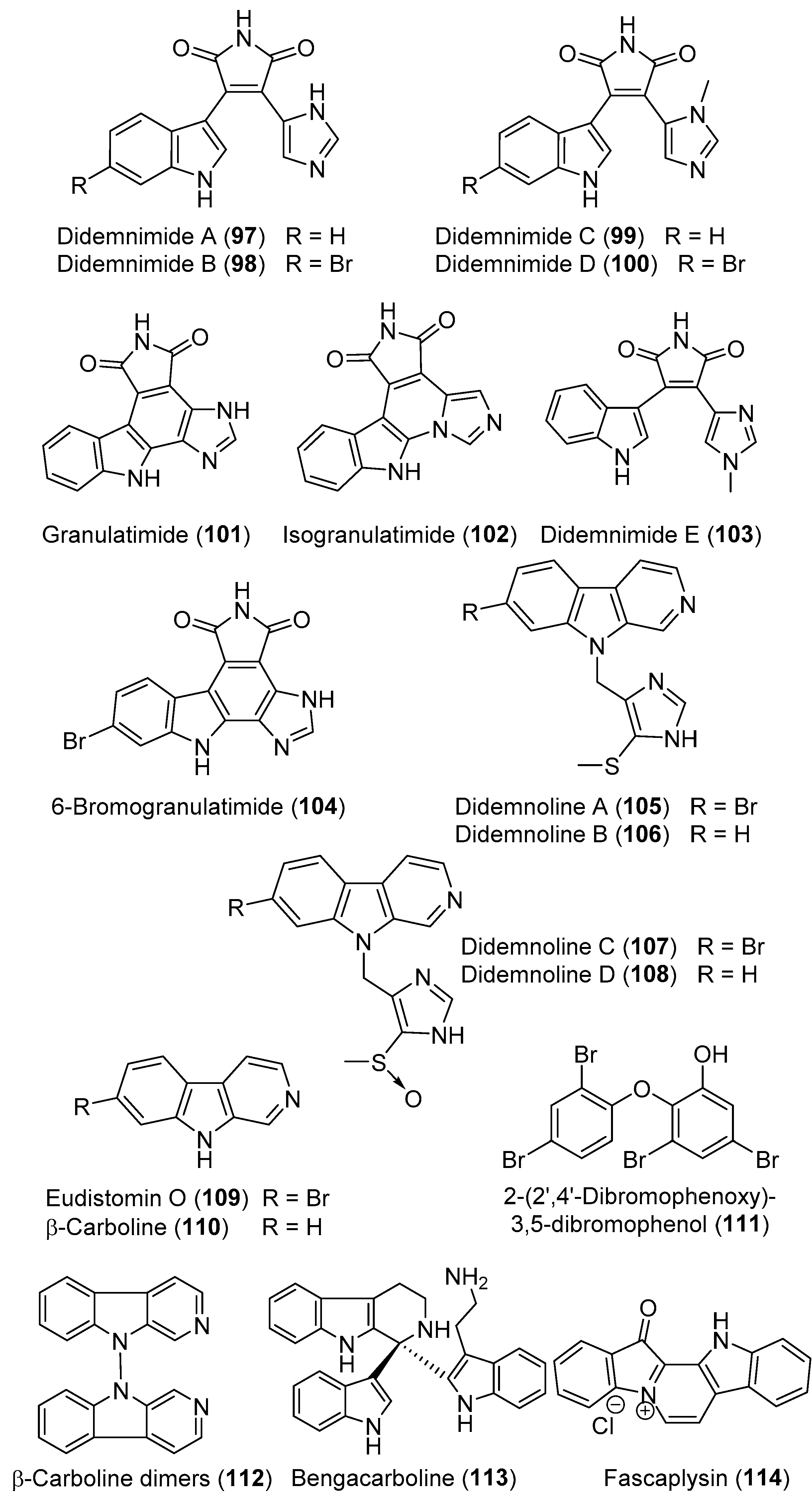
- Vervoort, H.C.; Richards-Gross, S.E.; Fenical, W.; Lee, A.Y.; Clardy, J. Didemnimides A−D: Novel, predator-deterrent alkaloids from the Caribbean mangrove ascidian Didemnum conchyliatum. J. Org. Chem. 1997, 62, 1486–1490. [Google Scholar] [CrossRef]
- Berlinck, R.G.; Britton, R.; Piers, E.; Lim, L.; Roberge, M.; Moreira da Rocha, R.; Andersen, R.J. Granulatimide and isogranulatimide, aromatic alkaloids with G2 checkpoint inhibition activity isolated from the Brazilian ascidian Didemnum granulatum: Structure elucidation and synthesis. J. Org. Chem. 1998, 63, 9850–9856. [Google Scholar] [CrossRef]
- Britton, R.; de Oliveira, J.H.; Andersen, R.J.; Berlinck, R.G. Granulatimide and 6-bromogranulatimide, minor alkaloids of the Brazilian ascidian Didemnum granulatum. J. Nat. Prod. 2001, 64, 254–255. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, R.W.; Davidson, B.S. Didemnolines A–D, new N9-substituted β-carbolines from the marine ascidian Didemnum sp. Tetrahedron 1995, 51, 10125–10130. [Google Scholar] [CrossRef]
- Kearns, P.S.; Coll, J.C.; Rideout, J.A. A β-carboline dimer from an ascidian, Didemnum sp. J. Nat. Prod. 1995, 58, 1075–1076. [Google Scholar] [CrossRef]
- Foderaro, T.A.; Barrows, L.R.; Lassota, P.; Ireland, C.M. Bengacarboline, a new β-carboline from a marine ascidian Didemnum sp. J. Org. Chem. 1997, 62, 6064–6065. [Google Scholar] [CrossRef]
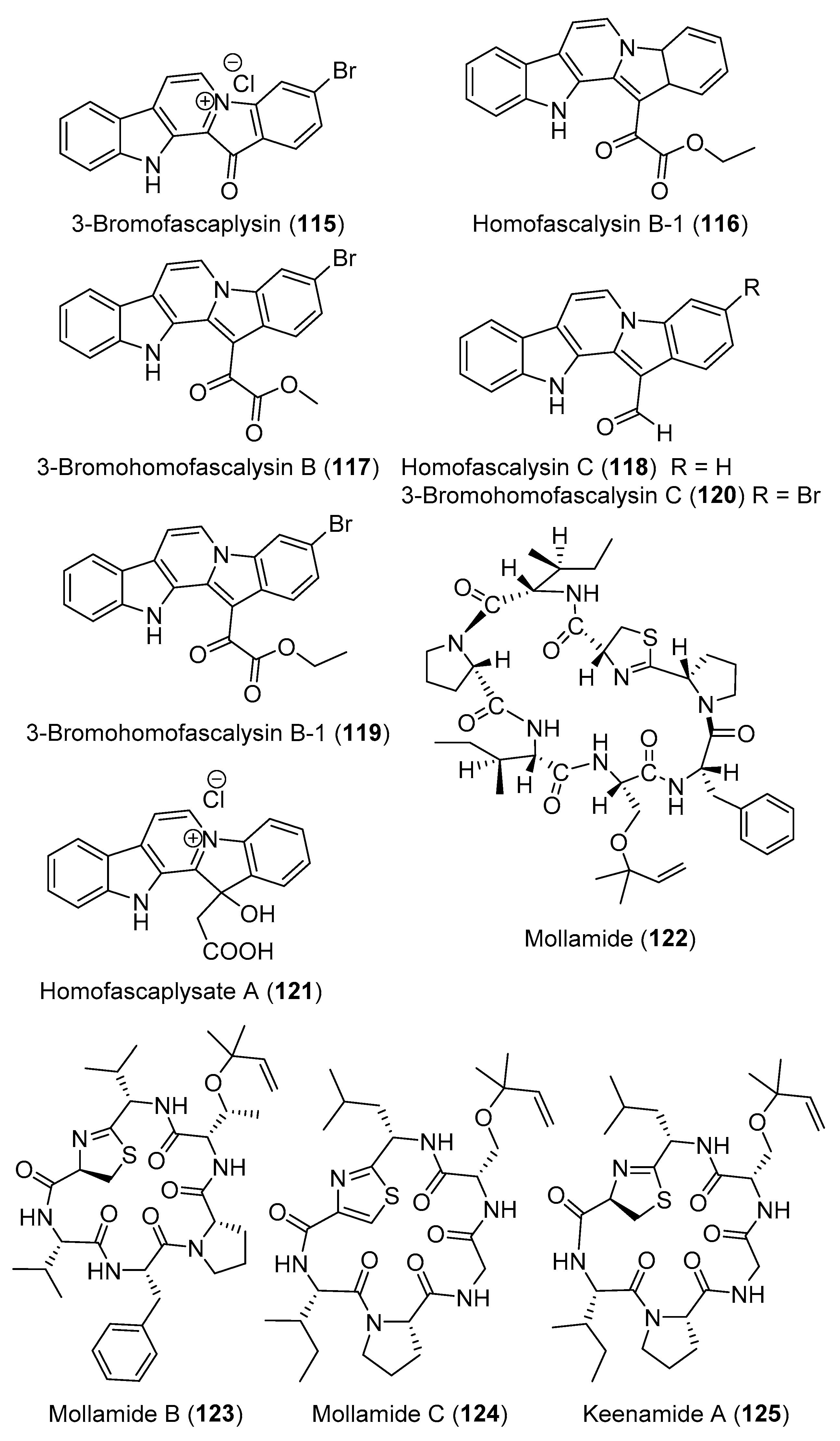
- Segraves, N.L.; Robinson, S.J.; Garcia, D.; Said, S.A.; Fu, X.; Schmitz, F.J.; Pietraszkiewicz, H.; Valeriote, F.A.; Crews, P. Comparison of fascaplysin and related alkaloids: A study of structures, cytotoxicities, and sources. J. Nat. Prod. 2004, 67, 783–792. [Google Scholar] [CrossRef]
- Carroll, A.R.; Bowden, B.F.; Coll, J.C.; Hockless, D.C.; Skelton, B.W.; White, A.H. Studies of Australian ascidians. IV. Mollamide, a cytotoxic cyclic heptapeptide from the compound ascidian Didemnum molle. Aust. J. Chem. 1994, 47, 61–69. [Google Scholar] [CrossRef]
- Donia, M.S.; Wang, B.; Dunbar, D.C.; Desai, P.V.; Patny, A.; Avery, M.; Hamann, M.T. Mollamides B and C, cyclic hexapeptides from the Indonesian tunicate Didemnum molle. J. Nat. Prod. 2008, 71, 941–945. [Google Scholar] [CrossRef] [Green Version]
- Oku, N.; Matsunaga, S.; Fusetani, N. Shishijimicins A−C, novel enediyne antitumor antibiotics from the ascidian Didemnum proliferum. J. Am. Chem. Soc. 2003, 125, 2044–2045. [Google Scholar] [CrossRef]
- Ford, P.W.; Davidson, B.S. Plakinidine D, a new pyrroloacridine alkaloid from the ascidian Didemnum rubeum. J. Nat. Prod. 1997, 60, 1051–1053. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.J.; Venables, D.A.; Hopmann, C.; Salomon, C.E.; Jompa, J.; Tahir, A.; Faulkner, D.J.; Ireland, C.M. Plakinidine D, a new pyrroloacridine alkaloid from two ascidians of the genus Didemnum. J. Nat. Prod. 1997, 60, 1048–1050. [Google Scholar] [CrossRef]
- Kobayash, J.I.; Cheng, J.-F.; Nakamura, H.; Ohizumi, Y.; Hirata, Y.; Sasaki, T.; Ohta, T.; Nozoe, S. Ascididemin, a novel pentacyclic aromatic alkaloid with potent antileukemic activity from the Okinawan tunicate Didemnum sp. Tetrahedron Lett. 1988, 29, 1177–1180. [Google Scholar] [CrossRef]
- Sesin, D.F.; Ireland, C.M. Iodinated phenethylamine products from a didemnid tunicate. Tetrahedron Lett. 1984, 25, 403–404. [Google Scholar] [CrossRef]
- Takeara, R.; Jimenez, P.C.; Wilke, D.V.; de Moraes, M.O.; Pessoa, C.; Lopes, N.P.; Lopes, J.L.C.; da Cruz Lotufo, T.M.; Costa-Lotufo, L.V. Antileukemic effects of Didemnum psammatodes (Tunicata: Ascidiacea) constituents. Comp. Biochem. Physiol. A 2008, 151, 363–369. [Google Scholar] [CrossRef]
- Liberio, M.; Sadowski, M.; Nelson, C.; Davis, R. Identification of eusynstyelamide B as a potent cell cycle inhibitor following the generation and screening of an ascidian-derived extract library using a real time cell analyzer. Mar. Drugs 2014, 12, 5222–5239. [Google Scholar] [CrossRef] [Green Version]
- Ireland, C.M.; Fernandez, A. Cyclic Peptide Antitumor Agent from an Ascidian. Ph.D. Thesis, University of Utah, Salt Lake City, UT, USA, 1998. [Google Scholar]
- Lindquist, N.; Fenical, W.; Sesin, D.F.; Ireland, C.M.; Van Duyne, G.D.; Forsyth, C.J.; Clardy, J. Isolation and structure determination of the didemnenones, novel cytotoxic metabolites from tunicates. J. Am. Chem. Soc. 1988, 110, 1308–1309. [Google Scholar] [CrossRef]
- Niwa, H.; Watanabe, M.; Inagaki, H.; Yamada, K. Didemnilactones A and B and neodidemnilactone three new fatty acid metabolites isolated from the tunicate Didemnum moseleyi (Herdman). Tetrahedron 1994, 50, 7385–7400. [Google Scholar] [CrossRef]
- Toske, S.G.; Fenical, W. Cyclodidemnamide: A new cyclic heptapeptide from the marine ascidian Didemnum molle. Tetrahedron Lett. 1995, 36, 8355–8358. [Google Scholar] [CrossRef]
- Rudi, A.; Aknin, M.; Gaydou, E.M.; Kashman, Y. Four new cytotoxic cyclic hexa-and heptapeptides from the marine ascidian Didemnum molle. Tetrahedron 1998, 54, 13203–13210. [Google Scholar] [CrossRef]
- Vázquez, M.J.; Quiñoá, E.; Riguera, R.; Ocampo, A.; Iglesias, T.; Debitus, C. Caledonin, a natural peptide bolaphile with ZnII and CuI complexing properties from the tunicate Didemnun rodriguesi. Tetrahedron Lett. 1995, 36, 8853–8856. [Google Scholar] [CrossRef]
- Exposito, M.; López, B.; Fernández, R.; Vazquez, M.; Debitus, C.; Iglesias, T.; Jiménez, C.; Quinoa, E.; Riguera, R. Minalemines AF: Sulfamic acid peptide guanidine derivatives isolated from the marine tunicate Didemnun rodriguesi. Tetrahedron 1998, 54, 7539–7550. [Google Scholar] [CrossRef]
- Teruya, T.; Sasaki, H.; Suenaga, K. Hexamollamide, a hexapeptide from an Okinawan ascidian Didemnum molle. Tetrahedron Lett. 2008, 49, 5297–5299. [Google Scholar] [CrossRef]
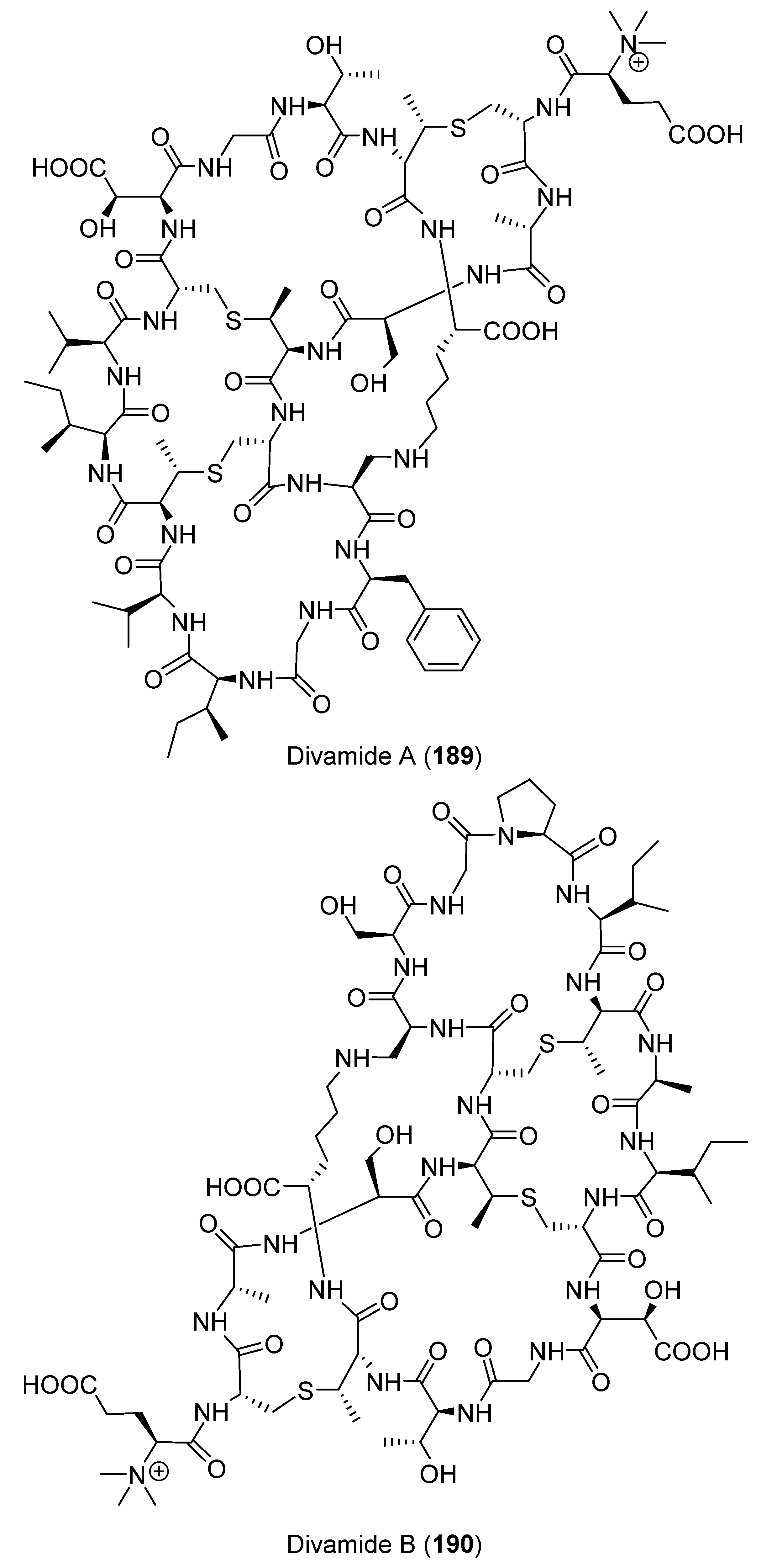
- Kossuga, M.H.; Lira, S.P.; McHugh, S.; Torres, Y.R.; Lima, B.A.; Gonçalves, R.; Veloso, K.; Ferreira, A.G.; Rocha, R.M.; Berlinck, R.G. Antibacterial modified diketopiperazines from two ascidians of the genus Didemnum. J. Braz. Chem. Soc. 2009, 20, 704–711. [Google Scholar] [CrossRef]
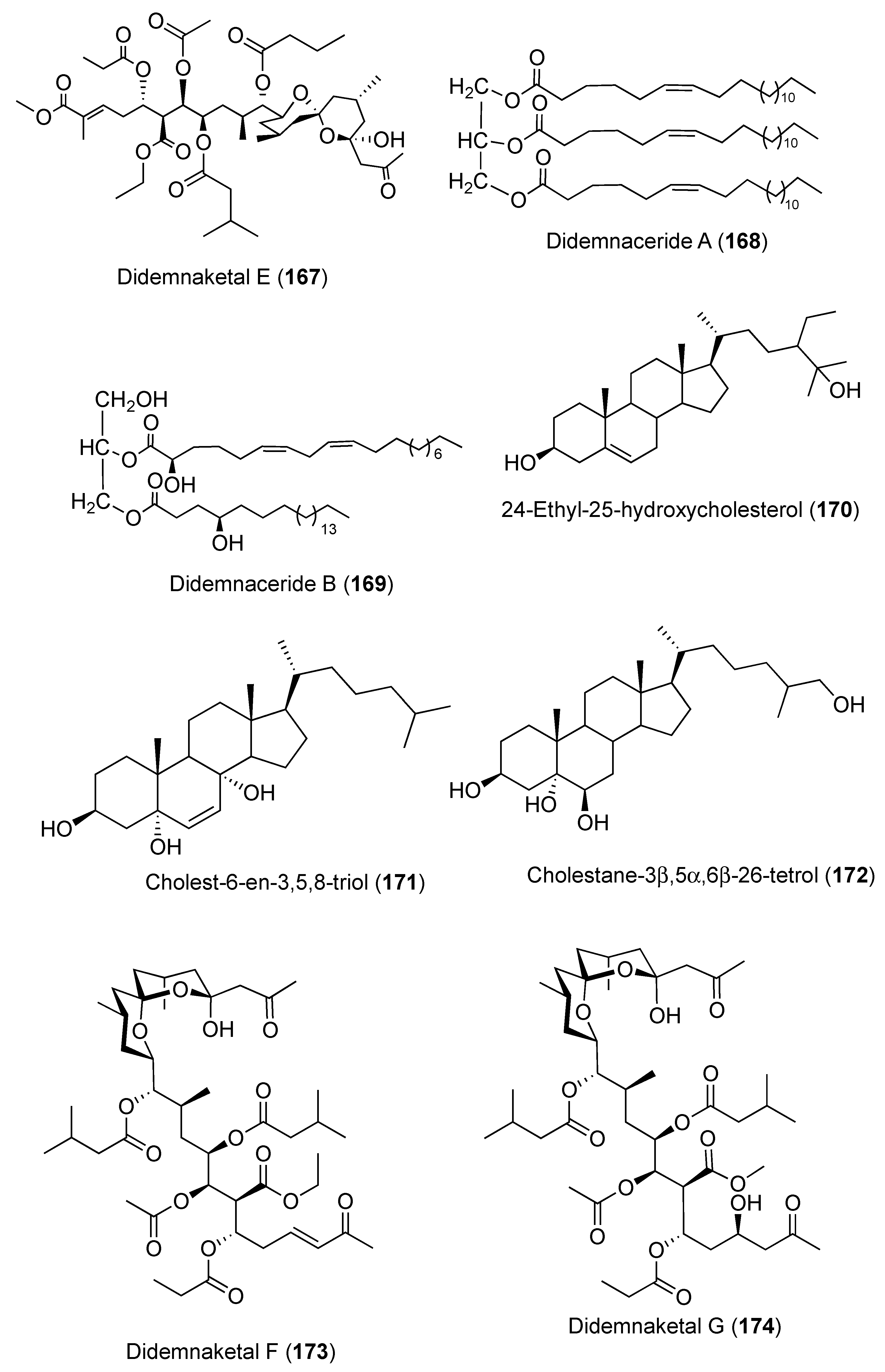
- Mohamed, G.A.; Ibrahim, S.R.; Badr, J.M.; Youssef, D.T. Didemnaketals D and E, bioactive terpenoids from a Red Sea ascidian Didemnum species. Tetrahedron 2014, 70, 35–40. [Google Scholar] [CrossRef]
- Ibrahim, S.R.; Mohamed, G.A.; Shaala, L.A.; Youssef, D.T.; Gab-Alla, A.A. Didemnacerides A and B: Two new glycerides from Red Sea ascidian Didemnum species. Nat. Prod. Res. 2014, 28, 1591–1597. [Google Scholar] [CrossRef] [PubMed]
- Shaala, L.; Youssef, D.; Ibrahim, S.; Mohamed, G.; Badr, J.; Risinger, A.; Mooberry, S. Didemnaketals F and G, new bioactive spiroketals from a Red Sea ascidian Didemnum species. Mar. Drugs 2014, 12, 5021–5034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
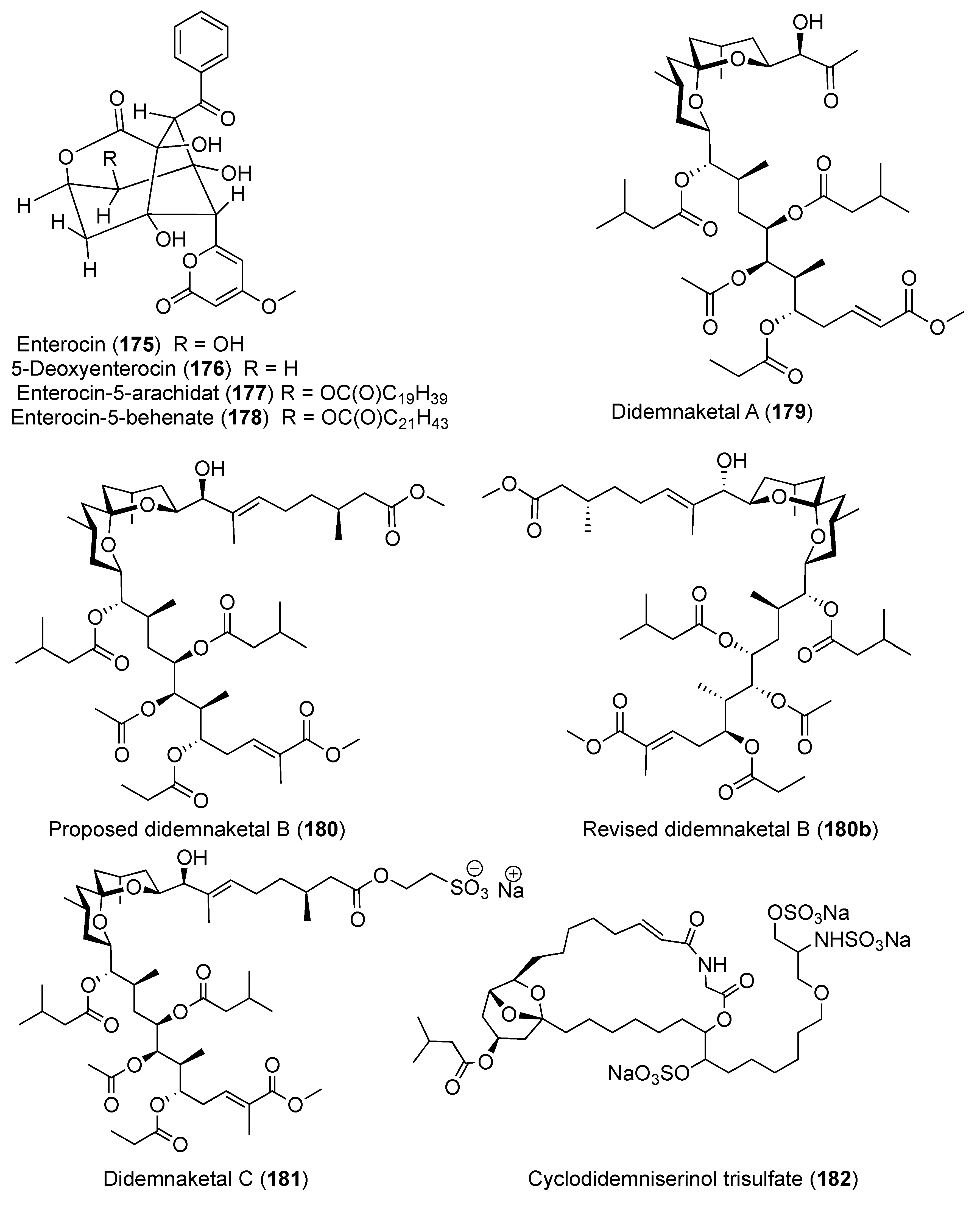
- Kang, H.; Jensen, P.R.; Fenical, W. Isolation of microbial antibiotics from a marine ascidian of the genus Didemnum. J. Org. Chem. 1996, 61, 1543–1546. [Google Scholar] [CrossRef]
- Pika, J.; Faulkner, D.J. A reinvestigation of the didemnaketals from the Palauan ascidian Didemnum sp. Nat. Prod. Lett. 1995, 7, 291–296. [Google Scholar] [CrossRef]
- Potts, B.C.; Faulkner, D.J.; Chan, J.A.; Simolike, G.C.; Offen, P.; Hemling, M.E.; Francis, T.A. Didemnaketals A and B, HIV-1 protease inhibitors from the ascidian Didemnum sp. J. Am. Chem. Soc. 1991, 113, 6321–6322. [Google Scholar] [CrossRef]
- Fuwa, H.; Muto, T.; Sekine, K.; Sasaki, M. Total synthesis and structure revision of didemnaketal B. Chem. Eur. J. 2014, 20, 1848–1860. [Google Scholar] [CrossRef] [PubMed]
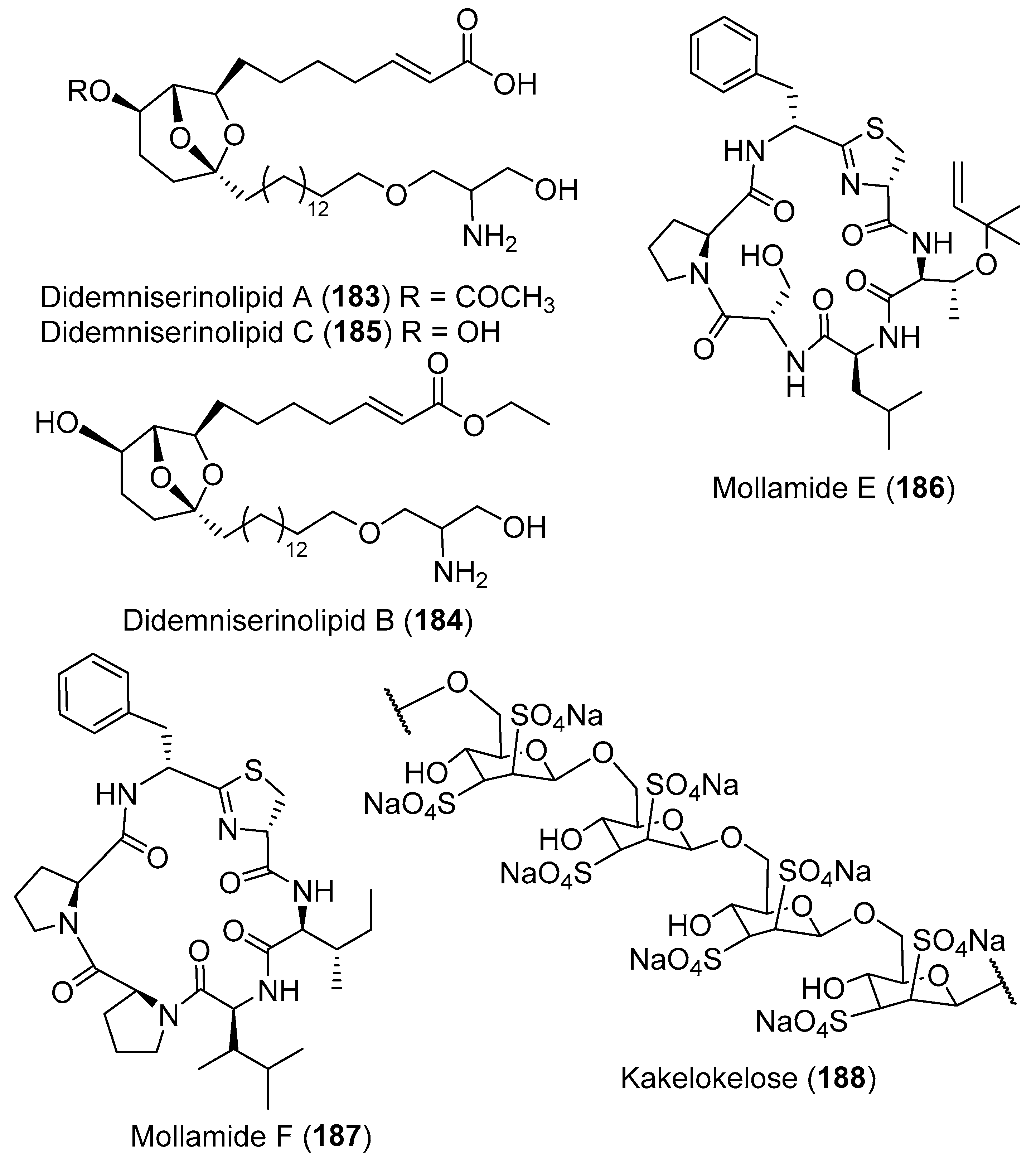
- Gonzalez, N.; Rodriguez, J.; Jiménez, C. Didemniserinolipids A−C, unprecedented serinolipids from the tunicate Didemnum sp. J. Org. Chem. 1999, 64, 5705–5707. [Google Scholar] [CrossRef]
- Mitchell, S.S.; Rhodes, D.; Bushman, F.D.; Faulkner, D.J. Cyclodidemniserinol trisulfate, a sulfated serinolipid from the Palauan ascidian Didemnum guttatum that inhibits HIV-1 integrase. Org. Lett. 2000, 2, 1605–1607. [Google Scholar] [CrossRef]
- Lu, Z.; Harper, M.K.; Pond, C.D.; Barrows, L.R.; Ireland, C.M.; Van Wagoner, R.M. Thiazoline peptides and a tris-phenethyl urea from Didemnum molle with anti-HIV activity. J. Nat. Prod. 2012, 75, 1436–1440. [Google Scholar] [CrossRef] [Green Version]
- Riccio, R.; Kinnel, R.B.; Bifulco, G.; Scheuer, P.J. Kakelokelose, a sulfated mannose polysaccharide with anti-HIV activity from the Pacific tunicate Didemnum molle. Tetrahedron Lett. 1996, 37, 1979–1982. [Google Scholar] [CrossRef]
- Smith, T.E.; Pond, C.D.; Pierce, E.; Harmer, Z.P.; Kwan, J.; Zachariah, M.M.; Harper, M.K.; Wyche, T.P.; Matainaho, T.K.; Bugni, T.S. Accessing chemical diversity from the uncultivated symbionts of small marine animals. Nat. Chem. Biol. 2018, 14, 179. [Google Scholar] [CrossRef] [PubMed]
- Searle, P.A.; Molinski, T.F. Structure and absolute configuration of (R)-(E)-1-aminotridec-5-en-2-ol, an antifungal amino alcohol from the ascidian Didemnum sp. J. Org. Chem. 1993, 58, 7578–7580. [Google Scholar] [CrossRef]
- Wright, A.D.; Goclik, E.; König, G.M.; Kaminsky, R. Lepadins D−F: Antiplasmodial and antitrypanosomal decahydroquinoline derivatives from the tropical marine tunicate Didemnum sp. J. Med. Chem. 2002, 45, 3067–3072. [Google Scholar] [CrossRef]
- Carroll, A.R.; Nash, B.D.; Duffy, S.; Avery, V.M. Albopunctatone, an antiplasmodial anthrone-anthraquinone from the Australian ascidian Didemnum albopunctatum. J. Nat. Prod. 2012, 75, 1206–1209. [Google Scholar] [CrossRef]
- Lu, Z.; Ding, Y.; Li, X.-C.; Djigbenou, D.R.; Grimberg, B.T.; Ferreira, D.; Ireland, C.M.; Van Wagoner, R.M. 3-Bromohomofascaplysin A, a fascaplysin analogue from a Fijian Didemnum sp. ascidian. Bioorg. Med. Chem. 2011, 19, 6604–6607. [Google Scholar] [CrossRef] [Green Version]
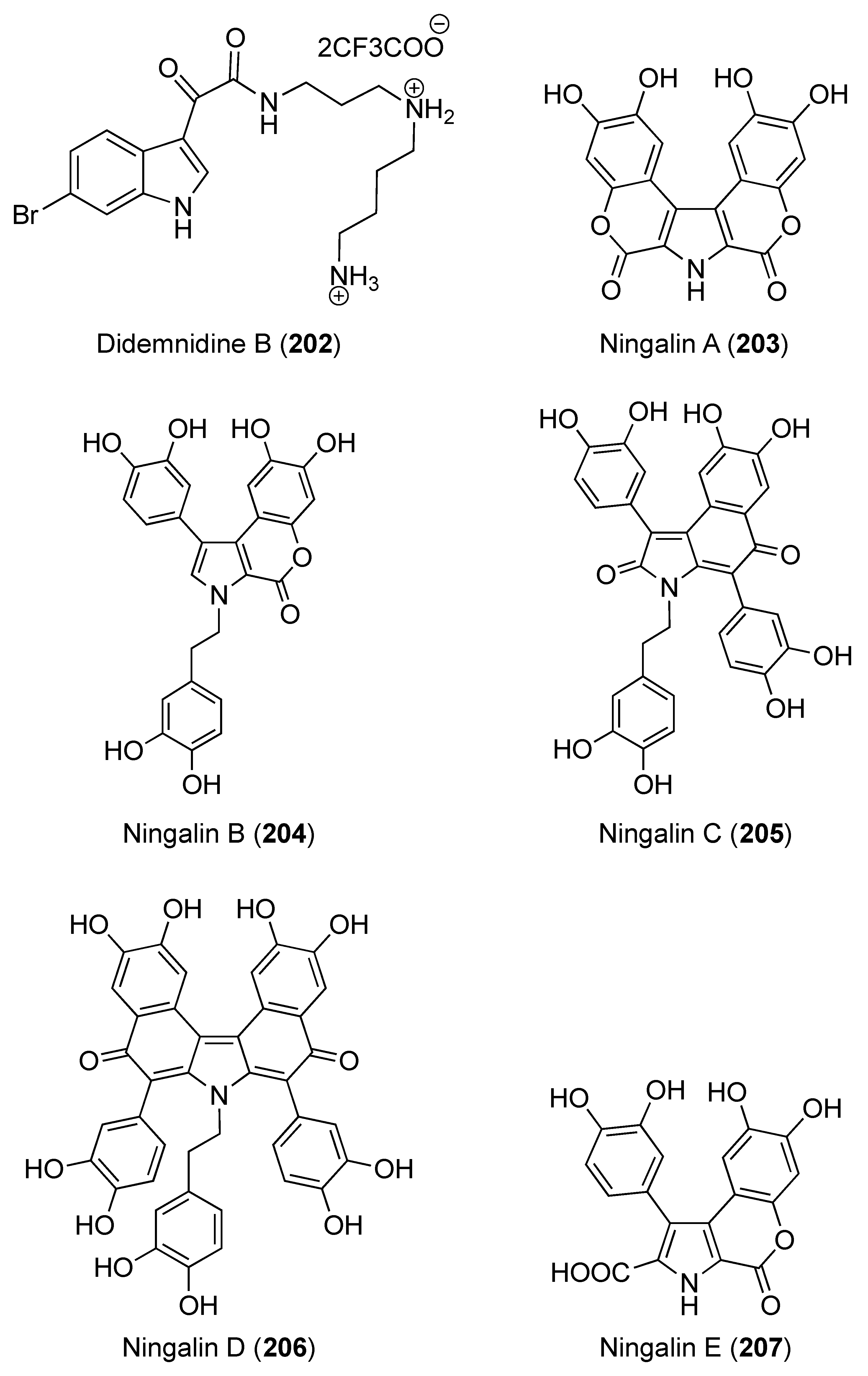
- Finlayson, R.; Pearce, A.N.; Page, M.J.; Kaiser, M.; Bourguet-Kondracki, M.-L.; Harper, J.L.; Webb, V.L.; Copp, B.R. Didemnidines A and B, indole spermidine alkaloids from the New Zealand ascidian Didemnum sp. J. Nat. Prod. 2011, 74, 888–892. [Google Scholar] [CrossRef]
- Choi, S.-S.; Cha, B.-Y.; Kagami, I.; Lee, Y.-S.; Sasaki, H.; Suenaga, K.; Teruya, T.; Yonezawa, T.; Nagai, K.; Woo, J.-T. N, N′-diphenethylurea isolated from Okinawan ascidian Didemnum molle enhances adipocyte differentiation in 3T3-L1 cells. J. Antibiot. 2011, 64, 277–280. [Google Scholar] [CrossRef]
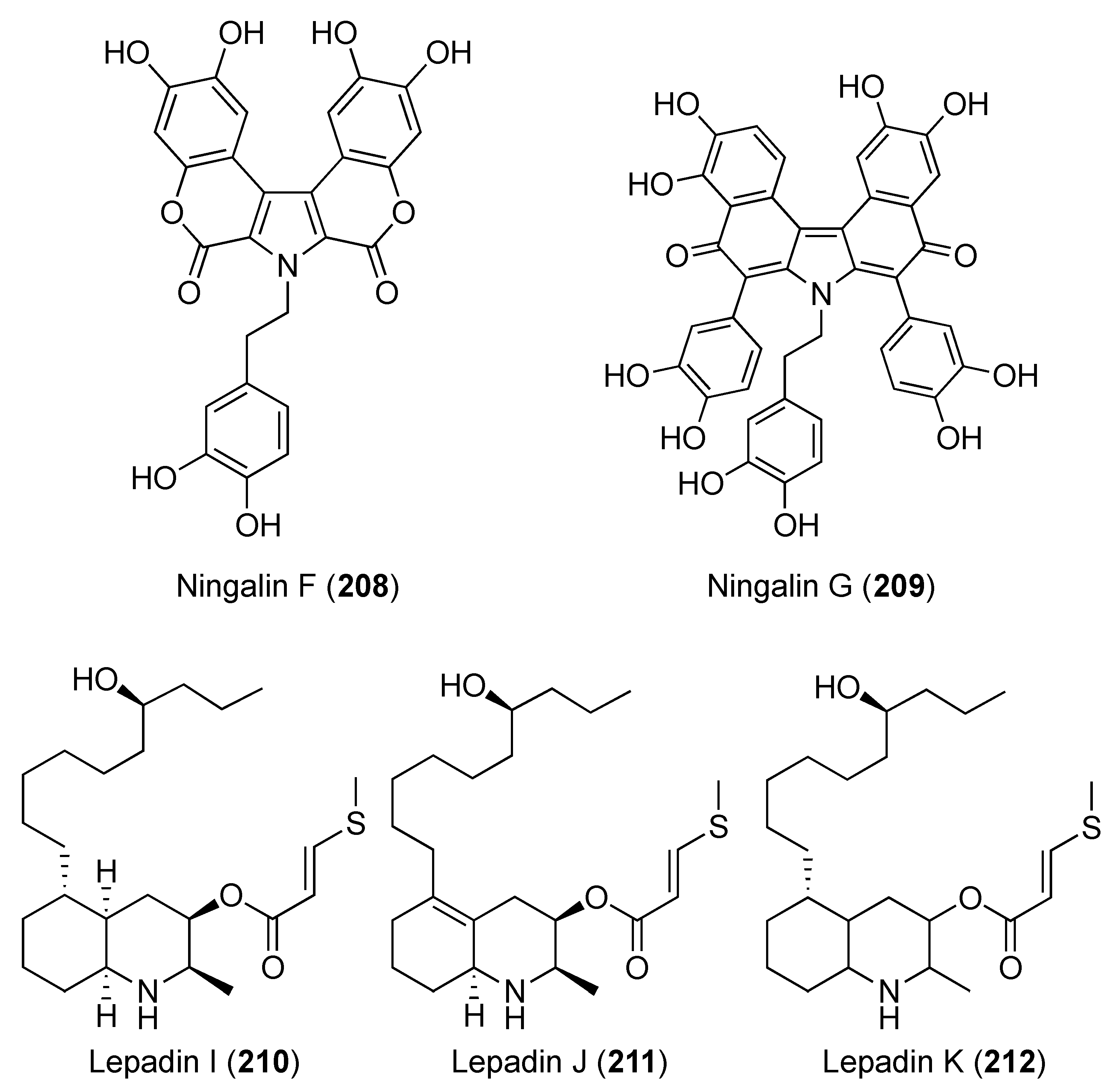
- Kang, H.; Fenical, W. Ningalins A−D: Novel aromatic alkaloids from a western Australian ascidian of the genus Didemnum. J. Org. Chem. 1997, 62, 3254–3262. [Google Scholar] [CrossRef]
- Plisson, F.; Conte, M.; Khalil, Z.; Huang, X.C.; Piggott, A.M.; Capon, R.J. Kinase inhibitor scaffolds against neurodegenerative diseases from a Southern Australian ascidian, Didemnum sp. ChemMedChem 2012, 7, 983–990. [Google Scholar] [CrossRef]
- Ómarsdóttir, S.; Wang, X.; Liu, H.-b.; Duggan, B.M.; Molinski, T.F. Lepadins I–K, 3-O-(3′-methylthio) acryloyloxy-decahydroquinoline esters from a Bahamian ascidian Didemnum sp. Assignment of absolute stereostructures. J. Org. Chem. 2018, 83, 13670–13677. [Google Scholar] [CrossRef]
- Bailly, C. Anticancer properties of lamellarins. Mar. Drugs 2015, 13, 1105–1123. [Google Scholar] [CrossRef] [Green Version]
- Andersen, R.J.; Faulkner, D.J.; He, C.H.; Van Duyne, G.D.; Clardy, J. Metabolites of the marine prosobranch mollusk Lamellaria sp. J. Am. Chem. Soc. 1985, 107, 5492–5495. [Google Scholar] [CrossRef]
- Schmidt, E.W.; Donia, M.S. Life in cellulose houses: Symbiotic bacterial biosynthesis of ascidian drugs and drug leads. Curr. Opin. Biotechnol. 2010, 21, 827–833. [Google Scholar] [CrossRef] [Green Version]
- Arnison, P.G.; Bibb, M.J.; Bierbaum, G.; Bowers, A.A.; Bugni, T.S.; Bulaj, G.; Camarero, J.A.; Campopiano, D.J.; Challis, G.L.; Clardy, J.; et al. Ribosomally synthesized and post-translationally modified peptide natural products: Overview and recommendations for a universal nomenclature. Nat. Prod. Rep. 2013, 30, 108–160. [Google Scholar] [CrossRef]
- Donia, M.S.; Fricke, W.F.; Partensky, F.; Cox, J.; Elshahawi, S.I.; White, J.R.; Phillippy, A.M.; Schatz, M.C.; Piel, J.; Haygood, M.G.; et al. Complex microbiome underlying secondary and primary metabolism in the tunicate-Prochloron symbiosis. Proc. Natl. Acad. Sci. USA 2011, 108, E1423–E1432. [Google Scholar] [CrossRef] [Green Version]
- Donia, M.S.; Ravel, J.; Schmidt, E.W. A global assembly line for cyanobactins. Nat. Chem. Biol. 2008, 4, 341. [Google Scholar] [CrossRef]
- Piel, J.; Hertweck, C.; Shipley, P.R.; Hunt, D.M.; Newman, M.S.; Moore, B.S. Cloning, sequencing and analysis of the enterocin biosynthesis gene cluster from the marine isolate ‘Streptomyces maritimus’: Evidence for the derailment of an aromatic polyketide synthase. Chem. Biol. 2000, 7, 943–955. [Google Scholar] [CrossRef] [Green Version]
- Shen, B.; Yan, X.; Huang, T.; Ge, H.; Yang, D.; Teng, Q.; Rudolf, J.D.; Lohman, J.R. Enediynes: Exploration of microbial genomics to discover new anticancer drug leads. Bioorg. Med. Chem. Lett. 2015, 25, 9–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, J.P.; Schmidt, E.W. The biosynthetic diversity of the animal world. J. Biol. Chem. 2019, 294, 17684–17692. [Google Scholar] [CrossRef] [Green Version]
- Torres, J.P.; Lin, Z.; Winter, J.M.; Krug, P.J.; Schmidt, E.W. Animal biosynthesis of complex polyketides in a photosynthetic partnership. Nat. Commun. 2020, 11, 2882. [Google Scholar] [CrossRef]
- Kwan, J.C.; Donia, M.S.; Han, A.W.; Hirose, E.; Haygood, M.G.; Schmidt, E.W. Genome streamlining and chemical defense in a coral reef symbiosis. Proc. Natl. Acad. Sci. USA 2012, 109, 20655–20660. [Google Scholar] [CrossRef] [Green Version]
- Kwan, J.C.; Tianero, M.D.; Donia, M.S.; Wyche, T.P.; Bugni, T.S.; Schmidt, E.W. Host control of symbiont natural product chemistry in cryptic populations of the tunicate Lissoclinum patella. PLoS ONE 2014, 9, e95850. [Google Scholar] [CrossRef]



























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathogenic Microbial Strain | MIC in µg/mL |
|---|---|
| Staphylococcus aureus ATCC 6538 | 62.5 |
| Staphylococcus aureus ATCC 259223 | 22.6 |
| ORSA 8 | 45.3 |
| ORSA 108 | 91.0 |
| Escherichia coli ATCCNTCC 861 | 125.0 |
| Escherichia coli ATCC 259222 | 45.6 |
| Pseudomonas aeruginosa ATCC 27853 | 22.6 |
| Pseudomonas aeruginosa 13 | 45.3 |
| Pseudomonas aeruginosa P1 | 4.3 |
| Candida albicans ATCC 10231 | n.a. |
| Candida albicans ATCC 36801 (serum type A) | 125.0 |
| Enterococcus faecalis ATCC 14506 | 125.0 |
| Streptococcus sanguinis ATCC 15300 | 125.0 |
| Streptococcus sobrinus ATCC 27607 | 125.0 |
| Streptococcus mutans UA 159 | 62.5 |
| Streptococcus mutans (clinical isolate 2.M7/4) | 31.2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Youssef, D.T.A.; Almagthali, H.; Shaala, L.A.; Schmidt, E.W. Secondary Metabolites of the Genus Didemnum: A Comprehensive Review of Chemical Diversity and Pharmacological Properties. Mar. Drugs 2020, 18, 307. https://doi.org/10.3390/md18060307
Youssef DTA, Almagthali H, Shaala LA, Schmidt EW. Secondary Metabolites of the Genus Didemnum: A Comprehensive Review of Chemical Diversity and Pharmacological Properties. Marine Drugs. 2020; 18(6):307. https://doi.org/10.3390/md18060307
Chicago/Turabian StyleYoussef, Diaa T. A., Hadeel Almagthali, Lamiaa A. Shaala, and Eric W. Schmidt. 2020. "Secondary Metabolites of the Genus Didemnum: A Comprehensive Review of Chemical Diversity and Pharmacological Properties" Marine Drugs 18, no. 6: 307. https://doi.org/10.3390/md18060307