Antioxidant Effect of a Marine Oligopeptide Preparation from Chum Salmon (Oncorhynchus keta) by Enzymatic Hydrolysis in Radiation Injured Mice


Abstract
:1. Introduction
2. Results and Discussion
2.1. Characterization of MOP
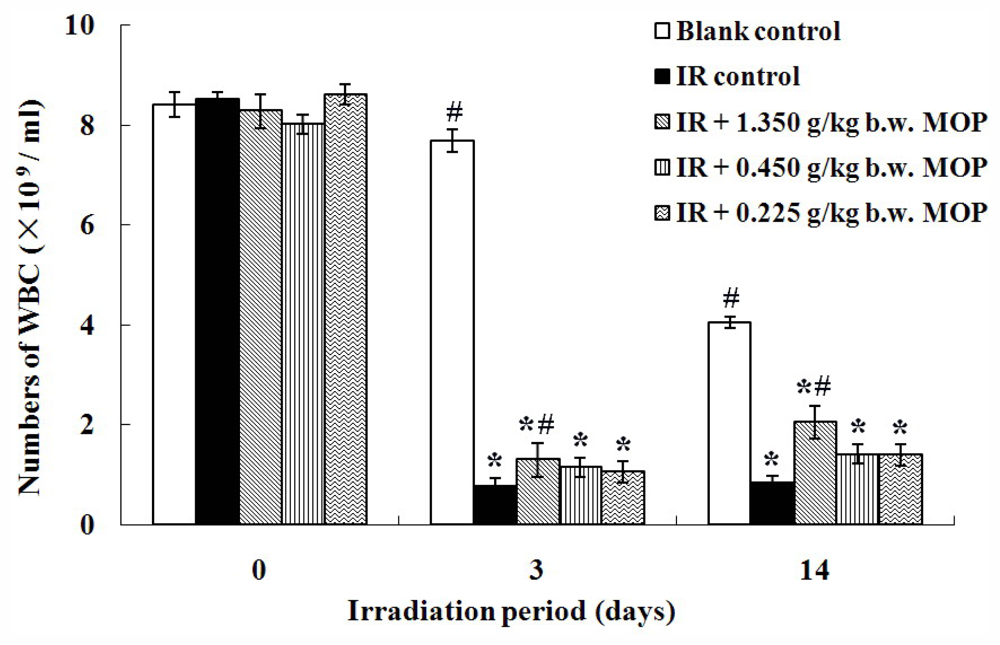
2.2. Effect of MOP on Irradiation-Reduced White Blood Cells Count
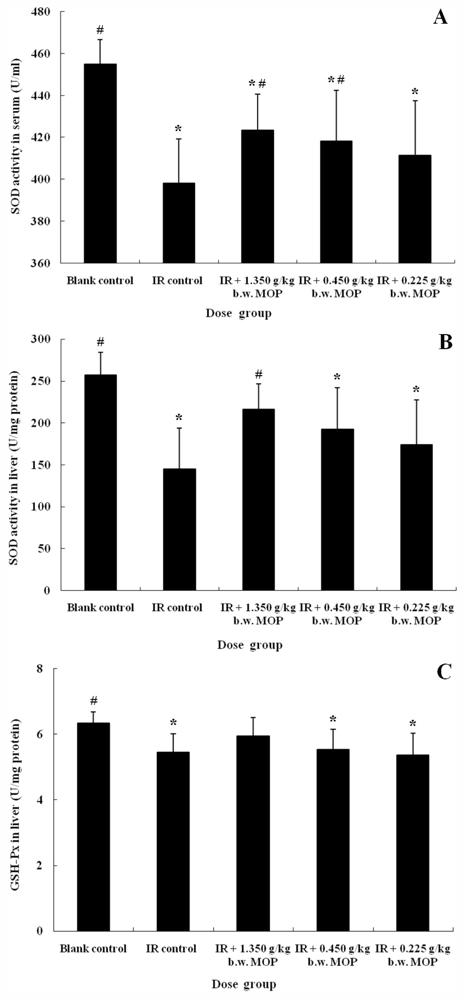
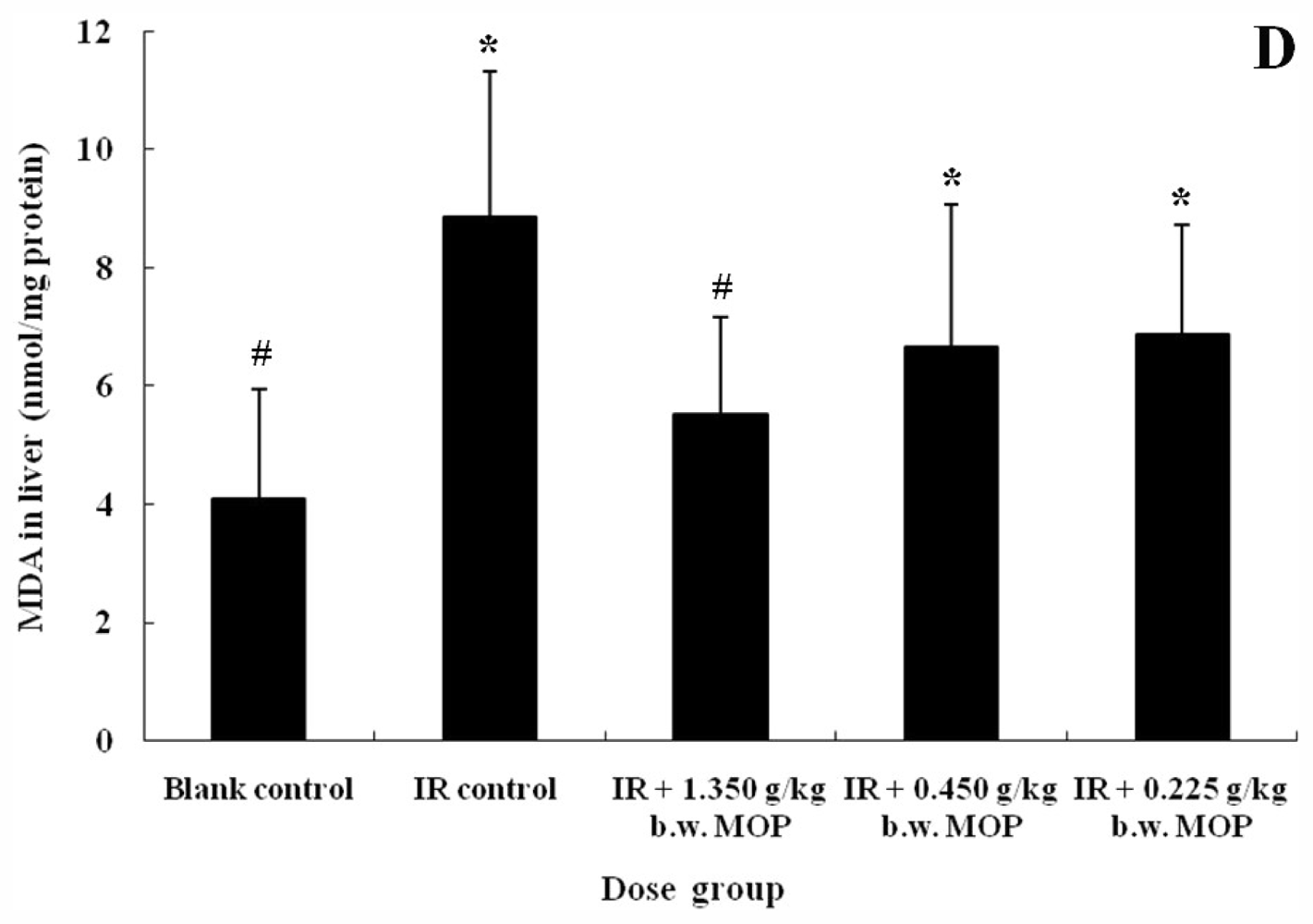
2.3. Effect of MOP on Superoxide Dismutase (SOD) Activity in Serum and Liver, Glutathione Peroxidase (GSH-Px) Activity and Malondialdehyde (MDA) Level in Liver after WBI
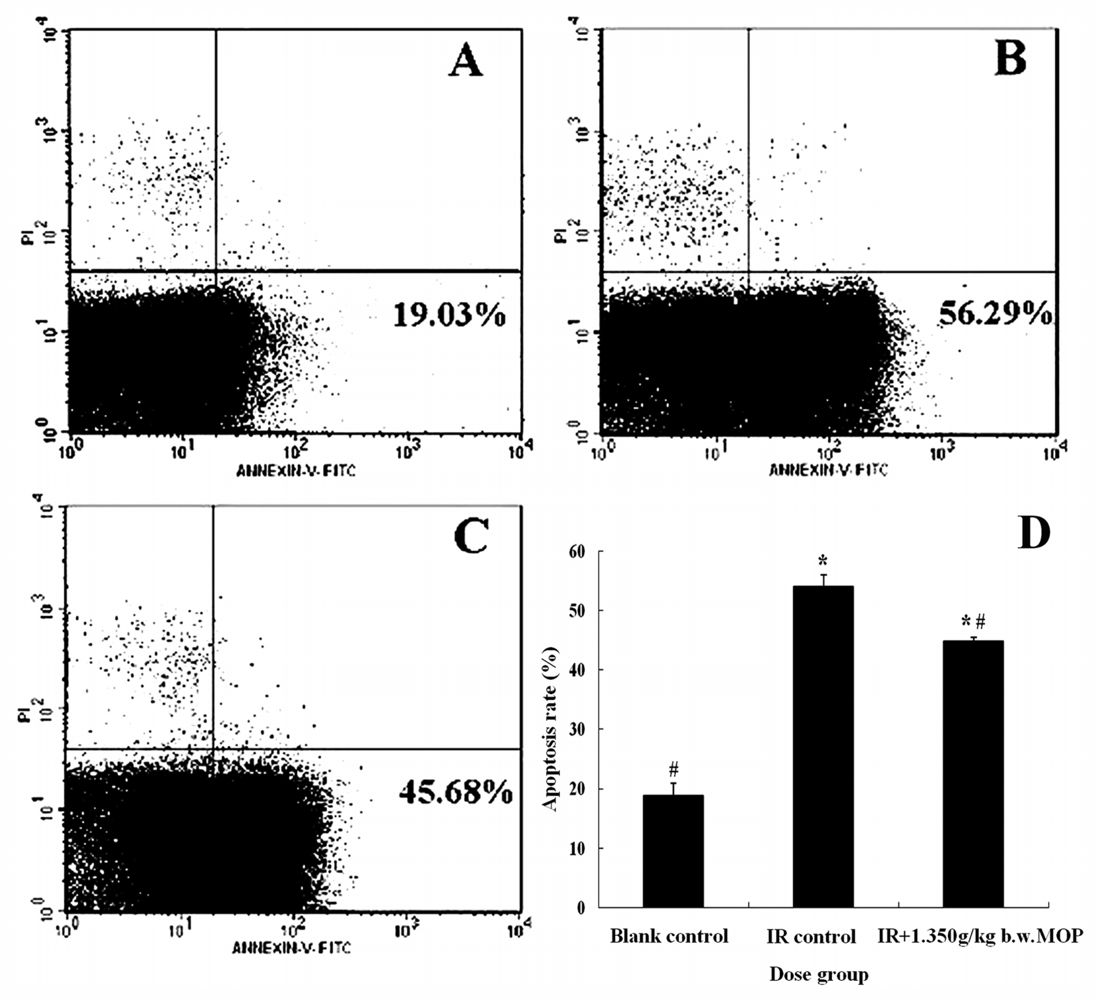
2.4. Effect of MOP on the Apoptosis Rate of Splenocytes after WBI
3. Experimental Section
3.1. Treatment of Mice with MOP
3.2. Irradiation of Animals with 60Co γ-Rays
3.3. Number of White Blood Cells
3.4. Measurement of Antioxidative Systems
3.5. Measurement of Oxidative Products
3.6. Apoptosis Rate Measurement
3.7. Statistical Analysis
4. Conclusions
Acknowledgements
- Samples Availability: Available from the authors.
References
- Korhonen, H.; Pihlanto, A. Food-derived bioactive peptides—Opportunities for designing future foods. Curr. Pharm. Des 2003, 9, 1297–1308. [Google Scholar]
- Pihlanto, A.; Korhonen, H. Bioactive peptides and proteins. Adv. Food Nutr. Res 2003, 47, 175–276. [Google Scholar]
- Rutherfurd-Markwick, K.J.; Moughan, P.J. Bioactive peptides derived from food. J. AOAC Int 2005, 88, 955–966. [Google Scholar]
- Meisel, H. Food-derived bioactive proteins and peptides as potential components of nutraceuticals. Curr. Pharm. Des 2007, 13, 873–874. [Google Scholar]
- Jung, W.K.; Mendis, E.; Je, J.Y.; Park, P.J.; Son, B.W.; Kim, H.C.; Choib, Y.K.; Kim, S.K. Angiotensin I-converting enzyme inhibitory peptide from yellowfin sole (Limanda aspera) frame protein and its antihypertensive effect in spontaneously hypertensive rats. Food Chem 2006, 94, 26–32. [Google Scholar]
- Kim, S.K.; Kim, Y.T.; Byun, H.G.; Nam, K.S.; Joo, D.S.; Shahidi, F. Angiotensin I converting enzyme inhibitory peptides purified from bovin skin gelatin hydrolysate. J. Agric. Food Chem 2001, 49, 2992–2997. [Google Scholar]
- Lordan, S.; Ross, R.P.; Stanton, C. Marine bioactives as functional food ingredients: Potential to reduce the incidence of chronic diseases. Mar. Drugs 2011, 9, 1056–1100. [Google Scholar]
- Liaset, B.; Julshamn, K.; Espe, M. Chemical composition and theoretical nutritional evaluation of the produced fractions from enzymic hydrolysis of salmon frames with Protamex(TM). Process Biochem 2003, 38, 1747–1159. [Google Scholar]
- Zhang, Z.; Zhao, M.; Wang, J.; Ding, Y.; Dai, X.; Li, Y. Oral administration of skin gelatin isolated from chum salmon (Oncorhynchus keta) enhances wound healing in diabetic rats. Mar. Drugs 2011, 9, 696–711. [Google Scholar]
- Gildberg, A.; Arnesen, J.A.; Carlehog, M. Utilisation of cod backbone by biochemical fractionation. Process Biochem 2002, 38, 475–480. [Google Scholar]
- Klompong, V.; Benjakul, S.; Kantachote, D.; Shahidi, F. Antioxidative activity and functional properties of protein hydrolysate of yellow stripe trevally (Selaroides leptolepis) as influenced by the degree of hydrolysis and enzyme type. Food Chem 2007, 102, 1317–1327. [Google Scholar]
- Jung, W.K.; Mendis, E.; Je, J.Y.; Park, P.J.; Son, B.W.; Kim, H.C.; Choi, Y.K.; Kim, S.K. Angiotensin I-converting enzyme inhibitory peptide from yellowfin sole (Limanda aspera) frame protein and its antihypertensive effect in spontaneously hypertensive rats. Food Chem 2006, 94, 26–32. [Google Scholar]
- Yang, R.; Pei, X.; Wang, J.; Zhang, Z.; Zhao, H.; Li, Q.; Zhao, M.; Li, Y. Protective effect of a marine oligopeptide preparation from Chum Salmon (Oncorhynchus keta) on radiation-induced immune suppression in mice. J. Sci. Food Agric 2010, 90, 2241–2248. [Google Scholar]
- Liaset, B.; Espe, M. Nutritional composition of soluble and insoluble fractions obtained by enzymatic hydrolysis of fish-raw materials. Process Biochem 2008, 43, 42–48. [Google Scholar]
- Li, X.; Zhou, A. Preparation of polysaccharides from Acanthopanax senticosus and its inhibition against irradiation-induced injury of rat. Carbohydr. Polym 2007, 67, 219–226. [Google Scholar]
- Lee, T.K.; Allison, R.R.; O’Brien, K.F.; Khazanie, P.G.; Johnke, R.M.; Brown, R.; Bloch, R.M.; Tate, M.L.; Dobbs, L.J.; Kragel, P.J. Ginseng reduces the micronuclei yield in lymphocytes after irradiation. Mutat. Res 2004, 557, 75–84. [Google Scholar]
- Sunila, E.S.; Kuttan, G. Protective effect of Piper longum fruit ethanolic extract on radiation induced damages in mice: A preliminary study. Fitoterapia 2005, 76, 649–655. [Google Scholar]
- Badr El-Din, N.K. Protective role of sanumgerman against γ-irradiation-induced oxidative stress in Ehrlich carcinoma-bearing mice. Nutr. Res 2004, 24, 271–291. [Google Scholar]
- Prabhakar, K.R.; Veerapur, V.P.; Bansal, P.; Parihar, V.K.; Reddy Kandadi, M.; Bhagath Kumar, P.; Priyadarsini, K.I.; Unnikrishnan, M.K. Antioxidant and radioprotective effect of the active fraction of Pilea microphylla (L.) ethanolic extract. Chem. Biol. Interact 2007, 165, 22–32. [Google Scholar]
- Halliwell, B.; Whiteman, M. Measuring reactive species and oxidative damage in vivo and in cellculture: How should you do it and what do the results mean? Br. J. Pharmacol 2004, 142, 231–255. [Google Scholar]
- Jacobson, M.D. Reactive oxygen species and programmed cell death. Trends Biochem. Sci 1996, 21, 83–86. [Google Scholar]
- Helzlsouer, K.J.; Huang, H.Y.; Alberg, A.J. Association between alpha-tocopherol, gamma-tocopherol, selenium, and subsequent prostate cancer. J. Natl. Cancer Inst 2000, 92, 2018–2023. [Google Scholar]
- Levi, F.; Pasche, C.; Lucchini, F.; Vecchia, C.L. Dietary intake of selected micronutrients and breast-cancer risk. Int. J. Cancer Inst 2001, 91, 260–263. [Google Scholar]
- Uchimura, E.; Watanabe, N.; Niwa, O.; Muto, M.; Kobayashi, Y. Transient infiltration of nutrophils into the thymus in association with apoptosis induced by whole-body X-irradiation. J. Leukoc. Biol 2000, 67, 780–784. [Google Scholar]
- Li, C.; Zhou, Z.; Zhu, D.; Sun, Y.; Dai, J.; Wang, S. Protective effect of paeoniflorin on irradiation-induced cell damage involved in modulation of reactive oxygen species and the mitogen-activated protein kinases. Int. J. Biochem. Cell Biol 2007, 39, 426–438. [Google Scholar]
- Yang, R.; Zhang, Z.; Pei, X.; Han, X.; Wang, J.; Wang, L.; Long, Z.; Shen, X.; Li, Y. Immunomodulatory effects of marine oligopeptide preparation from Chum Salmon (Oncorhynchus keta) in mice. Food Chem 2009, 113, 464–470. [Google Scholar]
- NIH Guide for the Care and Use of Laboratory Animals; NIH Publication No. 85-23; National Research Council: Washington, DC, USA, 1985.
- Fridovich, I. Superoxide radical: An endogenous toxicant. Annu. Rev. Pharmacol. Toxicol 1983, 23, 239–257. [Google Scholar]
- Schepetkin, I.A.; Faulkner, C.L.; Nelson-Overton, L.K.; Wiley, J.A.; Quinn, M.T. Macrophage immunomodulatory activity of polysaccharides isolated from Juniperus scopolorum. Int. Immunopharmacol 2005, 5, 1783–1799. [Google Scholar]
- Mihara, M.; Uchiyama, M. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal. Biochem 1978, 86, 271–278. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino acid | No. residues/100 residues |
|---|---|
| Arginine | 7.12 |
| Histidine | 3.39 |
| Isoleucine | 3.76 |
| Leucine | 7.71 |
| Lysine | 9.18 |
| Methionine | 3.52 |
| Phenylalanine | 4.46 |
| Threonine | 4.67 |
| Tryptophan | 0.19 |
| Valine | 5.17 |
| Indispensable AA (IAA) | 49.16 |
| Alanine | 5.70 |
| Aspartic acid | 10.45 |
| Cystine | 1.05 |
| Glutamic acid | 15.72 |
| Glycine | 6.56 |
| Proline | 3.61 |
| Serine | 4.10 |
| Tyrosine | 3.65 |
| Dispensable AA (DAA) | 50.84 |
© 2011 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yang, R.; Wang, J.; Liu, Z.; Pei, X.; Han, X.; Li, Y. Antioxidant Effect of a Marine Oligopeptide Preparation from Chum Salmon (Oncorhynchus keta) by Enzymatic Hydrolysis in Radiation Injured Mice. Mar. Drugs 2011, 9, 2304-2315. https://doi.org/10.3390/md9112304
Yang R, Wang J, Liu Z, Pei X, Han X, Li Y. Antioxidant Effect of a Marine Oligopeptide Preparation from Chum Salmon (Oncorhynchus keta) by Enzymatic Hydrolysis in Radiation Injured Mice. Marine Drugs. 2011; 9(11):2304-2315. https://doi.org/10.3390/md9112304
Chicago/Turabian StyleYang, Ruiyue, Junbo Wang, Zhigang Liu, Xinrong Pei, Xiaolong Han, and Yong Li. 2011. "Antioxidant Effect of a Marine Oligopeptide Preparation from Chum Salmon (Oncorhynchus keta) by Enzymatic Hydrolysis in Radiation Injured Mice" Marine Drugs 9, no. 11: 2304-2315. https://doi.org/10.3390/md9112304
APA StyleYang, R., Wang, J., Liu, Z., Pei, X., Han, X., & Li, Y. (2011). Antioxidant Effect of a Marine Oligopeptide Preparation from Chum Salmon (Oncorhynchus keta) by Enzymatic Hydrolysis in Radiation Injured Mice. Marine Drugs, 9(11), 2304-2315. https://doi.org/10.3390/md9112304