Metabolomic Investigations of American Oysters Using 1H-NMR Spectroscopy

Abstract
:1. Introduction
2. Results and Discussion
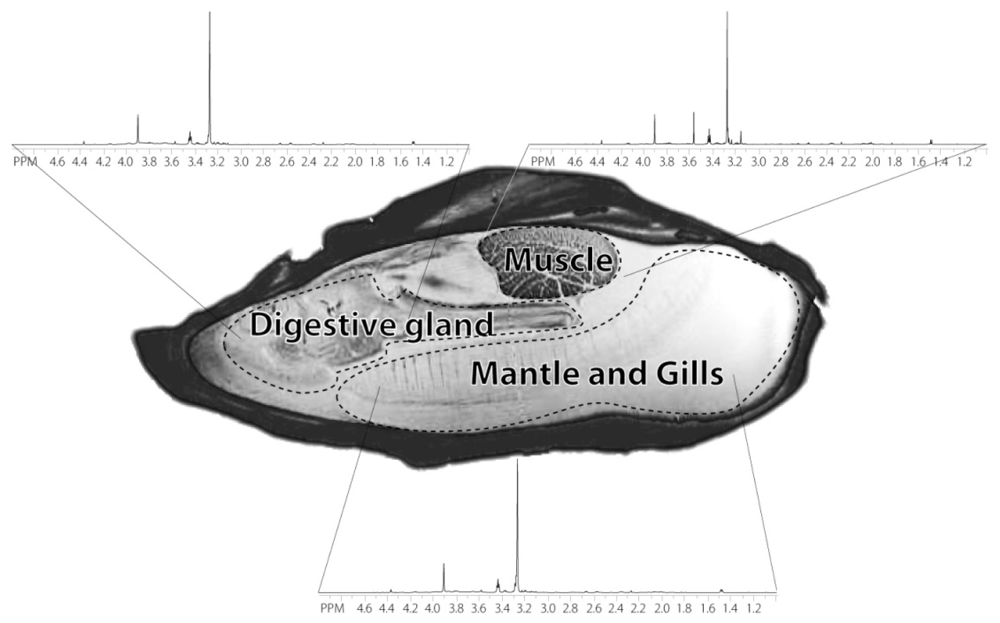
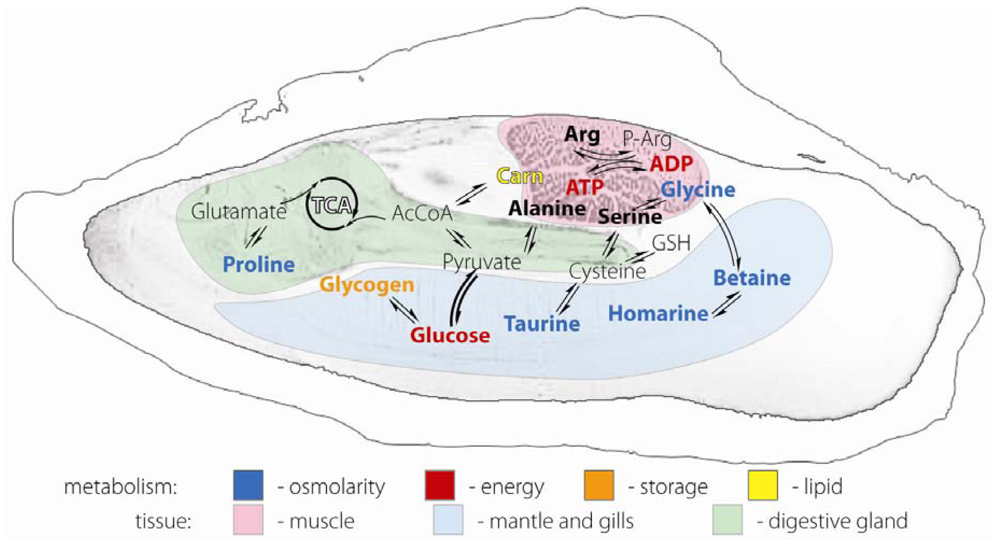
2.1. Anatomical definition of oyster body parts
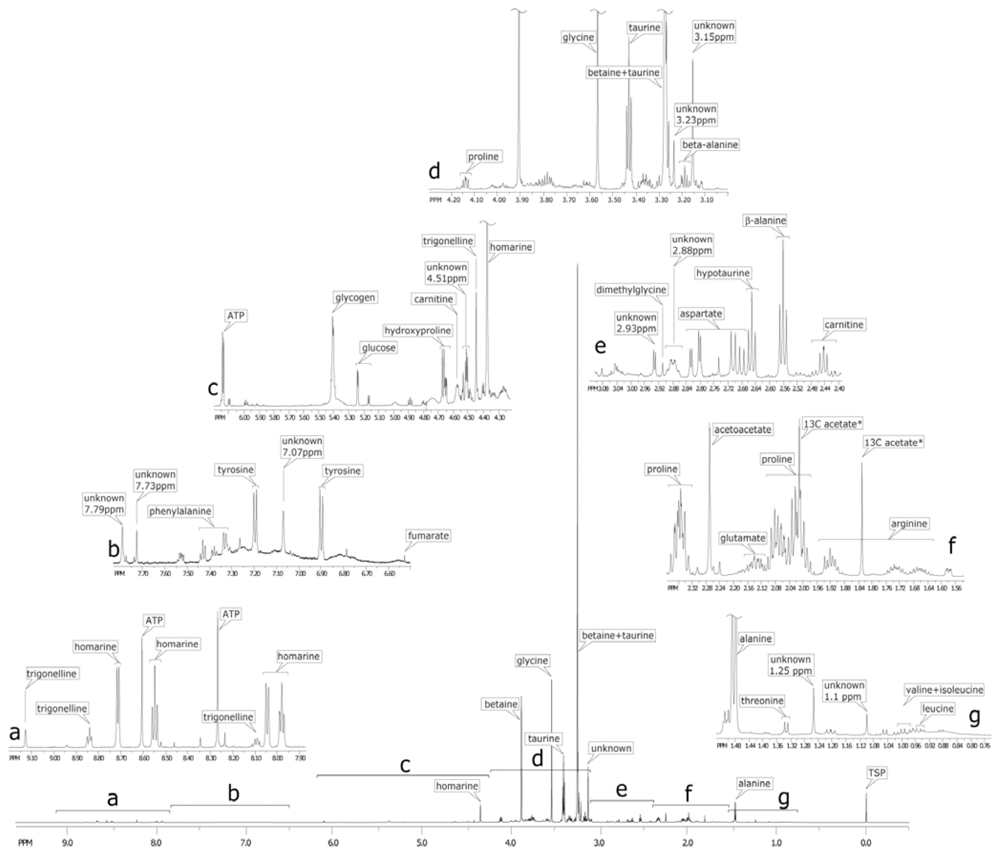
2.2. Identification of the metabolites on 1D 1H spectrum
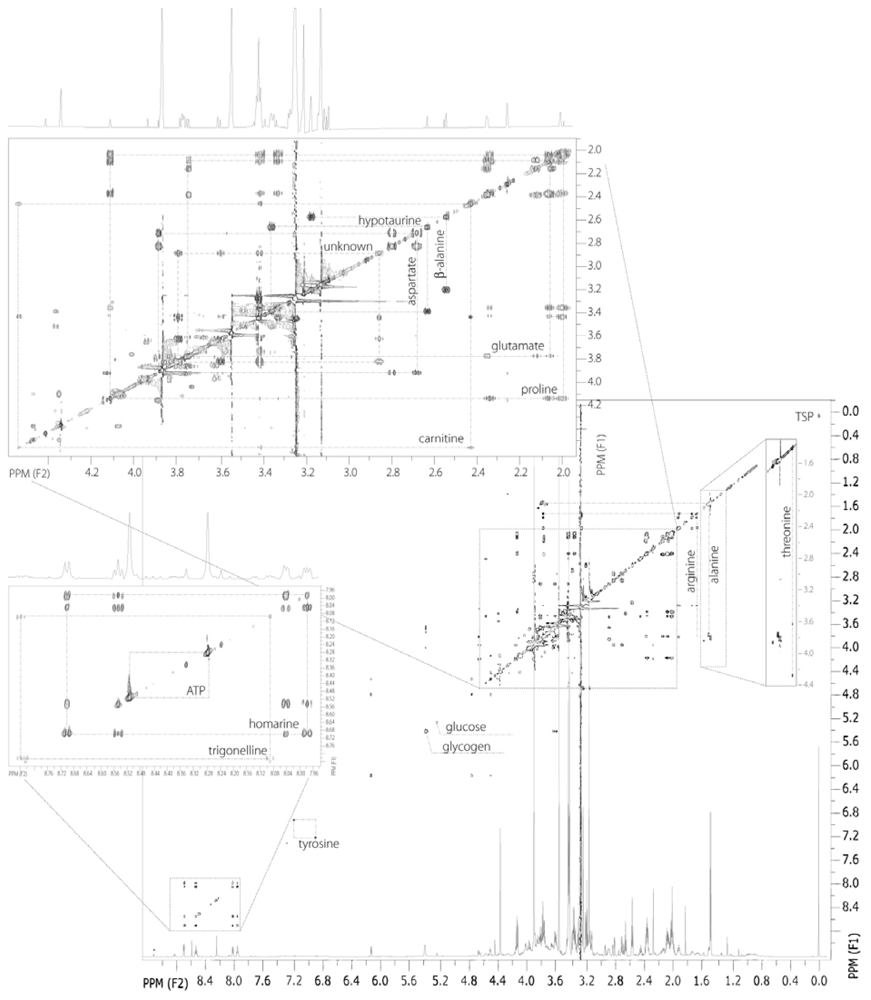
2.3. 2D 1H NMR spectra of oyster muscle
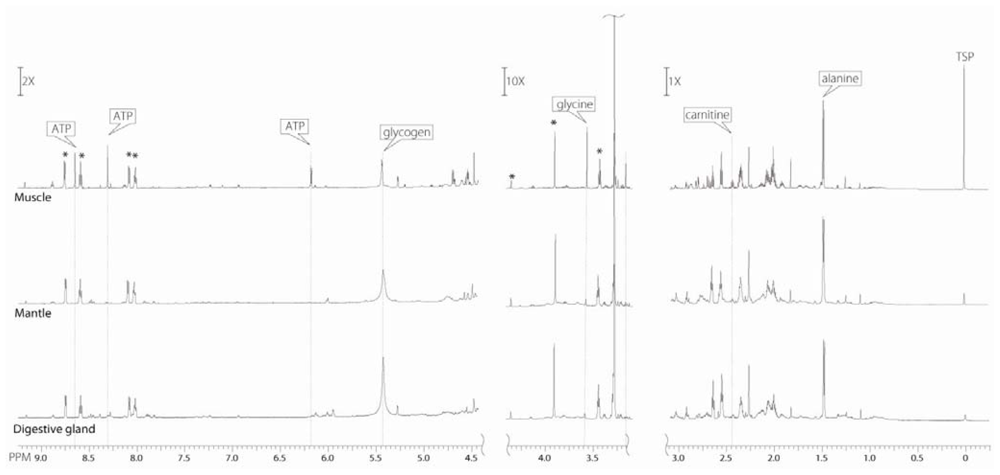
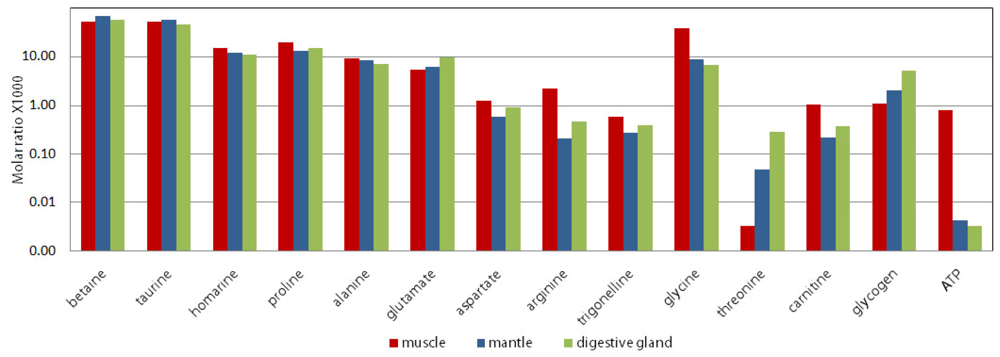
2.4. Differences in metabolites from the three oyster organ blocks
3. Experimental
4. Conclusions
Acknowledgements
- Samples Availability: Available from the authors.
References
- Hines, A; Oladiran, GS; Bignell, JP; Stentiford, GD; Viant, MR. Direct sampling of organisms from the field and knowledge of their phenotype: Key recommendations for environmental metabolomics. Environ. Sci. Technol 2007, 41, 3375–3381. [Google Scholar]
- Jones, OAH; Spurgeon, DJ; Svendsen, C; Griffin, JL. A metabolomics based approach to assessing the toxicity of the polyaromatic hydrocarbon pyrene to the earthworm Lumbricus rubellus. Chemosphere 2008, 71, 601–609. [Google Scholar]
- Tuffnail, W; Mills, GA; Cary, P; Greenwood, R. An environmental 1H NMR metabolomic study of the exposure of the marine mussel Mytilus edulis to atrazine, lindane, hypoxia and starvation. Metabolomics 2009, 5, 33–43. [Google Scholar]
- Viant, MR; Rosenblum, ES; Tjeerdema, RS. NMR-based metabolomics: a powerful approach for characterizing the effects of environmental stressors on organism health. Environ. Sci. Technol 2003, 37, 4982–4989. [Google Scholar]
- Willard, HH; Merritt, LL, Jr; Dean, JA; Settle, FA, Jr. Belmont, CA, Ed.; Instrumental Methods of Analysis, 7th ed; Wadsworth: Belmont, CA, USA, 1988. [Google Scholar]
- Fan, TWM. Metabolite profiling by one- and two-dimensional NMR analysis of complex mixtures. Prog. Nucl. Magn. Reson. Spectrosc 1996, 28, 161–219. [Google Scholar]
- Fan, TWM; Lane, AN. Structure-based profiling of metabolites and isotopomers by NMR. Prog. Nucl. Magn. Reson. Spectrosc 2008, 52, 69–117. [Google Scholar]
- Sabatini, P; Josupeit, H. Bivalves: Commodity Update; FAO GLOBEFISH: Rome, Italy, 2007. [Google Scholar]
- Allen, K. Amino acids in the Mollusca. Integr. Comp. Biol 1961, 1, 253–261. [Google Scholar]
- Allen, K. The effect of salinity on the amino acid concentration in Rangia cuneata (Pelecypoda). Biol. Bull 1961, 121, 419–424. [Google Scholar]
- Burg, MB; Ferraris, JD. Intracellular organic osmolytes: function and regulation. J. Biol. Chem 2008, 283, 7309–7313. [Google Scholar]
- Collicutt, JM; Hochachka, PW. The anaerobic oyster heart: coupling of glucose and aspartate fermentation. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol 1977, 115, 147–157. [Google Scholar]
- Hammen, CS. Metabolism of the oyster, Crassostrea virginica. Integr. Comp. Biol 1969, 9, 309–318. [Google Scholar]
- Hammen, CS; Miller, HF. Nitrogen excretion of Crassostrea virginica. Comp. Biochem. Physiol 1966, 17, 1199–1200. [Google Scholar]
- Ringwood, AH; Conners, DE; DiNovo, A. The effects of copper exposures on cellular responses in oysters. Mar. Environ. Res 1998, 46, 591–595. [Google Scholar]
- Fan, TWM; Higashi, RM; Macdonald, JM. Emergence and recovery response of phosphate metabolites and intracellular pH in intact Mytilus edulis as examined in situ by in vivo 31P-NMR. Biochim. Biophys. Acta 1991, 1092, 39–47. [Google Scholar]
- Higashi, RM; Fan, TWM; Macdonald, JM. Monitoring of metabolic responses of intact Haliotis (abalones) under salinity stress by 31P surface probe localized NMR. J. Exp. Zool 1989, 249, 350–356. [Google Scholar]
- Harris, JM. The presence, nature, and role of gut microflora in aquatic invertebrates: a synthesis. Microb. Ecol 1993, 25, 195–231. [Google Scholar]
- Kueh, CSW; Chan, K-y. Bacteria in bivalve shellfish with special reference to the oyster. J. Appl. Microbiol 1985, 59, 41–47. [Google Scholar]
- Ballantyne, JS; Moyes, CD. The effects of salinity acclimation on the osmotic properties of mitochondria from the gill of Crassostrea virginica. J. Exp. Biol 1987, 133, 449. [Google Scholar]
- Powell, EN; Kasschau, M; Chen, E; Koenig, M; Pecon, J. Changes in the free amino acid pool during environmental stress in the gill tissue of the oyster, Crassostrea virginica. Comp. Biochem. Physiol. A Physiol 1982, 71, 591–598. [Google Scholar]
- Carr, WES; Netherton, JC, III; Gleeson, RA; Derby, CD. Stimulants of feeding behavior in fish: analyses of tissues of diverse marine organisms. Biol. Bull 1996, 190, 149–160. [Google Scholar]
- Henry, RP; Mangum, CP. Salt and water balance in the oligohaline clam, Rangia cuneata III. Reduction of the free amino acid pool during low salinity adaptation. J. Exp. Zool 1980, 211, 25–32. [Google Scholar]
- Henry, RP; Mangum, CP; Webb, KL. Salt and water balance in the oligohaline clam, Rangia cuneata II. Accumulation of intracellular free amino acids during high salinity adaptation. J. Exp. Zool 1980, 211, 11–24. [Google Scholar]
- Lynch, MP; Wood, L. Effects of environmental salinity of free amino acids of Crassostrea virginica gmelin. Comp. Biochem. Physiol 1966, 19, 783–790. [Google Scholar]
- Sidwell, VD; Loomis, AL; Grodner, RM. Geographic and monthly variation in composition of oysters. Mar. Fish Rev 1979, 41, 13–17. [Google Scholar]
- Pierce, SK; Rowland-Faux, LM; O’Brien, SM. Different salinity tolerance mechanisms in Atlantic and Chesapeake Bay conspecific oyster: glycine betaine and amino acid pool variations. Mar. Biol 1992, 113, 107–115. [Google Scholar]
- Foreman, RA, III; Ellington, WR. Effects of inhibitors and substrate supplementation on anaerobic energy metabolism in the ventricle of the oyster, Crassostrea virginica. Comp. Biochem. Physiol. B 1983, 74, 543–547. [Google Scholar]
- Bremer, J. Carnitine-metabolism and functions. Physiol. Rev 1983, 63, 1420–1480. [Google Scholar]
- Bremer, J. Carnitine in intermediary metabolism. J. Biol. Chem 1963, 238, 2774–2779. [Google Scholar]
- Fritz, IB. Carnitine and its role in fatty acid metabolism. Adv. Lipid. Res 1963, 1, 285–334. [Google Scholar]
- Conners, DE; Ringwood, AH. Effects of glutathione depletion on copper cytotoxicity in oysters (Crassostrea virginica). Aquat. Toxicol 2000, 50, 341–349. [Google Scholar]
- Lamoureux, GL; Rusness, DG; Dolphin, D; Poulson, R; Avramovic, O. Dolphin, D, Poulson, R, Avramovic, O, Eds.; The role of glutathione and glutathione-S-transferases in pesticide metabolism, selectivity, and mode of action in plants and insects. In Glutathione: Chemical, Biochemical and Medical Aspects; John Wiley & Sons: New York, NY, USA, 1989; pp. 153–196. [Google Scholar]
- Ackermann, D. Uber das vorkommen von homarin, trigonellin und einer neuen base anemonin in der anthozoe anemonia-sulcata. Hoppe-Seyler’s Z. Physiol. Chem 1953, 295, 1–9. [Google Scholar]
- Yancey, PH; Clark, ME; Hand, SC; Bowlus, RD; Somero, GN. Living with water stress: evolution of osmolyte systems. Science 1982, 217, 1214–1222. [Google Scholar]
- Hu, YH; Lin, CL; Huang, YW; Liu, PE; Hwang, DF. Dietary amino acid taurine ameliorates liver injury in chronic hepatitis patients. Amino Acids 2008, 35, 469–473. [Google Scholar]
- Lee, N-H; Han, K-N; Choi, K-S. Effects of salinity and turbidity on the free amino acid composition in gill tissue of the Pacific oyster Crassostrea gigas. J. Shellfish Res 2004, 23, 129–133. [Google Scholar]
- Schuller-Levis, GB; Park, E. Taurine: new implications for an old amino acid. FEMS Microbiol. Lett 2003, 226, 195–202. [Google Scholar]
- Mendel, LB; Bradley, HC. Experimental studies on the physiology of the mollusks-third paper. Am. J. Physiol 1906, 17, 167–176. [Google Scholar]
- Karsten, H. Disquisitio microscopia et chemica hepatis et bilis crustaceorum et molluscorum. Nova acta Academiae Caesareae Leopoldino-Carolinae Germanicae Naturae Curiosorum 1845, 21, 295–326. [Google Scholar]
- Markwell, PJ; Earle, KE. Taurine: an essential nutrient for the cat. A brief review of the biochemistry of its requirement and the clinical consequences of deficiency. Nutr. Res 1995, 15, 53–58. [Google Scholar]
- Bouckenooghe, T; Remacle, C; Reusens, B. Is taurine a functional nutrient? Curr. Opin. Clin. Nutr. Metab. Care 2006, 9, 728–733. [Google Scholar]
- Toyohara, H; Ikeda, M; Goto, C; Sawada, H; Hosoi, M; Takeuchi, K; Hayashi, I; Imamura, S; Yamashita, M. Osmo-responsive expression of oyster amino acid transporter gene and analysis of the regulatory region involved. Fish. Sci 2005, 71, 465–470. [Google Scholar]
- Somero, GN; Bowlus, RD. Hochachka, PW, Ed.; Osmolytes and metabolic end products of molluscs: the design of compatible solute systems. In The Mollusca. Environmental Biochemistry and Physiology; Academic Press: New York; NY, USA, 1988; pp. 77–100. [Google Scholar]
- Affeld, S; Wagele, H; Avila, C; Kehraus, S; Konig, GM. Distribution of homarine in some Opisthobranchia (Gastropoda: Mollusca). Bonner Zoologische Beitrage 2006, 55, 181–190. [Google Scholar]
- Gasteiger, EL; Gergen, JA; Haake, PC. A study of the distribution of homarine (N-methyl picolinic acid). Biol. Bull 1955, 109, 345–346. [Google Scholar]
- Polychronopoulos, P; Magiatis, P; Skaltsounis, AL; Tillequin, F; Vardala-Theodorou, E; Tsarbopoulos, A. Homarine, a common metabolite in edible Mediterranean molluscs: Occurrence, spectral data and revision of a related structure. Nat. Prod. Res 2001, 15, 411–418. [Google Scholar]
- Linehan, LG; O’Connor, TP; Burnell, G. Seasonal variation in the chemical composition and fatty acid profile of Pacific oysters (Crassostrea gigas). Food Chem 1999, 64, 211–214. [Google Scholar]
- Berthelin, C; Kellner, K; Mathieu, M. Storage metabolism in the Pacific oyster (Crassostrea gigas) in relation to summer mortalities and reproductive cycle (West Coast of France). Comp. Biochem. Physiol. B, Biochem. Mol. Biol 2000, 125, 359–369. [Google Scholar]
- Lenoir, F; Robbins, I; Mathieu, M; Lubet, P; Gabbott, PA. Isolation, characterization and glucose metabolism of glycogen cells (=vesicular connective-tissue cells) from the labial palps of the marine mussel Mytilus edulis. Mar. Biol 1989, 101, 495–501. [Google Scholar]
- Gabbott, PA. Barnes, HB, Ed.; Storage cycles in marine bivalve molluscs: a hypothesis concerning the relationship between glycogen metabolism and gametogenesis. In Ninth European Marine Biology Symposium; Aberdeen University Press: Aberdeen, UK, 1975; pp. 191–211. [Google Scholar]
- Goddard, CK; Martin, AW. Wilbur, KM, Yonge, CM, Eds.; Carbohydrate metabolism. In Physiology of Mollusca; Academic Press: New York, NY, USA, 1966; Volume 2, pp. 275–308. [Google Scholar]
- Walne, PR. The seasonal variation of meat and glycogen content of seven populations of Oysters Ostrea edulis L. and a review of the literature. Fish. Invest. Lond. Ser. II 1970, 26, 1–35. [Google Scholar]
- Deslous-Paoli, JM; Heral, M. Biochemical composition and energy value of Crassostrea gigas (Thunberg) cultured in the bay of Marennes-Oleron. Aquat. Living Resour 1988, 1, 239–249. [Google Scholar]
- Patrick, S; Faury, N; Goulletquer, P. Seasonal changes in carbohydrate metabolism and its relationship with summer mortality of Pacific oyster Crassostrea gigas (Thunberg) in Marennes-Oleron bay (France). Aquaculture 2006, 252, 328–338. [Google Scholar]
- Perdue, JA; Beattie, JH; Chew, KK. Some relationships between gametogenic cycle and summer mortality phenomenon in the Pacific oyster (Crassostrea gigas) in Washington State. J. Shellfish Res 1981, 1, 9–16. [Google Scholar]
- Maurer, D; Borel, M. Croissance, engraissement et cycle sexuel de Crassostrea gigas dans le Bassin d’Arachon: comparaison des huitres agees de 1 et 2 ans. Haliotis 1985, 15, 125–134. [Google Scholar]
- Mathieu, M; Lubet, P. Storage tissue metabolism and reproduction in marine bivalves: a brief review. Invertebr. Reprod. Dev 1993, 23, 123–129. [Google Scholar]
- Ruiz, C; Abad, M; Sedano, F; Garcia-Martin, LO; Sanchez Lopez, JL. Influence of seasonal environmental changes on the gamete production and biochemical composition of Crassostrea gigas (Thunberg) in suspended culture in El Grove, Galicia, Spain. J. Exp. Mar. Biol. Ecol 1992, 155, 249–262. [Google Scholar]
- Gabbott, PA; Whittle, MA. Glycogen synthetase in the sea mussel Mytilus edulis L.—II. Seasonal changes in glycogen content and glycogen synthetase activity in the mantle tissue. Comp. Biochem. Physiol. B, Biochem. Mol. Biol 1986, 83, 197–207. [Google Scholar]
- Berthelin, CH; Fievet, B; Leclerc, G; Germain, P; Kellner, K; Mathieu, M. In vivo and in vitro approaches to the analysis of glycogen metabolism in the Pacific oyster, Crassostrea gigas. J. Shellfish Res 2003, 22, 715–720. [Google Scholar]
- Fando, JJ; Garcia-Fernandez, MC; Candela, JL. Glycogen metabolism in Ostrea edulis (L.)— factors affecting glycogen synthesis. Comp. Biochem. Physiol. B 1972, 43, 807–814. [Google Scholar]
- Wylie, V; Smith, M. Nucleotides of the Pacific oyster Crassostrea gigas (Thunberg). Biochem. Cell Biol 1964, 42, 1347–1351. [Google Scholar]
- Michaelidis, B; Haas, D; Grieshaber, MK. Extracellular and intracellular acid base status with regard to the energy metabolism in the oyster Crassostrea gigas during exposure to air. Physiol. Biochem. Zool 2005, 78, 373–383. [Google Scholar]
- Yokoyama, Y; Sakaguchi, M; Kawai, F; Kanamori, M. Changes in concentration of ATP-related compounds in various tissues of oyster during ice storage. Nippon Suisan Gakk 1992, 58, 2125–2136. [Google Scholar]
- Van Pilsum, JF; Stephens, GC; Taylor, D. Distribution of creatine, guanidinoacetate and the enzymes for their biosynthesis in the animal kingdom. Biochem. J 1972, 126, 325–345. [Google Scholar]
- Nakada, HI; Friedmann, B; Weinhouse, S. Pathways of glycine catabolism in rat liver. J. Biol. Chem 1955, 216, 583–592. [Google Scholar]
- Lehninger, AL; Nelson, DL; Cox, MM. Lehninger Principles of Biochemistry; W.H. Freeman: New York, NY, USA, 2005. [Google Scholar]
- Stryer, L. Biochemistry; W.H. Freeman: New York, NY, USA, 1995; pp. 86–88. [Google Scholar]
- Dewar, BJ; Keshari, K; Jeffries, R; Dzeja, P; Graves, LM; Macdonald, JM. Metabolic assessment of a novel chronic myelogenous leukemic cell line and an imatinib resistant subline by 1H NMR spectroscopy. Metabolomics 2010, 6, 439–450. [Google Scholar]
- Fan, TWM; Colmer, TD; Lane, AN; Higashi, RM. Determination of metabolites by 1H NMR and GC: analysis for organic osmolytes in crude tissue extracts. Anal. Biochem 1993, 214, 260–271. [Google Scholar]
- Fan, TWM; Higashi, RM; Lane, AN; Jardetzky, O. Combined use of 1H-NMR and GC-MS for metabolite monitoring and in vivo 1H-NMR assignments. Biochim. Biophys. Acta 1986, 882, 154–167. [Google Scholar]
- Keshari, KR; Kurhanewicz, J; Jeffries, RE; Wilson, DM; Dewar, BJ; Van Criekinge, M; Zierhut, M; Vigneron, DB; Macdonald, JM. Hyperpolarized 13C spectroscopy and an NMR-compatible bioreactor system for the investigation of real-time cellular metabolism. Magn. Reson. Med 2010, 63, 322–329. [Google Scholar]
- Lee, H; Tikunov, A; Stoskopf, MK; Macdonald, JM. Application of chemical shift imaging to marine sciences. Mar. Drugs 2010, 8, 2369–2383. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolites | Chemical shift and peak shape, ppm | Concentration | Reference |
|---|---|---|---|
| Amino acids | |||
| Alanine | 1.46 (d), 3.76 (m) | 1.30 ± 0.27 * | [21] |
| Arginine | 1.68 (m), 1.90 (m), 3.23 (t), 3.74 (t) | 0.16–0.24 † | [25] |
| Aspartate | 2.66 (dd), 2.79(dd), 3.87 (dd) | 2.59 ± 0.72 * | [21] |
| Glutamate | 2.08 (m), 2.34 (m), 3.74 (t) | 1.30 ± 0.19 * | [21] |
| Glutamine | 2.12 (m), 2.44 (m), 3.75 (t) | 0.53 ± 0.13 * | [21] |
| Glycine | 3.54 (s) | 1.29 ± 0.29 * | [21] |
| Histidine | 7.81 (s) 7.04 (s), 3.99 (dd), 3.24/3.15 (dd) | 0.089–0.255 † | [25] |
| Isoleucine | 0.92 (t), 1.00 (d), 1.26(m), 1.44 (m), 1.96 (m), 3.66 (d) | 0.001–0.008 † | [25] |
| Leucine | 0.94 (d), 0.96 (d), 1.66 (m), 3.71 (t) | 0.005–0.014 † | [25] |
| Phenylalanine | 3.98 (m), 7.31 (d), 7.36 (t), 7.41 (m) | 0.006–0.033 † | [25] |
| Proline | 1.99 (m), 2.06 (m), 2.34 (m), 3.33 (dt), 3.41 (dt), 4.12 (dd) | 0.020–0.582 † | [25] |
| Serine | 3.84 (m), 3.96 (m) | 0.023–0.057 † | [25] |
| Threonine | 1.33 (d), 3.578 (d), 4.25 (m) | 0.009–0.036 † | [25] |
| Tyrosine | 6.89 (d), 7.19 (d) | 0.005–0.046 † | [25] |
| Valine | 0.98 (d), 1.03 (d), 2.25 (m), 3.59 (d) | 0.004–0.018 † | [25] |
| Energy related | |||
| α-Glucose | 3.23 (dd), 3.40 (m), 3.46 (m), 3.52 (dd), 3.73 (m), 3.82 (m), 3.88 (dd), 4.63 (d), 5.22 (d) | n/a | - |
| ß-Glucose | 4.64 (d) | n/a | - |
| Glycogen | 3.40 (m), 3.60 (m), 3.80 (m), 3.96 (br. s.), 5.40 (br. s.) | 0.467–6.920 ‡ | [26] |
| ADP | 4.15 (m), 4.16 (m), 4.57 (m), 5.94 (m), 8.29 (s), 8.54 (s) | n/a | - |
| ATP | 4.21 (m), 4.28 (m), 4.39 (m), 4.51 (m), 4.62 (t), 6.13 (d), 8.24 (s), 8.53(s) | n/a | - |
| Osmolytes/Organic acids | |||
| Betaine | 3.25 (s), 3.89 (s) | 230 ± 30 □ | [27] |
| ß-alanine | 2.55 (t), 3.18 (t) | 0.25 ± 0.13 * | [21] |
| Homarine | 4.35 (s), 7.95 (dd), 8.02 (d), 8.53 (dd), 8.71 (d) | n/a | - |
| Hypotaurine | 2.64 (t), 3.36 (t) | n/a | - |
| Taurine | 3.25 (s), 3.41 (t) | 16.37 ± 3.39 * | [21] |
| Krebs cycle intermediates | |||
| Succinate | 2.41 (s) | 0.733 ± 0.288 □ | [28] |
| Fatty Acid Metabolism | |||
| Acetoacetate | 2.22 (s), 3.41 (m) | n/a | - |
| Carnitine | 2.43 (dd), 3.21 (s), 3.42 (m), 4.56 (s) | n/a | - |
| Alkaloids | |||
| Trigonelline | 4.43 ppm (s), 8.08 ppm (t), 8.84 ppm (t), 9.12 ppm (s) | n/a | - |
| Unknown Resonances | |||
| Unknown #1 | 1.1 ppm (s) | ||
| Unknown #2 | 1.25 ppm (s) | ||
| Unknown #3 | 2.88 (m)(Jab = 7.67 Hz, Jbc = 7.75), 3.44 (m), 3.62 (m)(Jab = 7.63 Hz, Jbc = 11.12), 3.82 (m) | ||
| Unknown #4 | 2.93 (d) or two (s) | ||
| Unknown #5 | 3.15 ppm (s) | ||
| Unknown #6 | 3.23 ppm (s) | ||
| Unknown #7 | 4.51 (t) (Jab = 4.41 Hz) | ||
| Unknown #8 | 7.73 ppm (s) | ||
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tikunov, A.P.; Johnson, C.B.; Lee, H.; Stoskopf, M.K.; Macdonald, J.M. Metabolomic Investigations of American Oysters Using 1H-NMR Spectroscopy. Mar. Drugs 2010, 8, 2578-2596. https://doi.org/10.3390/md8102578
Tikunov AP, Johnson CB, Lee H, Stoskopf MK, Macdonald JM. Metabolomic Investigations of American Oysters Using 1H-NMR Spectroscopy. Marine Drugs. 2010; 8(10):2578-2596. https://doi.org/10.3390/md8102578
Chicago/Turabian StyleTikunov, Andrey P., Christopher B. Johnson, Haakil Lee, Michael K. Stoskopf, and Jeffrey M. Macdonald. 2010. "Metabolomic Investigations of American Oysters Using 1H-NMR Spectroscopy" Marine Drugs 8, no. 10: 2578-2596. https://doi.org/10.3390/md8102578