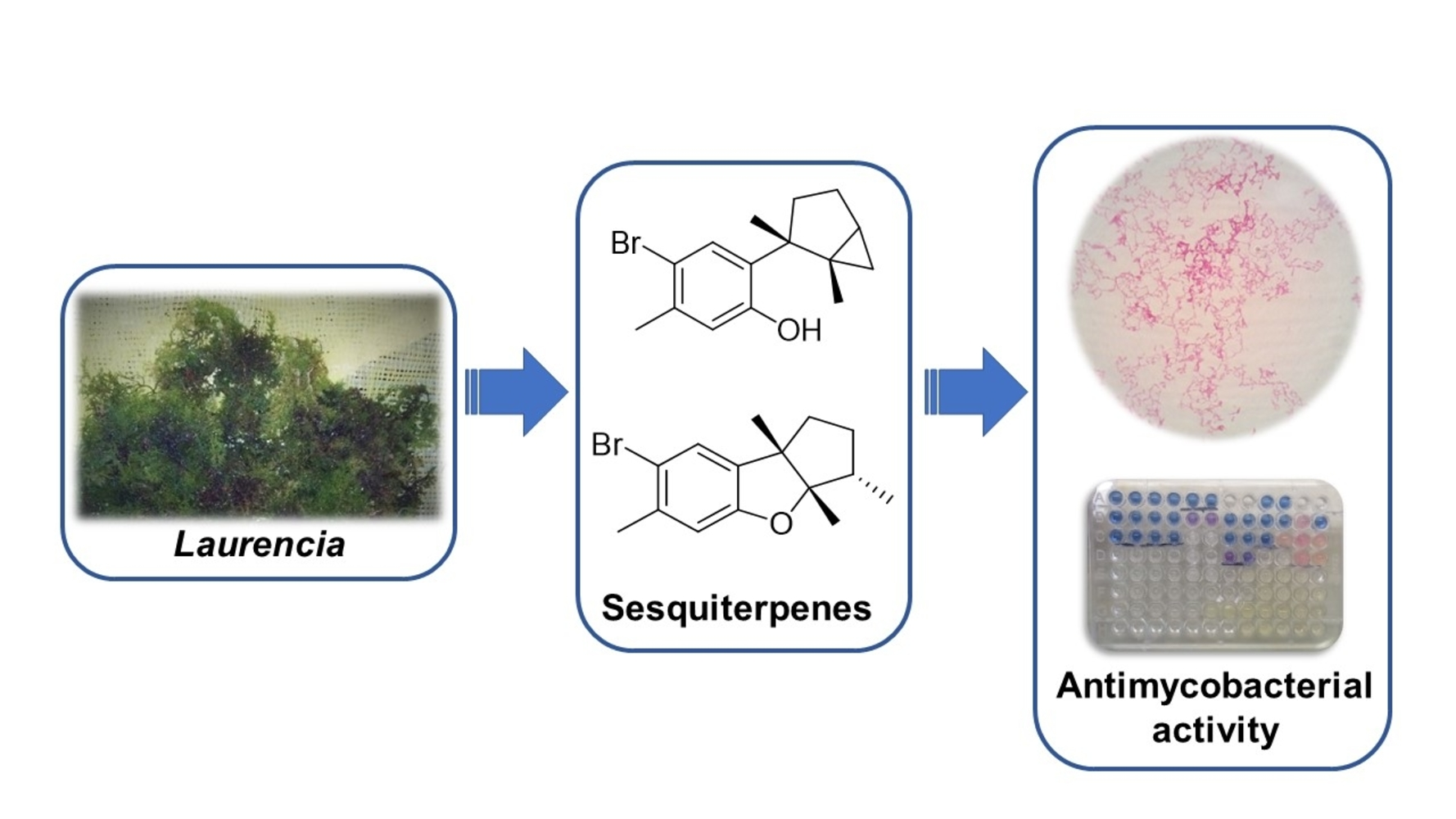
Antimycobacterial Activity of Laurinterol and Aplysin from Laurencia johnstonii



 ,
,  , and
, and
Abstract

1. Introduction
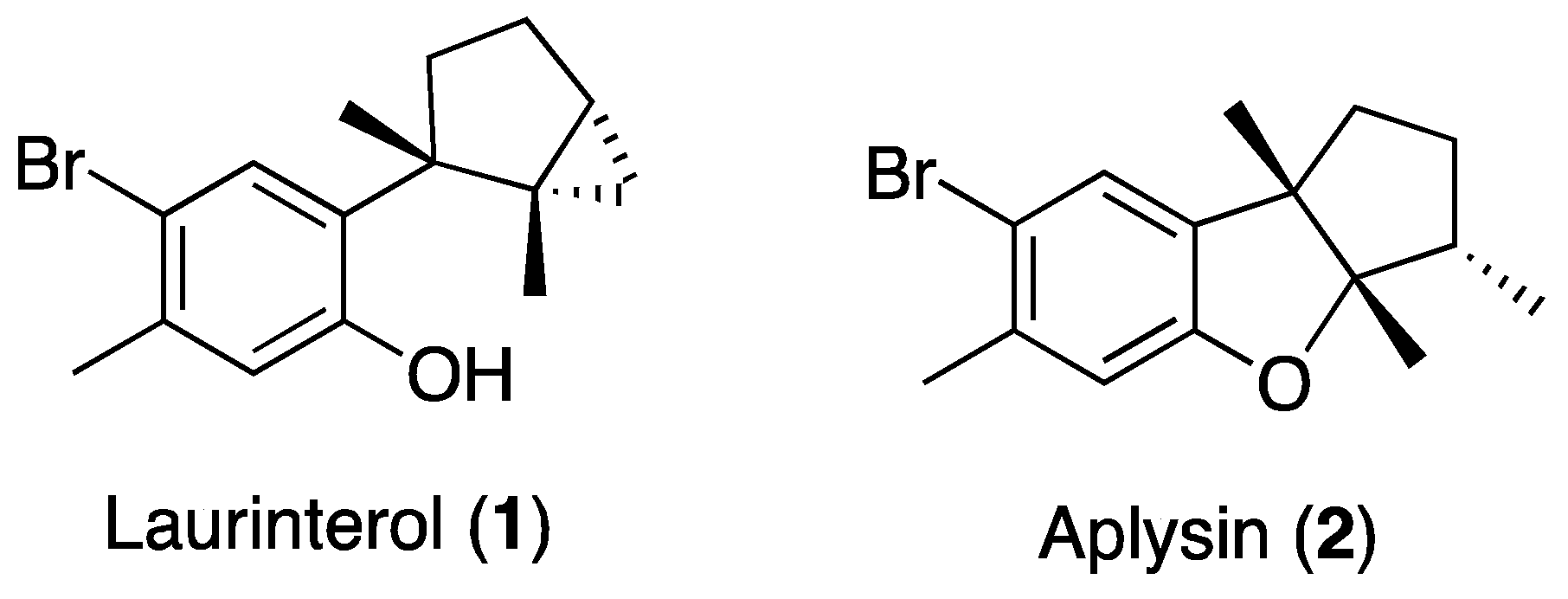
2. Results and Discussion
3. Materials and Methods
3.1. Extraction and Isolation
3.2. Mycobacterial Strains and Culture Conditions
3.3. Determination of the Minimum Inhibitory Concentration (MIC)
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. Antimicrobial Resistance. Available online: https://www.who.int/en/news-room/fact-sheets/detail/antimicrobial-resistance (accessed on 18 February 2020).
- WHO. Tuberculosis. Available online: https://www.who.int/en/news-room/fact-sheets/detail/tuberculosis (accessed on 18 February 2020).
- Espinal, M.A.; Laszlo, A.; Simonsen, L.; Boulahbal, F.; Kin, S.J.; Reniero, A.; Hoffner, S.; Rieder, H.L.; Binkin, N.; Dye, C.; et al. Global trends in resistance to antituberculosis drugs. World Health Organization-International Union against tuberculosis and lung disease working group on anti-tuberculosis drug resistance surveillance. N. Engl. J. Med. 2001, 344, 1294–1303. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, S.L.; Larsen, S.E.; Ordway, D.; Cassell, G.; Coler, R.N. The complexities and challenges of preventing and treating nontuberculous mycobacterial diseases. PLoS Negl. Trop. Dis. 2019, 13, e0007083. [Google Scholar] [CrossRef] [PubMed]
- Patridge, E.; Gareiss, P.; Kinch, M.S.; Hoyer, D. An analysis of FDA-approved drugs: Natural products and their derivatives. Drug Discov. Today 2016, 21, 204–207. [Google Scholar] [CrossRef]
- Wright, G.D. Opportunities for natural products in 21st century antibiotic discovery. Nat. Prod. Rep. 2017, 34, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Gerwick, W.H.; Moore, B.S. Lessons from the past and charting the future of marine natural drugs. Chem. Biol. 2012, 19, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.S. Marine Pharmaceuticals: The Clinical Pipeline. Available online: https://www.midwestern.edu/departments/marinepharmacology/clinical-pipeline.xml (accessed on 18 February 2020).
- Newman, D.J.; Cragg, G.M. Drugs and drug candidates from marine sources: An assessment of the current state of play. Planta Med. 2016, 82, 775–789. [Google Scholar] [CrossRef]
- Harizani, M.; Ioannou, E.; Roussis, V. The Laurencia paradox: An endless source of chemodiversity. Prog. Chem. Org. Nat. Prod. 2016, 102, 91–252. [Google Scholar] [CrossRef]
- Vairappan, C.S.; Suzuki, M.; Abe, T.; Masuda, M. Halogenated metabolites with antibacterial activity from the Okinawan Laurencia species. Phytochemistry 2001, 58, 517–523. [Google Scholar] [CrossRef]
- Vairappan, C.S.; Kawamoto, T.; Miwa, H.; Suzuki, M. Potent antibacterial activity of halogenated compounds against antibiotic-resistant bacteria. Planta Med. 2004, 70, 1087–1090. [Google Scholar] [CrossRef]
- Tsukamoto, S.; Yamashita, Y.; Ohta, T. New cytotoxic and antibacterial compounds isolated from sea hare, Aplysia kurodai. Mar. Drugs 2005, 3, 22–28. [Google Scholar] [CrossRef]
- Kladi, M.; Xenaki, H.; Vagias, C.; Papazafiri, P.; Roussis, V. New cytotoxic sesquiterpenes from the red algae Laurencia obtusa and Laurencia microcladia. Tetrahedron 2006, 62, 182–189. [Google Scholar] [CrossRef]
- García-Davis, S.; Viveros-Valdez, E.; Díaz-Marrero, A.R.; Fernández, J.J.; Valencia-Mercado, D.; Esquivel-Hernández, O.; Carranza-Rosales, P.; Carranza-Torres, I.E.; Guzmán-Delgado, N.E. Antitumoral effect of laurinterol on 3D culture of breast cancer explants. Mar. Drugs 2019, 17, 201. [Google Scholar] [CrossRef]
- Oguri, Y.; Watanabe, M.; Ishikawa, T.; Kamada, T.; Vairappan, C.; Matsuura, H.; Kaneko, K.; Ishii, T.; Suzuki, M.; Yoshimura, E.; et al. New marine antifouling compounds from the red alga Laurencia sp. Mar. Drugs 2017, 15, 267. [Google Scholar] [CrossRef] [PubMed]
- García-Davis, S.; Sifaoui, I.; Reyes-Batlle, M.; Viveros-Valdez, E.; Piñero, J.E.; Lorenzo-Morales, J.; Fernández, J.J.; Díaz-Marrero, A.R. Anti-Acanthamoeba activity of brominated sesquiterpenes from Laurencia johnstonii. Mar. Drugs 2018, 16, 443. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, Y.; Nitanda, N.; Ojika, M.; Sakagami, Y. Aplysiallene, a new bromoallene as an Na, K-ATPase inhibitor from the sea hare, Aplysia kurodai. Biosci. Biotechnol. Biochem. 2001, 65, 474–476. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.; Nagamine, T.; Nguyen, B.C.; Tawata, S. Insecticidal and repellent activities of laurinterol from the Okinawan red alga Laurencia nidifica. Rec. Nat. Prod. 2017, 11, 63–68. [Google Scholar]
- Liu, J.; Ma, N.; Liu, G.; Zheng, L.; Lin, X. Aplysin sensitizes cancer cells to TRAIL by suppressing P38 MAPK/survivin pathway. Mar. Drugs 2014, 12, 5072–5088. [Google Scholar] [CrossRef]
- Zhang, X.; Zhuang, T.; Liang, Z.; Li, L.; Xue, M.; Liu, J.; Liang, H. Breast cancer suppression by aplysin is associated with inhibition of PI3K/AK/FOXO3a pathway. Oncotarget 2017, 8, 63923–63934. [Google Scholar] [CrossRef]
- Gong, A.; Ge, N.; Yao, W.; Lu, L.; Liang, H. Aplysin enhances temozolomide sensitivity in glioma cells by increasing miR-181 level. Cancer Chemother. Pharmacol. 2014, 74, 531–538. [Google Scholar] [CrossRef]
- Gong, A.J.; Gong, L.L.; Yao, W.C.; Ge, N.; Lu, L.X.; Liang, H. Aplysin induces apoptosis in glioma cells through HSP90/AKT pathway. Exp. Biol. Med. 2015, 240, 639–644. [Google Scholar] [CrossRef]
- Novoa-Oponte, L.; Soto Ospina, C.Y. Mycobacterium tuberculosis P-type ATPases: Possible targets for drug or vaccine development. Biomed. Res. Int. 2014, 2014, 296986. [Google Scholar] [CrossRef]
- Lu, P.; Lill, H.; Bald, D. ATP synthase in mycobacteria: Special features and implications for a function as drug target. Biochim. Biophys. Acta 2014, 1837, 1208–1218. [Google Scholar] [CrossRef] [PubMed]
- Grooset, J.H.; Singer, T.G.; Bishai, W.R. New drugs for the treatment of tuberculosis: Hope and reality. Int. J. Tuberc. Lung Dis. 2012, 16, 1005–1014. [Google Scholar] [CrossRef] [PubMed]
- Palomino, J.C.; Martin, A. TMC207 becomes bedaquiline, a new anti-TB drug. Future Microbiol. 2013, 8, 1071–1080. [Google Scholar] [CrossRef]
- Aguilar-Ayala, D.A.; Cnockaert, M.; André, E.; Andries, K.; Gonzalez-Y-Merchand, J.A.; Vandamme, P.; Palomino, J.C.; Martin, A. In vitro activity of bedaquiline against rapidly growing nontuberulous mycobacteria. J. Med. Microbiol. 2017, 66, 1140–1143. [Google Scholar] [CrossRef]
- Ruth, M.M.; Sangen, J.J.N.; Remmers, K.; Pennings, L.J.; Svensson, E.; Aarnoutse, R.E.; Zweijpfenning, S.M.H.; Hoefsloot, W.; Kuipers, S.; Magis-Escurra, C.; et al. A bedaquiline/clofazimine combination regimen might add activity to the treatment of clinically relevant nontuberculous mycobacteria. J. Antimicrob. Chemother. 2018, 74, 935–943. [Google Scholar] [CrossRef]
- Copp, B.R. Antimycobacterial natural products. Nat. Prod. Rep. 2003, 20, 535–557. [Google Scholar] [CrossRef]
- Hou, X.M.; Wang, C.Y.; Gerwick, W.H.; Shao, C.-L. Marine natural products as potential anti-tubercular agents. Eur. J. Med. Chem. 2019, 165, 273–292. [Google Scholar] [CrossRef]
- Grobbelaar, M.; Louw, G.E.; Sampson, S.L.; van Helden, P.D.; Donald, P.R.; Warren, R.M. Evolution of rifampicin treatment for tuberculosis. Infect. Genet. Evol. 2019, 74, 103937. [Google Scholar] [CrossRef]
- Low, J.L.; Wu, M.L.; Aziz, D.B.; Laleu, B.; Dick, T. Screening of TB actives for activity against nontuberculous Mycobacteria delivers high hit rates. Front. Microbiol. 2017, 8, 1539. [Google Scholar] [CrossRef]
- Kaushik, A.; Makkar, N.; Pandey, P.; Parrish, N.; Singh, U.; Lamichhane, G. Carbapenems and rifampin exhibit synergy against Mycobacterium tuberculosis and Mycobacterium abscessus. Antimicrob. Agents Chemother. 2015, 59, 6561–6567. [Google Scholar] [CrossRef] [PubMed]
- Jaganath, D.; Lamichhane, G.; Shah, M. Carbapenems against Mycobacterium tuberculosis: A review of the evidence. Int. J. Tuberc. Lung Dis. 2016, 20, 1436–1447. [Google Scholar] [CrossRef]
- Alcalá, L.; Ruiz-Serrano, M.J.; Pérez-Fernández Turégano, C.; García de Viedma, D.; Díaz-Infantes, M.; Marín-Arriaza, M.; Bouza, E. In vitro activities of linezolid against clinical isolates of Mycobacterium tuberculosis that are susceptible or resistant to first-line antituberculous drugs. Antimicrob. Agents Chemother. 2003, 47, 416–417. [Google Scholar] [CrossRef] [PubMed]
- Tato, M.; de la Pedrosa, E.G.; Canton, R.; Gomez-Garcia, I.; Fortun, J.; Martin-Davila, P.; Baquero, F.; Gomez-Mampaso, E. In vitro activity of linezolid against Mycobacterium tuberculosis complex, including multidrug-resistant Mycobacterium bovis isolates. Int. J. Antimicrob. Agents 2006, 28, 75–78. [Google Scholar] [CrossRef] [PubMed]
- König, G.M.; Wright, A.D.; Franzblau, S.G. Assessment of antimycobacterial activity of a series of mainly marine derived natural products. Planta Med. 2000, 66, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Biá Ventura, T.L.; da SIlva Machado, F.L.; de Araujo, M.H.; de Souza Gestinari, L.M.; Kaiser, C.R.; Esteves, F.d.; Lasunskaia, E.B.; RIbeiro Soares, A.; Frazão Muzitano, M. Nitric oxide production inhibition and anti-mycobacterial activity of extracts and halogenated sesquiterpenes from the Brazilian red alga Laurencia dendroidea J. Agardh. Pharmacogn. Mag. 2015, 11, 611–618. [Google Scholar] [CrossRef]
- Appleton, D.R.; Babcock, R.C.; Copp, B.R. Novel tryptophan-derived dipeptides and bioactive metabolites from the sea hare Aplysia dactylomela. Tetrahedron 2001, 57, 10181–10189. [Google Scholar] [CrossRef]
- Soni, I.; De Groote, M.A.; Dasgupta, A.; Chopra, S. Challenges facing the drug discovery pipeline for non-tuberculous mycobacteria. J. Med. Microbiol. 2016, 65, 1–8. [Google Scholar] [CrossRef]
- Bryant, J.M.; Grogono, D.; Rodriguez, D.; Everall, I.; Brown, K.P.; Moreno, P.; Verma, D.; Hill, E.; Drijkoningen, J.; Gilligan, P.; et al. Emergence and spread of human -transmissible multidrug-resistant nontuberculous mycobacterium. Science 2016, 354, 751–757. [Google Scholar] [CrossRef]
- Wu, M.L.; Aziz, D.B.; Dartois, V.; Dick, T. NTM drug discovery: Status, gaps and the way forward. Drug Discov. Today 2018, 23, 1502–1519. [Google Scholar] [CrossRef]
- García, A.; Bocanegra-García, V.; Palma-Nicolás, J.P.; Rivera, G. Recent advances in antitubercular natural products. Eur. J. Med. Chem. 2012, 49, 1–23. [Google Scholar] [CrossRef]
- Salomon, C.E.; Schmidt, L.E. Natural products as leads for tuberculosis drug development. Curr. Top. Med. Chem. 2012, 12, 735–765. [Google Scholar] [CrossRef] [PubMed]
- Telenti, A.; Marchesi, F.; Balz, M.; Bally, F.; Böttger, E.C.; Bodmer, T. Rapid identification of mycobacteria to the species level by polymerase chain reaction and restriction enzyme analysis. J. Clin. Microbiol. 1993, 31, 175–178. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Susceptibility Testing of Mycobacteria, Nocardiae, and Other Aerobic Actinomycetes; Approved Standard, 2nd ed.; CLSI document M24-A2; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2011; Available online: https://community.clsi.org/media/1463/m24a2_sample.pdf (accessed on 18 February 2020).
- Franzblau, S.G.; Witzig, R.S.; McLaughlin, J.C.; Torres, P.; Madico, G.; Hernandez, A.; Degnan, M.T.; Cook, M.B.; Quenzer, V.K.; Ferguson, R.M.; et al. Rapid, low-technology MIC determination with clinical Mycobacterium tuberculosis isolates by using the microplate Alamar Blue assay. J. Clin. Microbiol. 1998, 36, 362–366. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Strain | Laurinterol µg/mL | Aplysin µg/mL | * Rifampicin µg/mL |
|---|---|---|---|
| M. tuberculosis H37Rv ATCC 27294 | 100 | Nt | 0.1 |
| M. tuberculosis CDC 1551 | 50 | Nt | 0.1 |
| M. tuberculosis LIID-28-99 | 50 | Nt | 0.1 |
| M. tuberculosis LIID-582-15 | 25 | >100 | 0.1 |
| M. tuberculosis LIID-619-15 | 25 | >100 | 1 |
| M. tuberculosis LIID-853-15 | 100 | >100 | 1 |
| M. tuberculosis CIPTIR-F296 | 25 | 50 | 32 |
| M. tuberculosis CIPTIR -D152 | 50 | >100 | 1 |
| M. tuberculosis CIPTIR -C131 | >100 | >100 | 0.1 |
| Strain | Laurinterol µg/mL | Aplysin µg/mL | * Linezolid µg/mL | * Imipenem µg/mL |
|---|---|---|---|---|
| Mycobacterium abscessus LIID- 01 | 6.2 | 100 | 1 | 32 |
| M. abscessus LIID- 02 | 6.2 | 100 | 1 | 32 |
| M. abscessus LIID-03 | 6.2 | >100 | 1 | 8 |
| Mycobacterium fortuitum LIID- 01 | 12.5 | >100 | 8 | 4 |
| Mycobacterium intracellulare LIID-01 | 12.5 | 50 | 2 | 1 |
| M. intracellulare LIID-02 | 25.0 | 100 | 8 | >64 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Davis, S.; Leal-López, K.; Molina-Torres, C.A.; Vera-Cabrera, L.; Díaz-Marrero, A.R.; Fernández, J.J.; Carranza-Rosales, P.; Viveros-Valdez, E. Antimycobacterial Activity of Laurinterol and Aplysin from Laurencia johnstonii. Mar. Drugs 2020, 18, 287. https://doi.org/10.3390/md18060287
García-Davis S, Leal-López K, Molina-Torres CA, Vera-Cabrera L, Díaz-Marrero AR, Fernández JJ, Carranza-Rosales P, Viveros-Valdez E. Antimycobacterial Activity of Laurinterol and Aplysin from Laurencia johnstonii. Marine Drugs. 2020; 18(6):287. https://doi.org/10.3390/md18060287
Chicago/Turabian StyleGarcía-Davis, Sara, Karla Leal-López, Carmen A. Molina-Torres, Lucio Vera-Cabrera, Ana R. Díaz-Marrero, José J. Fernández, Pilar Carranza-Rosales, and Ezequiel Viveros-Valdez. 2020. "Antimycobacterial Activity of Laurinterol and Aplysin from Laurencia johnstonii" Marine Drugs 18, no. 6: 287. https://doi.org/10.3390/md18060287
APA StyleGarcía-Davis, S., Leal-López, K., Molina-Torres, C. A., Vera-Cabrera, L., Díaz-Marrero, A. R., Fernández, J. J., Carranza-Rosales, P., & Viveros-Valdez, E. (2020). Antimycobacterial Activity of Laurinterol and Aplysin from Laurencia johnstonii. Marine Drugs, 18(6), 287. https://doi.org/10.3390/md18060287