Shotgun Proteomics of Ascidians Tunic Gives New Insights on Host–Microbe Interactions by Revealing Diverse Antimicrobial Peptides


 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Protein Identification
2.2. Gene Ontology (GO) Annotation of Proteins Identified with Proteome Discoverer
2.3. Kyoto Encyclopedia of Genes and Genomes Analyses
2.4. Associated Organisms from a Metaproteomic Perspective
2.5. Antimicrobial Peptides Identified with MaxQuant
3. Discussion
3.1. Proteins Identified by Shotgun Proteomics
3.2. Distribution of Bacterial Proteins among Species
3.3. Interaction Host–Microbes
4. Materials and Methods
4.1. Sampling and Protein Extraction
4.2. LC-MS/MS Analyses
4.3. Protein Identification
4.4. MaxQuant Analyses
4.5. Gene Ontology and Kyoto Encyclopedia of Genes and Genomes (KEGG) Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Frazão, B.; Campos, A.; Osório, H.; Thomas, B.; Leandro, S.; Teixeira, A.; Vasconcelos, V.; Antunes, A. Analysis of Pelagia noctiluca proteome Reveals a Red Fluorescent Protein, a Zinc Metalloproteinase and a Peroxiredoxin. Prot. J. 2017, 36, 77–97. [Google Scholar] [CrossRef] [PubMed]
- Frazão, B.; Antunes, A. Jellyfish bioactive compounds: Methods for wet-lab work. Mar. Drugs 2016, 14, 75. [Google Scholar]
- Alex, A.; Vasconcelos, V.; Tamagnini, P.; Santos, A.; Antunes, A. Unusual Symbiotic Cyanobacteria Association in the Genetically Diverse Intertidal Marine Sponge Hymeniacidon perlevis (Demospongiae, Halichondrida). PloS ONE 2012, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Alex, A.; Antunes, A. Whole Genome Sequencing of the Symbiont Pseudovibrio sp. from the Intertidal Marine Sponge Polymastia penicillus Revealed a Gene Repertoire for Host-Switching Permissive Lifestyle. Genome Biol. Evol. 2015, 7, 3022–3032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledoux, J.-B.; Antunes, A.; Haguenauer, A.; Pratlong, M.; Costantini, F.; Abbiati, M.; Aurelle, D. Molecular Forensics into the Sea: How Molecular Markers Can Help to Struggle Against Poaching and Illegal Trade in Precious Corals. In The Cnidaria, Past, Present and Future; Goffredo, S., Dubinsky, Z., Eds.; Springer International Publishing: Cham, Switzerland, 2016; ISBN 9783319313054. [Google Scholar]
- Ledoux, J.-B.; Antunes, A. Beyond the beaten path: Improving natural products bioprospecting using an eco-evolutionary framework-the case of the octocorals. Crit. Rev. Biotechnol. 2018, 38, 184–198. [Google Scholar] [CrossRef] [PubMed]
- Tsagkogeorga, G.; Turon, X.; Hopcroft, R.R.; Tilak, M.-K.; Feldstein, T.; Shenkar, N.; Loya, Y.; Huchon, D.; Douzery, E.J.P.; Delsuc, F. An updated 18S rRNA phylogeny of tunicates based on mixture and secondary structure models. Bmc Evol. Biol. 2009, 9, 187. [Google Scholar] [CrossRef] [Green Version]
- Di Gregorio, A.; Levine, M. Ascidian embryogenesis and the origins of the chordate body plan. Curr. Opin. Genet. Dev. 1998, 8, 457–463. [Google Scholar] [CrossRef]
- Lambert, G. Ecology and natural history of the protochordates. Can. J. Zool. 2005, 83, 34–50. [Google Scholar] [CrossRef]
- Aldred, N.; Clare, A.S. Mini-review: Impact and dynamics of surface fouling by solitary and compound ascidians. Biofouling 2014, 30, 259–270. [Google Scholar] [CrossRef]
- Shenkar, N.; Swalla, B.J. Global diversity of Ascidiacea. PloS ONE 2011, 6, e20657. [Google Scholar] [CrossRef]
- Hirose, E. Ascidian tunic cells: Morphology and functional diversity of free cells outside the epidermis. Invertebr. Biol. 2009, 128, 83–96. [Google Scholar] [CrossRef]
- Behrendt, L.; Larkum, A.W.D.; Trampe, E.; Norman, A.; Sørensen, S.J.; Kühl, M. Microbial diversity of biofilm communities in microniches associated with the didemnid ascidian Lissoclinum patella. ISME J. 2012, 6, 1222–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blasiak, L.C.; Zinder, S.H.; Buckley, D.H.; Hill, R.T. Bacterial diversity associated with the tunic of the model chordate Ciona intestinalis. ISME J. 2014, 8, 309–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donia, M.S.; Fricke, W.F.; Partensky, F.; Cox, J.; Elshahawi, S.I.; White, J.R.; Phillippy, A.M.; Schatz, M.C.; Piel, J.; Haygood, M.G.; et al. Complex microbiome underlying secondary and primary metabolism in the tunicate-Prochloron symbiosis. Proc. Natl. Acad. Sci. USA 2011, 108, E1423–E1432. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.S.; Erwin, P.M.; Shenkar, N.; López-Legentil, S. A comparison of prokaryotic symbiont communities in nonnative and native ascidians from reef and harbor habitats. FEMS Microbiol. Ecol. 2018, 77058, 1–31. [Google Scholar]
- Erwin, P.M.; Pineda, M.C.; Webster, N.; Turon, X.; López-Legentil, S. Down under the tunic: Bacterial biodiversity hotspots and widespread ammonia-oxidizing archaea in coral reef ascidians. ISME J.. 2014, 8, 575–588. [Google Scholar] [CrossRef] [Green Version]
- Rinehart, K.L.; Holt, T.G.; Fregeau, N.L.; Stroh, J.G.; Keifer, P.A.; Sun, F.; Li, L.H.; Martin, D.G. Ecteinascidins 729, 743, 745, 759A, 759B, and 770: Potent antitumor agents from the Caribbean tunicate Ecteinascidia turbinata. J. Org. Chem. 1990, 55, 4512–4515. [Google Scholar] [CrossRef]
- Rath, C.M.; Janto, B.; Earl, J.; Ahmed, A.; Hu, F.Z.; Hiller, L.; Dahlgren, M.; Kreft, R.; Yu, F.; Wolff, J.J.; et al. Meta-omic characterization of the marine invertebrate microbial consortium that produces the chemotherapeutic natural product ET-743. ACS Chem. Biol. 2011, 6, 1244–1256. [Google Scholar] [CrossRef] [Green Version]
- Hirose, E.; Hirabayashi, S.; Hori, K.; Kasai, F.; Watanabe, M.M. UV Protection in the Photosymbiotic Ascidian Didemnum molle Inhabiting Different Depths. Zool. Sci. 2006, 23, 57–63. [Google Scholar] [CrossRef]
- Hirose, E. Ascidian Photosymbiosis: Diversity of Cyanobacterial Transmission During Embryogenesis. Genesis 2015, 53, 121–131. [Google Scholar] [CrossRef]
- Lin, Z.; Torres, J.P.; Tianero, M.D.; Kwan, J.C.; Schmidt, E.W. Origin of chemical diversity in the Prochloron-Tunicate Symbiosis. Appl. Environ. Microbiol. 2016, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwan, J.C.; Donia, M.S.; Han, A.W.; Hirose, E.; Haygood, M.G.; Schmidt, E.W. Genome streamlining and chemical defense in a coral reef symbiosis. Proc. Natl. Acad. Sci. USA 2012, 109, 20655–20660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwan, J.C.; Schmidt, E.W. Bacterial endosymbiosis in a chordate host: Long-term co-evolution and conservation of secondary metabolism. PLoS ONE 2013, 8, e80822. [Google Scholar] [CrossRef] [PubMed]
- Donia, M.S.; Fricke, W.F.; Ravel, J.; Schmidt, E.W. Variation in tropical reef symbiont metagenomes defined by secondary metabolism. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alex, A.; Silva, V.; Vasconcelos, V.; Antunes, A. Evidence of unique and generalist microbes in distantly related sympatric intertidal marine sponges (Porifera: Demospongiae). PLoS ONE 2013, 8, e80653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alex, A.; Antunes, A. Pyrosequencing Characterization of the Microbiota from Atlantic Intertidal Marine Sponges Reveals High Microbial Diversity and the Lack of Co-Occurrence Patterns. PLoS ONE 2015, 10, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Behrendt, L.; Raina, J.-B.; Lutz, A.; Kot, W.; Albertsen, M.; Halkjær-Nielsen, P.; Sørensen, S.J.; Larkum, A.W.; Kühl, M. In situ metabolomic- and transcriptomic-profiling of the host-associated cyanobacteria Prochloron and Acaryochloris marina. ISME J. 2018, 12, 556–567. [Google Scholar] [CrossRef] [Green Version]
- Campagna, D.; Gasparini, F.; Franchi, N.; Vitulo, N.; Ballin, F.; Manni, L.; Valle, G.; Ballarin, L. Transcriptome dynamics in the asexual cycle of the chordate Botryllus schlosseri. BMC Genom. 2016, 17, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Ricci, L.; Chaurasia, A.; Lapébie, P.; Dru, P.; Helm, R.R.; Copley, R.R.; Tiozzo, S. Identification of differentially expressed genes from multipotent epithelia at the onset of an asexual development. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, D.; Sanders, E.N.; Farell, K.; Langenbacher, A.D.; Taketa, D.A.; Hopper, M.R.; Kennedy, M.; Gracey, A.; De Tomaso, A.W. Analysis of the basal chordate Botryllus schlosseri reveals a set of genes associated with fertility. Bmc Genom. 2014, 15, 1183. [Google Scholar] [CrossRef] [Green Version]
- Kawada, T.; Shiraishi, A.; Aoyama, M.; Satake, H. Transcriptomes of the Premature and Mature Ovaries of an Ascidian, Ciona intestinalis. Front. Endocrinol. 2017, 8, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, M.; Nakajima, A.; Inaba, K. Proteomic profiles of embryonic development in the ascidian Ciona intestinalis. Dev. Biol. 2009, 325, 468–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuoka, T.; Ikeda, T.; Fujimaki, K.; Satou, Y. Transcriptome dynamics in early embryos of the ascidian, Ciona intestinalis. Dev. Biol. 2013, 384, 375–385. [Google Scholar] [CrossRef]
- Lopez, C.E.; Sheehan, H.C.; Vierra, D.A.; Azzinaro, P.A.; Meedel, T.H.; Howlett, N.G.; Irvine, S.Q. Proteomic responses to elevated ocean temperature in ovaries of the ascidian Ciona intestinalis. Biol. Open 2017, 6, 943–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palanisamy, S.K.; Trisciuoglio, D.; Zwergel, C.; Del Bufalo, D.; Mai, A. Metabolite profiling of ascidian Styela plicata using LC–MS with multivariate statistical analysis and their antitumor activity. J. Enzym. Inhib. Med. Chem. 2017, 32, 614–623. [Google Scholar] [CrossRef] [Green Version]
- Kuplik, Z.; Novak, L.; Shenkar, N. Proteomic profiling of ascidians as a tool for biomonitoring marine environments. PLoS ONE 2019, 14, e0215005. [Google Scholar] [CrossRef] [PubMed]
- Brozovic, M.; Dantec, C.; Dardaillon, J.; Dauga, D.; Faure, E.; Gineste, M.; Louis, A.; Naville, M.; Nitta, K.R.; Piette, J.; et al. ANISEED 2017: Extending the integrated ascidian database to the exploration and evolutionary comparison of genome-scale datasets. Nucleic Acids Res. 2018, 46, D718–D725. [Google Scholar] [CrossRef]
- Tianero, M.D.B.; Kwan, J.C.; Wyche, T.P.; Presson, A.P.; Koch, M.; Barrows, L.R.; Bugni, T.S.; Schmidt, E.W. Species specificity of symbiosis and secondary metabolism in ascidians. ISME J. 2015, 9, 615–628. [Google Scholar] [CrossRef] [Green Version]
- Aguilera-Mendoza, L.; Marrero-Ponce, Y.; Tellez-Ibarra, R.; Llorente-Quesada, M.T.; Salgado, J.; Barigye, S.J.; Liu, J. Overlap and diversity in antimicrobial peptide databases: Compiling a non-redundant set of sequences. Bioinformatics 2015, 31, 2553–2559. [Google Scholar] [CrossRef] [Green Version]
- Kleiner, M. Metaproteomics: Much More than Measuring Gene Expression in Microbial Communities. mSystems 2019, 4, e00115–e00119. [Google Scholar] [CrossRef] [Green Version]
- Di Bella, M.A.; Fedders, H.; De Leo, G.; Leippe, M. Localization of antimicrobial peptides in the tunic of Ciona intestinalis (Ascidiacea, Tunicata) and their involvement in local inflammatory-like reactions. Results Immunol. 2011, 1, 70–75. [Google Scholar] [CrossRef] [Green Version]
- Franchi, N.; Ballarin, L. Immunity in Protochordates: The Tunicate Perspective. Front. Immunol. 2017, 8, 674. [Google Scholar] [CrossRef] [PubMed]
- Litman, G.W.; Cannon, J.P.; Dishaw, L.J. Reconstructing immune phylogeny: New perspectives. Nat. Rev. Immunol. 2005, 5, 866–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyholm, S.V.; Graf, J. Knowing your friends: Invertebrate innate immunity fosters beneficial bacterial symbioses. Nat. Rev. Microbiol. 2012, 10, 815–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiebe, M.S.; Traktman, P. Poxviral B1 Kinase Overcomes Barrier to Autointegration Factor, a Host Defense against Virus Replication. Cell Host Microbe 2007, 1, 187–197. [Google Scholar] [CrossRef] [Green Version]
- Immesberger, A.; Burmester, T. Putative phenoloxidases in the tunicate Ciona intestinalis and the origin of the arthropod hemocyanin superfamily. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2004, 174, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Cammarata, M.; Parrinello, N. The ascidian prophenoloxidase activating system. Invertebr. Surviv. J. 2009, 6, S67–S76. [Google Scholar]
- Cerenius, L.; Soderhall, K. The prophenoloxidase-activating system in invertebrates. Immunol. Rev. 2004, 198, 116–126. [Google Scholar] [CrossRef]
- Dishaw, L.J.; Giacomelli, S.; Melillo, D.; Zucchetti, I.; Haire, R.N.; Natale, L.; Russo, N.A.; De Santis, R.; Litman, G.W.; Pinto, M.R. A role for variable region-containing chitin-binding proteins (VCBPs) in host gut-bacteria interactions. Proc. Natl. Acad. Sci. USA 2011, 108, 16747–16752. [Google Scholar] [CrossRef] [Green Version]
- Matsushita, M.; Thiel, S.; Jensenius, J.C.; Terai, I.; Fujita, T. Proteolytic Activities of Two Types of Mannose-Binding Lectin-Associated Serine Protease. J. Immunol. 2000, 165, 2637–2642. [Google Scholar] [CrossRef] [Green Version]
- Fujita, T. Evolution of the lectin–complement pathway and its role in innate immunity. Nat. Rev. Immunol. 2002, 2, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.R.; Vasconcelos, V.M.; Antunes, A. The phosphoprotein phosphatase family of Ser/Thr phosphatases as principal targets of naturally occurring toxins. Crit. Rev. Toxicol. 2011, 41, 83–110. [Google Scholar] [CrossRef]
- Pereira, S.R.; Vasconcelos, V.M.; Antunes, A. Computational study of the covalent bonding of microcystins to cysteine residues-a reaction involved in the inhibition of the PPP family of protein phosphatases. FEBS J. 2013, 280, 674–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, P.; Swiatlo, E. A multifaceted role for polyamines in bacterial pathogens. Mol. Microbiol. 2008, 68, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Wallace, H.M. The polyamines: Past, present and future. Essays Biochem. 2009, 46, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Cohen, S.S. Spermidine and Other Bacterial Polyamines. In A Guide to the Polyamines; Oxford University Press: Oxford, United Kingdom, 1998; ISBN 9780195110647. [Google Scholar]
- Cárdenas, A.; Neave, M.J.; Haroon, M.F.; Pogoreutz, C.; Rädecker, N.; Wild, C.; Gärdes, A.; Voolstra, C.R. Excess labile carbon promotes the expression of virulence factors in coral reef bacterioplankton. ISME J. 2018, 12, 59–76. [Google Scholar] [CrossRef] [Green Version]
- Linhartová, I.; Bumba, L.; Mašín, J.; Basler, M.; Osička, R.; Kamanová, J.; Procházková, K.; Adkins, I.; Hejnová-Holubová, J.; Sadílková, L.; et al. RTX proteins: A highly diverse family secreted by a common mechanism. FEMS Microbiol. Rev. 2010, 34, 1076–1112. [Google Scholar] [CrossRef] [Green Version]
- Frey, J. The role of RTX toxins in host specificity of animal pathogenic Pasteurellaceae. Vet. Microbiol. 2011, 153, 51–58. [Google Scholar] [CrossRef]
- Welch, R.A. RTX Toxin Structure and Function: A Story of Numerous Anomalies and Few Analogies in Toxin Biology. In Current Topics in Microbiology and Immunology; van der Goot, F.G., Ed.; Springer: Berlin, Heidelberg, Germany, 2001; ISBN 978-3-642-56508-3. [Google Scholar]
- Eberl, G. A new vision of immunity: Homeostasis of the superorganism. Mucosal Immunol. 2010, 3, 450–460. [Google Scholar] [CrossRef]
- Riley, M.A.; Goldstone, C.M.; Wertz, J.E.; Gordon, D. A phylogenetic approach to assessing the targets of microbial warfare. J. Evol. Biol. 2003, 16, 690–697. [Google Scholar] [CrossRef]
- Ghequire, M.G.K.; De Mot, R. Ribosomally encoded antibacterial proteins and peptides from Pseudomonas. FEMS Microbiol. Rev. 2014, 38, 523–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCaughey, L.C.; Ritchie, N.D.; Douce, G.R.; Evans, T.J.; Walker, D. Efficacy of species-specific protein antibiotics in a murine model of acute Pseudomonas aeruginosa lung infection. Sci. Rep. 2016, 6, 30201. [Google Scholar] [CrossRef] [PubMed]
- Ghequire, M.G.K.; Swings, T.; Michiels, J.; Buchanan, S.K.; De Mot, R. Hitting with a BAM: Selective Killing by Lectin-Like Bacteriocins. mBio 2018, 9, e02138-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dischinger, J.; Josten, M.; Szekat, C.; Sahl, H.-G.; Bierbaum, G. Production of the Novel Two-Peptide Lantibiotic Lichenicidin by Bacillus licheniformis DSM 13. PloS ONE 2009, 4, e6788. [Google Scholar] [CrossRef] [PubMed]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins-a viable alternative to antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Conesa, A.; Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M. From genomics to chemical genomics: New developments in KEGG. Nucleic Acids Res. 2006, 34, D354–D357. [Google Scholar] [CrossRef]
- Moriya, Y.; Itoh, M.; Okuda, S.; Yoshizawa, A.C.; Kanehisa, M. KAAS: An automatic genome annotation and pathway reconstruction server. Nucleic Acids Res. 2007, 35, W182–W185. [Google Scholar] [CrossRef] [Green Version]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 1999, 27, 29–34. [Google Scholar] [CrossRef] [Green Version]
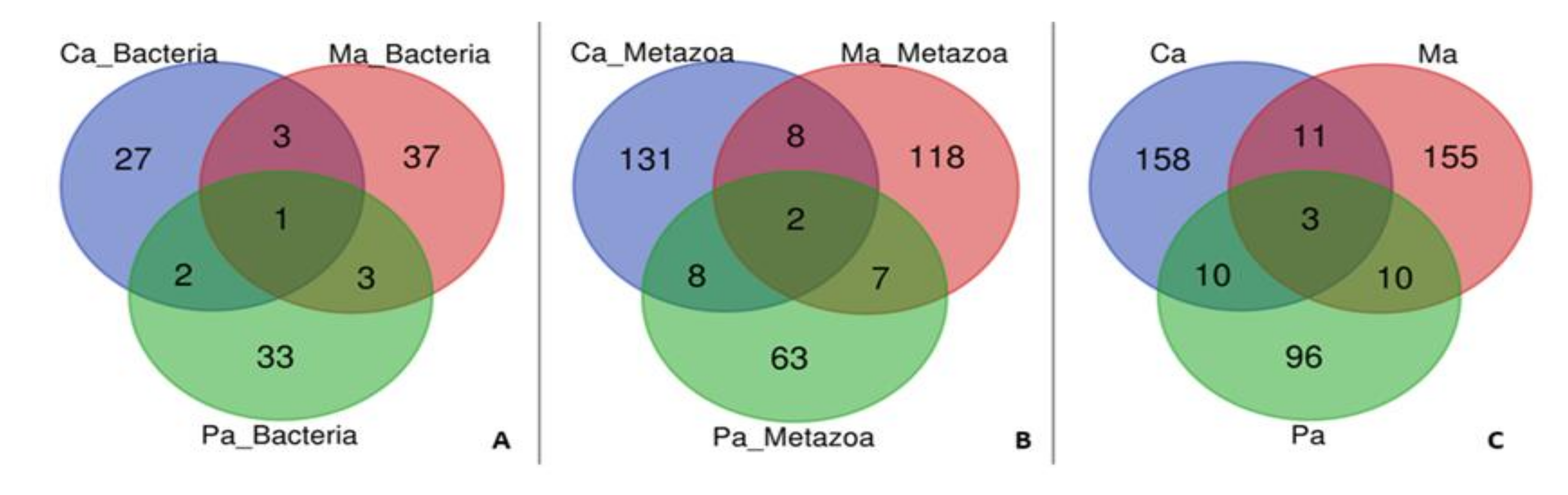
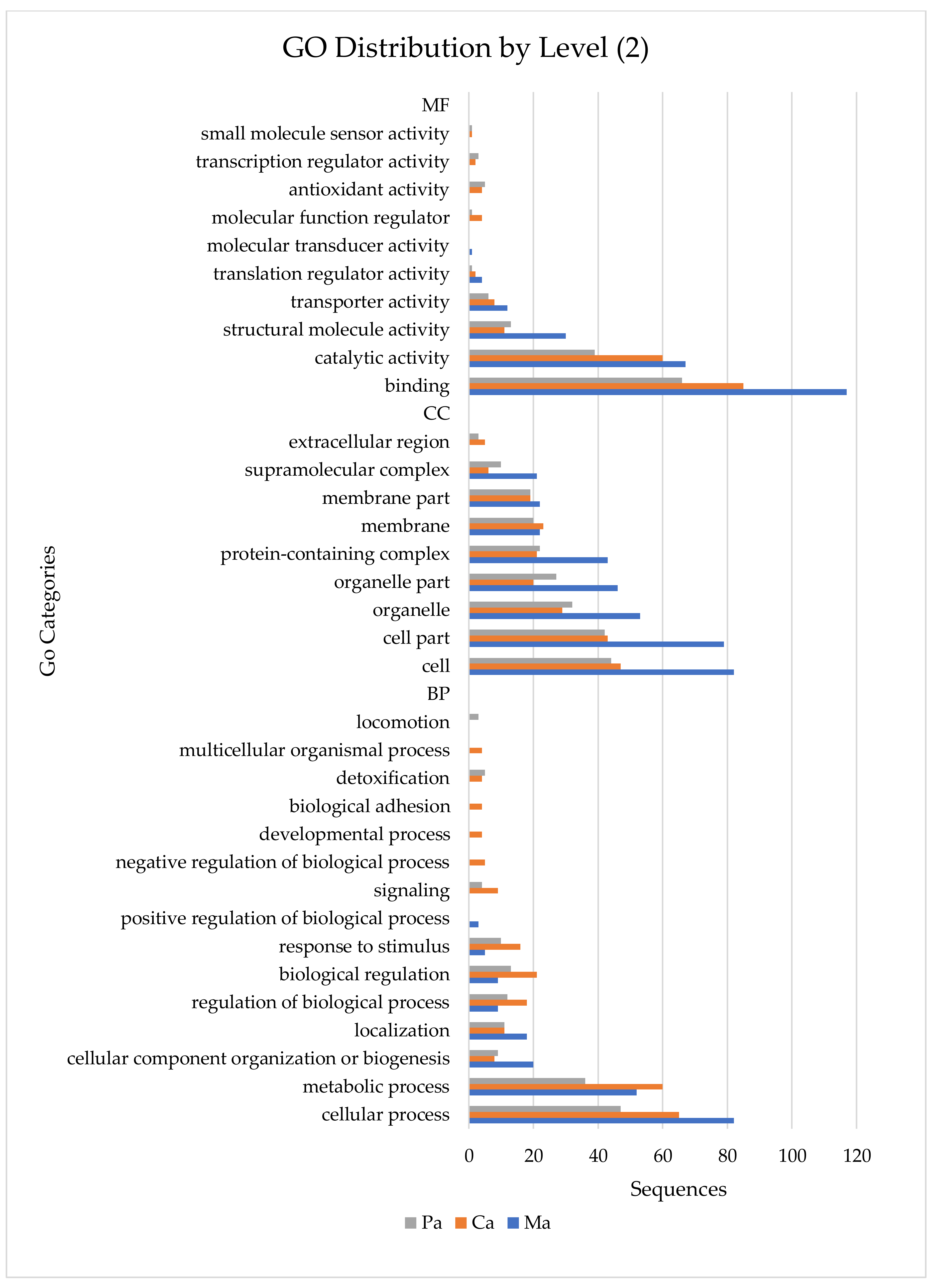
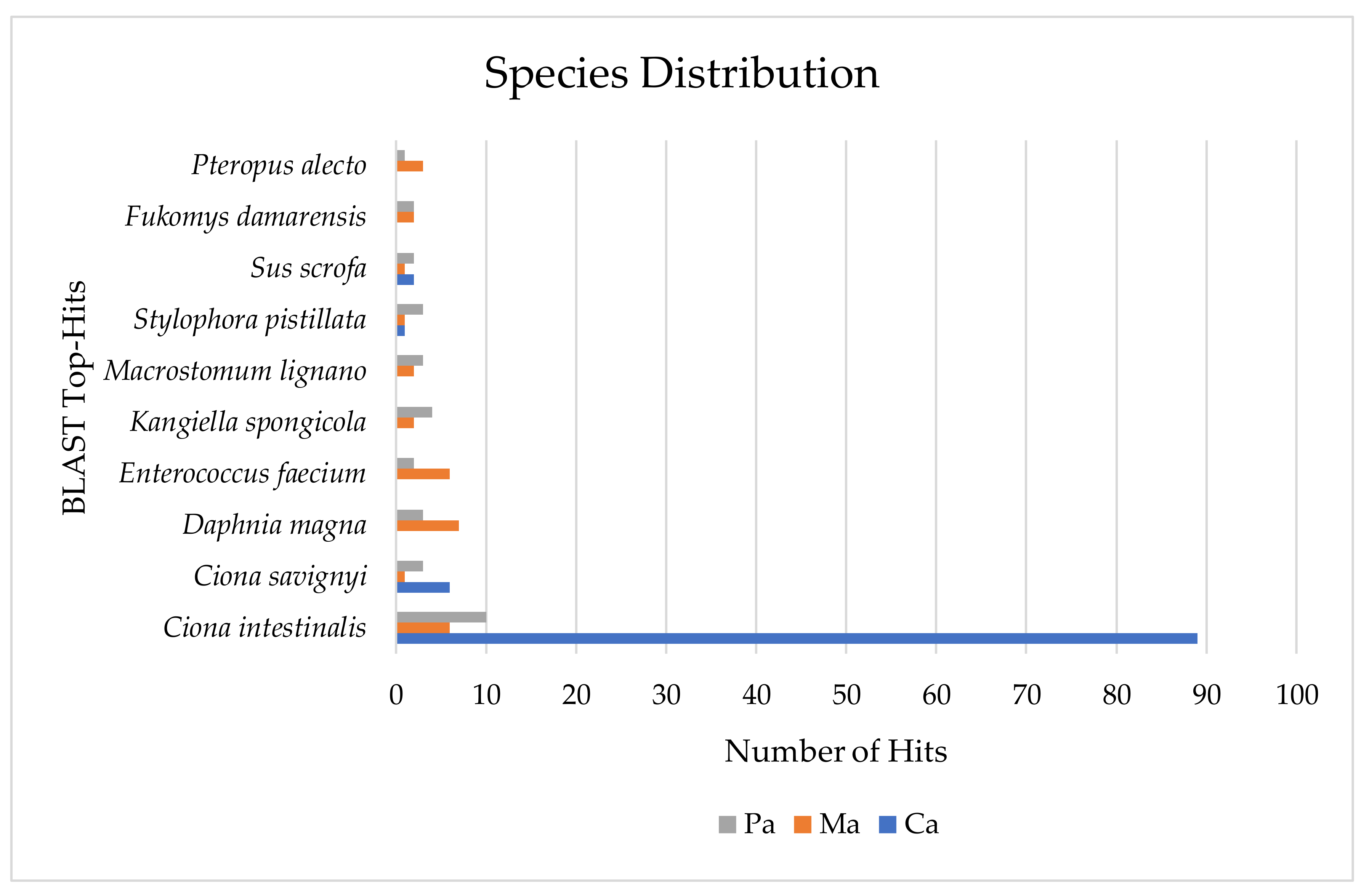


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UniProtKB Taxonomic Section 1 | Species 2 | Accession Number 3 | Protein Description 4 |
|---|---|---|---|
| Bacteria | Ca, Ma, Pa | A0A1E2TY30 | Actin, cytoplasmic 2 |
| Ca, Ma | A0A2I6SAN9 A0A2D9B6R8 A0A368ML82 | Ubiquitin Histone domain-containing protein Histidine kinase | |
| Ca, Pa | A0A0A8RA46 A0A293NCX8 | Lysyl endopeptidase, EC 3.4.21.50 TAF domain-containing protein | |
| Ma, Pa | A0A2V2P8C5 A0A318CZJ6 A0A1C7PFN9 | Tubulin domain-containing protein Myosin_tail_1 domain-containing protein Tubulin_C domain-containing protein | |
| Metazoa | Ca, Ma, Pa | A0A287B5W2 A0A1W5BGH3 | Trypsinogen isoform X1 Myosin-10 isoform X2 |
| Ca, Ma | A0A182L962 A0A2U3WDE6 A0A368GTV8 F6PP44 F6KMG7 F7D7P2 A0A2Y9E533 L7N0S7 | Uncharacterized protein Histone H2B Ribosomal protein S3 Fructose-bisphosphate aldolase, EC 4.1.2.13 Actin Uncharacterized protein Uncharacterized protein LOC101342124ATP synthase subunit alpha | |
| Ca, Pa | F6SM47 B0LXF7 A0A1W2W3E0 A0A1W5BMF0 A2SY09 A0A1W3JRI0 A0A1W3JCQ2 A0A1W3JCW3 | Dolichyl-diphosphooligosaccharide--protein glycosyltransferase 48 kDa subunit, Oligosaccharyl transferase 48 kDa subunit Aprotinin Tropomyosin, smooth muscle/fibroblast CTM1-like isoform X1 (tropomyosin, smooth muscle/fibroblast CTM1-like isoform X2) Talin-2 isoform X1 Actin (act protein isoform X1) r27a protein isoform X1 Glial fibrillary acidic protein isoform X1P-selectin-like | |
| Ma, Pa | L5K7V0 N6TCR8 A0A2U3Z9L8 Q95PQ7 A0A091CUF9 A0A212D7H3 A0A074ZCV8 | Tubulin beta-2C chain Uncharacterized protein LOW QUALITY PROTEIN: actin, gamma 1 Intermediate filament protein C Keratin, type II cytoskeletal 1 MYH1 Uncharacterized protein |
| Species | Ca | Ma | Pa | ||||
|---|---|---|---|---|---|---|---|
| Pathway | Enzymes in Pathway | Sequences of Enzymes | Enzymes in Pathway | Sequences of Enzymes | Enzymes in Pathway | Sequences of Enzymes | |
| Biosynthesis of antibiotics | 9 | 9 | 10 | 10 | 4 | 4 | |
| Purine metabolism | 5 | 13 | 6 | 40 | 1 | 16 | |
| Glycolysis/Gluconeogenesis | 4 | 4 | 6 | 6 | 1 | 1 | |
| Carbon fixation in photosynthetic organisms | 3 | 3 | 4 | 4 | N.F. | N.F. | |
| Methane metabolism | 3 | 3 | 2 | 2 | 1 | 1 | |
| Pentose phosphate pathway | 2 | 2 | 1 | 1 | 2 | 2 | |
| Glutathione metabolism | 2 | 3 | N.F. | N.F. | 2 | 3 | |
| Thiamine metabolism | 1 | 8 | 1 | 33 | 1 | 16 | |
| Drug metabolism-other enzymes | 2 | 2 | 1 | 1 | N.F. | N.F. | |
| Pyruvate metabolism | 2 | 2 | 1 | 1 | N.F. | N.F. | |
| Glyoxylate and dicarboxylate metabolism | 1 | 1 | 1 | 1 | 1 | 1 | |
| PD-L1 expression and PD-1 checkpoint pathway in cancer | 1 | 1 | 1 | 1 | 1 | 1 | |
| T cell receptor signaling pathway | 1 | 1 | 1 | 1 | 1 | 1 | |
| Th1 and Th2 cell differentiation | 1 | 1 | 1 | 1 | 1 | 1 | |
| Fructose and mannose metabolism | 2 | 2 | 1 | 1 | N.F. | N.F. | |
| Species | Ca | Ma | Pa | |
|---|---|---|---|---|
| Phylum | ||||
| Chordata | 112 | 56 | 34 | |
| Arthropoda | 10 | 41 | 18 | |
| Proteobacteria | 15 | 14 | 19 | |
| Nematoda | 9 | 12 | 5 | |
| Platyhelminthes | 2 | 9 | 7 | |
| Firmicutes | 1 | 7 | 3 | |
| Cnidaria | 4 | 1 | 4 | |
| AMP_ID 1 | PGs ID 2 (peptides) | PEP 3 | Original Database 4 | AMP Description/Main Activity 5 |
|---|---|---|---|---|
| Overall_15444|DAMPD_732|11\DAMPD\DAMPD_548|H2B_LITVA | 0 (4) | 6.959e-119 | DAMPD | Core histone H2A/H2B/H3/H4 Defence response to bacteria G+/– |
| Overall_11263|CAMP_Validated_1724|CAMPSQ4167|Histone H2B | CAMP | Core histone H2B Defence response to bacteria G+/– | ||
| Overall_15445|DAMPD_733|11\DAMPD\DAMPD_549|H2B_RHASC | DAMPD | Core histone H2B Response to bacteria G+/– | ||
| Overall_15492|DAMPD_780|11\DAMPD\DAMPD_591|ANN2_AREMA | 1 (1) | 0.006 | DAMPD | Arenicin-2 from Arenicola marina (Lugworm) Antibacterial G+/– |
| Overall_33701|Yadamp_1162|2043|Beta-defensin 20 | 2 (1) | 2.752e-05 | YADAMP | Beta defensin Antibacterial and Antiviral |
| Overall_31051|UniProtKb_1300|Q30KP3;Q8C5A7|DFB20_MOUSE | UniProtKB | Beta defensin Antibacterial | ||
| Overall_6716|Bagel_I_54|54.1|LichenicidinVK21A1 | 3 (1) | 0.028 | Bagel_I | Lantibiotic alpha Antibacterial |
| Overall_7055|Bagel_III_7|7.3|Bacteriocinalbusin_B | 4 (1) | 0.038 | Bagel_III | Bacteriocin Antibacterial G+ |
| Overall_32056|UniProtKb_2305|A1A547;E9QMH8;Q6R1Z2|PGRP3_MOUSE | 188 (1) | 0.038 | UniProtKB | Peptidoglycan recognition protein 3 (N-acetylmuramoyl-l-alanine amidase) Antibacterial G+ |
| Overall_4110|APD_2229|AP00908|Dermatoxin DA1 | 194 (1) | 0.037 | APD | Antibacterial peptide Bactericidal towards mollicutes (wall-less eubacteria) and Gram-positive eubacteria |
| Overall_4109|APD_2228|AP00907|Dermatoxin A1 | APD | Antibacterial peptide Bactericidal towards mollicutes (wall-less eubacteria) and Gram-positive eubacteria | ||
| Overall_496|AMPer_496|DMS1_PACDA|Dermaseptin PD-1-5 precursor | AMPer | Frog skin active peptide AMPs with a large spectrum of activities | ||
| Overall_495|AMPer_495|DMS1_AGAAN|Dermaseptin AA-1-1 precursor | AMPer | Frog skin active peptide AMPs with a large spectrum of activities | ||
| Overall_2680|APD_799|AP01717|Esculentin-2PRa | 195 (1) | 0.011 | APD | Antibacterial peptide Antibacterial |
| Overall_3280|APD_1399|AP02257|Lysozyme | 196 (1) | 4.469e-05 | APD | C-type lysozyme/alpha-lactalbumin family (Kinocidin) Antibacterial G+/-, Antifungal, Antiparasitic, Chemotactic |
| Overall_31618|UniProtKb_1867|P79294|LYSC_SAISC | UniProtKB | C-type lysozyme/alpha-lactalbumin family Bacteriolytic function (defence response to bacterium) | ||
| Overall_31613|UniProtKb_1862|P79239|LYSC_PONPY | UniProtKB | C-type lysozyme/alpha-lactalbumin family Bacteriolytic function (defence response to bacterium) | ||
| Overall_31593|UniProtKb_1842|P79180|LYSC_HYLLA | UniProtKB | C-type lysozyme/alpha-lactalbumin family Bacteriolytic function (defence response to bacterium) | ||
| Overall_31590|UniProtKb_1839|P79179|LYSC_GORGO | UniProtKB | C-type lysozyme/alpha-lactalbumin family Bacteriolytic function (defence response to bacterium) | ||
| Overall_31592|UniProtKb_1841| | UniProtKB | C-type lysozyme/alpha-lactalbumin family Bacteriolytic function (defence response to bacterium) | ||
| Overall_4269|APD_2388|AP02388|BPTI | 197 (1) | 4.185e-276 | APD | Bovine Pancreatic Trypsin Inhibitor Antibacterial |
| Overall_1040|AMSDb_52|APRFR_BOVIN|APROTININ ANTIBACTERIAL FRAGMENTS | AMSDb | Bovine Pancreatic Trypsin Inhibitor Antibacterial | ||
| Overall_6268|AVPdb_1892|AVP1895|Coronaviridae | 198 (1) | 0.012 | AVP | Coronavirus S2 glycoprotein Antiviral |
| Overall_6267|AVPdb_1891|AVP1894|Coronaviridae | AVP | Coronavirus S2 glycoprotein Antiviral | ||
| Overall_12077|CAMP_Validated_2538|CAMPSQ940|Defensin J1-1 | 206 (1) | 5.523e-12 | CAMP | Gamma-thionin family: Defensin Antibacterial and Antifungal |
| Overall_30115|UniProtKb_364|Q1KLX1|CAMP_PANTR | 249 (2) | 0.001 | UniProtKB | Cathelicidin AMP Antibacterial G+/– |
| Overall_546|AMPer_546|FAL39_HUMAN|Antibacterial protein FALL-39 precursor (FALL-39 peptide antibiotic); (Cationic antimicrobial protein CAP-18) (hCAP-18) [Contains:;Antibacterial protein LL-37] | AMPer | Cathelicidin Cathionic AMP Antibacterial G+/– | ||
| Overall_17064|DB|DBAASP_1120|1242|Cathelicidin antimicrobial peptide | DBAASP | Cathelicidin Cathionic AMP Antibacterial | ||
| Overall_21373|DBAASP_5429|5781|cgUbiquitin | 254 (2) | 0 | DBAASP | Ubiquitin family Immune response and inflammation and viral infection |
| Overall_3029|APD_1148|AP02030|cgUbiquitin | APD | Ubiquitin family Immune response and inflammation and viral infection | ||
| Overall_31667|UniProtKb_1916|O80288|LYS_BPPS3 | 290 (1) | 2.899e-60 | UniProtKB | Lysozyme Defence response to bacterium |
| Overall_31249|UniProtKb_1498|Q8CFB4;E9QJR4;Q8CFA4|GBP5_MOUSE | 305 (1) | 0.014 | UniProtKB | Lysozyme C Bacteriolytic function (defence response to bacterium) |
| Overall_5857|AVPdb_1481|AVP1484|Coronaviridae | 314 (1) | 0.006 | AVP | Synthetic peptide targeting critical sites on the SARS-associated coronavirus spike protein responsible for viral infection Antiviral |
| Overall_8456|CAMP_Patent_1315|Sequence 1188|US 6573361 | 317 (1) | 0.038 | CAMP | Guanylate-binding protein Response to bacterium and inflammatory process |
| Overall_14669|DADP_2528|SP_2629|Ranalexin-1Vb Ranalexin-Vb | 318 (1) | 0.001 | DADP | Frog antimicrobial peptide Antibacterial |
| Overall_12734|DADP_593|SP_C0ILB1|Nigroain-I | 319 (1) | 0.001 | DADP | Frog antimicrobial peptide Antibacterial |
| Overall_13517|DADP_1376|SP_E7EKC8|Hainanensin-1_2 | 320 (1) | 0.004 | DADP | Frog antimicrobial peptide Antibacterial |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matos, A.; Domínguez-Pérez, D.; Almeida, D.; Agüero-Chapin, G.; Campos, A.; Osório, H.; Vasconcelos, V.; Antunes, A. Shotgun Proteomics of Ascidians Tunic Gives New Insights on Host–Microbe Interactions by Revealing Diverse Antimicrobial Peptides. Mar. Drugs 2020, 18, 362. https://doi.org/10.3390/md18070362
Matos A, Domínguez-Pérez D, Almeida D, Agüero-Chapin G, Campos A, Osório H, Vasconcelos V, Antunes A. Shotgun Proteomics of Ascidians Tunic Gives New Insights on Host–Microbe Interactions by Revealing Diverse Antimicrobial Peptides. Marine Drugs. 2020; 18(7):362. https://doi.org/10.3390/md18070362
Chicago/Turabian StyleMatos, Ana, Dany Domínguez-Pérez, Daniela Almeida, Guillermin Agüero-Chapin, Alexandre Campos, Hugo Osório, Vitor Vasconcelos, and Agostinho Antunes. 2020. "Shotgun Proteomics of Ascidians Tunic Gives New Insights on Host–Microbe Interactions by Revealing Diverse Antimicrobial Peptides" Marine Drugs 18, no. 7: 362. https://doi.org/10.3390/md18070362