Differential Effect of Smoking on Gene Expression in Head and Neck Cancer Patients


 ,
, 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Cancer Genome Atlas Gene Expression Data for Head and Neck Squamous Cell Carcinomas
2.2. Molecular Classification for Gene Expression
2.3. Survival Analysis
3. Results
3.1. Differential Gene Expression in Tumor Tissues Versus Normal Tissues for Head and Neck Squamous Cell Carcinomas
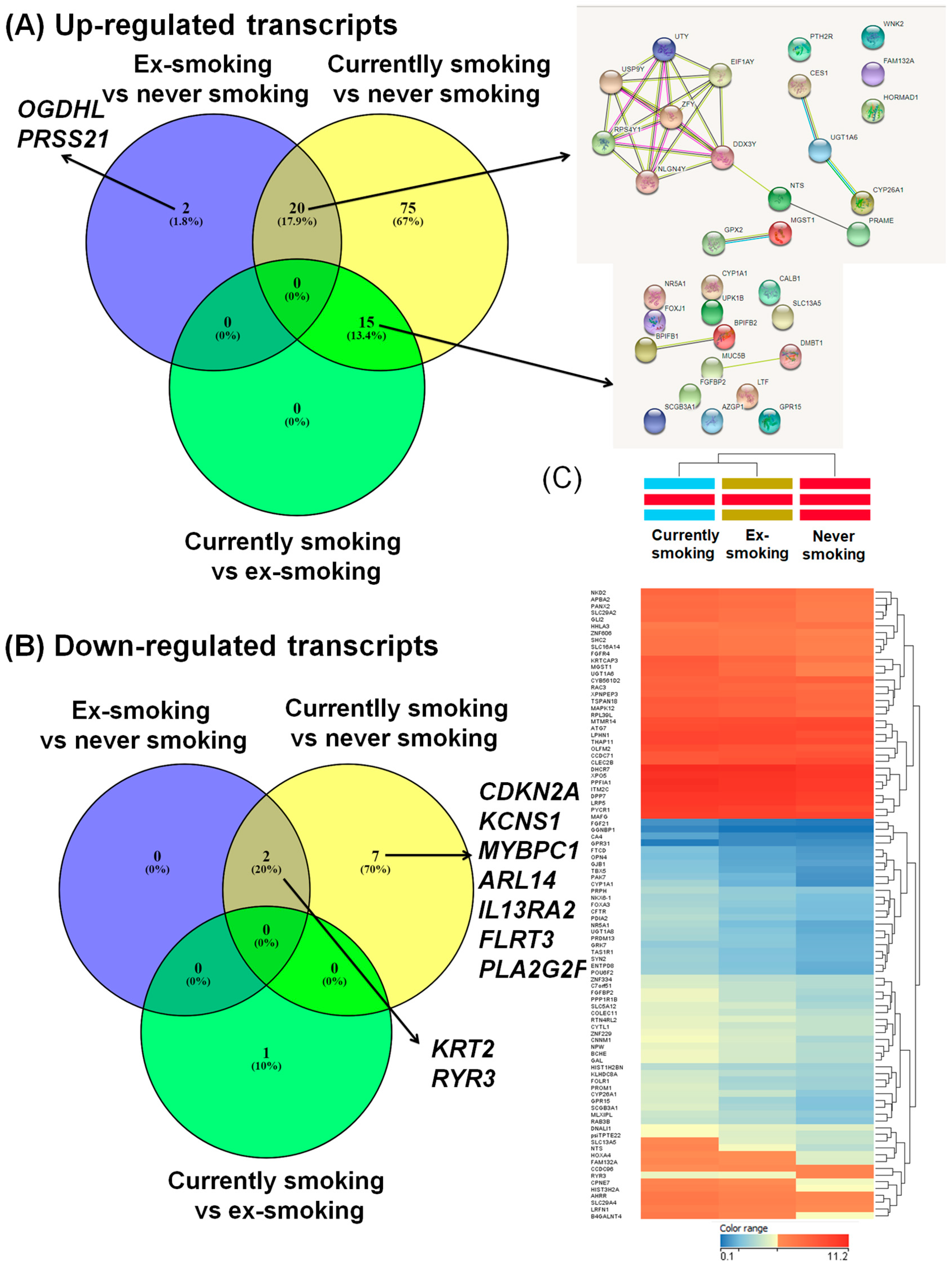
3.2. Differential Gene Expression Levels in Smokers Compared with Non-Smokers or Ex-Smokers in Head and Neck Squamous Cell Carcinomas
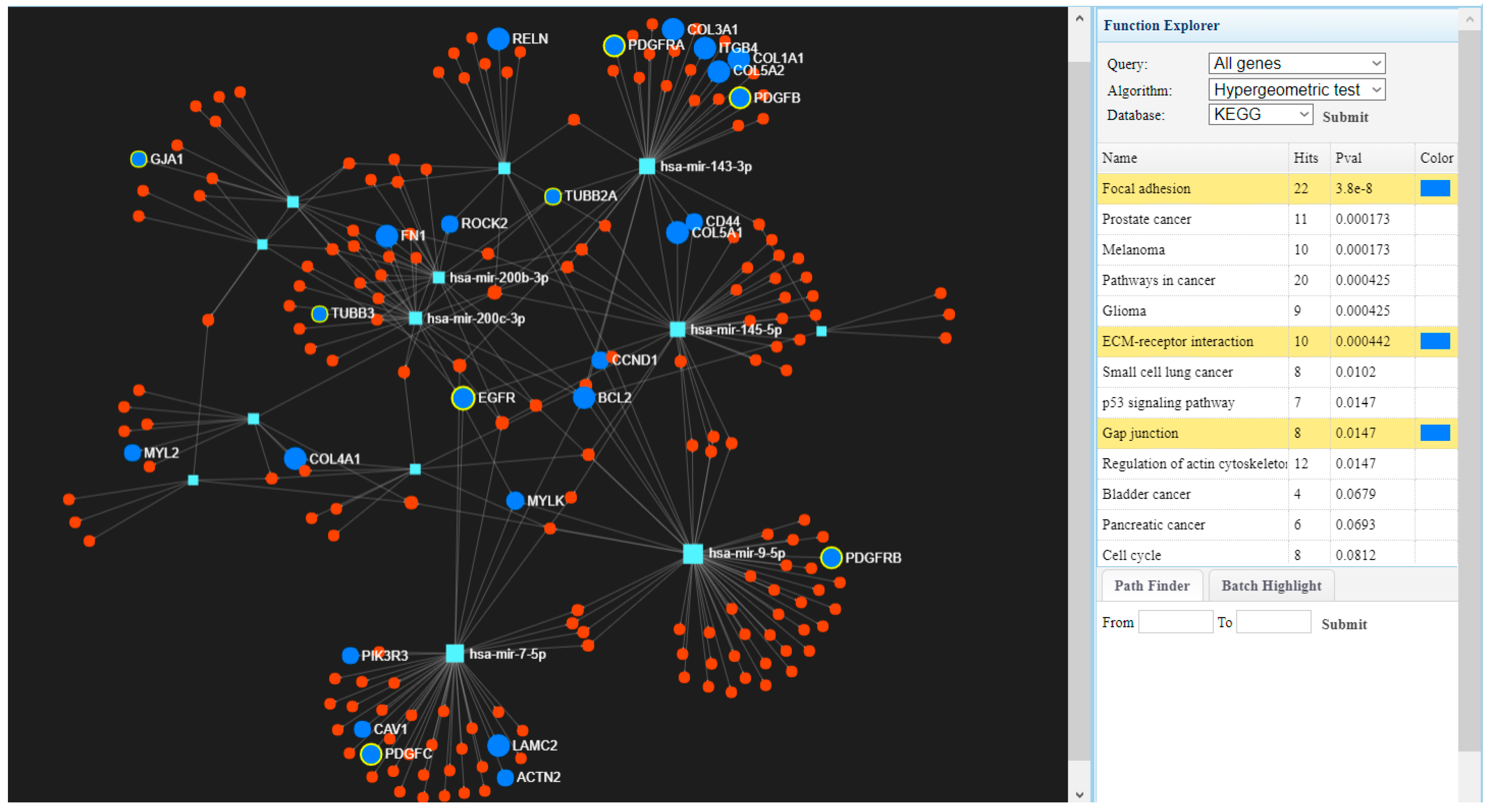
3.3. Molecular Classification for Altered Gene Expression Signature in Smoking versus Never Smoking Head and Neck Squamous Cell Carcinomas Patients
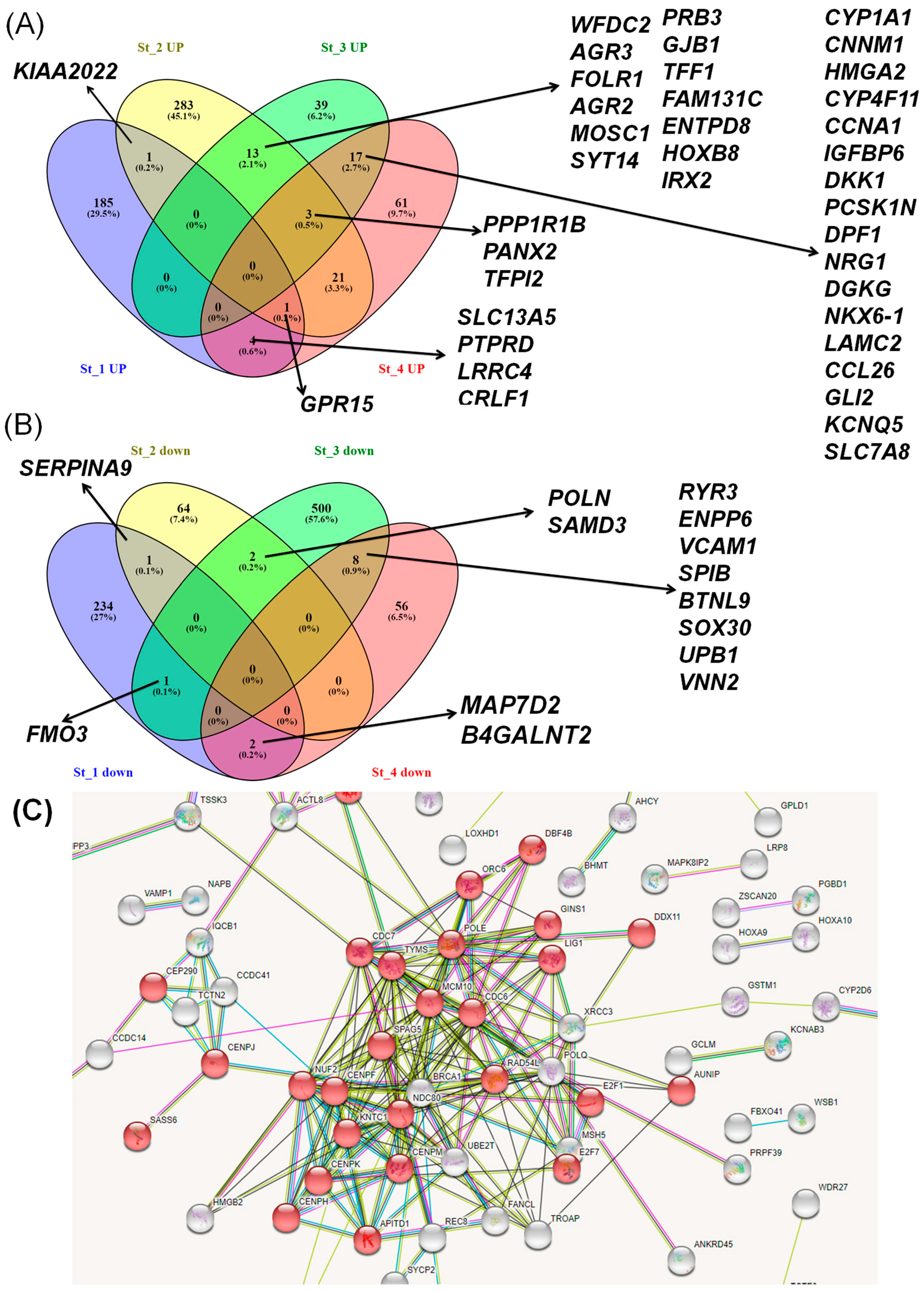
3.4. Effect of Smoking on Head and Neck Squamous Cell Carcinomas Stages
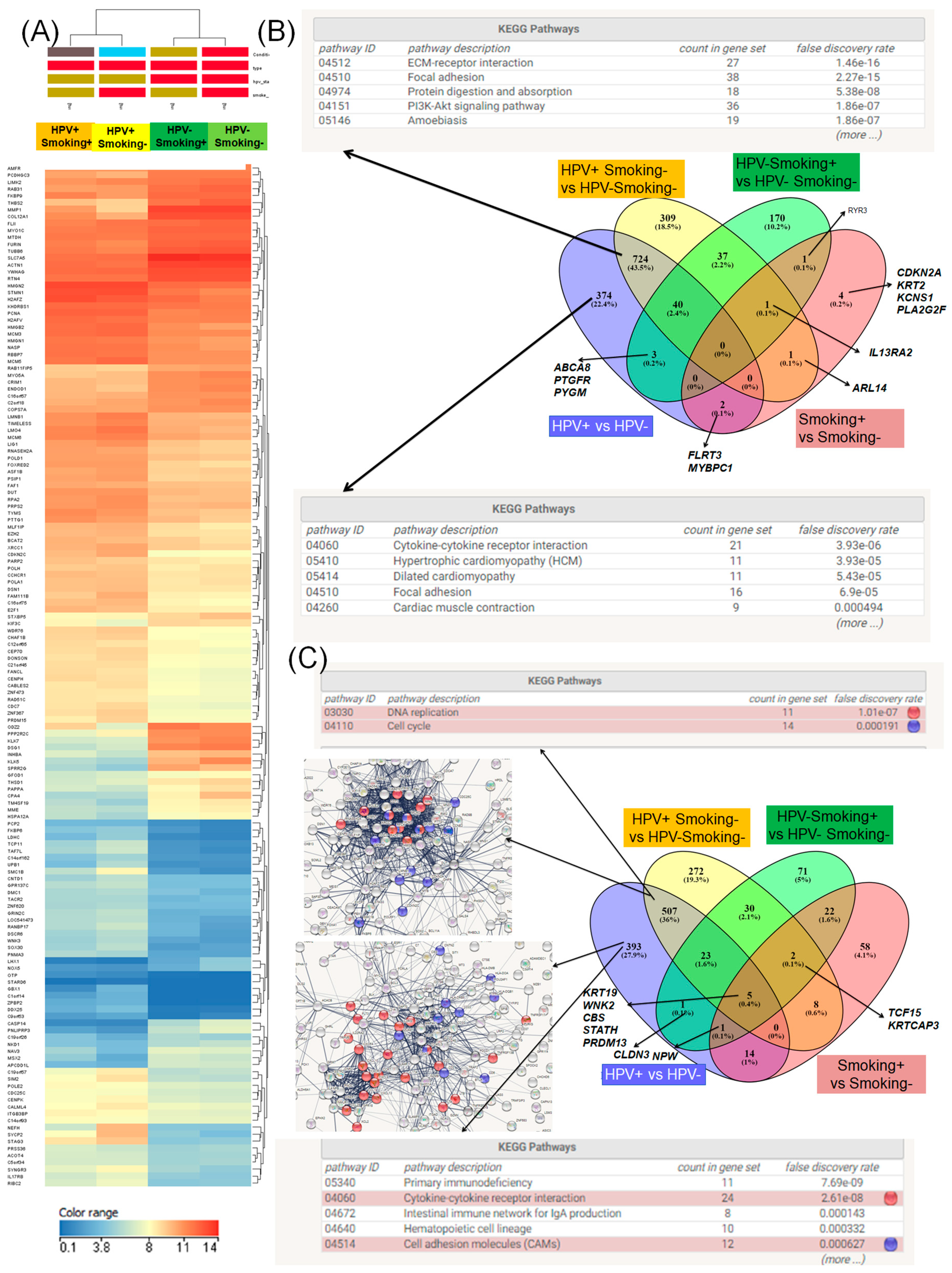
3.5. Evaluation of Gene Expression Signature Based on Human Papilloma Virus 16 Status with/without Correlation with Smoking in Head and Neck Squamous Cell Carcinomas Patients
3.6. Survival Prognosis Analysis Related to Smoking Status in Head and Neck Squamous Cell Carcinomas Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Abrahao, R.; Anantharaman, D.; Gaborieau, V.; Abedi-Ardekani, B.; Lagiou, P.; Lagiou, A.; Ahrens, W.; Holcatova, I.; Betka, J.; Merletti, F.; et al. The influence of smoking, age and stage at diagnosis on the survival after larynx, hypopharynx and oral cavity cancers in Europe: The ARCAGE study. Int. J. Cancer 2018, 143, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Irimie, A.I.; Braicu, C.; Cojocneanu-Petric, R.; Berindan-Neagoe, I.; Campian, R.S. Novel technologies for oral squamous carcinoma biomarkers in diagnostics and prognostics. Acta Odontol. Scand. 2015, 73, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Giraldi, L.; Leoncini, E.; Pastorino, R.; Wunsch-Filho, V.; de Carvalho, M.; Lopez, R.; Cadoni, G.; Arzani, D.; Petrelli, L.; Matsuo, K.; et al. Alcohol and cigarette consumption predict mortality in patients with head and neck cancer: A pooled analysis within the International Head and Neck Cancer Epidemiology (INHANCE) Consortium. Ann. Oncol. 2017, 28, 2843–2851. [Google Scholar] [CrossRef] [PubMed]
- Irimie, A.I.; Zimta, A.A.; Ciocan, C.; Mehterov, N.; Dudea, D.; Braicu, C.; Berindan-Neagoe, I. The Unforeseen Non-Coding RNAs in Head and Neck Cancer. Genes 2018, 9, 134. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Shin, H.R.; Bray, F.; Forman, D.; Mathers, C.; Parkin, D.M. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int. J. Cancer 2010, 127, 2893–2917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salyakina, D.; Tsinoremas, N.F. Non-coding RNAs profiling in head and neck cancers. NPJ Genom. Med. 2016, 1, 15004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irimie, A.I.; Braicu, C.; Sonea, L.; Zimta, A.A.; Cojocneanu-Petric, R.; Tonchev, K.; Mehterov, N.; Diudea, D.; Buduru, S.; Berindan-Neagoe, I. A looking-glass of non-coding RNAs in oral cancer. Int. J. Mol. Sci. 2017, 18, 2620. [Google Scholar] [CrossRef] [PubMed]
- Irimie, A.I.; Sonea, L.; Jurj, A.; Mehterov, N.; Zimta, A.A.; Budisan, L.; Braicu, C.; Berindan-Neagoe, I. Future trends and emerging issues for nanodelivery systems in oral and oropharyngeal cancer. Int. J. Nanomed. 2017, 12, 4593–4606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, H.-M.; Zhao, Y.; Zhang, J.-P.; Zhang, J.-H.; Jiang, P.; Li, B.; Wang, C. Expression and potential mechanism of metabolism-related genes and CRLS1 in non-small cell lung cancer. Oncol. Lett. 2018, 15, 2661–2668. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.H.; Hsia, S.M.; Shih, Y.H.; Shieh, T.M. Association of smoking, alcohol use, and betel quid chewing with epigenetic aberrations in cancers. Int. J. Mol. Sci. 2017, 18, 1210. [Google Scholar] [CrossRef] [PubMed]
- Leemans, C.R.; Snijders, P.J.F.; Brakenhoff, R.H. The molecular landscape of head and neck cancer. Nat. Rev. Cancer 2018, 18, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Vineis, P.; Chadeau-Hyam, M.; Gmuender, H.; Gulliver, J.; Herceg, Z.; Kleinjans, J.; Kogevinas, M.; Kyrtopoulos, S.; Nieuwenhuijsen, M.; Phillips, D.H.; et al. The exposome in practice: Design of the EXPOsOMICS project. Int. J. Hyg. Environ. Health 2017, 220, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Van Breda, S.G.J.; Wilms, L.C.; Gaj, S.; Jennen, D.G.J.; Briedé, J.J.; Kleinjans, J.C.S.; de Kok, T.M.C.M. The exposome concept in a human nutrigenomics study: Evaluating the impact of exposure to a complex mixture of phytochemicals using transcriptomics signatures. Mutagenesis 2015, 30, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Sivabalakrishnan, M. Feature selection of gene expression data for cancer classification: A review. Procedia Comput. Sci. 2015, 50, 52–57. [Google Scholar] [CrossRef]
- Zhang, X.; Cha, I.-H.; Kim, K.-Y. Highly preserved consensus gene modules in human papilloma virus 16 positive cervical cancer and head and neck cancers. Oncotarget 2017, 8, 114031–114040. [Google Scholar] [CrossRef] [PubMed]
- Braicu, C.; Catana, C.; Calin, G.A.; Berindan-Neagoe, I. NCRNA combined therapy as future treatment option for cancer. Curr. Pharm. Des. 2014, 20, 6565–6574. [Google Scholar] [CrossRef] [PubMed]
- UCSC Genome Browser. Available online: https://genome.ucsc.edu (accessed on 12 March 2018).
- STRING: Functional Protein Association Network, Version 10.5. Available online: https://string-db.org (accessed on 16 May 2018).
- KEGG Pathway Maps. Available online: http://www.genome.jp/kegg/pathway.html (accessed on 10 June 2018).
- Panther Clasiffication System. Available online: http://www.pantherdb.org (accessed on 14 June 2018).
- miRNet-Network-Based Visual Analysis of miRNAs, Targets and Functions. Available online: http://www.mirnet.ca/faces/home.xhtml (accessed on 24 June 2018).
- Campbell, J.D.; Yau, C.; Bowlby, R.; Liu, Y.; Brennan, K.; Fan, H.; Taylor, A.M.; Wang, C.; Walter, V.; Akbani, R.; et al. Genomic, pathway network, and immunologic features distinguishing squamous carcinomas. Cell Rep. 2018, 23, 194–212.e6. [Google Scholar] [CrossRef] [PubMed]
- Ferketich, A.K.; Niland, J.C.; Mamet, R.; Zornosa, C.; D’Amico, T.A.; Ettinger, D.S.; Kalemkerian, G.P.; Pisters, K.M.; Reid, M.E.; Otterson, G.A. Smoking status and survival in the national comprehensive cancer network non-small cell lung cancer cohort. Cancer 2013, 119, 847–853. [Google Scholar] [CrossRef] [PubMed]
- Bryant, A.; Cerfolio, R.J. Differences in epidemiology, histology, and survival between cigarette smokers and never-smokers who develop non-small cell lung cancer. Chest 2007, 132, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Koshiaris, C.; Aveyard, P.; Oke, J.; Ryan, R.; Szatkowski, L.; Stevens, R.; Farley, A. Smoking cessation and survival in lung, upper aero-digestive tract and bladder cancer: Cohort study. Br. J. Cancer 2017, 117, 1224–1232. [Google Scholar] [CrossRef] [PubMed]
- Pierce, J.P.; Patterson, R.E.; Senger, C.M.; Flatt, S.W.; Caan, B.J.; Natarajan, L.; Nechuta, S.J.; Poole, E.M.; Shu, X.-O.; Chen, W.Y. Lifetime cigarette smoking and breast cancer prognosis in the after breast cancer pooling project. JNCI J. Nat. Cancer Inst. 2014, 106, djt359. [Google Scholar] [CrossRef] [PubMed]
- Champion, M.; Brennan, K.; Croonenborghs, T.; Gentles, A.J.; Pochet, N.; Gevaert, O. Module analysis captures pancancer genetically and epigenetically deregulated cancer driver genes for smoking and antiviral response. EBioMedicine 2018, 27, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Yavorski, J.M.; Blanck, G. Smoking correlates with increased cytoskeletal protein-related coding region mutations in the lung and head and neck datasets of the cancer genome atlas. Physiol. Rep. 2016, 4, e13045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, M.; Quabius, E.S.; Tribius, S.; Hebebrand, L.; Gorogh, T.; Halec, G.; Kahn, T.; Hedderich, J.; Rocken, C.; Haag, J.; et al. Human papillomavirus infection in head and neck cancer: The role of the secretory leukocyte protease inhibitor. Oncol. Rep. 2013, 29, 1962–1968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quabius, E.S.; Moller, P.; Haag, J.; Pfannenschmidt, S.; Hedderich, J.; Gorogh, T.; Rocken, C.; Hoffmann, M. The role of the antileukoprotease SLPI in smoking-induced human papillomavirus-independent head and neck squamous cell carcinomas. Int. J. Cancer 2014, 134, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Osazuwa-Peters, N.; Adjei Boakye, E.; Chen, B.Y.; Tobo, B.B.; Varvares, M.A. Association Between Head and Neck Squamous Cell Carcinoma Survival, Smoking at Diagnosis, and Marital Status. JAMA Otolaryngol. 2018, 144, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Rudin, C.M.; Avila-Tang, E.; Harris, C.C.; Herman, J.G.; Hirsch, F.R.; Pao, W.; Schwartz, A.G.; Vahakangas, K.H.; Samet, J.M. Lung cancer in never smokers: Molecular profiles and therapeutic implications. Clin. Cancer Res. 2009, 15, 5646–5661. [Google Scholar] [CrossRef] [PubMed]
- Delagranda, A.; Leterme, G.; Chirpaz, E.; Ferdynus, C.; Fernandez, C.; Rubin, F. Epidemiological features of cancers of the oral cavity, oropharynx, hypopharynx and larynx cancer in Reunion Island. Eur. Ann. Otorhinolaryngol. Head Neck Dis. 2018, 135, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Cufari, M.E.; Proli, C.; De Sousa, P.; Raubenheimer, H.; Al Sahaf, M.; Chavan, H.; Shedden, L.; Niwaz, Z.; Leung, M.; Nicholson, A.G.; et al. Increasing frequency of non-smoking lung cancer: Presentation of patients with early disease to a tertiary institution in the UK. Eur. J. Cancer 2017, 84, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Farshadpour, F.; Roepman, P.; Hordijk, G.J.; Koole, R.; Slootweg, P.J. A gene expression profile for non-smoking and non-drinking patients with head and neck cancer. Oral Dis. 2012, 18, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Network. Comprehensive genomic characterization of head and neck squamous cell carcinomas. Nature 2015, 517, 576–582. [Google Scholar] [Green Version]
- Rouissi, K.; Ouerhani, S.; Hamrita, B.; Bougatef, K.; Marrakchi, R.; Cherif, M.; Ben Slama, M.R.; Bouzouita, M.; Chebil, M.; Ben Ammar Elgaaied, A. Smoking and polymorphisms in xenobiotic metabolism and DNA repair genes are additive risk factors affecting bladder cancer in Northern Tunisia. Pathol. Oncol. Res. 2011, 17, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Ihsan, R.; Chauhan, P.S.; Mishra, A.K.; Yadav, D.S.; Kaushal, M.; Sharma, J.D.; Zomawia, E.; Verma, Y.; Kapur, S.; Saxena, S. Multiple analytical approaches reveal distinct gene-environment interactions in smokers and non smokers in lung cancer. PLoS ONE 2011, 6, e29431. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, P.S.; Ihsan, R.; Mishra, A.K.; Yadav, D.S.; Saluja, S.; Mittal, V.; Saxena, S.; Kapur, S. High order interactions of xenobiotic metabolizing genes and P53 codon 72 polymorphisms in acute leukemia. Environ. Mol. Mutagen. 2012, 53, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Petros, W.P.; Younis, I.R.; Ford, J.N.; Weed, S.A. Effects of tobacco smoking & nicotine on cancer treatment. Pharmacotherapy 2012, 32, 920–931. [Google Scholar] [PubMed]
- Gümüş, Z.H.; Du, B.; Kacker, A.; Boyle, J.O.; Bocker, J.M.; Mukherjee, P.; Subbaramaiah, K.; Dannenberg, A.J.; Weinstein, H. Effects of tobacco smoke on gene expression and cellular pathways in a cellular model of oral leukoplakia. Cancer Prev. Res. 2008, 1, 100–111. [Google Scholar] [CrossRef] [PubMed]
- Irimie, A.I.; Braicu, C.; Zanoaga, O.; Pileczki, V.; Gherman, C.; Berindan-Neagoe, I.; Campian, R.S. Epigallocatechin-3-gallate suppresses cell proliferation and promotes apoptosis and autophagy in oral cancer SSC-4 cells. Onco Targets Ther. 2015, 8, 461–470. [Google Scholar] [PubMed]
- Braicu, C.; Mehterov, N.; Vladimirov, B.; Sarafian, V.; Nabavi, S.M.; Atanasov, A.G.; Berindan-Neagoe, I. Nutrigenomics in cancer: Revisiting the effects of natural compounds. Semin. Cancer Biol. 2017, 46, 84–106. [Google Scholar] [CrossRef] [PubMed]
- Budisan, L.; Gulei, D.; Zanoaga, O.M.; Irimie, A.I.; Sergiu, C.; Braicu, C.; Gherman, C.D.; Berindan-Neagoe, I. Dietary intervention by phytochemicals and their role in modulating coding and non-coding genes in cancer. Int. J. Mol. Sci. 2017, 18, 1178. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.W.; Lee, H.M.; Lee, K.; Kim, B.; Lee, M.Y.; Choi, K.C. Effects of cigarette smoke extracts on cell cycle, cell migration and endocrine activity in human placental cells. Reprod. Toxicol. 2017, 73, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Mirghani, H.; Ugolin, N.; Ory, C.; Lefevre, M.; Baulande, S.; Hofman, P.; St Guily, J.L.; Chevillard, S.; Lacave, R. A predictive transcriptomic signature of oropharyngeal cancer according to HPV16 status exclusively. Oral Oncol. 2014, 50, 1025–1034. [Google Scholar] [CrossRef] [PubMed]
- Scott, D.A.; Palmer, R.M. The influence of tobacco smoking on adhesion molecule profiles. Tob. Induc. Dis. 2003, 1, 7–25. [Google Scholar] [CrossRef] [PubMed]
- Zahra, A.; Rubab, I.; Malik, S.; Khan, A.; Khan, M.J.; Fatmi, M.Q. Meta-Analysis of miRNAs and their involvement as biomarkers in oral cancers. BioMed Res. Int. 2018, 2018, 8439820. [Google Scholar] [CrossRef] [PubMed]
- Palmer, R.M.; Stapleton, J.A.; Sutherland, G.; Coward, P.Y.; Wilson, R.F.; Scott, D.A. Effect of nicotine replacement and quitting smoking on circulating adhesion molecule profiles (sICAM-1, sCD44v5, sCD44v6). Eur. J. Clin. Investig. 2002, 32, 852–857. [Google Scholar] [CrossRef]
- Scott, D.A.; Stapleton, J.A.; Palmer, R.M.; Wilson, R.F.; Sutherland, G.; Coward, P.Y.; Gustavsson, G.; Odell, E.W.; Poston, R.N. Plasma concentrations of reputed tumor-associated soluble CD44 isoforms (v5 and v6) in smokers are dose related and decline on smoking cessation. Cancer Epidemiol. Biomark. Prev. 2000, 9, 1211–1214. [Google Scholar]
- Ionescu, C.; Braicu, C.; Chiorean, R.; Cojocneanu Petric, R.; Neagoe, E.; Pop, L.; Chira, S.; Berindan-Neagoe, I. TIMP-1 expression in human colorectal cancer is associated with SMAD3 gene expression levels: A pilot study. JGLD 2014, 23, 413–418. [Google Scholar] [PubMed]
- Braicu, C.; Tudoran, O.; Balacescu, L.; Catana, C.; Neagoe, E.; Berindan-Neagoe, I.; Ionescu, C. The significance of PDGF expression in serum of colorectal carcinoma patients—Correlation with Duke’s classification. Can PDGF become a potential biomarker? Chirurgia 2013, 108, 849–854. [Google Scholar] [PubMed]
- Ionescu, S.; Barbu, E.; Ionescu, C.; Costache, A.; Balasoiu, M. Giant gastrointestinal stromal tumor of the stomach. Rom. J. Morphol. Embryol. 2015, 56, 239–245. [Google Scholar] [PubMed]
- Scott, D.A.; Todd, D.H.; Coward, P.Y.; Wilson, R.F.; Odell, E.W.; Poston, R.N.; Matthews, J.P.; Palmer, R.M. The acute influence of tobacco smoking on adhesion molecule expression on monocytes and neutrophils and on circulating adhesion molecule levels in vivo. Addict. Biol. 2000, 5, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Shiels, M.S.; Katki, H.A.; Freedman, N.D.; Purdue, M.P.; Wentzensen, N.; Trabert, B.; Kitahara, C.M.; Furr, M.; Li, Y.; Kemp, T.J.; et al. Cigarette smoking and variations in systemic immune and inflammation markers. J. Natl. Cancer Inst. 2014, 106, dju294. [Google Scholar] [CrossRef] [PubMed]
- Mandal, R.; Şenbabaoğlu, Y.; Desrichard, A.; Havel, J.J.; Dalin, M.G.; Riaz, N.; Lee, K.-W.; Ganly, I.; Hakimi, A.A.; Chan, T.A.; et al. The head and neck cancer immune landscape and its immunotherapeutic implications. JCI Insight 2016, 1, e89829. [Google Scholar] [CrossRef] [PubMed]
- Desrichard, A.; Kuo, F.; Chowell, D.; Lee, K.W.; Riaz, N.; Wong, R.J.; Chan, T.A.; Morris, L.G.T. Tobacco Smoking-Associated Alterations in the Immune Microenvironment of Squamous Cell Carcinomas. J. Natl. Cancer Inst. 2018. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Taneja, V.; Vassallo, R. Cigarette smoking and inflammation: Cellular and molecular mechanisms. J. Dent. Res. 2012, 91, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Solomon, B.; Young, R.J.; Rischin, D. Head and neck squamous cell carcinoma: Genomics and emerging biomarkers for immunomodulatory cancer treatments. Semin. Cancer Biol. 2018, in press. [Google Scholar] [CrossRef] [PubMed]
- Dok, R.; Nuyts, S. HPV positive head and neck cancers: Molecular pathogenesis and evolving treatment strategies. Cancers 2016, 8, 41. [Google Scholar] [CrossRef] [PubMed]
- Thibodeau, B.J.; Geddes, T.J.; Fortier, L.E.; Ahmed, S.; Pruetz, B.L.; Wobb, J.; Chen, P.; Wilson, G.D.; Akervall, J.A. Gene expression Characterization of HPV positive head and neck cancer to predict response to chemoradiation. Head Neck Pathol. 2015, 9, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Bishop, J.A.; Lewis, J.S., Jr.; Rocco, J.W.; Faquin, W.C. HPV-related squamous cell carcinoma of the head and neck: An update on testing in routine pathology practice. Semin. Diagn. Pathol. 2015, 32, 344–351. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Parameters | Patients (n = 519) | Females/Males | |
|---|---|---|---|
| Sex | Females | 134 | |
| Males | 374 | ||
| Undeclared | 11 | ||
| Age | Median, range | 61, 19–90 | |
| Median, range males | 59, 19–88 | ||
| Median, range females | 64.5, 24–90 | ||
| Undeclared | 11 | ||
| Clinical stage | 1 | 20 | 9/11 |
| 2 | 98 | 33/65 | |
| 3 | 101 | 26/75 | |
| 4 | 275 | 64/211 | |
| Unknown | 25 | ||
| Clinical TNM | T1N0M0 | 18 | 7/11 |
| T1N1M0 | 5 | 2/3 | |
| T1N2M0 | 7 | 1/6 | |
| T1N2M1 | 1 | -/1 | |
| T1NxM0 | 2 | 2/- | |
| T1N0M0 | 96 | 33/63 | |
| T1N0Mx | 1 | -/1 | |
| T2N1M0 | 12 | 3/9 | |
| T2N1Mx | 1 | 1/- | |
| T2N2M0 | 33 | 6/27 | |
| T2N2M1 | 1 | -/1 | |
| T2N2Mx | 3 | -/3 | |
| T1N3M0 | 1 | -/1 | |
| T2NxM0 | 1 | -/1 | |
| T3N0M0 | 58 | 15/43 | |
| T3N0Mx | 1 | 1/- | |
| T2N1M0 | 20 | 4/16 | |
| T3N1M1 | 1 | 1/- | |
| T3N2M0 | 46 | 8/38 | |
| T3N2M1 | 1 | 1/- | |
| T3N2Mx | 1 | -/1 | |
| T3N3M0 | 1 | -/1 | |
| T3NxM0 | 2 | -/2 | |
| T4N0M0 | 66 | 17/49 | |
| T4N0Mx | 1 | 1/- | |
| T4N1M0 | 39 | 13/26 | |
| T4N2M0 | 62 | 14/48 | |
| T4N2Mx | 1 | -/1 | |
| T4N3M0 | 7 | 2/5 | |
| T4NxM0 | 3 | -/3 | |
| TxN1M0 | 1 | -/1 | |
| TxN2M0 | 1 | -/1 | |
| TxNxMx | 9 | -/9 | |
| Unknown | 16 | ||
| Smoking history | Smoker | 174 | 35/139 |
| Reformed smoker <15 years | 134 | 26/108 | |
| Reformed smoker >15 years | 72 | 19/53 | |
| Reformed, unknown years | 2 | 0/2 | |
| Lifelong non-smoker | 114 | 49/65 | |
| Unknown | 23 | ||
| Anatomic neoplasm subdivision | Alveolar ridge | 18 | |
| Base of tongue | 27 | ||
| Buccal mucosa | 20 | ||
| Floor of mouth | 60 | ||
| Hard palate | 7 | ||
| Hypopharynx | 9 | ||
| Larynx | 114 | ||
| Lip | 3 | ||
| Oral cavity | 71 | ||
| Oral tongue | 127 | ||
| Oropharynx | 10 | ||
| Tonsil | 42 | ||
| Unknown | 11 | ||
| HPV | Positive | 72 | |
| Negative | 37 | ||
| Unknown | 410 |
| Gene | FC (abs) | p-Value | Regulation | Gene | FC (abs) | p-Value | Regulation |
|---|---|---|---|---|---|---|---|
| CDKN2A | −2.59737 | 0.000603 | Down | EPHA7 | 2.39461 | 0.000111 | Up |
| RYR3 | −2.35924 | 1.45 × 10−6 | Down | psiTPTE22 | 2.394392 | 6.22 × 10−6 | Up |
| KRT2 | −2.31761 | 0.002097 | Down | POU6F2 | 2.393242 | 3.2 × 10−6 | Up |
| KCNS1 | −2.31495 | 0.000321 | Down | SOHLH1 | 2.390684 | 0.000256 | Up |
| MYBPC1 | −2.10091 | 0.046549 | Down | LTF | 2.380863 | 0.012372 | Up |
| ARL14 | −2.08283 | 0.000138 | Down | MLXIPL | 2.373807 | 8 × 10−6 | Up |
| IL13RA2 | −2.05906 | 8.82 × 10−5 | Down | GLI2 | 2.372254 | 6.52 × 10−7 | Up |
| FLRT3 | −2.02315 | 0.002341 | Down | NLGN4Y | 2.358872 | 0.00092 | Up |
| PLA2G2F | −2.01307 | 0.002746 | Down | PAK7 | 2.355639 | 6.22 × 10−6 | Up |
| NTS | 5.224654 | 8 × 10−6 | Up | FIBCD1 | 2.345015 | 0.000181 | Up |
| RPS4Y1 | 4.924049 | 7.27 × 10−5 | Up | GATA4 | 2.343389 | 0.000117 | Up |
| UGT1A6 | 4.505707 | 3.3 × 10−7 | Up | PANX2 | 2.319508 | 1.31 × 10−6 | Up |
| UPK1B | 4.336869 | 4.15 × 10−5 | Up | PCYT1B | 2.314619 | 3.29 × 10−5 | Up |
| MGST1 | 4.296636 | 4.68 × 10−7 | Up | FGF19 | 2.31125 | 3.18 × 10−5 | Up |
| CYP1A1 | 4.269958 | 5.3 × 10−13 | Up | SLC44A4 | 2.29202 | 0.000356 | Up |
| C20orf114 | 3.985658 | 0.001123 | Up | SCN2A | 2.28985 | 0.00011 | Up |
| SCGB3A1 | 3.985451 | 3.93 × 10−5 | Up | PROM1 | 2.2842 | 0.001556 | Up |
| GPR15 | 3.913692 | 2.69 × 10−15 | Up | CYorf15A | 2.257771 | 0.002582 | Up |
| DDX3Y | 3.748438 | 0.000214 | Up | TFPI2 | 2.25193 | 0.000222 | Up |
| MUC5B | 3.573662 | 0.001207 | Up | MSI1 | 2.249305 | 2.06 × 10−5 | Up |
| CNNM1 | 3.508977 | 3.02 × 10−8 | Up | ADD2 | 2.237741 | 6.45 × 10−5 | Up |
| CES1 | 3.41544 | 1.21 × 10−5 | Up | ALDH1A1 | 2.234898 | 0.000377 | Up |
| PRAME | 3.289014 | 0.000277 | Up | ERN2 | 2.210954 | 0.000992 | Up |
| GPX2 | 3.252181 | 3.95 × 10−6 | Up | LGI3 | 2.206128 | 0.000183 | Up |
| CYP26A1 | 3.241253 | 2.07 × 10−5 | Up | PRKY | 2.206114 | 0.002395 | Up |
| NR5A1 | 3.186291 | 1.24 × 10−5 | Up | SALL1 | 2.200458 | 8.32 × 10−6 | Up |
| PPP1R1B | 3.18047 | 1.38 × 10−5 | Up | TBX5 | 2.194891 | 8.95 × 10−6 | Up |
| FGFBP2 | 3.156294 | 2.95 × 10−6 | Up | HOXA4 | 2.181679 | 4.15 × 10−6 | Up |
| SLC13A5 | 3.142512 | 6.04 × 10−6 | Up | TNNI3 | 2.18105 | 4.06 × 10−5 | Up |
| DMBT1 | 3.102893 | 0.000721 | Up | PLUNC | 2.179797 | 0.009085 | Up |
| KRTCAP3 | 3.077278 | 7.92 × 10−12 | Up | NGB | 2.177274 | 0.000323 | Up |
| BPIL1 | 3.011847 | 0.002582 | Up | FOLR1 | 2.175127 | 0.000402 | Up |
| UGT1A8 | 3.00819 | 1.43 × 10−5 | Up | GDA | 2.163271 | 0.00378 | Up |
| PTH2R | 2.849531 | 2.55 × 10−5 | Up | AKR1C3 | 2.150982 | 8.66 × 10−5 | Up |
| KRT19 | 2.812564 | 0.001704 | Up | AZGP1 | 2.142351 | 0.004176 | Up |
| GAL | 2.792954 | 6.04 × 10−6 | Up | CCNA1 | 2.138528 | 0.001642 | Up |
| EIF1AY | 2.761805 | 0.001147 | Up | PCDH19 | 2.131033 | 0.000887 | Up |
| WNK2 | 2.756387 | 3.74 × 10−5 | Up | GJB7 | 2.128482 | 0.000382 | Up |
| B4GALNT4 | 2.731352 | 5.73 × 10−7 | Up | WDR72 | 2.119921 | 0.003053 | Up |
| RAB3B | 2.721629 | 2.09 × 10−7 | Up | CLDN8 | 2.118758 | 0.001164 | Up |
| FAM132A | 2.67193 | 2.95 × 10−6 | Up | CBS | 2.117995 | 2.03 × 10−5 | Up |
| HOXA7 | 2.666479 | 3.29 × 10−5 | Up | MSMB | 2.117211 | 0.003146 | Up |
| PIGR | 2.652387 | 0.004096 | Up | CFTR | 2.112852 | 0.00034 | Up |
| BCHE | 2.639082 | 3.2 × 10−6 | Up | NTRK2 | 2.112379 | 0.001737 | Up |
| UGT8 | 2.635306 | 5.71 × 10−5 | Up | FGF13 | 2.108137 | 2.2 × 10−5 | Up |
| USP9Y | 2.629508 | 0.001642 | Up | RPL39L | 2.085114 | 5.8 × 10−6 | Up |
| PDIA2 | 2.613751 | 2.12 × 10−6 | Up | SLC29A4 | 2.079458 | 4.68 × 10−7 | Up |
| ZFY | 2.558155 | 0.0013 | Up | ADH7 | 2.07414 | 0.012696 | Up |
| CALB1 | 2.545223 | 0.002413 | Up | PIWIL2 | 2.068965 | 0.000399 | Up |
| AKR1C1 | 2.540783 | 7.2 × 10−6 | Up | CYP1B1 | 2.057724 | 8.81 × 10−5 | Up |
| UTY | 2.533913 | 0.00193 | Up | CPNE7 | 2.055357 | 1.57 × 10−6 | Up |
| ATP13A5 | 2.530633 | 0.00011 | Up | BRDT | 2.041222 | 0.001179 | Up |
| SLC5A12 | 2.504015 | 2.95 × 10−6 | Up | CHGA | 2.033392 | 2.12 × 10−5 | Up |
| FOXJ1 | 2.491234 | 0.000516 | Up | ABO | 2.032663 | 0.001283 | Up |
| PRDM13 | 2.479601 | 4.03 × 10−7 | Up | STATH | 2.023424 | 0.020511 | Up |
| HORMAD1 | 2.437306 | 0.001219 | Up | SCN9A | 2.018196 | 0.00048 | Up |
| UCHL1 | 2.434416 | 7.96 × 10−6 | Up | ADAMTS20 | 2.011134 | 0.000245 | Up |
| NPW | 2.41423 | 6.04 × 10−6 | Up | RBM11 | 2.010622 | 4.12 × 10−5 | Up |
| PNCK | 2.396283 | 8.83 × 10−5 | Up | ZNF556 | 2.009657 | 1.24 × 10−5 | Up |
| TCF15 | 2.007499 | 4.31 × 10−5 | Up |
| Ontology Function | Type | No. Molecules | Percent (%) |
|---|---|---|---|
| Molecular function | binding (GO:0005488) | 33 | 36.7% |
| catalytic activity (GO:0003824) | 30 | 33.3% | |
| transporter activity (GO:0005215) | 14 | 15.6% | |
| receptor activity (GO:0004872) | 4 | 4.4% | |
| signal transducer activity (GO:0004871) | 4 | 4.4% | |
| structural molecule activity (GO:0005198) | 3 | 3.3% | |
| translation regulator activity (GO:0045182) | 1 | 1.1% | |
| antioxidant activity (GO:0016209) | 1 | 1.1% | |
| Biological process | cellular process (GO:0009987) | 48 | 28.1% |
| metabolic process (GO:0008152) | 32 | 18.7% | |
| biological regulation (GO:0065007) | 23 | 13.5% | |
| developmental process (GO:0032502) | 16 | 9.4% | |
| response to stimulus (GO:0050896) | 15 | 8.8% | |
| multicellular organismal process (GO:0032501) | 13 | 7.6% | |
| localization (GO:0051179) | 8 | 4.7% | |
| cellular component organization or biogenesis (GO:0071840) | 7 | 4.1% | |
| biological adhesion (GO:0022610) | 4 | 2.3% | |
| locomotion (GO:0040011) | 3 | 1.8% | |
| immune system process (GO:0002376) | 1 | 0.6% | |
| reproduction (GO:0000003) | 1 | 0.6% | |
| Protein class | transporter (PC00227) | 10 | 13.5% |
| hydrolase (PC00121) | 9 | 12.2% | |
| oxidoreductase (PC00176) | 8 | 10.8% | |
| transcription factor (PC00218) | 8 | 10.8% | |
| nucleic acid binding (PC00171) | 7 | 9.5% | |
| signaling molecule (PC00207) | 6 | 8.1% | |
| transferase (PC00220) | 5 | 6.8% | |
| enzyme modulator (PC00095) | 5 | 6.8% | |
| receptor (PC00197) | 3 | 4.1% | |
| extracellular matrix protein (PC00102) | 2 | 2.7% | |
| cytoskeletal protein (PC00085) | 2 | 2.7% | |
| transfer/carrier protein (PC00219) | 2 | 2.7% | |
| cell junction protein (PC00070) | 2 | 2.7% | |
| lyase (PC00144) | 1 | 1.4% | |
| calcium-binding protein (PC00060) | 1 | 1.4% | |
| defense/immunity protein (PC00090) | 1 | 1.4% | |
| membrane traffic protein (PC00150) | 1 | 1.4% | |
| isomerase (PC00135) | 1 | 1.4% |
| Ontology Function | Type | No. Molecules | Percent (%) |
|---|---|---|---|
| Molecular function | binding (GO:0005488) | 1433 | 37.5% |
| catalytic activity (GO:0003824) | 1262 | 33.0% | |
| transporter activity (GO:0005215) | 429 | 11.2% | |
| receptor activity (GO:0004872) | 255 | 6.7% | |
| signal transducer activity (GO:0004871) | 231 | 6.0% | |
| structural molecule activity (GO:0005198) | 157 | 4.1% | |
| antioxidant activity (GO:0016209) | 23 | 0.6% | |
| translation regulator activity (GO:0045182) | 17 | 0.4% | |
| channel regulator activity (GO:0016247) | 13 | 0.3% | |
| Biological process | cellular process (GO:0009987) | 2479 | 29.5% |
| metabolic process (GO:0008152) | 1589 | 18.9% | |
| biological regulation (GO:0065007) | 911 | 10.9% | |
| response to stimulus (GO:0050896) | 696 | 8.3% | |
| localization (GO:0051179) | 583 | 6.9% | |
| cellular component organization or biogenesis (GO:0071840) | 581 | 6.9% | |
| developmental process (GO:0032502) | 534 | 6.4% | |
| multicellular organismal process (GO:0032501) | 523 | 6.2% | |
| immune system process (GO:0002376) | 146 | 1.7% | |
| locomotion (GO:0040011) | 120 | 1.4% | |
| biological adhesion (GO:0022610) | 110 | 1.3% | |
| reproduction (GO:0000003) | 83 | 1.0% | |
| rhythmic process (GO:0048511) | 32 | 0.4% | |
| cell killing (GO:0001906) | 6 | 0.1% | |
| Protein class | nucleic acid binding (PC00171) | 565 | 15.2% |
| transcription factor (PC00218) | 504 | 13.5% | |
| hydrolase (PC00121) | 410 | 11.0% | |
| receptor (PC00197) | 355 | 9.5% | |
| transporter (PC00227) | 269 | 7.2% | |
| signaling molecule (PC00207) | 251 | 6.7% | |
| transferase (PC00220) | 240 | 6.4% | |
| cytoskeletal protein (PC00085) | 192 | 5.2% | |
| enzyme modulator (PC00095) | 179 | 4.8% | |
| oxidoreductase (PC00176) | 149 | 4.0% | |
| extracellular matrix protein (PC00102) | 84 | 2.3% | |
| membrane traffic protein (PC00150) | 83 | 2.2% | |
| ligase (PC00142) | 79 | 2.1% | |
| calcium-binding protein (PC00060) | 72 | 1.9% | |
| structural protein (PC00211) | 63 | 1.7% | |
| isomerase (PC00135) | 42 | 1.1% | |
| lyase (PC00144) | 36 | 1.0% | |
| defense/immunity protein (PC00090) | 35 | 0.9% | |
| cell adhesion molecule (PC00069) | 31 | 0.8% | |
| cell junction protein (PC00070) | 31 | 0.8% | |
| chaperone (PC00072) | 24 | 0.6% | |
| transfer/carrier protein (PC00219) | 19 | 0.5% | |
| transmembrane receptor regulatory/adaptor protein (PC00226) | 11 | 0.3% |
| Ontology Function | Type | No. Molecules | Percent (%) |
|---|---|---|---|
| Molecular function | binding (GO:0005488) | 6900 | 41.1% |
| catalytic activity (GO:0003824) | 5156 | 30.7% | |
| transporter activity (GO:0005215) | 1474 | 8.8% | |
| receptor activity (GO:0004872) | 1450 | 8.6% | |
| signal transducer activity (GO:0004871) | 895 | 5.3% | |
| structural molecule activity (GO:0005198) | 826 | 4.9% | |
| translation regulator activity (GO:0045182) | 53 | 0.3% | |
| antioxidant activity (GO:0016209) | 23 | 0.1% | |
| binding (GO:0005488) | 6900 | 41.1% | |
| Biological process | cellular process (GO:0009987) | 10968 | 28.1% |
| metabolic process (GO:0008152) | 6589 | 16.9% | |
| biological regulation (GO:0065007) | 4065 | 10.4% | |
| response to stimulus (GO:0050896) | 3467 | 8.9% | |
| developmental process (GO:0032502) | 3319 | 8.5% | |
| multicellular organismal process (GO:0032501) | 2954 | 7.6% | |
| cellular component organization or biogenesis (GO:0071840) | 2626 | 6.7% | |
| localization (GO:0051179) | 2125 | 5.4% | |
| immune system process (GO:0002376) | 975 | 2.5% | |
| biological adhesion (GO:0022610) | 898 | 2.3% | |
| locomotion (GO:0040011) | 668 | 1.7% | |
| reproduction (GO:0000003) | 289 | 0.7% | |
| rhythmic process (GO:0048511) | 28 | 0.1% | |
| growth (GO:0040007) | 27 | 0.1% | |
| Protein class | hydrolase (PC00121) | 2235 | 13.2% |
| nucleic acid binding (PC00171) | 1676 | 9.9% | |
| signaling molecule (PC00207) | 1598 | 9.5% | |
| transcription factor (PC00218) | 1551 | 9.2% | |
| enzyme modulator (PC00095) | 1462 | 8.7% | |
| receptor (PC00197) | 1324 | 7.8% | |
| cytoskeletal protein (PC00085) | 987 | 5.9% | |
| transferase (PC00220) | 987 | 5.9% | |
| transporter (PC00227) | 977 | 5.8% | |
| oxidoreductase (PC00176) | 667 | 4.0% | |
| extracellular matrix protein (PC00102) | 632 | 3.7% | |
| cell adhesion molecule (PC00069) | 560 | 3.3% | |
| calcium-binding protein (PC00060) | 362 | 2.1% | |
| membrane traffic protein (PC00150) | 331 | 2.0% | |
| cell junction protein (PC00070) | 307 | 1.8% | |
| defense/immunity protein (PC00090) | 278 | 1.6% | |
| ligase (PC00142) | 185 | 1.1% | |
| structural protein (PC00211) | 173 | 1.0% | |
| chaperone (PC00072) | 157 | 0.9% | |
| transmembrane receptor regulatory/adaptor protein (PC00226) | 147 | 0.9% | |
| lyase (PC00144) | 134 | 0.8% | |
| isomerase (PC00135) | 66 | 0.4% | |
| transfer/carrier protein (PC00219) | 63 | 0.4% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Irimie, A.I.; Braicu, C.; Cojocneanu, R.; Magdo, L.; Onaciu, A.; Ciocan, C.; Mehterov, N.; Dudea, D.; Buduru, S.; Berindan-Neagoe, I. Differential Effect of Smoking on Gene Expression in Head and Neck Cancer Patients. Int. J. Environ. Res. Public Health 2018, 15, 1558. https://doi.org/10.3390/ijerph15071558
Irimie AI, Braicu C, Cojocneanu R, Magdo L, Onaciu A, Ciocan C, Mehterov N, Dudea D, Buduru S, Berindan-Neagoe I. Differential Effect of Smoking on Gene Expression in Head and Neck Cancer Patients. International Journal of Environmental Research and Public Health. 2018; 15(7):1558. https://doi.org/10.3390/ijerph15071558
Chicago/Turabian StyleIrimie, Alexandra Iulia, Cornelia Braicu, Roxana Cojocneanu, Lorand Magdo, Anca Onaciu, Cristina Ciocan, Nikolay Mehterov, Diana Dudea, Smaranda Buduru, and Ioana Berindan-Neagoe. 2018. "Differential Effect of Smoking on Gene Expression in Head and Neck Cancer Patients" International Journal of Environmental Research and Public Health 15, no. 7: 1558. https://doi.org/10.3390/ijerph15071558
APA StyleIrimie, A. I., Braicu, C., Cojocneanu, R., Magdo, L., Onaciu, A., Ciocan, C., Mehterov, N., Dudea, D., Buduru, S., & Berindan-Neagoe, I. (2018). Differential Effect of Smoking on Gene Expression in Head and Neck Cancer Patients. International Journal of Environmental Research and Public Health, 15(7), 1558. https://doi.org/10.3390/ijerph15071558