
Automatic Fed-Batch Cultivation Enhances Microbial Lipid Production from Volatile Fatty Acids

Gwangju Clean Energy Research Center, Korea Institute of Energy Research, Gwangju 61003, Republic of Korea
*
Author to whom correspondence should be addressed.
Energies 2023, 16(4), 1996; https://doi.org/10.3390/en16041996
Submission received: 11 January 2023
/
Revised: 3 February 2023
/
Accepted: 16 February 2023
/
Published: 17 February 2023
(This article belongs to the Collection Bioenergy and Biofuel)
Abstract
:Organic waste is generated worldwide, and its disposal and recycling are becoming a challenge. Due to its high carbon content, however, it may be converted into valuable products. Carbon neutrality is essential, and unstable international oil prices stress the increasing importance of biofuels significantly. Volatile fatty acids (VFA) derived from organic waste can be converted to microbial lipids by oleaginous yeast using it as a carbon source. When VFA is consumed by oleaginous yeast, the pH of the medium rises; hence, acidic agents have to be added to the medium to maintain the broth’s pH. In this study, we enhanced microbial lipid productivity by automatic fed-batch cultivation using VFA as an acidic agent, and the modified cultivation showed 48.9% and 69.0% higher biomass and lipid productivity than manual multi-fed culture. At a VFA concentration of 5 g/L and pH 7.0, a lipid yield of 0.25 g/g alongside lipid productivity of 0.11 g/L/h was obtained from an automatic fed-batch system. Oleic acid accounted for the largest proportion of microbial lipids, and the fatty acid composition was suitable for biodiesel production.
1. Introduction
Organic waste is one of the major concerns at present, and it has been increasing with the increase in population worldwide. According to the Food and Agriculture Organization (FAO), approximately 1.3 billion tons of food are lost and wasted every year, costing the world economy 750 billion dollars [1]. Wasted food is currently landfilled or incinerated, together with other combustible municipal wastes, for the generation of heat or energy in many countries [2]. However, both methods produce environmental stress and carbon emission [3]. Organic waste can also be utilized as a useful resource through various fermentation processes due to its nutrients; therefore, research is underway for using organic waste as a feedstock for producing biofuels, such as biogas, hydrogen, ethanol, and biodiesel, as a final product [4,5].
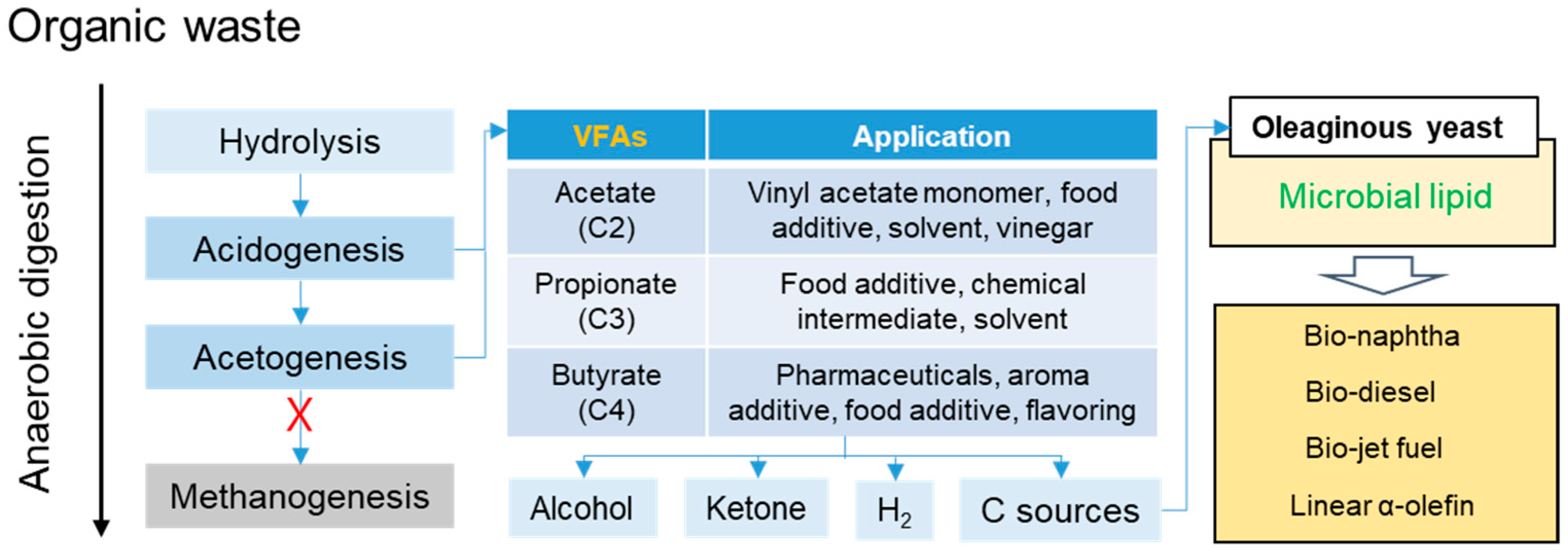
Organic waste can be converted to valuable products via anaerobic digestion (AD) of its high carbohydrate content. AD involves four steps, namely hydrolysis, acidogenesis, acetogenesis, and methanogenesis (Figure 1) [6]. The first step of AD involves the hydrolysis of organic matter, whereby organic material is broken down into smaller molecules. The next step of acidogenesis is the conversion of these small molecules to organic acids, carbon dioxide, and hydrogen. Acetic acid (AA), propionic acid (PA), butyric acid (BA), and ethanol are produced. Finally, methane and carbon dioxide are produced from acetate, ethanol, and other intermediates during the methanogenesis step [5]. The final product is methane, and the process is relatively slow for its high retention time. With the addition of methanogen inhibitors (chloroform, BES, iodoform, and β-cyclodextrin), volatile fatty acids (VFA) concentration can be determined during acidogenesis and acetogenesis. VFA is a short-chain carboxylic acid with AA, PA, and BA as the main components from AD. Each VFA has a number of industrial applications such as food additives, solvents, chemical intermediates, etc. However, the separation of mixed VFA in media broth into single VFA is a challenge for its azeotrope distillation and economic problem. Therefore, the VFA conversion research is focus on these days.
Considering VFA as an intermediate biofuel and biochemical, it is a versatile product that can play a crucial role in bioprocesses. VFA can be converted into mixed alcohols by the addition of hydrogen, ketone by thermal conversion, and hydrogen by VFA reforming. It can also be used as a carbon source by many microorganisms. The disadvantage of VFA is that it can be utilized by only some microbial cells, while glucose can be utilized by almost all microbial cells. However, oleaginous yeast can utilize VFA as a carbon source and accumulate lipids over 20% of its cell mass [7,8,9,10].
Oleaginous yeast can accumulate lipids through the conversion of a given substrate by either de novo synthesis or ex novo synthesis [11]. Additionally, oleaginous yeasts can convert organic acids directly into acetyl-CoA through acetyl coenzyme A synthetase. Acetyl-CoA is a building block of fatty acids used for lipid accumulation and ATP synthesis for biomass production in the TCA cycle. Furthermore, the microbial lipid is a potential platform chemical; it can be converted to biodiesel, linear alpha olefin, bio-naphtha, and jet fuel by further catalytic reaction (Figure 1) [12,13,14,15].
Microbial lipid is an intracellular product, and high cell density with high lipid production is important for the reduction of further downstream process costs. For high cell density culture, substrate feeding control would be essential to avoid substrate inhibition and increase cell mass and microbial lipid during cultivation [16]. Some studies have been published regarding the online measurement of in situ VFA concentration by mid-infrared spectroscopy [17,18]. Nevertheless, accurate analysis remains a challenge, and some indirect feedback methods for feeding strategies in fed-batch culture, including pH-stat, dissolved oxygen (DO)-stat, and exponential feeding, are known to exist [19]. Research on several feeding strategies for oleaginous yeast cultivation has been conducted using glucose, lingo-cellulosic biomass hydrolysate, and glycerol as carbon sources [20,21,22,23,24]. However, very few studies have shown VFA feeding strategies for microbial lipid production.
The current study aimed to develop a new feeding strategy for VFA as a sole carbon source. It is based on the fact that pH rises due to the consumption of VFA. The oleaginous yeast, Cryptococcus curvatus (currently named Cutaneotrichosporon oleaginosus), was cultivated for microbial lipid production at an acidogenesis stage pH environment (pH 5.5 and 7.0) and for automatic fed-batch cultivation with VFA solution for high cell density culture.
2. Materials and Methods
2.1. Microorganism
Cryptococcus curvatus (ATCC 20509, KCTC 7225) was obtained from the Korea Biological Resource Center (Daejeon, Republic of Korea). The strain was maintained by monthly subculture on potato dextrose agar (0.4% potato infusion, 2% dextrose, and 1.5% agar). Seed culture was cultivated for 1 day on 50 mL of liquid YPD medium (2% peptone, 1% yeast extract, and 2% dextrose) in a 250 mL flask at 25 °C.
2.2. Flask Culture Condition
Flask culture experiments were performed using a minimal medium (pH 5.5) containing the following (per liter of distilled water): 2.7 g KH2PO4, 0.95 g Na2HPO4, 0.2 g MgSO4·7H2O, 0.5 g yeast extract, and 0.1 g EDTA. The culture was supplemented with a 10 mL stock solution containing (per liter) 4 g CaCl2·2H2O, 0.55 g FeSO4·7H2O, 0.52 g citric acid·H2O, 0.10 g ZnSO4·7H2O, 0.076 g MnSO4·H2O, and 100 μL 18 M H2SO4. Glucose (30 g/L) was used as a carbon source, and NaNO3 or NH4Cl was added as a nitrogen source for a C/N ratio of 40. The cultivation conditions were 25 °C and 180 revolutions per minute (RPM).
2.3. Fed-Batch Cultivation in 5 L Fermenter
A 5 L fermenter (CNS, Republic of Korea), fitted with pH (FermProbe, Broadley James, Irvine, CA, USA) and DO electrodes (InPro 6820, Mettler toledo, Switzerland), was used for yeast cultivation. Two types of fed-batch culture experiments were conducted. One was a normal repeated fed-batch by manual feeding of VFA. pH was automatically controlled in manual fed-batch by adding 5 N HCl and 5 N NaOH and in automatic fed-batch by adding synthetic VFA solution and 5 N NaOH. VFA solution ratio was 6:1:3 (AA:PA:BA), which is typical of that fermented from food waste [5,25]. The medium composition was the same as that of the flask culture without carbon and nitrogen sources. NaNO3 was used as a nitrogen source and added at C/N of 5. VFA consumption was determined by weighing the difference using a balance. The vessel was sterilized by autoclaving at 121 °C for 15 min; 10% inoculum was transferred to the fermenter, and the working volume was 3 L. Cultures were grown at 25 °C with an aeration rate of 1 volume of air/volume of medium per minute (VVM) and DO concentration was maintained above 30% air saturation for increasing RPM.
2.4. Analysis
Optical density (OD) was measured at 600 nm using a UV spectrophotometer (UV-2600, Shimadzu, Kyoto, Japan). The yeast biomass concentration was determined by dry-cell weight (DCW) measurement [26]. DCW was then determined by harvesting and transferring the culture samples to a pre-weighed centrifuge tube after centrifugation. Subsequent to washing with distilled water, the centrifuge tubes were dried at 85 °C overnight to attain a constant weight. The liquid samples were analyzed by high-performance liquid chromatography (Agilent 1260 Infinity II LC system, Agilent Technologies, Santa Clara, CA, USA) with a refractive index detector and Aminex HPX-87H column (Bio-Rad Laboratories, Hercules, CA, USA). The temperature was 65 °C, and 5 mM of H2SO4 was used as the mobile phase at a flow rate of 0.6 mL/min. The fatty acid composition was estimated according to the fatty acid methyl ester (FAME) measurement by gas chromatography (Agilent 7890A GC System, Agilent Technologies) analysis through direct transesterification [26]. Lipid extraction was conducted using the Folch method with minor modifications. Briefly, 10 mg of dry biomass (after lyophilization) was mixed with 2 mL Folch solvent (2:1 v/v chloroform and methanol); this mixture was then sonicated for 30 min to break the cell walls. For FAME analysis, 1 mL methanol, 300 µL sulfuric acid (H2SO4), and 1 mL chloroform containing lauric acid (C12:0) were used as the internal standard. This mixture was then incubated at 100 °C for 10 min using a heating block for the trans-esterification reaction. A reaction tube supplemented with 1 mL distilled water and the lipid extract fraction in the bottom layer was filtered through a 0.2 µm PTFE syringe filter and analyzed using a capillary HP-INNOWAX column (30 mm × 0.32 mm × 0.5 µm, Agilent Technologies) with a flame ionization detector (FID).
3. Results and Discussion
3.1. Effect of Nitrogen Source on pH of the Medium in Flask Culture
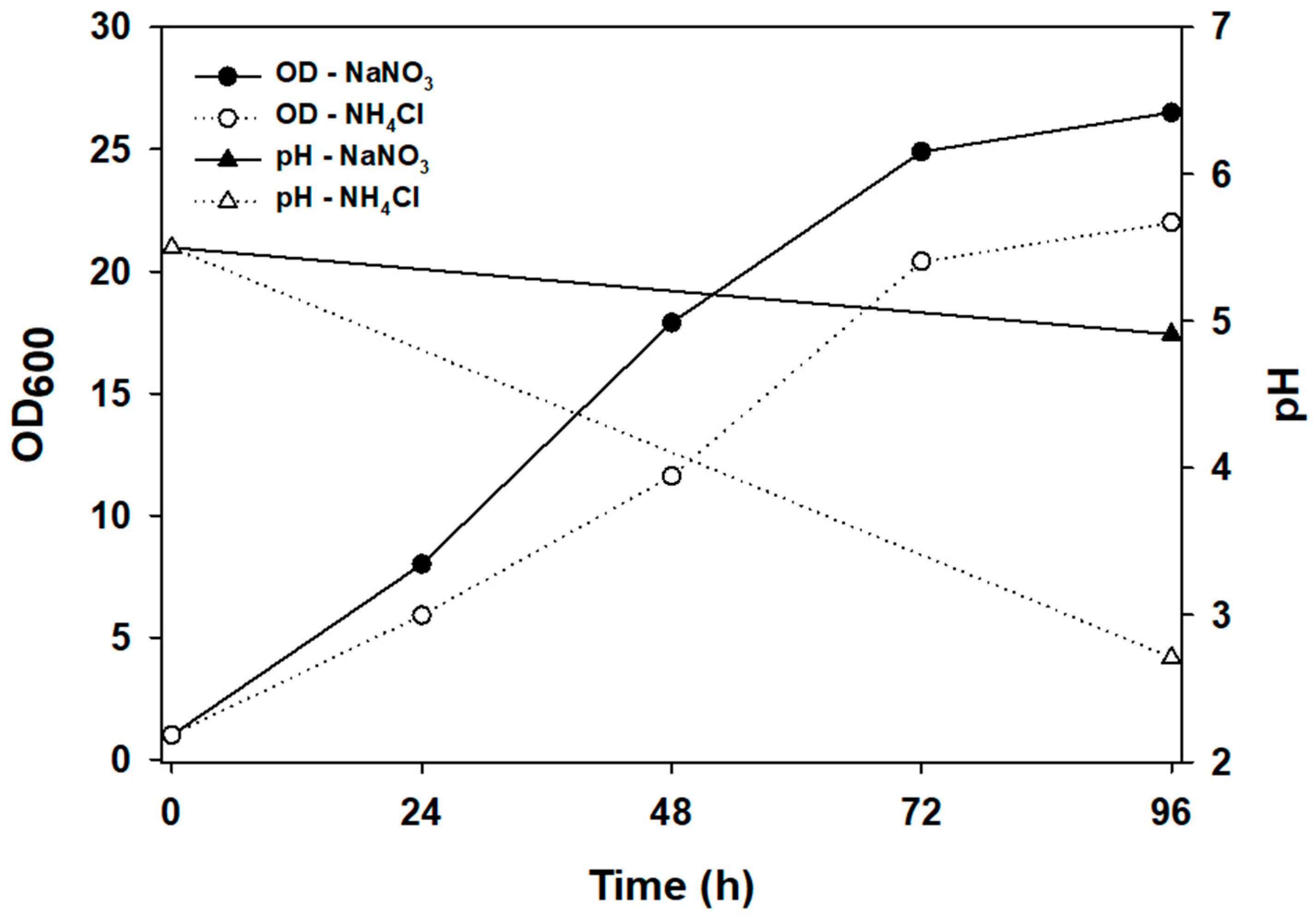
Ideal automatic fed-batch requires the weight of VFA supplied to be equal to the weight of VFA consumed in order to maintain the VFA concentration in broth. If the supply of VFA as an acidic agent and VFA consumption is different, the VFA concentration will not be maintained in the broth. If the VFA supplied as an acid agent exceeds the VFA consumption of oleaginous yeast, the VFA concentration of the broth would increase. Furthermore, if the VFA consumption were higher than that provided by VFA as an acid agent, the VFA concentration would decrease. Hence, the pH of the medium minimizes change from the consumption of nutrients except for VFA. Yeast cultivation used two nitrogen sources (NH4Cl and NaNO3) to find the pH drop effect without a carbon source. The OD values were 26.5 and 22.0 when using NH4Cl and NaNO3 as nitrogen sources, respectively. Hence, cell growth using NaNO3 as a nitrogen source showed 20% higher optical density than that using NH4Cl, and the final pH of the medium with NaNO3 or NH4Cl as the nitrogen source was 4.91 or 2.71, respectively (Figure 2).
The pH of the medium is decreased by the production of hydrogen ions as a result of the consumption of ammonium ions [27]. A similar trend was observed when 20 g/L of glucose was used as the sole carbon source, and Chlorella sp. cultivation from 10 g/L of glucose was used [26,27]. Although it had slightly lower than the initial pH, NaNO3 as a nitrogen source was relatively more acceptable for automatic fed-batch cultivation to maintain substrate concentration. Hence, NaNO3 was used as a nitrogen source in subsequent studies.
3.2. Yeast Cultivation with Manual and Automatic Fed-Batch in 5 L Fermenter
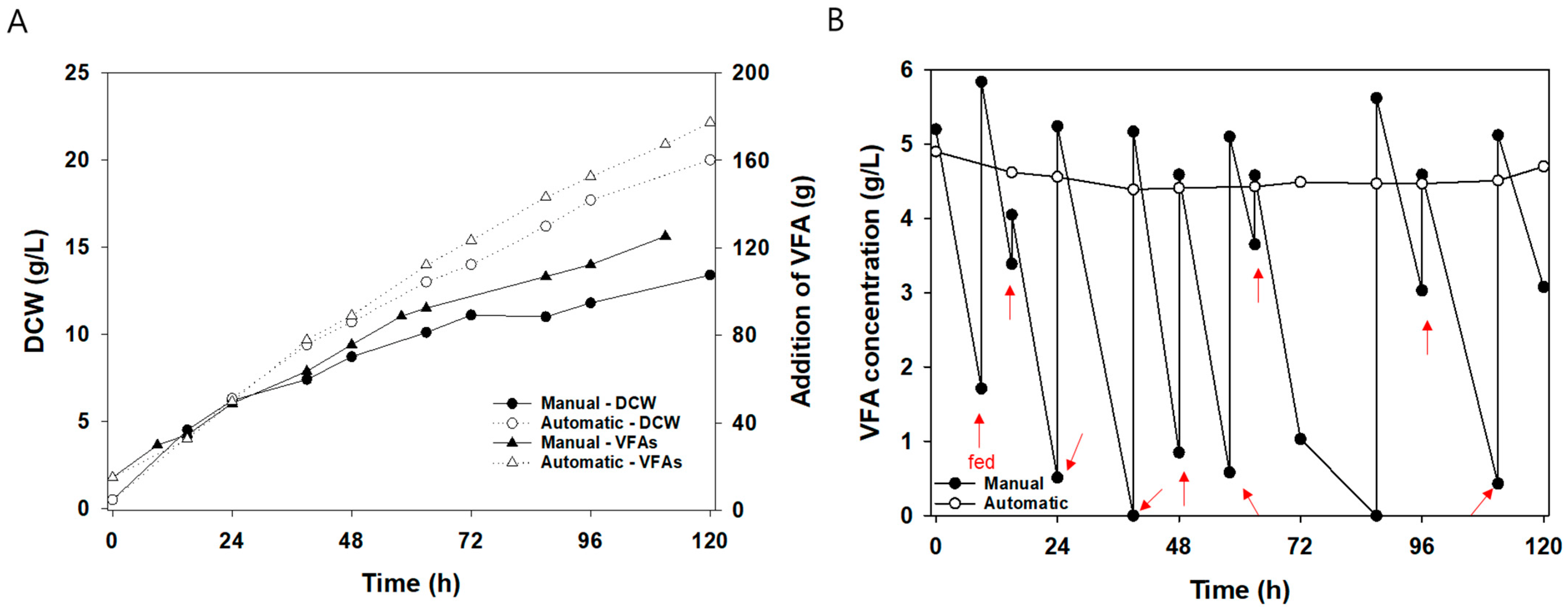
An automatic fed-batch uses the acidic agent for VFA solution when the broth of pH increases by consuming yeast as a carbon source. In order to maintain the broth pH and prevent VFA depletion, acidic agents, including VFA, were added to the fermenter. Two fed-batch (manual and automatic) cultivation modes were used, and the DCW of automatic fed-batch cultivation was 49% higher than that of manual fed-batch at 120 h. Manual and automatic fed-batch had similar biomass yields; however, the reason underlying the DCW difference was that automatic fed-batch had a 41.6% higher addition of VFA than manual fed-batch (Figure 3A). Automatic fed-batch maintained VFA concentration over 4.4 g/L throughout the whole cultivation, and VFA was constantly added to the broth for pH-stat (Figure 3B).
In the manual fed-batch mode, VFA was added to the broth to maintain a 5 g/L concentration, and the pH was adjusted to 7.0 with NaOH. However, manual fed-batch showed depletion of VFA even with two or three feeding times per day. VFA solution was added ten times to reduce VFA concentration across 120 h. This meant that the sampling cycle number had to be increased further. That depletion caused the inhibition of growth and lipid accumulation. The lipid content of automatic fed-batch was higher than that of manual fed-batch, and the lipid produced in automatic fed-batch was 69% higher than that in manual fed-batch during 120 h of cultivation. Lipid yield was 0.21 g/g for the manual fed-batch and 0.25 g/g for the automatic fed-batch. The summarized results are shown in Table 1.
To prevent carbon source depletion in manual fed-batch cultivation, increasing the frequency of manual feeding might enhance lipid production. However, it would lead to extra labor and analysis costs and is an uncomfortable process. Moreover, there was no need to control pH, with HCl as an acidic agent, for pH-stat in automatic fed-batch cultivation. It reduced the process cost of accumulation of Cl− ions for long-term operation. As per our strategy, Shin et al. used formic acid as an acid agent for hydrogen production. In this pH stat, fed-batch cultivation was 1.4-fold higher than that in batch culture [28]. Chi et al. used potassium acetate as the sole carbon source for oleaginous yeast [29]. It is challenging to control the fed-batch manually for different optimized feed concentrations and periods. Thus, the feeding system was changed to automatic using a VFA solution as the acidic agent.
3.3. Effects of VFA Concentration and Initial pH on 5 L Automatic Fed-Batch
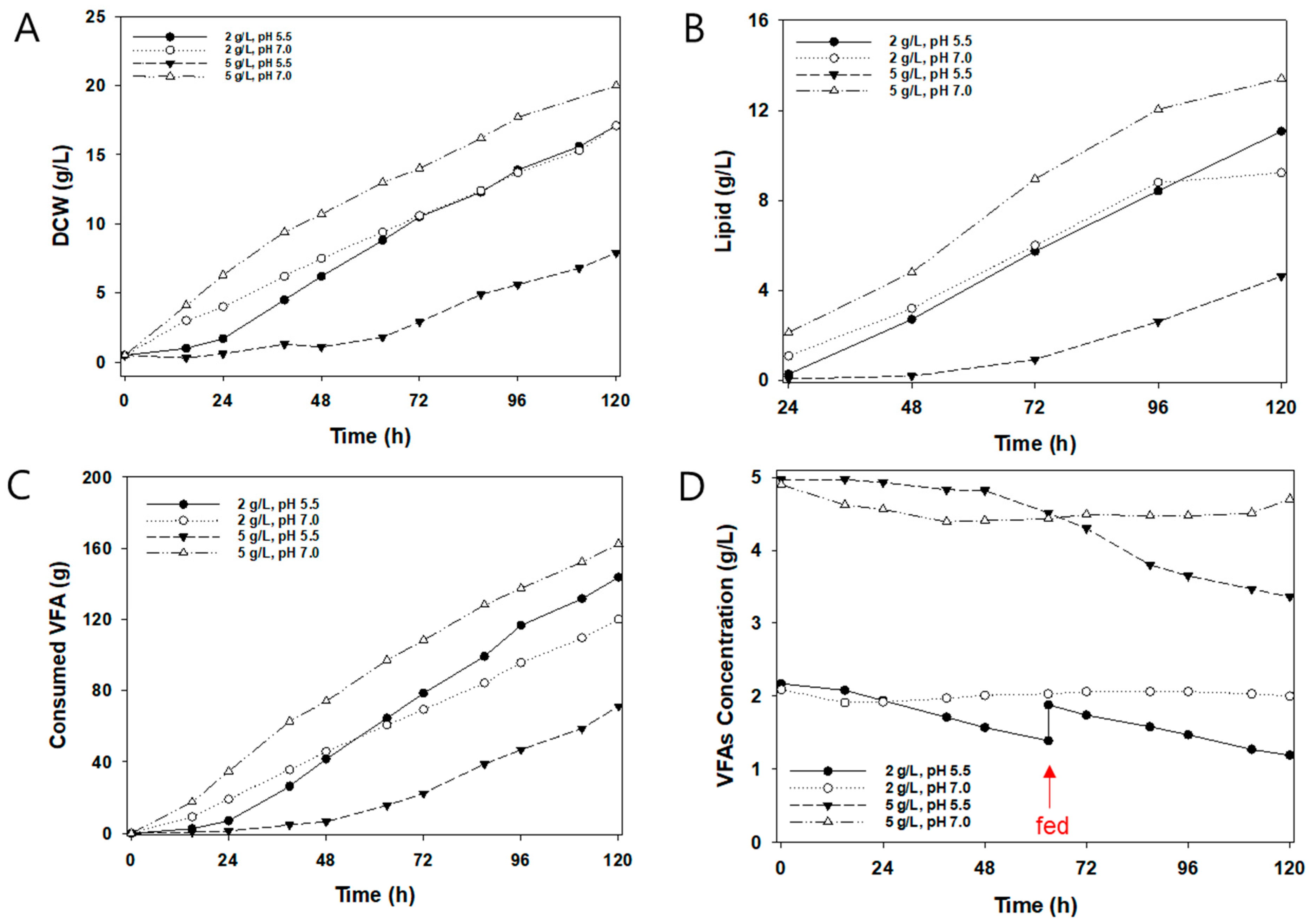
Oleaginous yeast has an optimal culture pH range between 5.0 and 6.0 during lipid production from glucose and glycerol [30]. However, a pH of 7.0 resulted in a higher DCW and lipid than a pH of 5.5 at a VFA concentration of 5 g/L (Figure 4A,B). Moreover, 5 g/L VFA and pH 5.5 represented 24% of the biomass productivity by 5 g/L VFA and pH 7.0 and had a long lag time due to substrate inhibition. The pKa values of AA, PA, and BA are 4.75, 4.88, and 4.82, respectively, and from the Henderson–Hasselbalch equation, acetic acid (undissociated acid) accounts for 15.1% and 0.6% at pH 5.5 and 7.0, respectively. The undissociated form of the acid molecule leads to intracellular acidification, which inhibits cell proliferation [31,32]. Furthermore, C. curvatus did not grow at over 7 g/L AA, 4 g/L PA, or 4 g/L BA at pH 5.5 [26]. At pH 7.0, 5 g/L of VFA resulted in a higher DCW and increased lipids from relatively sufficient carbon and nitrogen sources than those obtained by 2 g/L VFA. The final DCW was almost the same at pH 5.5 and 7.0 from 2 g/L VFA. At pH 5.5 and 2 g/L VFA, the volume of VFA consumed was lower than that consumed at pH 7.0 and 2 g/L VFA until 48 h, after which the opposite trend was observed (Figure 4C,D). At pH 5.5, VFA concentration was difficult to maintain, and required additional manual feeding to avoid VFA depletion. (Figure 4D).
Table 2 shows a comparative summary of the results for each experimental condition of automatic fed-batch cultivation. DCW had a similar value at both pH 5.5 and pH 7.0 at 2 g/L; however, pH 7.0 had a 19% higher biomass yield than pH 5.5. At a low pH, yeast cultivation increased the maintenance energy required to use ATP for energy maintenance [33]. To avoid substrate inhibition, Liu et al. carried out yeast cultivation under pH 7.0 with mixed VFAs and varying concentrations of AA, PA, and BA 0–40 g/L [34]. In this study, the highest lipid productivity obtained was 0.11 g/L/h at pH 7.0 with a 5 g/L of VFA concentration. This was significantly higher than that found for oleaginous yeast cultivation using VFA. Furthermore, Xu et al. obtained 0.011 g/L/h of lipid productivity from microalgae fermentation derived through VFA by C. curvatus [35]. In addition, Fontanille et al. researched fed-batch cultivation with AA, PA, and BA. Lipid productivity was observed to be 0.012, 0.006, and 0.006 g/L/h from AA, PA, and BA, respectively [36]. Moreover, Annamalai et al. produced VFA derived from waste paper with C.curvatus and subsequently obtained a lipid productivity of 0.037 g/L/h [37].
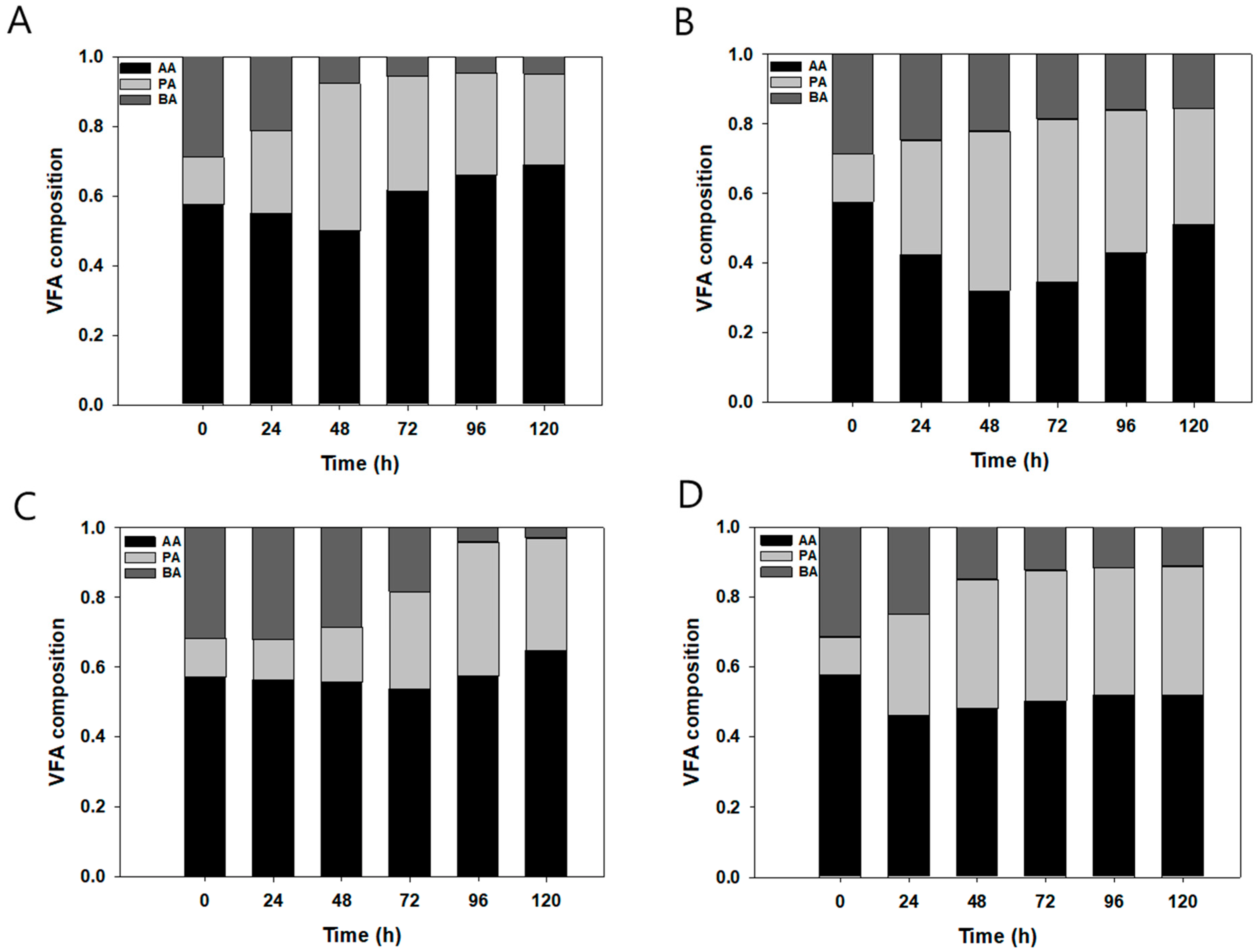
Figure 5 shows the composition of VFA under each experimental condition. The initial VFA and fed-VFA composition ratio was 6:1:3 (AA:PA:BA); however, the final VFA composition ratios were 6.89: 2.61: 0.50 at pH 5.5 and VFA 2 g/L; 5.11: 3.32: 1.57 at pH 5.5 and VFA 7.0 g/L; 6.47: 3.24: 0.30 at pH 7.0 and VFA 5.5 g/L; and 5.19: 3.68: 1.13 at pH 7.0 and VFA 5 g/L. The proportion of BA decreased, and that of PA increased at 120 h. At pH 5.5, the proportion of BA was less than 10% of the total VFA. At pH 5.5 and VFA 5/L, the VFA concentration was not maintained after 48 h and decreased to 3.36 g/L. At that time, butyrate consumption continued to increase after 48 h, with only 3% of butyrate remaining after 120 h. In addition, the propionate ratio was increased compared with the initial ratio under all experimental conditions. Hence, the high butyrate and low propionate content of VFA could enhance both DCW and lipid productivity for its high consumption rate than others. The consumption ratio between 0 and 24 h was 6.44: 3.1: 32.5 of AA: PA: BA at pH 7.0 and VFA 5 g/L. Hence, the PA consumption rate was lowest during the initial period. Furthermore, the consumption ratio between 96 and 120 h was 6.01: 0.91: 3.08 of AA: PA: BA with the same conditions. In automatic fed-batch cultivation, a 5 g/L VFA concentration at pH 7.0 was the optimum culture condition with a 6:1:3 ratio of VFA with 0.25 g/g of lipid and 0.45 g/L/h VFA consumption rate.
Table 3 shows the fatty acid profile and consumption of each VFA on automatic fed-batch cultivation. The major fatty acids were palmitic acid (C16:0), heptadecanoic acid (C17:0), heptadecenoic acid (C17:1), stearic acid (C18:0), oleic acid (C18:1), linoleic acid (C18:2) and, linolenic acid (C18:3). Fatty acid composition profiles were similar under all conditions, and the sum of C18:0 to C18:3 accounted for over 70%. Odd-number fatty acids concentrations were generally not higher than 5% of the total fatty acids in nature (animal or vegetable origin). Considering that propionic acid, odd numbers (C17:0 and C17:1) existed in 7–11% of total fatty acids and was converted to propionyl-CoA before entering the TCA cycle [26,38,39]. The BA consumption ratio was higher at 5 g/L VFA than 2 g/L VFA, whilst palmitic acid (C16:0) increased with the increasing BA consumption ratio. Zheng et al. obtained higher palmitic acid composition when using BA as the sole carbon source rather than PA and AA [38]. Additionally, Liu et al. also reported the lipid production from mixed VFAs and each VFA individually [34]. The lowest biomass and lipid yield was observed with higher PA proportions. Furthermore, higher BA consumption caused an increase in C16:0 fatty acid in microbial lipids. The oleic acid (C18:1) represented the highest proportion of total fatty acids and is a suitable biodiesel due to its high oxidation stability [40]. However, although the instantaneous consumption varied for each organic acid (Figure 5), the actual total amount of organic acid consumption was insignificant since the organic acid supplied remained at a constant ratio of 6:1:3.
4. Conclusions
VFA is a promising feedstock derived from organic waste and can be the sole carbon source for oleaginous yeast. The strain, C. curvatus, requires a set pH control under optimal VFA concentration in order to reduce inhibition due to undissociated acid. A lipid yield of 0.25 g/g alongside lipid productivity of 0.11 g/L/h was observed with an automatic feeding strategy under conditions of 5 g/L VFA and pH 7.0. Automatic fed-batch showed higher biomass productivity and lipid productivity along with better comfort than a manual multi-feeding system and no need for an additional chemical acid agent for pH adjustment. Hence, automatic fed-batch culture is a potential method for utilizing VFA for long-term stable cultivation and obtaining high lipid productivity. As scale-up research progresses further, the VFA-based microbial lipid could be a useful production platform for biofuels and biochemicals. This bioprocess may contribute to carbon neutrality by replacing conventional petroleum-based fuels or chemicals.
Author Contributions
Conceptualization, S.S. and G.W.P.; methodology, S.S. and J.H.G.; software, J.H.G.; formal analysis, S.S.; writing—original draft preparation, S.S. and G.W.P.; writing—review and editing, J.H.G., M.M. and G.W.P.; funding acquisition, G.W.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was part of a project (20210639) titled “the Development of Biohydrogen Plant Operation Optimization System”, funded by the Ministry of Oceans and Fisheries in the Republic of Korea and C1 Gas Refinery Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science and ICT (2021M3D3A1A01079479).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Withanage, S.V.; Dias, G.M.; Habib, K. Review of household food waste quantification methods: Focus on composition analysis. J. Clean. Prod. 2021, 279, 123722. [Google Scholar] [CrossRef]
- Kiran, E.U.; Trzcinski, A.P.; Ng, W.J.; Liu, Y. Bioconversion of food waste to energy: A review. Fuel 2014, 134, 389–399. [Google Scholar] [CrossRef]
- Gao, A.; Tian, Z.; Wang, Z.; Wennersten, R.; Sun, Q. Comparison between the technologies for food waste treatment. Energy Procedia 2017, 105, 3915–3921. [Google Scholar] [CrossRef]
- Caporusso, A.; Capece, A.; De Bari, I. Oleaginous yeasts as cell factories for the sustainable production of microbial lipids by the valorization of agri-food wastes. Fermentation 2021, 7, 50. [Google Scholar] [CrossRef]
- Chang, H.N.; Kim, N.-J.; Kang, J.; Jeong, C.M. Biomass-derived volatile fatty acid platform for fuels and chemicals. Biotechnol. Bioprocess Eng. 2010, 15, 1–10. [Google Scholar] [CrossRef]
- Park, G.W.; Seo, C.; Jung, K.; Chang, H.N.; Kim, W.; Kim, Y.-C. A comprehensive study on volatile fatty acids production from rice straw coupled with microbial community analysis. Bioprocess Biosyst. Eng. 2015, 38, 1157–1166. [Google Scholar] [CrossRef]
- Grubišić, M.; Mihajlovski, K.; Gruičić, A.M.; Beluhan, S.; Šantek, B.; Ivančić Šantek, M. Strategies for Improvement of Lipid Production by Yeast Trichosporon oleaginosus from Lignocellulosic Biomass. J. Fungi 2021, 7, 934. [Google Scholar] [CrossRef]
- Patel, A.; Sarkar, O.; Rova, U.; Christakopoulos, P.; Matsakas, L. Valorization of volatile fatty acids derived from low-cost organic waste for lipogenesis in oleaginous microorganisms-A review. Bioresour. Technol. 2021, 321, 124457. [Google Scholar] [CrossRef]
- Park, G.W.; Kim, N.J.; Chang, H.N. Microbial Lipid Production from Volatile Fatty Acids by Oleaginous Yeast. Emerg. Areas Bioeng. 2018, 1, 203–213. [Google Scholar]
- Park, G.W.; Son, S.; Moon, M.; Sin, S.; Min, K.; Lee, J.-S.; Chang, H.N. Volatile Fatty Acids from Lipid-Extracted Yeast Provide Additional Feedstock for Microbial Lipid Production. Catalysts 2021, 11, 1009. [Google Scholar] [CrossRef]
- Robles-Iglesias, R.; Naveira-Pazos, C.; Fernández-Blanco, C.; Veiga, M.C.; Kennes, C. Factors affecting the optimisation and scale-up of lipid accumulation in oleaginous yeasts for sustainable biofuels production. Renew. Sustain. Energy Rev. 2023, 171, 113043. [Google Scholar] [CrossRef]
- Scaldaferri, C.A.; Pasa, V.M.D. Hydrogen-free process to convert lipids into bio-jet fuel and green diesel over niobium phosphate catalyst in one-step. Chem. Eng. J. 2019, 370, 98–109. [Google Scholar] [CrossRef]
- Fei, Q.; Chang, H.N.; Shang, L.; Choi, J.-D.-R. Exploring low-cost carbon sources for microbial lipids production by fed-batch cultivation of Cryptococcus albidus. Biotechnol. Bioprocess Eng. 2011, 16, 482–487. [Google Scholar] [CrossRef]
- Llamas, M.; Magdalena, J.A.; González-Fernández, C.; Tomás-Pejó, E. Volatile fatty acids as novel building blocks for oil-based chemistry via oleaginous yeast fermentation. Biotechnol. Bioeng. 2020, 117, 238–250. [Google Scholar] [CrossRef]
- van der Klis, F.; Le Nôtre, J.; Blaauw, R.; van Haveren, J.; van Es, D.S. Renewable linear alpha olefins by selective ethenolysis of decarboxylated unsaturated fatty acids. Eur. J. Lipid Sci. Technol. 2012, 114, 911–918. [Google Scholar] [CrossRef]
- Beitel, S.M.; Coelho, L.F.; Contiero, J. Efficient conversion of agroindustrial waste into D(−) lactic acid by Lactobacillus delbrueckii using fed-batch fermentation. BioMed Res. Int. 2020, 2020, 4194052. [Google Scholar] [CrossRef] [Green Version]
- Falk, H.M.; Reichling, P.; Andersen, C.; Benz, R. Online monitoring of concentration and dynamics of volatile fatty acids in anaerobic digestion processes with mid-infrared spectroscopy. Bioprocess Biosyst. Eng. 2015, 38, 237–249. [Google Scholar] [CrossRef]
- Li, X.-Y.; Feng, Y.; Duan, J.-L.; Feng, L.-J.; Wang, Q.; Ma, J.-Y.; Liu, W.-Z.; Yuan, X.-Z. Model-based mid-infrared spectroscopy for on-line monitoring of volatile fatty acids in the anaerobic digester. Environ. Res. 2022, 206, 112607. [Google Scholar] [CrossRef]
- Kim, B.S.; Lee, S.C.; Lee, S.Y.; Chang, Y.K.; Chang, H.N. High cell density fed-batch cultivation of Escherichia coli using exponential feeding combined with pH-stat. Bioprocess Biosyst. Eng. 2004, 26, 147–150. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, X.; Drogui, P.; Tyagi, R.D. The pH-based fed-batch for lipid production from Trichosporon oleaginosus with crude glycerol. Bioresour. Technol. 2018, 259, 237–243. [Google Scholar] [CrossRef]
- Fei, Q.; O’Brien, M.; Nelson, R.; Chen, X.; Lowell, A.; Dowe, N. Enhanced lipid production by Rhodosporidium toruloides using different fed-batch feeding strategies with lignocellulosic hydrolysate as the sole carbon source. Biotechnol. Biofuels 2016, 9, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anschau, A.; Xavier, M.C.; Hernalsteens, S.; Franco, T.T. Effect of feeding strategies on lipid production by Lipomyces starkeyi. Bioresour. Technol. 2014, 157, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Gorte, O.; Kugel, M.; Ochsenreither, K. Optimization of carbon source efficiency for lipid production with the oleaginous yeast Saitozyma podzolica DSM 27192 applying automated continuous feeding. Biotechnol. Biofuels 2020, 13, 181. [Google Scholar] [CrossRef] [PubMed]
- Park, G.W.; Shin, S.; Kim, S.J.; Lee, J.-S.; Moon, M.; Min, K. Rice straw-derived lipid production by HMF/furfural-tolerant oleaginous yeast generated by adaptive laboratory evolution. Bioresour. Technol. 2023, 367, 128220. [Google Scholar] [CrossRef]
- Ryu, B.-G.; Kim, W.; Heo, S.-W.; Kim, D.; Choi, G.-G.; Yang, J.-W. Advanced treatment of residual nitrogen from biologically treated coke effluent by a microalga-mediated process using volatile fatty acids (VFAs) under stepwise mixotrophic conditions. Bioresour. Technol. 2015, 191, 488–495. [Google Scholar] [CrossRef]
- Park, G.W.; Chang, H.N.; Jung, K.; Seo, C.; Kim, Y.-C.; Choi, J.H.; Woo, H.C.; Hwang, I.-J. Production of microbial lipid by Cryptococcus curvatus on rice straw hydrolysates. Process Biochem. 2017, 56, 147–153. [Google Scholar] [CrossRef]
- Kim, H.S.; Park, W.-K.; Lee, B.; Seon, G.; Suh, W.I.; Moon, M.; Chang, Y.K. Optimization of heterotrophic cultivation of Chlorella sp. HS2 using screening, statistical assessment, and validation. Sci. Rep. 2019, 9, 19383. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.-H.; Yoon, J.H.; Lee, S.H.; Park, T.H. Hydrogen production from formic acid in pH-stat fed-batch operation for direct supply to fuel cell. Bioresour. Technol. 2010, 101, S53–S58. [Google Scholar] [CrossRef]
- Chi, Z.; Zheng, Y.; Ma, J.; Chen, S. Oleaginous yeast Cryptococcus curvatus culture with dark fermentation hydrogen production effluent as feedstock for microbial lipid production. Int. J. Hydrogen Energy 2011, 36, 9542–9550. [Google Scholar] [CrossRef]
- Gong, Z.; Shen, H.; Zhou, W.; Wang, Y.; Yang, X.; Zhao, Z.K. Efficient conversion of acetate into lipids by the oleaginous yeast Cryptococcus curvatus. Biotechnol. Biofuels 2015, 8, 189. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhou, W.; He, Q.; Zhao, M.; Gong, Z. Microbial Lipid Production from High Concentration of Volatile Fatty Acids via Trichosporon cutaneum for Biodiesel Preparation. Appl. Biochem. Biotechnol. 2022, 194, 2968–2979. [Google Scholar] [CrossRef]
- Palmqvist, E.; Hahn-Hägerdal, B. Fermentation of lignocellulosic hydrolysates. II: Inhibitors and mechanisms of inhibition. Bioresour. Technol. 2000, 74, 25–33. [Google Scholar] [CrossRef]
- Hakkaart, X.; Liu, Y.; Hulst, M.; El Masoudi, A.; Peuscher, E.; Pronk, J.; van Gulik, W.; Daran-Lapujade, P. Physiological responses of Saccharomyces cerevisiae to industrially relevant conditions: Slow growth, low pH, and high CO2 levels. Biotechnol. Bioeng. 2020, 117, 721–735. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Yuan, M.; Liu, J.-N.; Huang, X.-F. Bioconversion of mixed volatile fatty acids into microbial lipids by Cryptococcus curvatus ATCC 20509. Bioresour. Technol. 2017, 241, 645–651. [Google Scholar] [CrossRef]
- Xu, X.; Kim, J.Y.; Cho, H.U.; Park, H.R.; Park, J.M. Bioconversion of volatile fatty acids from macroalgae fermentation into microbial lipids by oleaginous yeast. Chem. Eng. J. 2015, 264, 735–743. [Google Scholar] [CrossRef]
- Fontanille, P.; Kumar, V.; Christophe, G.; Nouaille, R.; Larroche, C. Bioconversion of volatile fatty acids into lipids by the oleaginous yeast Yarrowia lipolytica. Bioresour. Technol. 2012, 114, 443–449. [Google Scholar] [CrossRef]
- Annamalai, N.; Sivakumar, N.; Fernandez-Castane, A.; Oleskowicz-Popiel, P. Production of microbial lipids utilizing volatile fatty acids derived from wastepaper: A biorefinery approach for biodiesel production. Fuel 2020, 276, 118087. [Google Scholar] [CrossRef]
- Zheng, Y.; Chi, Z.; Ahring, B.K.; Chen, S. Oleaginous yeast Cryptococcus curvatus for biofuel production: Ammonia’s effect. Biomass Bioenergy 2012, 37, 114–121. [Google Scholar] [CrossRef]
- Gao, R.; Li, Z.; Zhou, X.; Bao, W.; Cheng, S.; Zheng, L. Enhanced lipid production by Yarrowia lipolytica cultured with synthetic and waste-derived high-content volatile fatty acids under alkaline conditions. Biotechnol. Biofuels 2020, 13, 3. [Google Scholar] [CrossRef]
- Liu, B.; Zhao, Z. Biodiesel production by direct methanolysis of oleaginous microbial biomass. J. Chem. Technol. Biotechnol. 2007, 82, 775–780. [Google Scholar] [CrossRef]
Figure 1.
Product technology of VFAs and microbial lipid platforms.

Figure 2.
Effects of cell growth and pH change on the different nitrogen sources. Glucose (30 g/L) was used as the sole carbon source at a C/N of 40 in flask culture. Initial pH and rotation speed were 5.5 and 180 rpm, respectively.
Figure 2.
Effects of cell growth and pH change on the different nitrogen sources. Glucose (30 g/L) was used as the sole carbon source at a C/N of 40 in flask culture. Initial pH and rotation speed were 5.5 and 180 rpm, respectively.

Figure 3.
Two fed-batch cultivation modes with C. curvatus: (A) Dry cell weight and addition of VFA amount; (B) VFA concentration in broth. pH-stat for manual feeding used 5 N HCl as an acidic agent, and VFA solution was added ten times for fed-batch. The red arrows indicate when a VFA was added. Automatic feeding used 500 g/L VFA solution to obtain a pH of 7.0. The initial VFA concentration was 5 g/L.
Figure 3.
Two fed-batch cultivation modes with C. curvatus: (A) Dry cell weight and addition of VFA amount; (B) VFA concentration in broth. pH-stat for manual feeding used 5 N HCl as an acidic agent, and VFA solution was added ten times for fed-batch. The red arrows indicate when a VFA was added. Automatic feeding used 500 g/L VFA solution to obtain a pH of 7.0. The initial VFA concentration was 5 g/L.

Figure 4.
Effects of initial VFA concentration and pH on the growth of C. curvatus using the automatic fed-batch mode cultivation: (A) dry cell weight, (B) lipid, (C) consumed VFA, and (D) VFA concentration in broth. The red arrow indicates when a VFA was added.
Figure 4.
Effects of initial VFA concentration and pH on the growth of C. curvatus using the automatic fed-batch mode cultivation: (A) dry cell weight, (B) lipid, (C) consumed VFA, and (D) VFA concentration in broth. The red arrow indicates when a VFA was added.

Figure 5.
VFA composition under each condition (initial VFA composition was 60% AA, 10% PA, and 30% BA): (A) 2 g/L and pH 5.5, (B) 2 g/L and pH 7.0, (C) 5 g/L and pH 5.5, and (D) 5 g/L and pH 7.0. VFA solutions were fed manually after 63 h in order to maintain the concentration at 2 g/L and pH 5.5.
Figure 5.
VFA composition under each condition (initial VFA composition was 60% AA, 10% PA, and 30% BA): (A) 2 g/L and pH 5.5, (B) 2 g/L and pH 7.0, (C) 5 g/L and pH 5.5, and (D) 5 g/L and pH 7.0. VFA solutions were fed manually after 63 h in order to maintain the concentration at 2 g/L and pH 5.5.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Comparison of manual and automatic fed-batch cultivation.
| Category | Manual Fed-Batch | Automatic Fed-Batch |
|---|---|---|
| DCW (g/L) | 13.40 | 19.95 |
| Biomass yield (g/g) | 0.35 | 0.37 |
| Lipid content (%) | 59 | 67 |
| Lipid (g/L) * | 7.91 | 13.37 |
| VFA consumption rate (g/L/h) | 0.32 | 0.45 |
* Lipid means amount of lipid in yeast cells per 1 L broth. Hence, lipid = DCW × lipid content.
Table 2.
Comparison of different automatic fed-batch cultivation conditions.
| Category | Cultivation Condition (VFA Concentration, pH) | |||
|---|---|---|---|---|
| 2 g/L, pH 5.5 | 2 g/L, pH 7.0 | 5 g/L, pH 5.5 | 5 g/L, pH 7.0 | |
| Biomass yield (g/g) | 0.35 | 0.43 | 0.31 | 0.37 |
| Lipid yield (g/g) | 0.23 | 0.23 | 0.18 | 0.25 |
| VFA consumption rate (g/L/h) | 0.41 | 0.33 | 0.21 | 0.45 |
| Biomass productivity (g/L/h) | 0.14 | 0.14 | 0.04 | 0.17 |
| Lipid productivity (g/L/h) | 0.09 | 0.08 | 0.07 | 0.11 |
Table 3.
Fatty acid profile and consumed VFA of C. curvatus from automatic fed-batch cultivation.
| Category | Cultivation Condition (VFA Concentration, pH) | ||||
|---|---|---|---|---|---|
| 2 g/L, pH 5.5 | 2 g/L, pH 7.0 | 5 g/L, pH 5.5 | 5 g/L, pH 7.0 | ||
| Fatty acid composition a (%) | C16:0 | 11.08 | 11.56 | 15.34 | 14.96 |
| C17:0 | 6.32 | 8.68 | 6.65 | 7.09 | |
| C17:1 | 1.23 | 2.19 | 1.62 | 1.50 | |
| C18:0 | 14.18 | 11.71 | 11.21 | 14.80 | |
| C18:1 | 53.64 | 49.88 | 50.17 | 47.67 | |
| C18:2 | 7.81 | 9.72 | 9.39 | 8.40 | |
| C18:3 | 2.62 | 3.29 | 2.36 | 2.84 | |
| Consumed VFA (g) | AA | 88.37 | 72.58 | 44.69 | 98.63 |
| PA | 14.47 | 10.87 | 5.50 | 12.63 | |
| BA | 45.24 | 36.88 | 25.79 | 51.77 | |
| Consumed VFA ratio | 59.7:9.8:30.6 | 60.3:9.0:30.6 | 58.8:7.2:34 | 60.5:7.7:31.8 | |
a Cell was harvested at 120 h, and each fatty acid composition was analyzed.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shin, S.; Go, J.H.; Moon, M.; Park, G.W. Automatic Fed-Batch Cultivation Enhances Microbial Lipid Production from Volatile Fatty Acids. Energies 2023, 16, 1996. https://doi.org/10.3390/en16041996
AMA Style
Shin S, Go JH, Moon M, Park GW. Automatic Fed-Batch Cultivation Enhances Microbial Lipid Production from Volatile Fatty Acids. Energies. 2023; 16(4):1996. https://doi.org/10.3390/en16041996
Chicago/Turabian StyleShin, Subin, Jae Hee Go, Myounghoon Moon, and Gwon Woo Park. 2023. "Automatic Fed-Batch Cultivation Enhances Microbial Lipid Production from Volatile Fatty Acids" Energies 16, no. 4: 1996. https://doi.org/10.3390/en16041996
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.