How Much Should We Involve Genetic and Environmental Factors in the Risk Assessment of Mycotoxins in Humans?

Abstract
:Introduction
Materials and Methods
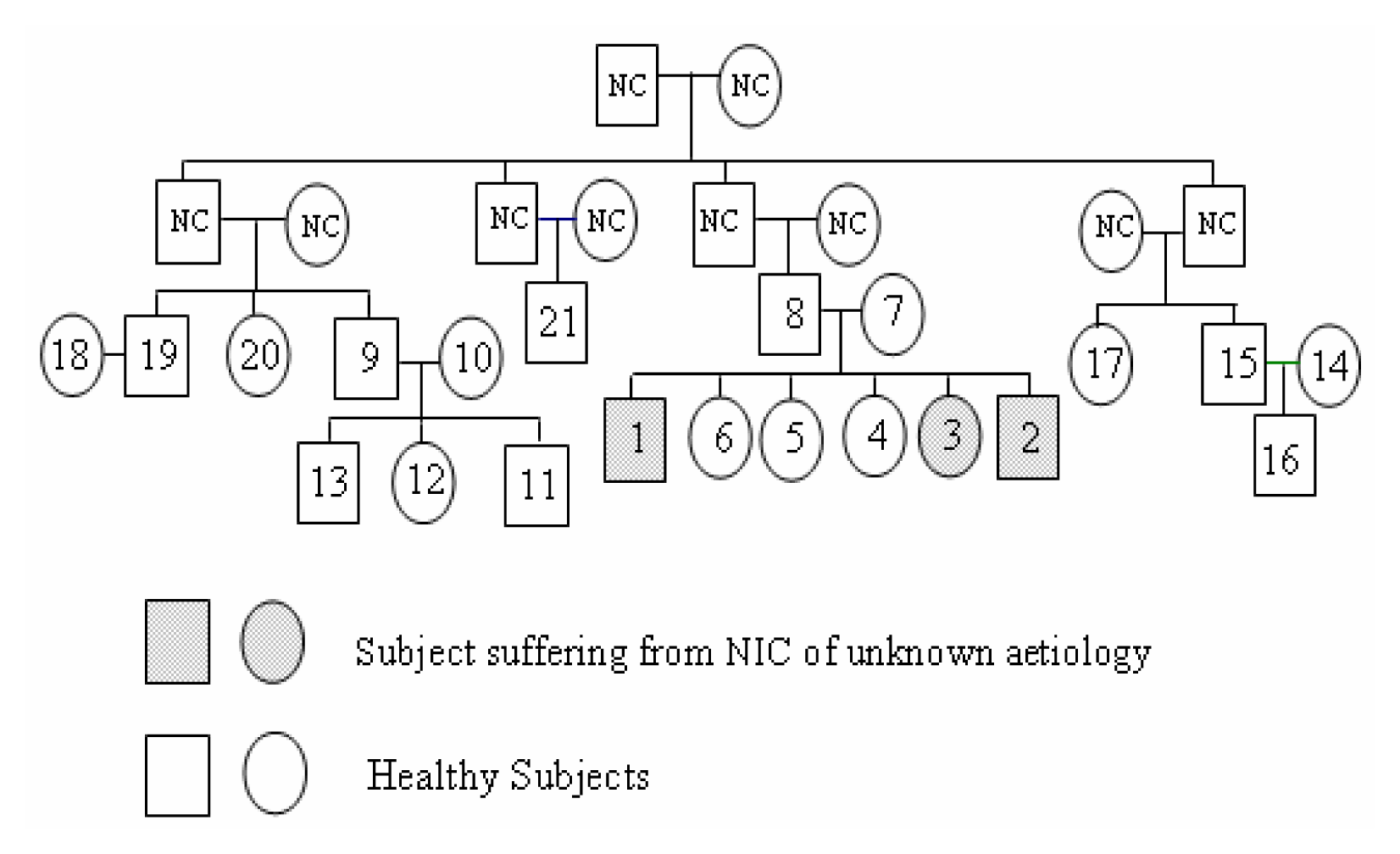
The household
Diagnosis of Chronic Interstitial Nephropathy of Unknown Aetiology
Bio Analysis
β2-Microglobulinuria Assay on Urines
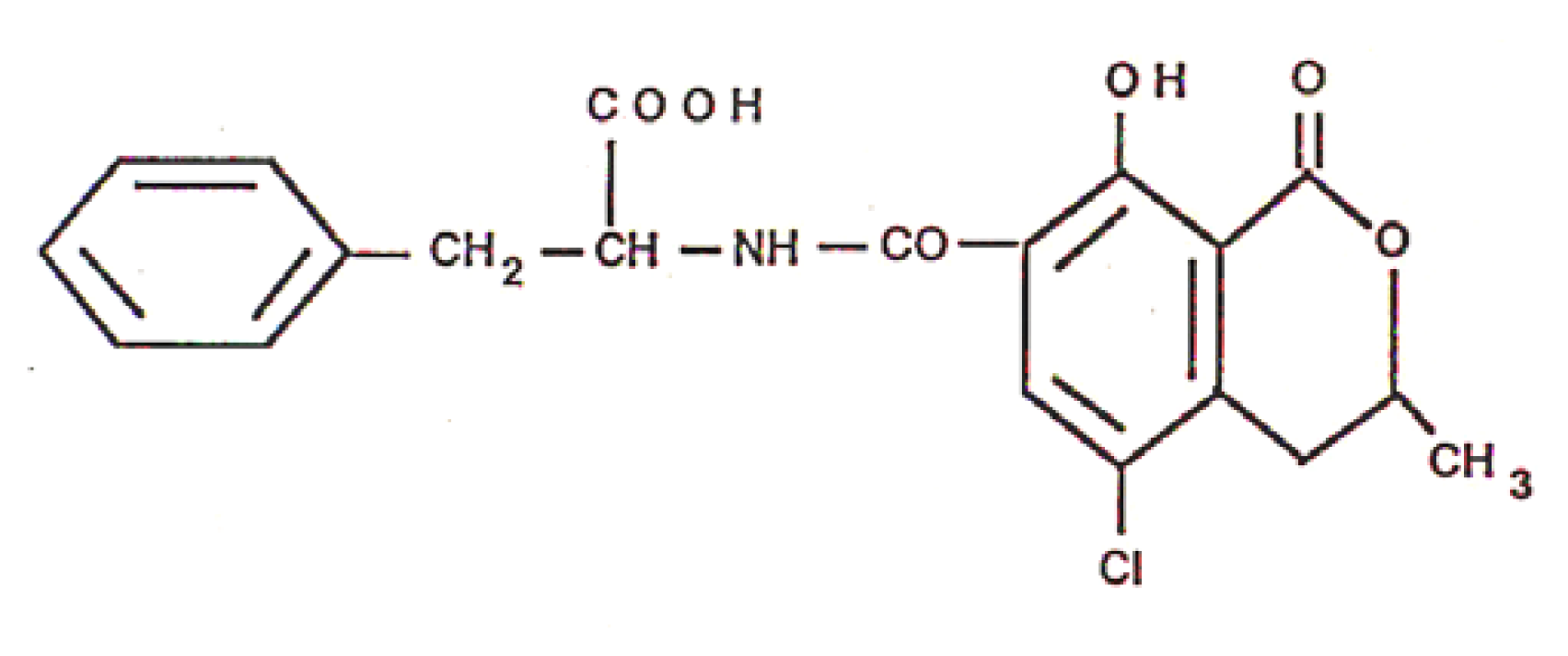
Analysis of Ochratoxin A in Blood, Urine and Food Samples
Histology
HLA Antigens Determination
Results
Presence of Ochratoxin A in Food, Blood and Urine
Genetic Imprinting of HLA
Discussion



{kind=link}
{kind=link}
| Case N° | OTA in Food μg/kg | OTA in Blood, ng/ml (OTA in Urine, ng/ml) | β2-Microglobulinuria μg/l | CIN of Unknown Etiology | |
|---|---|---|---|---|---|
| Family I | 1 | 0.88 ± 0.45 | 505.83 (10.18) | 10400 | + |
| 2 | 102.36 (94.4) | 440 | + | ||
| 3 | 1023 (limit) | 360 | + | ||
| 4 | 378 (limit) | 230 | − | ||
| 5 | 20.4 (limit) | 260 | − | ||
| 6 | 0 | 220 | − | ||
| 7 | 1332 (limit) | 300 | − | ||
| 8 | 15 (limit) | 850 | − | ||
| Family II | 9 | 5.29 ±7.19 | 1348 (limit) | 300 | − |
| 10 | 702(limit) | 260 | − | ||
| 11 | 1334 (41.5) | 1300 | − | ||
| 12 | 8 (limit) | 240 | − | ||
| 13 | 15 (limit) | 230 | − | ||
| Family III | 14 | 0.94 ± 0.45 | 967 (limit) | 225 | − |
| 15 | 218 (limit) | 700 | − | ||
| 16 | 210 (1000) | 250 | − | ||
| 17 | 33 (limit) | 210 | − | ||
| 18 | 0.31 ± 0.21 | 0 | 260 | − | |
| 19 | 9 (limit) | 280 | − | ||
| 20 | NC | 268 (limit) | 310 | − | |
| 21 | 8 (limit) | 300 | − | ||
| Case N° | OTA in Food μg/kg | OTA in Blood/ng/ml | OTA Daily intake ng/kg bw/Day | CIN BEN-Like | HLA Determinant | |
|---|---|---|---|---|---|---|
| Family I | 1 | 0.88 ± 0.45 | 505.8 | 658 | + | A3/28; B 27/35; DR 7/52 |
| 2 | 102.4 | 133 | + | A3/28; B 27/35; DR 7/11 | ||
| 3 | 1023 | 1330 | + | A3/40; B 27/35; DR 7/52 | ||
| 4 | 378 | 491 | − | |||
| 5 | 20.4 | 27 | − | |||
| 6 | 0 | 0 | − | |||
| 7 | 1332 | 1732 | − | |||
| 8 | 15 | 20 | − | |||
| Family II | 9 | 5.29 ±7.19 | 1348 | 1752 | − | |
| 10 | 702 | 912.6 | − | |||
| 11 | 1334 | 1734 | − | |||
| 12 | 8 | 10.4 | − | |||
| 13 | 15 | 20 | − | |||
| Family III | 14 | 0.94 ± 0.45 | 967 | 1257 | − | |
| 15 | 218 | 283 | − | |||
| 16 | 210 | 273 | − | |||
| 17 | 33 | 42.9 | − | |||
| 18 | 0.31 ± 0.21 | 0 | 0 | − | ||
| 19 | 9 | 12 | − | |||
| 20 | ND | 268 | 348 | − | ||
| 21 | 8 | 10 | − | |||
References
- Van der Merwe, K. J.; Steyn, P. S.; Fourie, L.; Scott, D. B.; Theron, J. J. Ochratoxin A, a toxic metabolite produced by Aspergillus ochraceus Wilh. Nature 1965, 205, 1112–1113. [Google Scholar]
- Creppy, E. E. Update of survey, regulation and toxic effects of mycotoxins in Europe. Toxicol. Lett 2002, 127, 19–28. [Google Scholar]
- Krogh, P. Role of ochratoxin in disease causation. Food Chem. Toxicol 1992, 30, 213–224. [Google Scholar]
- Kuiper-Goodman, T.; Scott, P. M. Risk assessment of the mycotoxin ochratoxin A. Biomed. Environ. Sci 1989, 2, 179–248. [Google Scholar]
- Abid, S.; Hassen, W.; Achour, A.; Skhiri, H.; Maaroufi, K.; Ellouz, F.; Creppy, E.; Bacha, H. Ochratoxin A and human chronic nephropathy in Tunisia: is the situation endemic? Hum. Exp. Tox 2003, 22, 77–84. [Google Scholar]
- McGlynn, K. A.; Hunter, K.; LeVoyer, T.; Roush, J.; Wise, P.; Michielli, R. A.; Shen, F. M.; Evans, A. A.; London, W. T.; Buetow, K. H. Susceptibility to aflatoxin B1-related primary hepatocellular carcinoma in mice and humans. Cancer Res 2003, 63, 4594–4601. [Google Scholar]
- Hassen, W.; Abid, S.; Achour, A.; Maaroufi, K.; Creppy, E.; Bacha, H. Ochratoxin A and human nephropathy in Tunisia: a ten year survey. Ann. Toxicol. Anal 2003, 15, 21–29. [Google Scholar]
- Ueno, Y. The toxicology of mycotoxins. Crit. Rev. Toxicol 1985, 14, 99–132. [Google Scholar]
- Creppy, E. E.; Kane, A.; Dirheimer, G.; Lafarge-Frayssinet, C.; Mousset, S.; Frayssinet, C. Genotoxicity of ochratoxin A in mice: DNA single-strand break evaluation in spleen, liver and Kidney. Toxicol. Lett 1985, 28, 29–35. [Google Scholar]
- Tiemersma, E. W.; Omer, R. E.; Bunschoten, A.; van’t Veer, P.; Kok, F. J.; Idris, M. O.; Kadaru, A. M.; Fedail, S. S.; Kampman, E. Role of genetic polymorphism of glutathione-S-transferase T1 and microsomal epoxide hydrolase in aflatoxin-associated hepatocellular carcinoma. Cancer Epidemiol Biomarkers Prev 2001, 10, 785–791. [Google Scholar]
- Simarro-Doorten, A. Y.; Bull, S.; van der Doelen, M. A. M.; Fink-Gremmels, J. Metabolism-mediated cytotoxicity of ochratoxin A. Toxicology in Vitro 2004, 18, 271–277. [Google Scholar]
- Creppy, E. E.; Chiarappa, P.; Baudrimont, I.; Borraci, P.; Moukha, S.; Carratu, M. R. Synergistic effects of fumonisin B1 and ochratoxin A: are in vitro cytotoxicity data predictive of in vivo acute toxicity? Toxicology 2004, 201, 115–123. [Google Scholar]
- FAO/WHO Expert Committee on Food Additives. Evaluation of certain mycotoxins in food. Fifty-sixth report of the Joint World Health Organ. Tech. Rep. Ser 2002, 906, 1–62.
- Mobio, T.A.; Anane, R.; Baudrimont, I.; Carratu, M.R.; Shier, T.W.; Dano, S.D.; Ueno, Y.; Creppy, E.E. Epigenetic properties of fumonisin B1: cell cycle arrest and DNA base modification in C6 glioma cells. Toxicol Appl Pharmacol 2000, 164, 91–96. [Google Scholar]
- Horvath, A.; Upham, B. L.; Ganev, V.; Trosko, J. E. Determination of the epigenetic effects of ochratoxin in a human kidney and a rat liver epithelial cell line. Toxicon 2002, 40, 273–82. [Google Scholar]
- Assaf, H.; Azouri, H.; Pallardy, M. Ochratoxin A induces apoptosis in human lymphocytes through down regulation of Bcl-xL. Toxicol Sci 2004, 79, 335–344. [Google Scholar]
- Lühe, A.; Hildebrand, H.; Bach, U.; Dingermann, T.; Ahr, H-J. A new approach to studying ochratoxin A(OTA)-induced nephrotoxicity: expression profiling in vivo and in vitro employing cDNA microarrays. Toxico. Sci 2003, 73, 315–328. [Google Scholar]
- Ouanes, Z.; Abid, S.; Ayed, I.; Anane, R.; Mobio, T.; Creppy, E. E.; Bacha, H. Induction of micronuclei by zearalenone in Vero monkey kidney cells and in bone marrow cells of mice: protective effect of vitamin E. Mut Res 2003, 538, 63–70. [Google Scholar]
- Stoloff, L.; Van Egmond, H. P.; Park, D. L. Rationales for the establishment of limits and regulations for mycotoxins in mammals. Food Addit Contam 1999, 36, 175–181. [Google Scholar]
- Pfohl-Leszkowicz, A.; Grosse, Y.; Castegnaro, M.; Nicolov, I. G.; Chernozemsky, I. N.; Bartsch, H.; Betbeder, A. M.; Creppy, E. E.; Dirheimer, G. Ochratoxin A-related DNA adducts in urinary tract tumours of Bulgarian subjects. IARC Sci Publ 1993, 124, 141–148. [Google Scholar]
- Hassen, W.; Abid, S.; Achour, A.; Creppy, E.; Bacha, H. Ochratoxin A and β2-microglobulinuria in healthy individuals and in chronic interstitial nephropathy patients in the centre of Tunisia: a hot spot of Ochratoxin A exposure. Toxicology 2004, 199, 185–93. [Google Scholar]
- Baudrimont, I.; Betbeder, A-M.; Creppy, E. E. Protective effect of aspartame on the karyomegaly induced by ochratoxin a in rat kidney. Toxicol Lett 1996, 88, 21–22. [Google Scholar]
- Mihatsch, M. J.; Gudat, F.; Zollinger, H. U.; Heierli, C.; Tholen, H.; Reutter, F. W. Systemic karyomegaly associated with chronic interstitial nephritis. A new disease entity? Clin Nephrol 1979, 12, 54–62. [Google Scholar]
- Spoendlin, M.; Moch, H.; Brunner, F.; Brunner, W.; Burger, H. R.; Kiss, D.; Wegmann, W.; Dalquen, P.; Oberholzer, M.; Thiel, G. Karyomegalic interstitial nephritis: further support for a distinct entity and evidence for a genetic defect. Am J Kidney Dis 1995, 25, 242–252. [Google Scholar]
- Godin, M.; Francois, A.; Le Roy, F.; Morin, J. P.; Creppy, E. E.; Hemet, J.; Fillastre, J. P. Karyomegalic interstitial nephritis. Am J Kidney Dis 1996, 27(1), 166. [Google Scholar]
- Hanada, K.; Yewdell, J. W.; Yang, J. C. Immune recognition of a human renal cancer antigen through post-translational protein splicing. Nature 2004, 427, 252–256. [Google Scholar]
- Morozov, V. N.; Morozova, T.; Bray, P.; Hranisavljevic, J.; Vucelic, D. Survey of small molecule and ion binding to beta 2-microglobulin--possible relation to BEN. Kidney Int Suppl 1991, 34, S85–88. [Google Scholar]
- Andonova, I. E.; Sarueva, R. B.; Horvath, A. D.; Simeonov, V. A.; Dimitrov, P. S.; Petropoulos, E. A.; Ganev, V. S. Balkan endemic nephropathy and genetic variants of glutathione S-transferases. J. Nephrol 2004, 17, 390–398. [Google Scholar]
- Gautier, J. P.; Holzhaeuser, D.; Markovic, J.; Germaud, E.; Schilter, B.; Ad Turesky, R. J. Oxidative damage and stress response from ochratoxin A exposure in rats. Free Radical Biology and Medicine 2001, 30, 1089–1098. [Google Scholar]
© 2005 MDPI. All rights reserved.
Share and Cite
Creppy, E.E.; Moukha, S.; Bacha, H.; Carratu, M.R. How Much Should We Involve Genetic and Environmental Factors in the Risk Assessment of Mycotoxins in Humans? Int. J. Environ. Res. Public Health 2005, 2, 186-193. https://doi.org/10.3390/ijerph2005010186
Creppy EE, Moukha S, Bacha H, Carratu MR. How Much Should We Involve Genetic and Environmental Factors in the Risk Assessment of Mycotoxins in Humans? International Journal of Environmental Research and Public Health. 2005; 2(1):186-193. https://doi.org/10.3390/ijerph2005010186
Chicago/Turabian StyleCreppy, Edmond E., Serge Moukha, Hassen Bacha, and Maria Rosaria Carratu. 2005. "How Much Should We Involve Genetic and Environmental Factors in the Risk Assessment of Mycotoxins in Humans?" International Journal of Environmental Research and Public Health 2, no. 1: 186-193. https://doi.org/10.3390/ijerph2005010186
APA StyleCreppy, E. E., Moukha, S., Bacha, H., & Carratu, M. R. (2005). How Much Should We Involve Genetic and Environmental Factors in the Risk Assessment of Mycotoxins in Humans? International Journal of Environmental Research and Public Health, 2(1), 186-193. https://doi.org/10.3390/ijerph2005010186