Improved Buffering Capacity and Methane Production by Anaerobic Co-Digestion of Corn Stalk and Straw Depolymerization Wastewater


Abstract
:
1. Introduction
2. Results and Discussion
2.1. Methane Production
2.2. Variation of SCOD and Sulfate Concentrations
2.3. Variation in VFA Concentration and pH
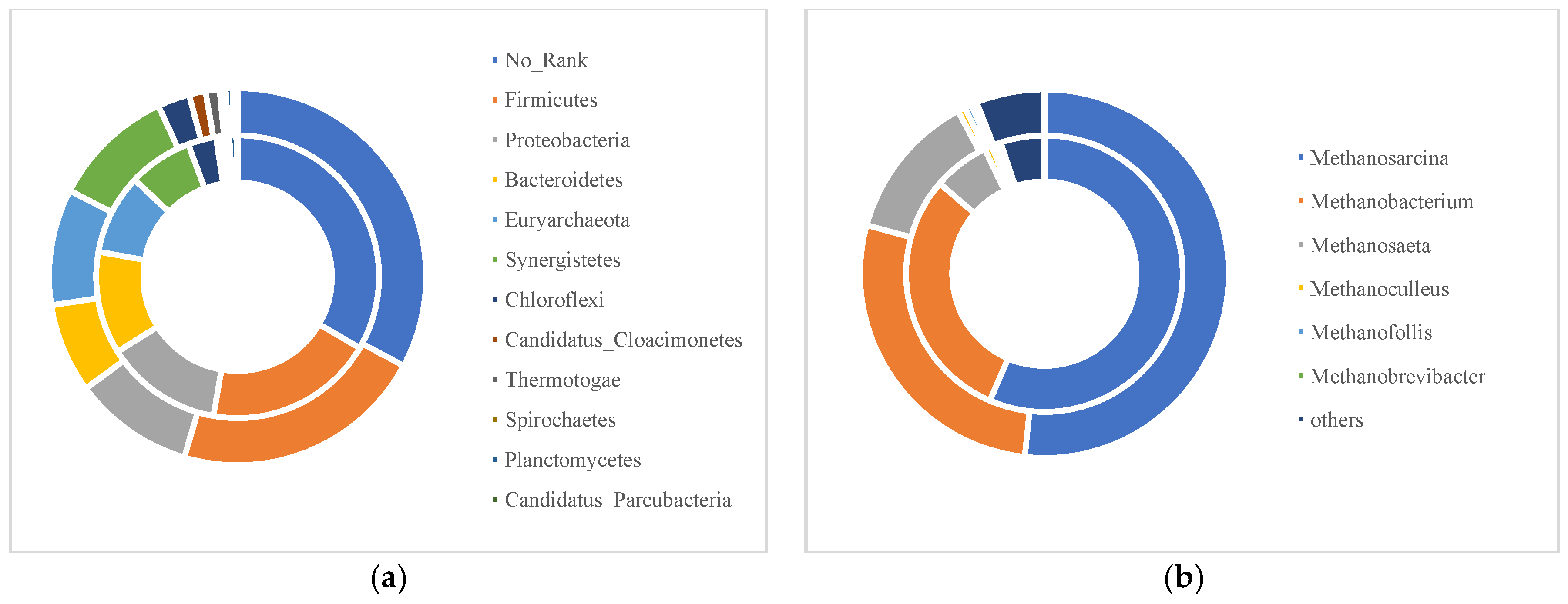
2.4. Analysis of Microbial Communities
3. Materials and Methods
3.1. Feedstocks and Inoculum
3.2. Experimental Setup and Procedure
3.3. Analytical Methods
4. Conclusions
- (1)
- Anaerobic co-digestion of CS and SDW can significantly increase the buffering capacity and methane production. The lag time for methane generation from the anaerobic co-digestion of CS and SDW was 10 days, however, the lag time for mono-digestion of CS was 23 days for a substrate concentration of 36.2 g/L. Maximum methane production from anaerobic co-digestion of CS and SDW of 214.81 mL/g-VS was about 7.76% higher than for mono-digestion of CS which gave a value of 199.34 mL/g-VS. Increasing the substrate concentration to 45.3 g/L resulted in methanogenic processes being significantly inhibited during the initial anaerobic digestion stage. However, the fermentation performance and buffering capacity observed for anaerobic co-digestion of CS and SDW were significantly better than levels obtained for mono-digestion of CS.
- (2)
- The removal rate of sulfate increased from 10.43% to 58.40% when the substrate concentration was increased from 25.9 to 45.3 mg/L for anaerobic co-digestion of CS and SDW.
- (3)
- 16S rDNA sequencing results showed that anaerobic co-digestion of CS and SDW promoted growth of methanogens, the relative abundance of Euryarchaeota (mainly Methanosarcina, Methanobacterium, and Methanosaeta) for anaerobic co-digestion of CS and SDW being 9.84%, which was 8.25% higher than the 9.09% of Euryarchaeota present in mono-digestion of CS. SOB and SRB could both improve mutual transformation of sulfate and sulfide, as well as increase the overall removal rate of organic matter.
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Guo, P.; Mochidzuki, K.; Cheng, W.; Zhou, M.; Gao, H.; Zheng, D. Effects of different pretreatment strategies on corn stalk acidogenic fermentation using a microbial consortium. Bioresour. Technol. 2011, 102, 7526–7531. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wu, S.B.; Zhang, W.Q.; Pang, C.; Deng, Y.; Dong, R.J. Effect of White-rot Fungi Pretreatment on Methane Production from Anaerobic Digestion of Corn Stover. Trans. Chin. Soc. Agric. Mach. 2013, 44, 124–129. [Google Scholar]
- Zhu, J.Y.; Wan, C.X.; Li, Y. Enhanced solid-state anaerobic digestion of corn stover by alkaline pretreatment. Bioresour. Technol. 2010, 101, 7523–7528. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhao, Y.; Liu, B.; Zhao, Y.; Wu, J.; Yuan, X. Accelerated acidification by inoculation with a microbial consortia in a complex open environment. Bioresour. Technol. 2016, 216, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Zhong, M.; Duan, N.; Lin, C.; Zhang, D.; Liang, S.; Sun, H. Effects of organic loading rate and additive on corn stalk anaerobic digestion. J. Biobased Mater. Bioenergy 2016, 10, 219–224. [Google Scholar] [CrossRef]
- Chen, G.; Zheng, Z.; Yang, S.; Fang, C.; Zou, X.; Luo, Y. Experimental co-digestion of corn stalk and vermicompost to improve biogas production. Waste Manag. 2010, 30, 1834–1840. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Zhang, Z.; Luo, Y.; Qiao, W.; Xiao, M.; Zhang, M. Biogas productivity by co-digesting taihu blue algae with corn straw as an external carbon source. Bioresour. Technol. 2012, 114, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.T.; Wang, B.H.; Han, H.J. Analysis and Progress in Treatment of Furfural Wastewater. Liaoning Chem. Ind. 2010, 39, 862–865. [Google Scholar]
- Wang, F.; Xuan, C.; Ma, Y.; Sun, Y.; Wu, Q. Products and mechanism of corn stalk after depolymerization with acid. J. Chin. Cereals Oils Assoc. 2015, 30, 1–5. [Google Scholar]
- Xu, Z.; Li, W.; Du, Z.; Wu, H.; Jameel, H.; Chang, H.M. Conversion of corn stalk into furfural using a novel heterogeneous strong acid catalyst in γ-valerolactone. Bioresour. Technol. 2015, 198, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, Y.Y.; Qiao, W.; Wang, X.; Takayanagi, K. Sulfate addition as an effective method to improve methane fermentation performance and propionate degradation in thermophilic anaerobic co-digestion of coffee grounds, milk and waste activated sludge with anmbr. Bioresour. Technol. 2015, 185, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Cheng, J.J.; Creamer, K.S. Inhibition of anaerobic digestion process: A review. Bioresour. Technol. 2008, 99, 4044–4064. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.L.; Wang, J.; Yue, Z.B.; Tao, W.; Yang, H.B.; Zhou, Y.F.; Chen, T.H. Simultaneous chemical oxygen demand removal, methane production and heavy metal precipitation in the biological treatment of landfill leachate using acid mine drainage as sulfate resource. J. Biosci. Bioeng. 2017, 124, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Ren, N.Q.; Wang, A.J.; Zhao, Y.G. Ecology of Sulfate-Reducing Bacteria in Anaerobic Biotreatment Processes; SciencePress: Beijing, China, 2009; pp. 132–133. [Google Scholar]
- Barton, L.L. Sulfate-Reducing Bacteria; Plenum Press: London, UK; Springer: New York, NY, USA, 1995; pp. 150–180. [Google Scholar]
- Zhou, X.F.; Ren, N.Q. Acid resistance of methanogenic bacteria in two—Stage anaerobic process treating high concentration methanol waste water. Acta Sci. Circumst. 2004, 24, 633–636. [Google Scholar]
- Preez, L.A.D.; Odendaal, J.P.; Maree, J.P.; Ponsonby, M. Biological removal of sulphate from industrial effluents using producer gas as energy source. Environ. Technol. Lett. 1992, 13, 875–882. [Google Scholar] [CrossRef]
- Peng, J.W.; Kang, C.L.; Cui, Y.B.; Liu, X.C.; Han, X.K. Application on Wastewater Treatment of Produced Furfural by Free Water Surface Flow Constructed Wetland. J. Jilin Univ. (Earth Sci. Ed.) 2010, 40, 1419–1424. [Google Scholar]
- Zou, X.N.; Tan, H.L.; Zhan, B.; He, J.L. Study on Treatment of Furfural Wastewater by Double-effect Evaporation Process. Ind. Saf. Environ. Prot. 2008, 34, 1–4. [Google Scholar]
- Qiao, W.; Takayanagi, K.; Shofie, M.; Niu, Q.; Yu, H.Q.; Li, Y.Y. Thermophilic anaerobic digestion of coffee grounds with and without waste activated sludge as co-substrate using a submerged AnMBR: System amendments and membrane performance. Bioresour. Technol. 2013, 150, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Vela, F.J.; Zaiat, M.; Foresti, E. Influence of the COD to sulphate ratio on theanaerobic organic matter degradation kinetics. Water SA 2002, 28, 213–216. [Google Scholar] [CrossRef]
- Speece, R.E. Anaerobic biotechnology for industrial wastewater treatment. Environ. Sci. Technol. 1983, 17, 416–427. [Google Scholar] [CrossRef] [PubMed]
- Ward, D.M.; Olson, G.J. Terminal processes in the anaerobic degradation of an algal-bacterial mat in a high-sulfate hot spring. Appl. Environ. Microbiol. 1980, 40, 67–74. [Google Scholar] [PubMed]
- Li, Y.Y.; Lam, S.; Fang, H.H.P. Interactions between methanogenic, sulfatereducing and syntrophic acetogenic bacteria in the anaerobic degradation of benzoate. Water Res. 1996, 30, 1555–1562. [Google Scholar] [CrossRef]
- Muyzer, G.; Stams, A. The ecology and biotechnology of sulphate-reducing bacteria. Rev. Microbiol. 2008, 6, 441–454. [Google Scholar] [CrossRef] [PubMed]
- Gevertz, D.; Telang, A.J.; Voordouw, G.; Jenneman, G.E. Isolation and Characterization of Strains CVO and FWKOB, Two Novel Nitrate-Reducing, Sulfide-Oxidizing Bacteria Isolated from Oil Field Brine. Appl. Environ. Microbiol. 2000, 66, 2491–2501. [Google Scholar] [CrossRef] [PubMed]
- Beristain-Cardoso, R.; Texier, A.C.; Alpuche-Solís, A.; Gómez, J.; Razo-Flores, E. Phenol and sulfide oxidation in a denitrifying biofilm reactor and its microbial community analysis. Process Biochem. 2009, 44, 23–28. [Google Scholar] [CrossRef]
- Haveman, S.A.; Greene, E.A.; Stilwell, C.P.; Voordouw, J.K.; Voordouw, G. Physiological and gene expression analysis of inhibition of desulfovibrio vulgaris hildenborough by nitrite. J. Bacteriol. 2004, 186, 7944–7950. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Zhao, M.; Chen, Y.; Yu, M.; Ruan, W. Tolerance response to in situ ammonia stress in a pilot-scale anaerobic digestion reactor for alleviating ammonia inhibition. Bioresour. Technol. 2015, 198, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Demirel, B.; Scherer, P. The roles of acetotrophic and hydrogenotrophic methanogens during anaerobic conversion of biomass to methane: a review. Rev. Environ. Sci. Biotechnol. 2008, 7, 173–190. [Google Scholar] [CrossRef]
- Riviere, D.; Desvignes, V.; Pelletier, E.; Chaussonnerie, S.; Guermazi, S.; Weissenbach, J.; Li, T.; Camacho, P.; Sghir, A. Towards the definition of a core of microorganisms involved in anaerobic digestion of sludge. ISME J. 2009, 3, 700–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sieber, J.R.; Mcinerney, M.J.; Gunsalus, R.P. Genomic insights into syntrophy: The paradigm for anaerobic metabolic cooperation. Annu. Rev. Microbiol. 2012, 66, 429–452. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.M.; Kong, X.Y.; Xing, T.; Sun, Y.M.; Zhang, Y.; Luo, X.J.; Sun, Y. Digestion performance and microbial metabolic mechanism in thermophilic and mesophilic anaerobic digesters exposed to elevated loadings of organic fraction of municipal solid waste. Energies 2018, 11, 952. [Google Scholar] [CrossRef]
- Kindaichi, T.; Ito, T.; Okabe, S. Ecophysiological interaction between nitrifying bacteria and heterotrophic bacteria in autotrophic nitrifying biofilms as determined by microautoradiography-fluorescence in situ hybridization. Appl. Environ. Microbiol. 2004, 70, 1641–1650. [Google Scholar] [CrossRef] [PubMed]
- Moran, J.J.; House, C.H.; Freeman, K.H.; Ferry, J.G. Trace methane oxidation studied in several euryarchaeota under diverse conditions. Archaea 2005, 1, 303. [Google Scholar] [CrossRef] [PubMed]
- Kern, T.; Linge, M.; Rother, M. Methanobacterium aggregans sp nov., a hydrogenotrophic methanogenic archaeon isolated from an anaerobic digester. Int. J. Syst. Evol. Microbiol. 2015, 65, 1975–1980. [Google Scholar] [CrossRef] [PubMed]
- De Lucena, R.M.; Gavazza, S.; Florencio, L.; Kato, M.T.; De Morais, M.A. Study of the microbial diversity in a full-scale UASB reactor treating domestic wastewater. World J. Microbiol. Biotechnol. 2011, 27, 2893–2902. [Google Scholar] [CrossRef]
- Kodama, Y.; Watanabe, K. Isolation and characterization of a sulfur-oxidizing chemolithotroph growing on crude oil under anaerobic conditions. Appl. Environ. Microbiol. 2003, 69, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Claudia, G.; Alberto, O.; Claudio, G.; Selene, M.; Fernando, B.; Paola, B. Enhanced sulfate reduction and trichloroethylene (TCE) biodegradation in a UASB reactor operated with a sludge developed from hydrothermal vents sediments: process and microbial ecology. Int. Biodeterior. Biodegrad. 2014, 94, 182–191. [Google Scholar]
- Wu, S.J.; Dang, Y.; Qiu, B.; Liu, Z.; Sun, D.Z. Effective treatment of fermentation wastewater containing high concentration of sulfate by two-stage expanded granular sludge bed reactors. Int. Biodeterior. Biodegrad. 2015, 104, 15–20. [Google Scholar] [CrossRef]
- Kiyuna, L.S.; Fuess, L.T.; Zaiat, M. Unraveling the influence of the COD/sulfate ratio on organic matter removal and methane production from the biodigestion of sugarcane vinasse. Bioresour. Technol. 2017, 232, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Luo, L.; Li, W.; Wang, X.; Sun, Y.; Sun, Y. Optimization of mixing ratio of ammoniated rice straw and food waste co-digestion and impact of trace element supplementation on biogas production. J. Mater. Cycles Waste Manag. 2017, 2, 1–9. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate Concentration | Methane Production | ||||
|---|---|---|---|---|---|
| P∞ a | Rmb | λc | R2 | ||
| 25.9 g/L (Figure 1a) | co-digestion of CS with SDW | 181.18 ± 3.73 | 32.57 ± 0.92 | 0.69 ± 0.02 | 0.9908 ± 0.0034 |
| mono-digestion of CS | 183.97 ± 4.32 | 21.38 ± 1.61 | 0.21 ± 0.00 | 0.9883 ± 0.0011 | |
| 36.2 g/L (Figure 1b) | co-digestion of CS with SDW | 214.81 ± 1.89 | 10.86 ± 0.92 | 5.23 ± 0.06 | 0.9750 ± 0.0023 |
| mono-digestion of CS | 189.20 ± 3.69 | 6.21 ± 0.51 | 23.82 ± 0.99 | 0.9465 ± 0.0072 | |
| 45.9 g/L (Figure 1c) | co-digestion of CS with SDW | 171.62 ± 1.43 | 4.66 ± 0.02 | 27.32 ± 1.78 | 0.9285 ± 0.0102 |
| mono-digestion of CS | 139.18 ± 0.68 | 5.61 ± 0.42 | 35.69 ± 3.62 | 0.9061 ± 0.0003 | |
| Parameters | SDW | Corn Stalk | Inoculum |
|---|---|---|---|
| TS (%) | 4.87 ± 0.02 | 94.07 ± 0.50 | 4.46 ± 0.02 |
| VS (%) | 1.66 ± 0.01 | 88.95 ± 0.91 | 3.12 ± 0.01 |
| pH | 2.25 ± 0.01 | - | 6.94 ± 0.01 |
| SCOD (g/L) | 12.15 ± 0.77 | - | 3.30 ± 0.21 |
| Sulphate (g/L) | 1.94 ± 0.10 | - | 0.01 ± 0.00 |
| Nitrate (mg/L) | 3.02 ± 0.02 | - | 1.534 ± 0.02 |
| Methanol (g/L) | 0.124 ± 0.00 | - | - |
| Acetic acid (g/L) | 1.83 ± 0.02 | - | - |
| Propionic acid (g/L) | 0.14 ± 0.00 | - | - |
| Butyric acid (g/L) | 0.38 ± 0.00 | - | - |
| C (%) | 10.48 ± 0.07 | 38.63 ± 0.02 | 30.04 ± 0.04 |
| N (%) | 1.65 ± 0.00 | 0.78 ± 0.03 | 2.55 ± 0.10 |
| Cellulose (%) | - | 34.10 ± 0.11 | 11.59 ± 0.06 |
| Hemi-cellulose (%) | - | 28.36 ± 0.08 | 24.99 ± 0.00 |
| Lignin (%) | - | 5.57 ± 0.03 | 25.04 ± 0.01 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, F.; Li, W.; Sun, M.; Li, Q.; Wang, M.; Sun, Y. Improved Buffering Capacity and Methane Production by Anaerobic Co-Digestion of Corn Stalk and Straw Depolymerization Wastewater. Energies 2018, 11, 1751. https://doi.org/10.3390/en11071751
Yang F, Li W, Sun M, Li Q, Wang M, Sun Y. Improved Buffering Capacity and Methane Production by Anaerobic Co-Digestion of Corn Stalk and Straw Depolymerization Wastewater. Energies. 2018; 11(7):1751. https://doi.org/10.3390/en11071751
Chicago/Turabian StyleYang, Fuli, Wenzhe Li, Mingchao Sun, Qiang Li, Mengyi Wang, and Yong Sun. 2018. "Improved Buffering Capacity and Methane Production by Anaerobic Co-Digestion of Corn Stalk and Straw Depolymerization Wastewater" Energies 11, no. 7: 1751. https://doi.org/10.3390/en11071751