
Photocurable Hydrogel Substrate—Better Potential Substitute on Bone-Marrow-Derived Dendritic Cells Culturing

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fabrication of GelMA Hydrogels
2.2. Fabrication of Hydrogel Solutions
2.3. Mechanical Testing
2.4. Routine Culture of Mouse Bone-Marrow-Derived Dendritic Cells (BMDCs) on GelMA-30 and Cell Morphology Observation
2.5. Isolation and Culture of T Cells from Mouse Spleen
2.6. MACS Magnetic Beads Separate BMDCs and T Cells
2.7. DCs Activation Test In Vitro
2.8. The Secretion of Cytokines Was Detected by ELISA
2.9. The Phenotype of BMDCs Was Detected by FACS
2.10. T Cell Proliferation Test
2.11. Real-Time PCR
| IRF4 | Forward Primer | CTTTGAGGAATTGGTCGAGAGG |
| Reverse Primer | GAGAGCCATAAGGTGCTGTCA | |
| IRF8 | Forward Primer | AGACCATGTTCCGTATCCCCT |
| Reverse Primer | CACAGCGTAACCTCGTCTTCC | |
| Batf3 | Forward Primer | CAGAGCCCCAAGGACGATG |
| Reverse Primer | GCACAAAGTTCATAGGACACAGC | |
| β-Actin | Forward Primer | AGATCAAGATCATTGCTCCTCCT |
| Reverse Primer | ACGCAGCTCAGTAACAGTCC |
2.12. Phagocytosis Assay and Flow Cytometry
2.13. Statistical Analysis
3. Result
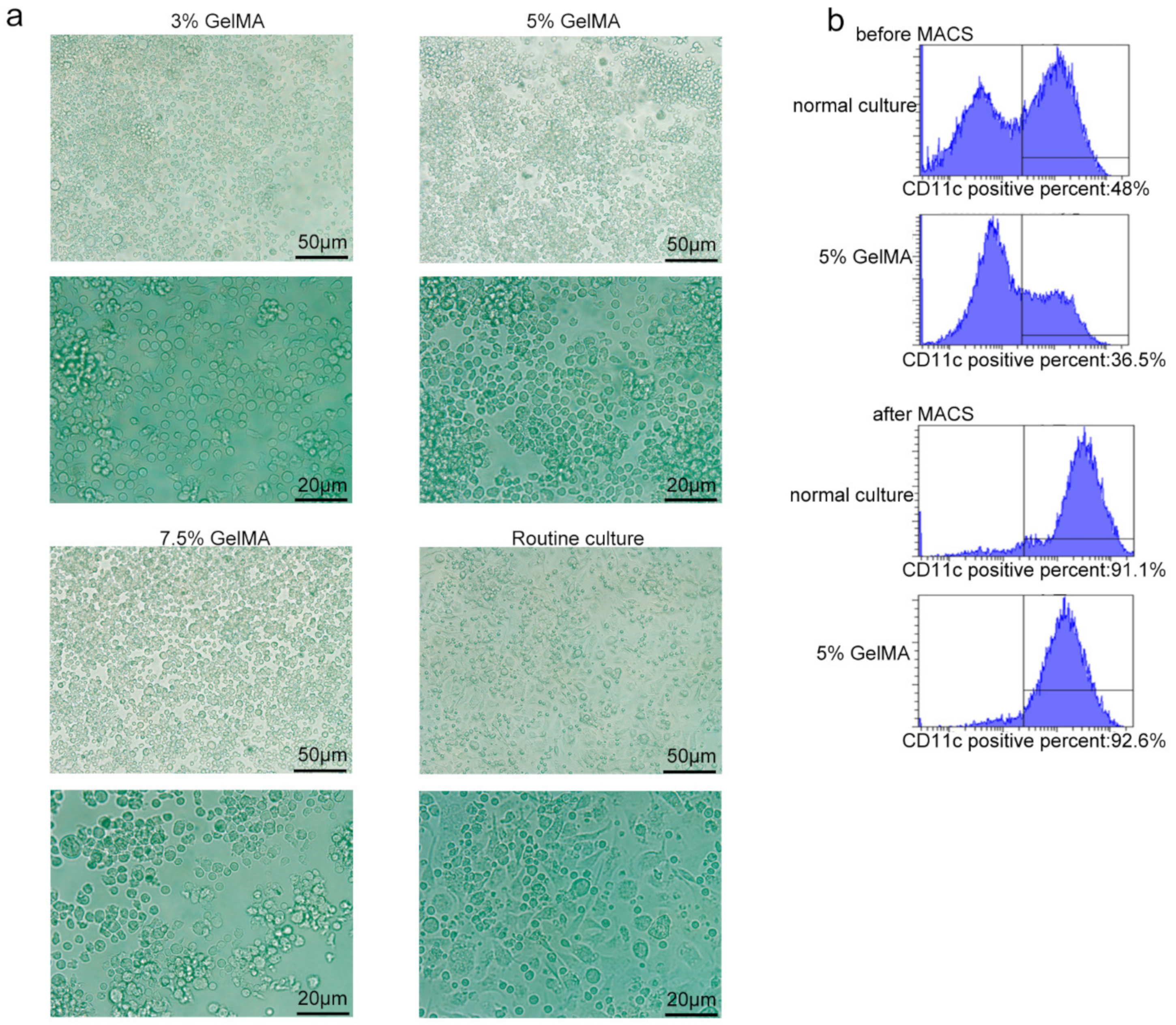
3.1. BMDCs on GelMA-30 Substrates

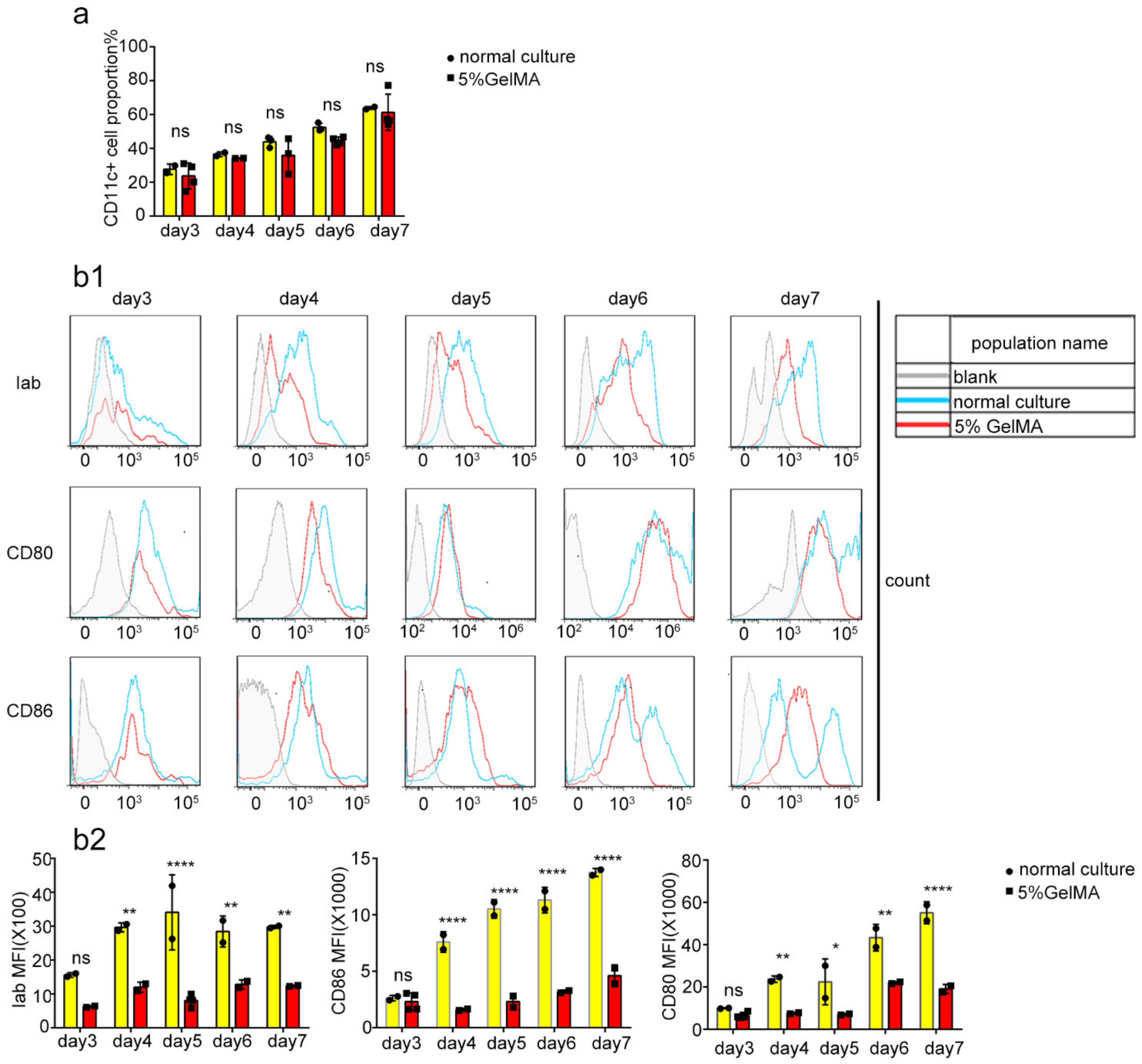
3.2. BMDCs Were Induced and Cultured Successfully on 5% GelMA-30
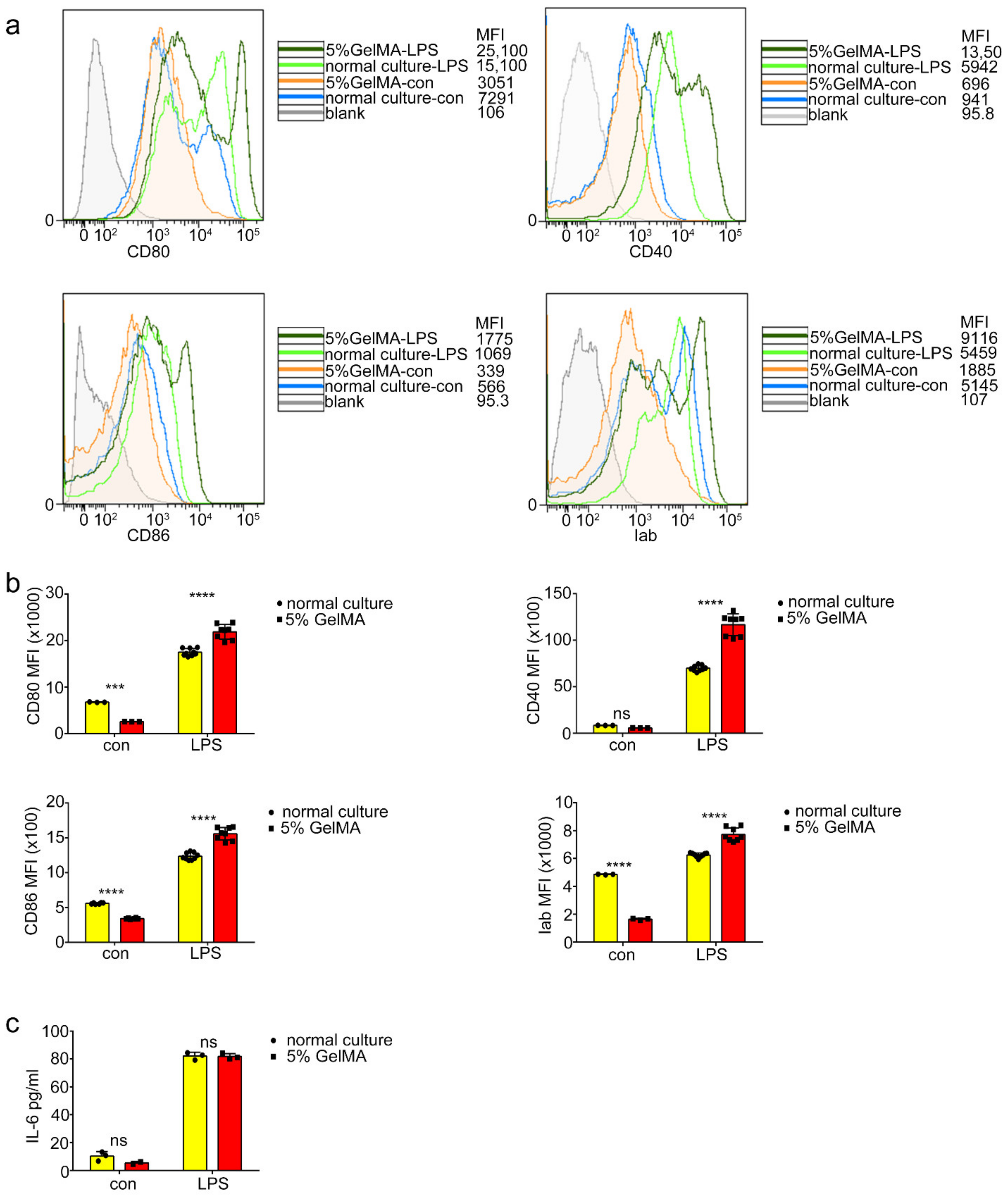
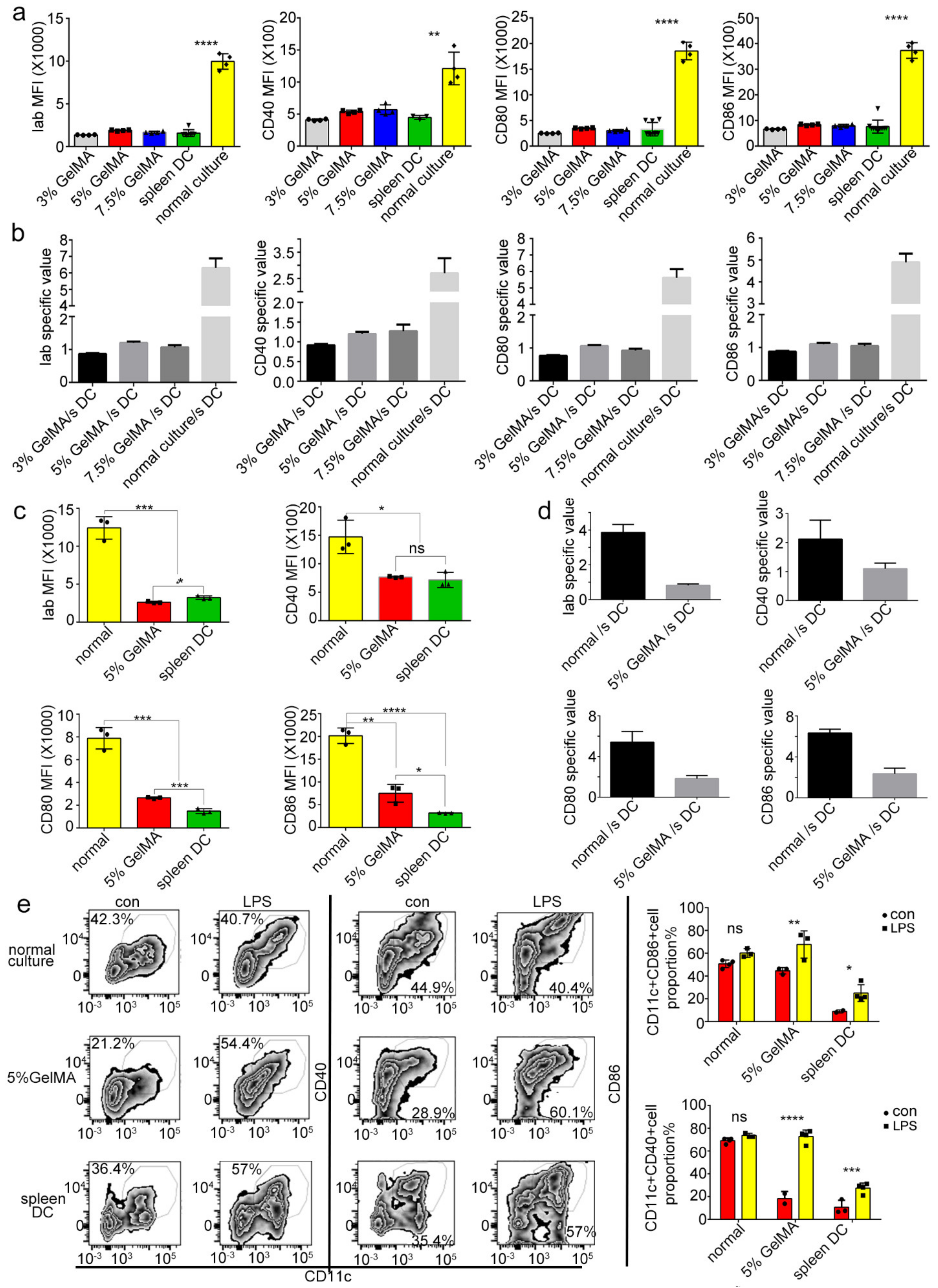
3.3. BMDCs on 5% GelMA-30 with Low Expression Levels of Costimulatory Molecules Were Activated by LPS
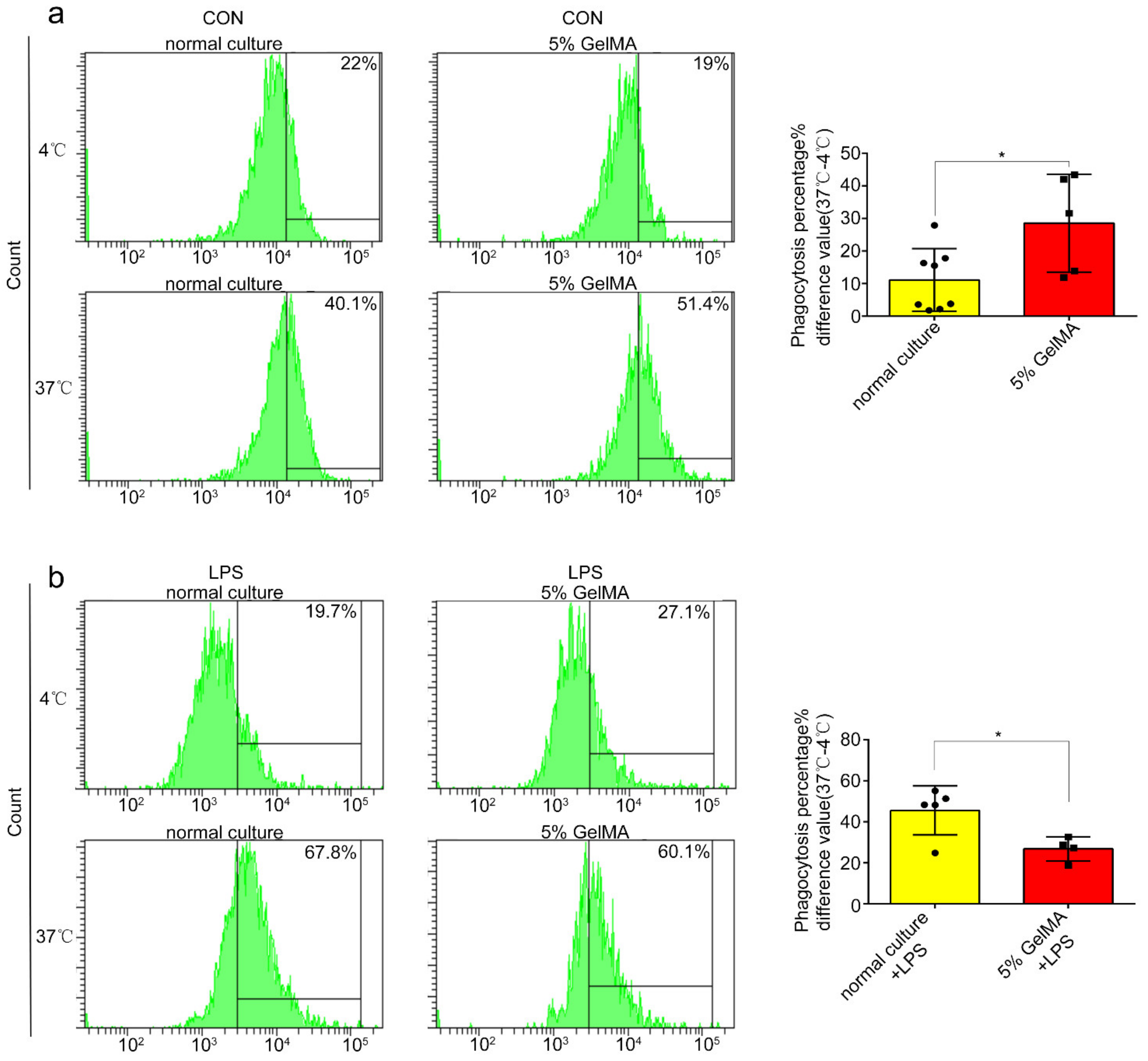
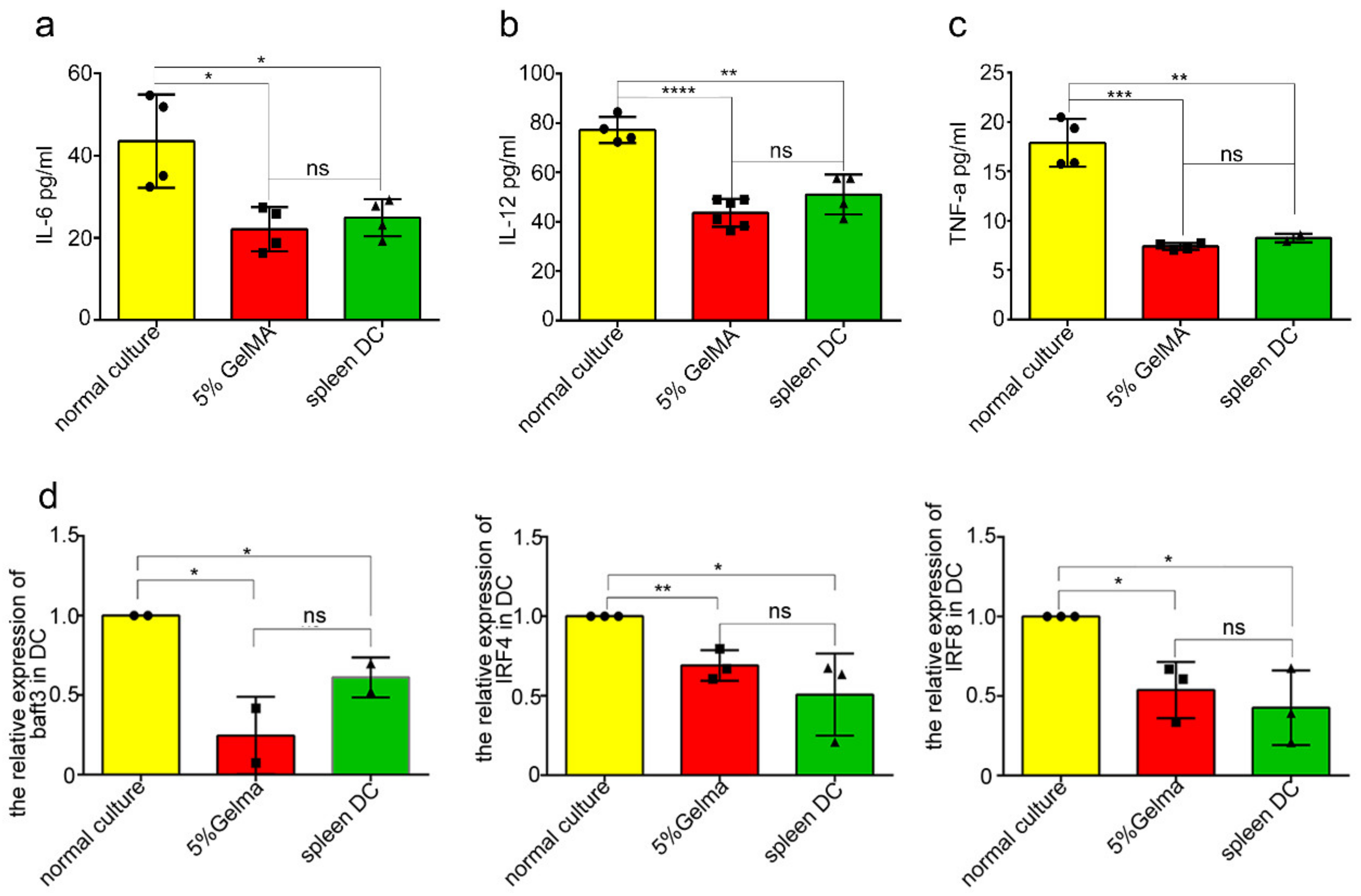
3.4. BMDCs on GelMA-30 Approached Spleen DCs
3.5. BMDCs on 5% GelMA-30 Were Closer to Spleen DCs in Terms of the Developmental State
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CCHS | BMDCs culturing with hydrogel substrate |
| BMDCs | Bone marrow-derived Dendritic cells |
| DCs | Dendritic cells |
| cDCs | Conventional Dendritic cells |
| LPS | Lipopolysaccharide |
| PBS | Phosphate buffer saline |
| GelMA-30 | Gelatin Methacrylate-30 |
| LAP | Lithium phenyl-2,4,6-trimethylbenzoylphosphinate |
| GM-CSF | Granulocyte/macrophage colony stimulating factor (GM-CSF) |
| TNF-a | Tumor necrosis factor-α |
| IL | Interleukin |
| IRF | Interferon regulatory factor |
| Batf | Basic leucine zipper ATF-like transcription factor |
| ID2 | DNA-binding2 |
| Nfil3 | Nuclear factor, interleukin 3 regulated |
| Bcl6 | B cell lymphoma 6 |
References
- Choo, E.H.; Lee, J.H.; Park, E.H.; Park, H.E.; Jung, N.C.; Kim, T.H.; Koh, Y.S.; Kim, E.; Seung, K.B.; Park, C.; et al. Infarcted Myocardium-Primed Dendritic Cells Improve Remodeling and Cardiac Function After Myocardial Infarction by Modulating the Regulatory T Cell and Macrophage Polarization. Circulation 2017, 135, 1444–1457. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.W.; Yang, W.; Gao, L.; Kang, J.R.; Qin, J.J.; Liu, Y.P.; Lu, J.Y. Adoptive transfer of bone marrow-derived dendritic cells decreases inhibitory and regulatory T-cell differentiation and improves survival in murine polymicrobial sepsis. Immunology 2015, 145, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Tran, S.D.; Sumita, Y.; Khalili, S. Bone marrow-derived cells: A potential approach for the treatment of xerostomia. Int. J. Biochem. Cell Biol. 2011, 43, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Ge, C.; Li, R.; Song, H.; Geng, T.; Yang, J.; Tan, Q.; Song, L.; Wang, Y.; Xue, Y.; Li, Z.; et al. Phase I clinical trial of a novel autologous modified-DC vaccine in patients with resected NSCLC. BMC Cancer 2017, 17, 884. [Google Scholar] [CrossRef] [PubMed]
- Chia, W.K.; Wang, W.W.; Teo, M.; Tai, W.M.; Lim, W.T.; Tan, E.H.; Leong, S.S.; Sun, L.; Chen, J.J.; Gottschalk, S.; et al. A phase II study evaluating the safety and efficacy of an adenovirus-DeltaLMP1-LMP2 transduced dendritic cell vaccine in patients with advanced metastatic nasopharyngeal carcinoma. Ann. Oncol. 2012, 23, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Mao, Q.; Li, L.; Zhang, C.; Sun, Y.; Liu, S.; Cui, S. Clinical effects of immunotherapy of DC-CIK combined with chemotherapy in treating patients with metastatic breast cancer. Pak. J. Pharm. Sci. 2015, 28, 1055–1058. [Google Scholar]
- Lee, J.H.; Lee, Y.; Lee, M.; Heo, M.K.; Song, J.S.; Kim, K.H.; Lee, H.; Yi, N.J.; Lee, K.W.; Suh, K.S.; et al. A phase I/IIa study of adjuvant immunotherapy with tumour antigen-pulsed dendritic cells in patients with hepatocellular carcinoma. Br. J. Cancer 2015, 113, 1666–1676. [Google Scholar] [CrossRef] [Green Version]
- Xi, H.B.; Wang, G.X.; Fu, B.; Liu, W.P.; Li, Y. Survivin and PSMA Loaded Dendritic Cell Vaccine for the Treatment of Prostate Cancer. Biol. Pharm. Bull. 2015, 38, 827–835. [Google Scholar] [CrossRef] [Green Version]
- Sabado, R.L.; Bhardwaj, N. Directing dendritic cell immunotherapy towards successful cancer treatment. Immunotherapy 2010, 2, 37–56. [Google Scholar] [CrossRef] [Green Version]
- Bol, K.F.; Schreibelt, G.; Rabold, K.; Wculek, S.K.; Schwarze, J.K.; Dzionek, A.; Teijeira, A.; Kandalaft, L.E.; Romero, P.; Coukos, G.; et al. The clinical application of cancer immunotherapy based on naturally circulating dendritic cells. J. Immunother. Cancer 2019, 7, 109. [Google Scholar] [CrossRef]
- Chung, J.T.; Lau, C.M.L.; Chau, Y. The effect of polysaccharide-based hydrogels on the response of antigen-presenting cell lines to immunomodulators. Biomater. Sci. 2021, 9, 6542–6554. [Google Scholar] [CrossRef]
- Hume, P.S.; He, J.; Haskins, K.; Anseth, K.S. Strategies to reduce dendritic cell activation through functional biomaterial design. Biomaterials 2012, 33, 3615–3625. [Google Scholar] [CrossRef] [Green Version]
- Molzer, C.; Shankar, S.P.; Griffith, M.; Islam, M.M.; Forrester, J.V.; Kuffova, L. Activation of dendritic cells by crosslinked collagen hydrogels (artificial corneas) varies with their composition. J. Tissue Eng. Regen. Med. 2019, 13, 1528–1543. [Google Scholar] [CrossRef]
- Park, J.; Babensee, J.E. Differential functional effects of biomaterials on dendritic cell maturation. Acta Biomater. 2012, 8, 3606–3617. [Google Scholar] [CrossRef] [Green Version]
- Nikkhah, M.; Eshak, N.; Zorlutuna, P.; Annabi, N.; Castello, M.; Kim, K.; Dolatshahi-Pirouz, A.; Edalat, F.; Bae, H.; Yang, Y.; et al. Directed endothelial cell morphogenesis in micropatterned gelatin methacrylate hydrogels. Biomaterials 2012, 33, 9009–9018. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.; Niu, X.; Shao, L.; Zhou, L.; Lin, Z.; Sun, A.; Fu, J.; Chen, Z.; Hu, J.; Liu, Y.; et al. 3D printing of complex GelMA-based scaffolds with nanoclay. Biofabrication 2019, 11, 035006. [Google Scholar] [CrossRef]
- Shin, S.R.; Jung, S.M.; Zalabany, M.; Kim, K.; Zorlutuna, P.; Kim, S.B.; Nikkhah, M.; Khabiry, M.; Azize, M.; Kong, J.; et al. Carbon-nanotube-embedded hydrogel sheets for engineering cardiac constructs and bioactuators. ACS Nano. 2013, 7, 2369–2380. [Google Scholar] [CrossRef] [Green Version]
- Saini, H.; Navaei, A.; Van Putten, A.; Nikkhah, M. 3D cardiac microtissues encapsulated with the co-culture of cardiomyocytes and cardiac fibroblasts. Adv. Healthc. Mater. 2015, 4, 1961–1971. [Google Scholar] [CrossRef]
- Ovsianikov, A.; Deiwick, A.; Van Vlierberghe, S.; Dubruel, P.; Moller, L.; Drager, G.; Chichkov, B. Laser fabrication of three-dimensional CAD scaffolds from photosensitive gelatin for applications in tissue engineering. Biomacromolecules 2011, 12, 851–858. [Google Scholar] [CrossRef]
- Dolatshahi-Pirouz, A.; Nikkhah, M.; Gaharwar, A.K.; Hashmi, B.; Guermani, E.; Aliabadi, H.; Camci-Unal, G.; Ferrante, T.; Foss, M.; Ingber, D.E.; et al. A combinatorial cell-laden gel microarray for inducing osteogenic differentiation of human mesenchymal stem cells. Sci. Rep. 2014, 4, 3896. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.C.; Lin, R.Z.; Qi, H.; Yang, Y.; Bae, H.; Melero-Martin, J.M.; Khademhosseini, A. Functional Human Vascular Network Generated in Photocrosslinkable Gelatin Methacrylate Hydrogels. Adv. Funct. Mater. 2012, 22, 2027–2039. [Google Scholar] [CrossRef] [Green Version]
- Bertassoni, L.E.; Cecconi, M.; Manoharan, V.; Nikkhah, M.; Hjortnaes, J.; Cristino, A.L.; Barabaschi, G.; Demarchi, D.; Dokmeci, M.R.; Yang, Y.; et al. Hydrogel bioprinted microchannel networks for vascularization of tissue engineering constructs. Lab Chip 2014, 14, 2202–2211. [Google Scholar] [CrossRef] [Green Version]
- Peela, N.; Sam, F.S.; Christenson, W.; Truong, D.; Watson, A.W.; Mouneimne, G.; Ros, R.; Nikkhah, M. A three dimensional micropatterned tumor model for breast cancer cell migration studies. Biomaterials 2016, 81, 72–83. [Google Scholar] [CrossRef]
- Donaldson, A.R.; Tanase, C.E.; Awuah, D.; Vasanthi Bathrinarayanan, P.; Hall, L.; Nikkhah, M.; Khademhosseini, A.; Rose, F.; Alexander, C.; Ghaemmaghami, A.M. Photocrosslinkable Gelatin Hydrogels Modulate the Production of the Major Pro-inflammatory Cytokine, TNF-alpha, by Human Mononuclear Cells. Front. Bioeng. Biotechnol. 2018, 6, 116. [Google Scholar] [CrossRef]
- Chakraborty, M.; Chu, K.; Shrestha, A.; Revelo, X.S.; Zhang, X.; Gold, M.J.; Khan, S.; Lee, M.; Huang, C.; Akbari, M.; et al. Mechanical Stiffness Controls Dendritic Cell Metabolism and Function. Cell Rep 2021, 34, 108609. [Google Scholar] [CrossRef]
- Jimenez, R.V.; Wright, T.T.; Jones, N.R.; Wu, J.; Gibson, A.W.; Szalai, A.J. C-Reactive Protein Impairs Dendritic Cell Development, Maturation, and Function: Implications for Peripheral Tolerance. Front. Immunol. 2018, 9, 372. [Google Scholar] [CrossRef]
- Zheng, H.; Liu, L.; Zhang, H.; Kan, F.; Wang, S.; Li, Y.; Tian, H.; Meng, S. Dendritic cells pulsed with placental gp96 promote tumor-reactive immune responses. PLoS ONE 2019, 14, e0211490. [Google Scholar] [CrossRef]
- Tan, J.K.; O’Neill, H.C. Maturation requirements for dendritic cells in T cell stimulation leading to tolerance versus immunity. J. Leukoc. Biol. 2005, 78, 319–324. [Google Scholar] [CrossRef]
- Garcia-Nieto, S.; Johal, R.K.; Shakesheff, K.M.; Emara, M.; Royer, P.J.; Chau, D.Y.; Shakib, F.; Ghaemmaghami, A.M. Laminin and fibronectin treatment leads to generation of dendritic cells with superior endocytic capacity. PLoS ONE 2010, 5, e10123. [Google Scholar] [CrossRef] [Green Version]
- Viebig, N.K.; Andrews, K.T.; Kooyk, Y.; Lanzer, M.; Knolle, P.A. Evaluation of the role of the endocytic receptor L-SIGN for cytoadhesion of Plasmodium falciparum-infected erythrocytes. Parasitol. Res. 2005, 96, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Geijtenbeek, T.B.; Groot, P.C.; Nolte, M.A.; van Vliet, S.J.; Gangaram-Panday, S.T.; van Duijnhoven, G.C.; Kraal, G.; van Oosterhout, A.J.; van Kooyk, Y. Marginal zone macrophages express a murine homologue of DC-SIGN that captures blood-borne antigens in vivo. Blood 2002, 100, 2908–2916. [Google Scholar] [CrossRef] [PubMed]
- Takahara, K.; Yashima, Y.; Omatsu, Y.; Yoshida, H.; Kimura, Y.; Kang, Y.S.; Steinman, R.M.; Park, C.G.; Inaba, K. Functional comparison of the mouse DC-SIGN, SIGNR1, SIGNR3 and Langerin, C-type lectins. Int. Immunol. 2004, 16, 819–829. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, S.F.; Ramsdell, F.; Alderson, M.R. The activation antigen CD69. Stem Cells 1994, 12, 456–465. [Google Scholar] [CrossRef]
- Bensinger, S.J.; Walsh, P.T.; Zhang, J.; Carroll, M.; Parsons, R.; Rathmell, J.C.; Thompson, C.B.; Burchill, M.A.; Farrar, M.A.; Turka, L.A. Distinct IL-2 receptor signaling pattern in CD4+ CD25+ regulatory T cells. J. Immunol. 2004, 172, 5287–5296. [Google Scholar] [CrossRef] [Green Version]
- Schiavoni, G.; Mattei, F.; Sestili, P.; Borghi, P.; Venditti, M.; Morse, H.C., 3rd; Belardelli, F.; Gabriele, L. ICSBP is essential for the development of mouse type I interferon-producing cells and for the generation and activation of CD8α+ dendritic cells. J. Exp. Med. 2002, 196, 1415–1425. [Google Scholar] [CrossRef] [Green Version]
- Aliberti, J.; Schulz, O.; Pennington, D.J.; Tsujimura, H.; Reis e Sousa, C.; Ozato, K.; Sher, A. Essential role for ICSBP in the in vivo development of murine CD8α+ dendritic cells. Blood 2003, 101, 305–310. [Google Scholar] [CrossRef] [Green Version]
- Hildner, K.; Edelson, B.T.; Purtha, W.E.; Diamond, M.; Matsushita, H.; Kohyama, M.; Calderon, B.; Schraml, B.U.; Unanue, E.R.; Diamond, M.S.; et al. Batf3 deficiency reveals a critical role for CD8α+ dendritic cells in cytotoxic T cell immunity. Science 2008, 322, 1097–1100. [Google Scholar] [CrossRef] [Green Version]
- Edelson, B.T.; Kc, W.; Juang, R.; Kohyama, M.; Benoit, L.A.; Klekotka, P.A.; Moon, C.; Albring, J.C.; Ise, W.; Michael, D.G.; et al. Peripheral CD103+ dendritic cells form a unified subset developmentally related to CD8α+ conventional dendritic cells. J. Exp. Med. 2010, 207, 823–836. [Google Scholar] [CrossRef] [Green Version]
- Hacker, C.; Kirsch, R.D.; Ju, X.S.; Hieronymus, T.; Gust, T.C.; Kuhl, C.; Jorgas, T.; Kurz, S.M.; Rose-John, S.; Yokota, Y.; et al. Transcriptional profiling identifies Id2 function in dendritic cell development. Nat. Immunol. 2003, 4, 380–386. [Google Scholar] [CrossRef]
- Kobayashi, T.; Walsh, P.T.; Walsh, M.C.; Speirs, K.M.; Chiffoleau, E.; King, C.G.; Hancock, W.W.; Caamano, J.H.; Hunter, C.A.; Scott, P.; et al. TRAF6 is a critical factor for dendritic cell maturation and development. Immunity 2003, 19, 353–363. [Google Scholar] [CrossRef] [Green Version]
- Kashiwada, M.; Pham, N.L.; Pewe, L.L.; Harty, J.T.; Rothman, P.B. NFIL3/E4BP4 is a key transcription factor for CD8α+ dendritic cell development. Blood 2011, 117, 6193–6197. [Google Scholar] [CrossRef]
- Ohtsuka, H.; Sakamoto, A.; Pan, J.; Inage, S.; Horigome, S.; Ichii, H.; Arima, M.; Hatano, M.; Okada, S.; Tokuhisa, T. Bcl6 is required for the development of mouse CD4+ and CD8α+ dendritic cells. J. Immunol. 2011, 186, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Kc, W.; Satpathy, A.T.; Rapaport, A.S.; Briseno, C.G.; Wu, X.; Albring, J.C.; Russler-Germain, E.V.; Kretzer, N.M.; Durai, V.; Persaud, S.P.; et al. L-Myc expression by dendritic cells is required for optimal T-cell priming. Nature 2014, 507, 243–247. [Google Scholar] [CrossRef]
- Dwyer, K.D.; Coulombe, K.L.K. Cardiac mechanostructure: Using mechanics and anisotropy as inspiration for developing epicardial therapies in treating myocardial infarction. Bioact. Mater. 2021, 6, 2198–2220. [Google Scholar] [CrossRef]
- Sridharan, R.; Cavanagh, B.; Cameron, A.R.; Kelly, D.J.; O’Brien, F.J. Material stiffness influences the polarization state, function and migration mode of macrophages. Acta Biomater. 2019, 89, 47–59. [Google Scholar] [CrossRef]
- Turnbull, E.; MacPherson, G. Immunobiology of dendritic cells in the rat. Immunol. Rev. 2001, 184, 58–68. [Google Scholar] [CrossRef]
- McIlroy, D.; Gregoire, M. Optimizing dendritic cell-based anticancer immunotherapy: Maturation state does have clinical impact. Cancer Immunol. Immunother. 2003, 52, 583–591. [Google Scholar] [CrossRef]
- Banchereau, J.; Briere, F.; Caux, C.; Davoust, J.; Lebecque, S.; Liu, Y.J.; Pulendran, B.; Palucka, K. Immunobiology of dendritic cells. Annu. Rev. Immunol. 2000, 18, 767–811. [Google Scholar] [CrossRef]
- Xu, Y.; Tang, X.; Yang, M.; Zhang, S.; Li, S.; Chen, Y.; Liu, M.; Guo, Y.; Lu, M. Interleukin 10 Gene-Modified Bone Marrow-Derived Dendritic Cells Attenuate Liver Fibrosis in Mice by Inducing Regulatory T Cells and Inhibiting the TGF-β/Smad Signaling Pathway. Mediat. Inflamm. 2019, 2019, 4652596. [Google Scholar] [CrossRef] [Green Version]
- Figliuolo da Paz, V.; Jamwal, D.R.; Gurney, M.; Midura-Kiela, M.; Harrison, C.A.; Cox, C.; Wilson, J.M.; Ghishan, F.K.; Kiela, P.R. Rapid Downregulation of DAB2 by Toll-Like Receptor Activation Contributes to a Pro-Inflammatory Switch in Activated Dendritic Cells. Front. Immunol. 2019, 10, 304. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, J.; Xie, Y.; Shen, J.; Gao, Q.; He, J.; Ma, H.; Ji, Y.; He, Y.; Xiang, M. Photocurable Hydrogel Substrate—Better Potential Substitute on Bone-Marrow-Derived Dendritic Cells Culturing. Materials 2022, 15, 3322. https://doi.org/10.3390/ma15093322
Deng J, Xie Y, Shen J, Gao Q, He J, Ma H, Ji Y, He Y, Xiang M. Photocurable Hydrogel Substrate—Better Potential Substitute on Bone-Marrow-Derived Dendritic Cells Culturing. Materials. 2022; 15(9):3322. https://doi.org/10.3390/ma15093322
Chicago/Turabian StyleDeng, Jiewen, Yao Xie, Jian Shen, Qing Gao, Jing He, Hong Ma, Yongli Ji, Yong He, and Meixiang Xiang. 2022. "Photocurable Hydrogel Substrate—Better Potential Substitute on Bone-Marrow-Derived Dendritic Cells Culturing" Materials 15, no. 9: 3322. https://doi.org/10.3390/ma15093322