Diversity of Phytophthora Species Associated with Quercus ilex L. in Three Spanish Regions Evaluated by NGS



1
Instituto Agroforestal Mediterráneo, Universitat Politècnica de València, Camino de Vera s/n, 46022 Valencia, Spain
2
Instituto de Ciencias de la Vid y del Vino, Consejo Superior de Investigaciones Científicas–Universidad de la Rioja–Gobierno de La Rioja, Ctra. de Burgos km. 6, 26007 Logroño, Spain
*
Author to whom correspondence should be addressed.
Forests 2019, 10(11), 979; https://doi.org/10.3390/f10110979
Submission received: 16 September 2019
/
Revised: 10 October 2019
/
Accepted: 1 November 2019
/
Published: 5 November 2019
(This article belongs to the Special Issue Phytophthora Infestations in Forest Ecosystems)
Abstract
:The diversity of Phytophthora species in declining Fagaceae forests in Europe is increasing in the last years. The genus Quercus is one of the most extended Fagaceae genera in Europe, and Q. ilex is the dominant tree in Spain. The introduction of soil-borne pathogens, such as Phytophthora in Fagaceae forests modifies the microbial community present in the rhizosphere, and has relevant environmental and economic consequences. A better understanding of the diversity of Phytophthora spp. associated with Q. ilex is proposed in this study by using Next Generation Sequencing (NGS) in six Q. ilex stands located in three regions in Spain. Thirty-seven Phytophthora phylotypes belonging to clades 1 to 12, except for clades 4, 5 and 11, are detected in this study, which represents a high diversity of Phytophthora species in holm oak Spanish forests. Phytophthora chlamydospora, P. citrophthora, P. gonapodyides, P. lacustris, P. meadii, P. plurivora, P. pseudocryptogea, P. psychrophila and P. quercina were present in the three regions. Seven phylotypes could not be associated with known Phytophthora species, so they were putatively named as Phytophthora sp. Most of the detected phylotypes corresponded to terrestrial Phytophthora species but aquatic species from clades 6 and 9 were also present in all regions.
1. Introduction
The genus Phytophthora comprises nowadays more than 150 species with a broad host range, and includes well-known plant pathogens that are devastating natural ecosystems [1,2,3]. Fagaceae forests, with Phytophthora hosts such as Quercus, Castanea, Lithocarpus or Fagus species, are an example of declining forests affected by Phytophthora worldwide [4,5,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26]. In Europe, Castanea, Fagus and Quercus are the most extended Fagaceae genera [13]. The diversity of Phytophthora species found in the last thirty years associated with the rhizosphere of these genera, reveals a complex syndrome in the decline of these forests [10,13,27,28,29,30,31]. On one hand, there is the difficulty of diagnosing which Phytophthora species present is the primary pathogen. On the other hand, it is also known that depending on which Phytophthora species is established first, the damage on the host varies [32]. Furthermore, all Phytophthora species present in the rhizosphere compromise forests regeneration, as they cause seedling damping off [13]. Within the genus Phytophthora we can find typical species of riparian ecosystems and/or water bodies that belong to clades 6 and 9. These species behave as saprotrophs or opportunistic organisms, causing in some cases tree decline, nevertheless its role is not well understood [32,33,34,35]. The remaining Phytophthora species included in the other clades (1 to 12, except 6 and 9) have a terrestrial life cycle, although they often end up in watercourses by runoff [13,35,36,37]. Either aquatic or terrestrial Phytophthora species are related to Fagaceae forests’ decline [13].
Global trade increases the risk of unnoticed introductions of alien species into natural ecosystems [38,39]. The introduction of soil-borne pathogens, such as Phytophthora in Fagaceae forests modifies the microbial community present in the rhizosphere, and has relevant environmental and economic consequences [3]. Soil properties, land use, environmental conditions, the host plants and/or the microbial background determine the microbiota composition from a site [40]. Introduced pathogens have to compete with other microorganisms for available resources, which can lead to a decrease of the native microbiota. Hosts that co-evolved with soil microorganisms can adapt more easily to biotic and abiotic stresses, but a shift in the microbiota structure can trigger the host decline [3,40].
Next generation sequencing (NGS) technologies have stood out as an essential tool for environmental and ecological studies [41,42]. Pyrosequencing the Internal Transcribed Spacer (ITS1) is an efficient and accurate NGS technique for the detection and identification of Phytophthora spp. in environmental samples [29,35,43,44,45]. A better understanding of the diversity of Phytophthora spp. associated with a host can be provided by metagenomics, even if these Phytophthora spp. are not the most prevalent pathogens [3,45,46,47]. Nevertheless, a holistic study, combining biological and molecular identification tools, can substantially improve the diagnosis, because in many cases establishing species boundaries via molecular methods it is not an easy task [2,39,43]. In this context, it is interesting to note that samples from the current study were previously subjected to traditional isolation, baiting and real-time polymerase chain reaction (PCR) methods to detect Phytophthora spp. [48]. The objective of the present study was to unravel the Phytophthora community present in these previously studied areas using NGS technology. Thus, the diversity and abundance of Phytophthora spp. in six holm oak forests located in southwestern and eastern Spain were investigated using an amplicon pyrosequencing approach and the implications are further discussed.
2. Materials and Methods
2.1. Study Site and Sampling
The six study areas are holm oak forests located in three different Spanish regions: Holm oak rangelands (“dehesas”) in Extremadura in southwestern Spain (province of Cáceres), four holm oak stands in the Comunidad Valenciana region in eastern Spain (two in Valencia province, one in Castellón province and one in Alicante province) and one holm oak stand in Cataluña in northeastern Spain (Barcelona province). Samplings were conducted in the autumn (fall) and winter season in different years (2012–2015), and in some areas, it was repeated for two consecutive years.
Soil samples (0.5–1 kg approx.) were collected in all the surveys, consisting in a mixture of four subsamples taken from four different points 1 m around the selected holm oak. In the surveys conducted from 2013 to 2015, along with the soil samples, roots from the rhizosphere were also taken to be analyzed. Soil and roots samples were conserved at 5 °C until DNA extraction was performed. Baiting with leaflets of Rhododendron sp., Viburnum tinus L, Quercus ilex L, Quercus suber L, Ceratonia siliqua L. and/or Dianthus caryophyllus L. petals was performed as described in a previous study [48], and the vegetal material from the baitings was conserved at -80 °C until DNA extraction.
In the Extremadura region in 2012, 60 soil samples from the declining and non-declining Q. ilex rhizosphere were collected from 10 “dehesas” during the autumn. In 2013, 54 soil and root samples and 216 baiting samples from 15 “dehesas” were analyzed. In the Comunidad Valenciana region, during 2014 and 2015, holm oak stands were surveyed, generating 26 soil and roots samples and 104 baiting samples from declining trees. In the Cataluña region in 2013, 15 soil and root samples and 45 baiting samples were processed from declining holm oaks.
2.2. DNA Extraction, PCR, and Preparation of the Amplicon Libraries
Each soil sample was passed through a 2 mm sieve to remove the organic matter and gravel. Once it was homogenized, 50–80 g per subsample was lyophilized overnight and pulverized using a FRITSCH Variable Speed Rotor Mill-PULVERISETTE 14 (ROSH, Oberstein, Germany). DNA from each sample was extracted by duplicate with the Power Soil DNA Isolation Kit (MO BIO Laboratories, Carlsbad, CA, USA) following the manufacturer´s protocol. Roots and baiting vegetal material samples were firstly ground using a mortar and pestle under liquid nitrogen.
Healthy leaflets of the different vegetal species used as baitings were included as negative controls. Once homogenized, DNA extraction was performed from 60–80 mg of material per sample using the Power Plant Pro DNA Isolation Kit (MO BIO Laboratories, Carlsbad, CA, USA).
Amplicon libraries were generated with a nested polymerase chain reaction (PCR) according to the methodology described by Català et al. [35]. The expected size of the amplicons ranged from 280 up to 450 bp. The duplicate amplicons obtained from each sample were pooled if they were positive. Negative samples were retried, firstly diluting 10 times the first round PCR product for the second round PCR. If the retried samples kept on being negative, DNA was diluted ten times, and nested PCR was performed without diluting the first round product. Positive amplicons were double purified using the Agencourt AMPureXP Bead PCR Purification protocol (Beckman Coulter Genomics, MA, USA). Samples were sequenced on the GS Junior 454 system (Roche 454 Life Science, Brandford, CT, USA) at the genotyping service facility from the Universitat de València (Burjassot, Spain).
2.3. Bioinformatics Processing and Statistical Analysis
The sequence reads were processed as described in Català et al. [35]. MOTU (Molecular Operational Taxonomic Unit) clustering of the reads was done with the 90% length coverage threshold and a 99% score coverage threshold using BLASTCLUST software [49]. The consensus sequences of the MOTUs were identified using BLAST tool in the Phytophthora Database [50], the MegaBLAST tool from the GenBank database [49] and a customized database. Afterwards, our query sequences from each survey and type of material (soil/vegetal) were aligned separately using MUSCLE [51]. Phylogenetic analyses consisting of maximum likelihood were performed in MEGA 6.06 [52] with the suggested suitable model, where gaps and missing data were treated as complete deletions. The robustness of the topology was evaluated by 1000 bootstrap replications [53].
A Phytophthora operational taxonomic unit (OTU) table was created excluding other genus reads, unaligned sequences and singletons. OTUs represented globally by less than five reads were removed [54]. The resulting quality-filtered dataset was used for the assessment of diversity and richness. The relationship in OTU composition among samples were investigated by calculating Bray Curtis metrics, and visualized by means of PCoA plots. Samples without Phytophthora reads were excluded for the diversity analyses. Biodiversity indices and principle statistics analyses on taxonomic profiles were analyzed in the tool MicrobiomeAnalyst [55].
3. Results
3.1. Sequencing Results
In the Extremadura region, 66,732 total ITS1 sequences were generated from the pyrosequenced soil samples collected in 2012. These had an average length of 308 bp, and only 61,576 high quality sequences were considered for the analysis after trimming, and excluding singletons; 48.7% of the sequences were identified from declining trees, and 51.3% from non-declining trees.
From samples collected in 2013 in the Extremadura region, 377,799 ITS1 sequences reads were obtained with an average length of 302 bp. After trimming and excluding singletons, 317,961 high quality sequences were used for the analysis; 103,333 from the root samples (45.05% from declining trees and 54.95% from non-declining trees), 56,112 sequences from the soil samples (45.3% from declining trees and 54.7% from non-declining trees) and 158,516 sequences from the baiting samples (51.68% from declining trees and 48.32% from non-declining trees). No Phytophhtora phylotypes are detected from the negative baiting controls.
In the Cataluña region, 63,105 total ITS1 sequences were generated from the pyrosequenced samples. These had an average length of 283 bp, and only 55,230 high quality sequences were considered for the analysis after trimming and excluding singletons (28,381 from roots, 9887 from soils and 16,962 from baitings).
In the Comunidad Valenciana region, 177,398 ITS1 raw sequences with an average length of 309 bp were obtained. After trimming and excluding singletons, 78,962 high quality sequences were considered for the analysis (3,977 from sampled roots, 18,861 from soils and 56,124 from baitings).
3.2. Identification of Phytophthora Phylotypes
3.2.1. Extremadura Region
A total of 33 different Phytophthora phylotypes are identified in the Extremadura region during the surveys conducted in 2012 and 2013 (Table 1). Detected phylotypes belong to all clades except for clades 4, 5 and 11, the clade 6 having the highest number of Phytophthora phylotypes detected (Table 1). In 2012, 20 Phytophthora phylotypes were obtained, and 15 of them identified to the species level: Phytophthora bilorbang Aghighi, G.E. Hardy, J.K. Scott & T.I. Burgess, Phytophthora botryosa Chee, Phytophthora cambivora (Petri) Buisman, Phytophthora chlamydospora Brasier and Hansen, Phytophthora cinnamomi Rands., Phytophthora cryptogea Pethybr. and Laff., Phytophthora gemini Man in ’t Veld, Rosendahl, Brouwer and de Cock, Phytophthora gonapodyides (H.E. Petersen) Buisman, Phytophthora hydropathica Hong and Gallegly, Phytophthora lacustris Brasier, Cacciola, Nechwatal, Jung and Bakonyi, Phytophthora lagoariana, Phytophthora multivora Scott and Jung, Phytophthora plurivora Jung and Burgess, Phytophthora psychrophila Jung and Hansen, Phytophthora quercina Jung and Phytophthora riparia Reeser, Sutton and Hansen. One phylotype belong to complex Phytophthora uliginosa-europaea as the ITS1 region used in the assay could not resolve its identity. Three phylotypes do not correspond to any Phytophthora sequence included in databases; therefore, these putative new species are named Phytophthora sp.1, Phytophthora sp.2 and Phytophthora sp.3.
In 2013, 25 Phytophthora phylotypes were recovered in Extremadura and 22 of them were identified to the species level: Phytophthora taxon ballota, P. bilorbang, P. chlamydospora, P. cinnamomi, Phytophthora citrophthora (R.E. Sm. and E.H. Sm.) Leonian, Phytophthora clandestina Taylor, Pascoe and Greenhalgh, P. cryptogea, Phytophthora gallica Jung and Nechwatal, P. gonapodyides, P. hydropathica, Phytophthora insolita Ann and Ko, Phytophthora sp. kelmania, P. lacustris, Phytophthora lactucae Bertier, Brouwer and de Cock, P. lagoariana, Phytophthora meadii McRae, Phytophthora megasperma Drechsler, Phytophthora sp. palustris, P. plurivora, Phytophthora pseudocryptogea Safaiefarahani, Mostowfizadeh, G.E. Hardy, and T.I. Burgess, P. psychrophila, P. quercina and Phytophthora rosacearum Hansen and Wilcox. The ITS1 region used in the assay could not resolve the identity of one Phytophthora phylotype complex: Phytophthora uliginosa-europaea. One Phytophthora phylotype does not match with any Phytophthora sequence included in the databases, therefore this putative new species is named as Phytophthora sp.4.
3.2.2. Cataluña Region
Fourteen Phytophthora phylotypes are identified in the Cataluña region during the survey conducted in 2013, belonging to clades 2, 3, 6, 7, 8, 9 and 12 (Table 1). Thirteen phylotypes are identified to the species level: P. chlamydospora, P. cinnamomi, P. citrophthora, P. gonapodyides, P. hydropathica, P. insolita, P. lacustris, P. lagoariana, P. meadii, P. plurivora P. pseudocryptogea, P. psychrophila and P. quercina. One Phytophthora phylotype does not match with any Phytophthora sequence included in the databases, therefore this putative new species is named Phytophthora sp.5.
3.2.3. Comunidad Valenciana Region
The Phytophthora phylotypes identified in Comunidad Valenciana during the surveys conducted in the period 2014-2015 is shown in Table 1. Sixteen Phytophthora phylotypes are identified in Comunidad Valenciana belonging to clades 1, 2, 3, 6, 7, 8 and 12 (Table 1). Thirteen phylotypes are identified to the species level: P. taxon ballota, P. chlamydospora, P. citrophthora, P. gonapodyides, P. sp. kelmania, P. lacustris, P. meadii, P. megasperma, P. plurivora P. pseudocryptogea, P. psychrophila, P. quercina and Phytophthora tentaculata Kröber and Marwitz. The ITS1 region used in the assay could not resolve the identity of one Phytophthora phylotype complex: P. uliginosa-europaea. Two Phytophthora phylotypes do not match with any Phytophthora sequences included in the databases and thus, these putative new species are named as Phytophthora sp.6. and Phytophthora sp.7.
3.2.4. Phytophthora Diversity
The relative abundance of Phytophthora species detected from 2013 to 2015 in the three studied regions is shown in Figure 1A. The Extremadura region presents the greatest diversity of Phytophthora species. According to the type of material used for Phytophthora isolation, baiting material has the highest relative abundance of Phytophthora species, followed by roots and soils (Figure 1B).
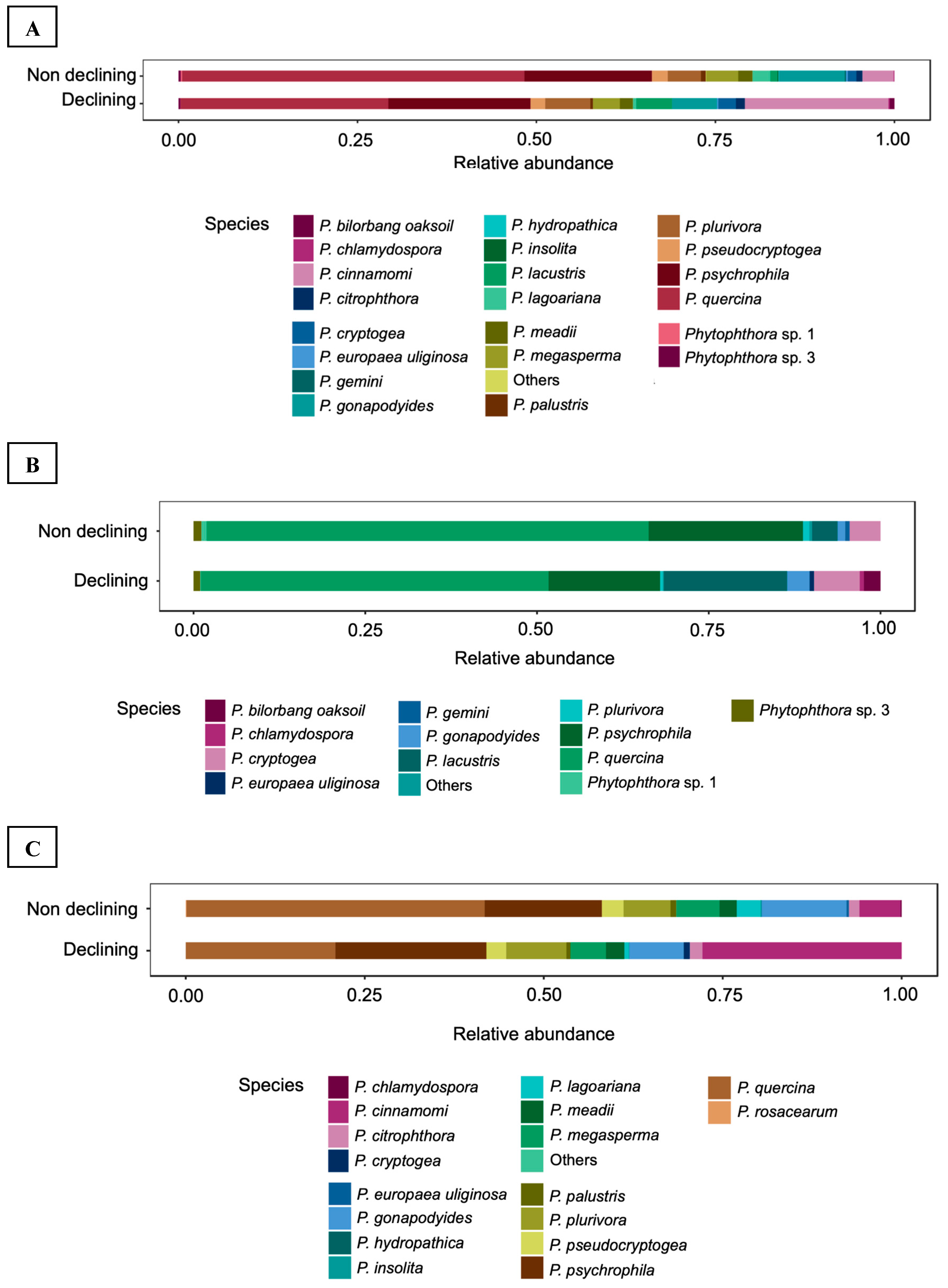
Regarding symptomatology, non-declining holm oak samples in the Extremadura present a higher diversity of Phytophthora species either in the 2013 survey or analyzing 2012 and 2013 together, than samples from declining trees (Figure 2A,C), but this diversity was slightly higher in declining samples in the 2012 survey (Figure 2B).
The ANOVA shows that the alpha diversity was significantly higher in the Extremadura region, followed by Comunidad Valenciana and Cataluña (Figure 3). Both the Chao 1 estimator (p value < 0.05) and the Shannon index (p value < 0.01) were significant according to the factor region (Table 2). Regarding the factor material, the Shannon index was significant (p value = 0.033), showing greater diversity of Phytophthora species when using baiting material, while the Chao 1 estimator was non-significant (p value = 0.091) (Table 2).
In the Extremadura region where the factor symptomatology was studied, in 2012 alpha diversity measured by the Chao1 estimator results was significant for the factor symptomatology (p value = 0.0218), but the Shannon Index was non-significant (p value = 0.1303). Nevertheless, in 2013 the ANOVA shows that both estimators were significant for the factor material and non-significant for the factor symptomatology (Table 3).
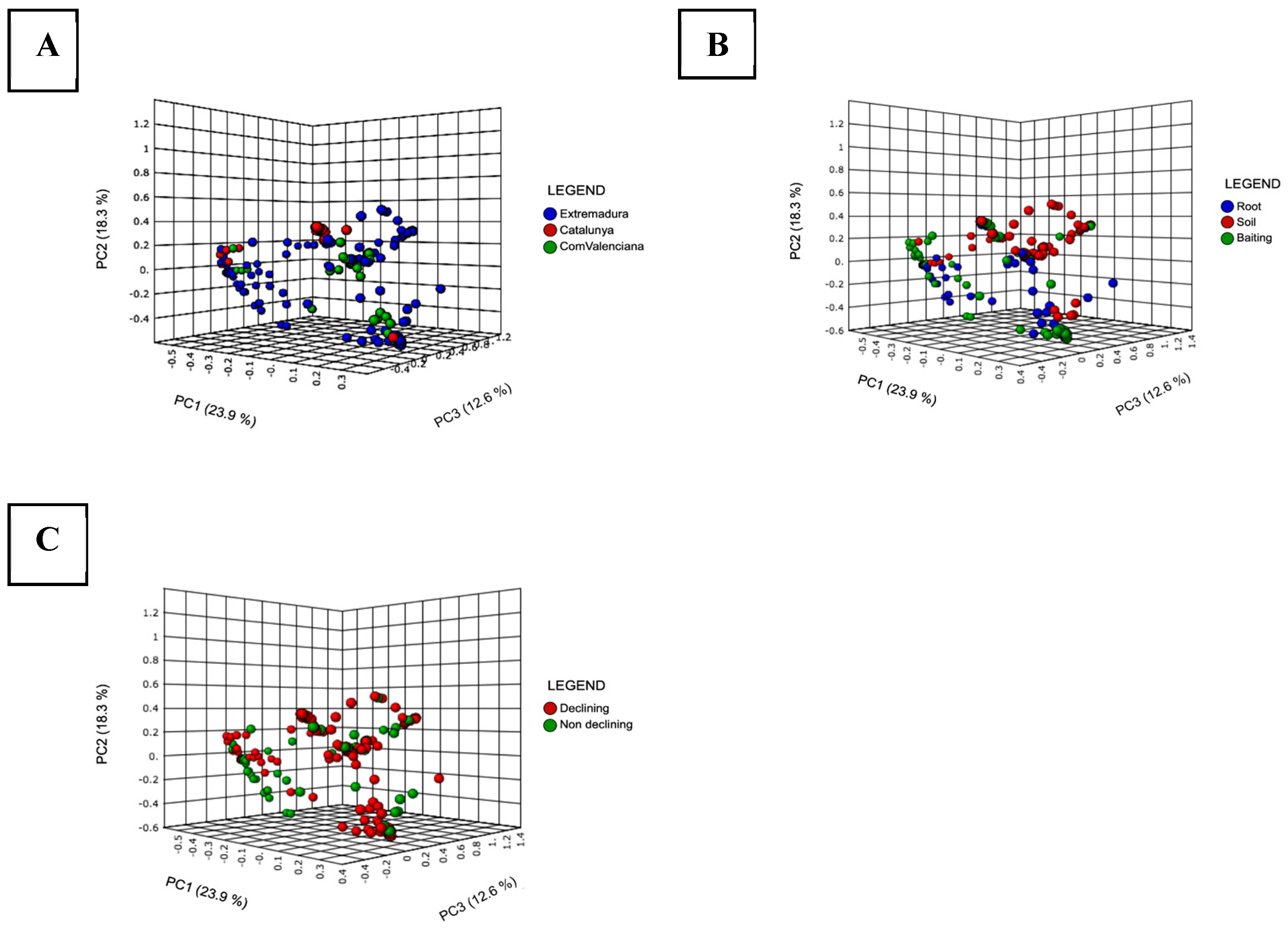
The principal coordinates analysis (PCoAs) of Bray Curtis show that the Phytophthora population in Extremadura had more variation in the number of species present, although there were similarities with the populations of the other two regions (Figure 4A). There are no significant differences in the diversity of Phytophthora species detected in the type of material used or the symptomatology of the sampled trees (Figure 4B,C).
Nine Phytophthora phylotypes were detected in the three regions (24.3%): P. citrophthora, P. meadii, P. plurivora, P. psychrophila, P. quercina, P. chlamydospora, P. gonapodyides, P. lacustris and P. pseudocryptogea (Figure 5A). Regarding the type of material, 11 Phytophthora phylotypes were common to root, soil and baiting material (28.9%): P. plurivora, P. psychrophila, P. quercina, P. chlamydospora, P. gonapodyides, P. lacustris, P. megasperma, P. cinnamomi, P. uliginosa-europaea, P. insolita and P. lagoariana (Figure 5B). Finally, 20 Phytophthora phylotypes were present in declining and non-declining holm oak trees, which represent 54.05% of the detected Phytophthora phylotypes (Figure 5C).
4. Discussion
Thirty-seven Phytophthora phylotypes belonging to clades 1 to 12, except for clades 4, 5 and 11, were detected in this study, which represents a high diversity of Phytophthora species in holm oak Spanish forests compared to previous Spanish NGS studies [44,56]. Phytophthora chlamydospora, P. citrophthora, P. gonapodyides, P. lacustris, P. meadii, P. plurivora, P. pseudocryptogea, P. psychrophila and P. quercina were present in the three regions. Seven of these phylotypes cannot be associated with known Phytophthora species, so they are putatively named as Phytophthora sp. Most of the detected phylotypes correspond to terrestrial Phytophthora species, but aquatic species from clades 6 and 9 were also present in all regions. In general, the most abundant phylotypes in the study are P. quercina, followed by P. psychrophila, P. cinnamomi and P. plurivora; all of them terrestrial species. Our results concur with Català et al. [44], which reported P. quercina and P. psychrophila as the most relevant species associated with Q. ilex in eastern Spain. Nevertheless, they differ from Ruiz Gómez et al. [56] describing P. plurivora, P. quercina, P. cinnamomi and P. cactorum as the most frequent detected species in Andalucía holm oak rangelands (dehesas) located in southern Spain.
In the Extremadura region, all Phytophthora clades are present with the exception of clades 4, 5 and 11. Terrestrial species dominate the Phytophthora community as P. quercina 30%, P. psychrophila 17% and P. cinnamomi 15%, are the most abundant species. Nevertheless, also aquatic species are identified, such as P. gonapodyides 12% and P. megasperma 7%, which follow in abundance. Hence, a mixture of Q. ilex pathogenic terrestrial and aquatic species is identified in the Extremadura region. In the Cataluña region, the Phytophthora community associated with the studied Q. ilex stand was dominated again by terrestrial species (P. plurivora 42%, P. quercina 22% and P. psychrophila 8%), followed by aquatic species (P. hydropathica 11%, P. lagoariana 6% and P. gonapodyides 5%). Clades 1, 4, 5, 8, 10 and 11 were not detected in the studied holm oaks. In the Comunidad Valenciana region, the Phytophthora community is made up primarily by terrestrial Phytophthora species from clades 1, 2, 3, 7, 8 and 12 (P. psychrophila 27%, P. quercina 26%, P. taxon ballota 11%, P. plurivora 10%); although there are also present in lower abundance aquatic species from clade 6 such as P. chlamydospora 7%, P. gonapodyides 2% or P. megasperma 3%.
The weather conditions of the Extremadura region were more favorable for Phytophthora development in 2013 than in 2012, since in 2012 Extremadura received less precipitation than in 2013, as reported in 2018 by Mora-Sala et al. [48]. Redondo et al. [57] state that there is a decrease in the diversity of terrestrial Phytophthora communities when temperature and precipitation decreases, precipitation being the main driving factor, except for clades 2 and 8, in which temperature was more conditioning. According to Redondo et al. [57], aquatic Phytophthora species are inversely conditioned by temperature and precipitation. The Phytophthora diversity found in Extremadura in 2013 was higher than in 2012, which fits with the parameters stated in the study of Redondo et al. [57]. Moreover, more types of sampling materials (roots and baitings) were used for DNA extraction in 2013 than in 2012, which might potentially increase the phylotype diversity. Redondo et al. [57] conclude that the land use is only significant for aquatic species, having more diversity in urban or agricultural sites than in forests. In our study, we found more species belonging to the clade 6 in Extremadura oak rangelands, such as P. gonapodyides or P. megasperma, which are man-made forests, although they are also identified in lower abundance in Cataluña and Comunidad Valenciana. Clade 9 species (P. hydropathica and P. lagoariana) are also detected in Extremadura and in Cataluña.
Phytophthora cinnamomi is an important and devastating Quercus pathogen in Spain [27,58,59,60,61], although it is not the most abundant species detected in the Quercus forests studied. The pathogen is significantly more abundant in declining trees than in non-declining trees, as previously reported by Mora-Sala et al. using qPCR and traditional isolation methods [48], and that also corroborates the metabarcoding study of Ruiz Gómez et al. [56]. Phytophthora cinnamomi is not detected in Comunidad Valenciana.
This is in agreement with previously published reports, and may be due to the unsuitable conditions for the pathogen development in the area consisting on primarily calcareous soils with a high pH [26,44,48,62].
Phytophthora quercina is a specific oak pathogen, present in the three studied regions, that probably has co-evolved with Q. ilex in Spain, rotting the trees slowly and progressively. As P. quercina was not described until 1999 [8], it could not be associated with the studies of the decline of Spanish holm oak conducted in the past. In addition, P. quercina is a difficult pathogen to isolate, due to its slow growth, thus its detection was not always possible in previous studies [8,15,26,30]. Moreover, as generally P. cinnamomi was present, and P. quercina is not a fast growing pathogen, the decline was associated with P. cinnamomi. Although recent studies report the presence of P. quercina, it is thought of as not being as frequent as P. cinnamomi. Mora-Sala et al. demonstrate that P. quercina is more frequent than P. cinnamomi, not only in Comunidad Valenciana, but also in the Extremadura and Cataluña regions by qPCR [48]. Ruiz Gómez et al. [56] report P. quercina as the fourth-most abundant oomycete in Andalucía holm oak rangelands, while P. cinnamomi stands in the ninth position. The present study based on amplicon pyrosequencing supports our previous results using qPCR and verifies that this pathogen is highly abundant in the studied Quercus Spanish regions.
Phytophthora psychrophila is detected in high abundance in the three regions sampled, especially in Comunidad Valenciana, either in declining or in non-declining trees. This species was previously related to Quercus spp. dieback in Spain where it is apparently well distributed and it is only able to be isolated during the winter and spring/autumn periods [10,26,56]. As Pérez-Sierra et al. report in 2013, it is perfectly adapted to xeric Mediterranean conditions due to the thick wall of its resting structures that enables it to overcome unsuitable environmental conditions. Pathogenicity tests with Q. ilex seedlings demonstrate that P. psychrophila is considered an aggressive pathogen, causing dieback of the root system, mainly the fine roots, necrotic lesions and open cankers [26,48].
Phytophthora plurivora is significantly detected in roots and it is even present in all regions, resulting more abundantly in the Cataluña region, where it is the most frequent detected species. Phytophthora plurivora is a widespread species in Europe, which had been already detected in Q. ilex in Spain [44,56], and its pathogenicity to Q. ilex is demonstrated in a previous study [63] causing the absence of fine roots, necrotic lesions, open cankers, dieback of the whole root system and collar rot. Ruiz Gómez et al. [56] report P. plurivora as the most abundant oomycete detected in the Spanish oak rangelands surveyed in their study. Phytophthora plurivora in our study seems to be more associated with declining Q. ilex trees, as Ruiz Gómez et al. [56] report. The homothallic behavior of Phytophthora species, as P. plurivora or P. psychrophila, facilitates its reproduction and establishment in new areas, boosting the risk of forest decline [10,26,29,36,57,64].
From the aquatic species detected, P. gonapodyides was previously detected affecting Q. ilex in Spain [26,33,48]. Phytophthora megasperma is reported to reduce the root system [26,63] although this species is considered an opportunistic oak pathogen present in oak forests [9,15,16]. The role of the remaining aquatic Phytophthora species detected in the Q. ilex decline is still unknown.
The sequencing results support the presence of P. taxon ballota in two forests of the Comunidad Valenciana and in Extremadura regions. This uncultured phylotype was previously detected in oak forests in Comunidad Valenciana [44], but it is the first time that it has been detected outside the Comunidad Valenciana. The designation of new phylotypes from environmental DNA remains a committed issue, but it is reported in other studies [65,66,67]. Both in previous studies and the present study, no Phytophthora culture was isolated that coincided with this proposed phylotype using traditional methods. Further surveys targeting this organism should be performed to try the isolation.
This study detected the presence of P. pseudocryptogea in all three regions, and it has also been isolated from other Fagaceae such as Castanea sativa in the North of Spain (Mora-Sala and Català unpublished), suggesting that it is well established in Spain and probably in the past many isolates identified as P. cryptogea, were actually P. pseudocryptogea. In 2018, P. pseudocryptogea was firstly reported in Extremadura region from Q. ilex rhizosphere [48] and it has just been reported in Sicily affecting Q. ilex [36], so it seems to be well established in the Mediterranean basin.
The pathogenicity on Q. ilex has not been tested, but its pathogenicity on other Fagaceae (Mora-Sala and Català, unpublished), suggests that this species could contribute to the oak decline.
Phytophthora community diversity recovered from Fagaceae forests in Europe has increased in the last years [8,9,10,13,16,20,21,23,24,26,29,30,36,44,64,68]. The implementation of NGS technologies to forests surveys helps to improve the knowledge about the Phytophthora spp. diversity associated with Fagaceae forests and to identify possible new introduced species [3,29,44,45,47]. Phytophthora spp. can adapt to a wide variety of environmental conditions [38]. In Spain, the oak decline due to Phytophthora spp. is related to the effect of the water stress; seasonal droughts followed by floods enhance the root damage induced by Phytophthora species. The versatility of these species to cope with the changing scenarios and the increase of extreme weather conditions that are occurring nowadays, focuses the attention on this destructive genus. The composition of Phytophthora species in these ecosystems is changing because of their adaptation to new environmental conditions and new species introduction. New technologies help us improve knowledge about species diversity in these new scenarios. In the present study, NGS reveals a higher Phytophthora species diversity than previously detected by traditional isolation, baiting and qPCR. However, the best approach should combine all available methodologies for a correct Phytophthora diagnosis, facilitating a quick answer facing the potential introduction of new and/or quarantine organisms.
5. Conclusions
The use of amplicon pyrosequencing reveals a high diversity of Phytophthora species associated with Q. ilex. Baiting material has the highest relative abundance of Phytophthora species. The highest alpha diversity is obtained in the region of Extremadura, the diversity of Comunidad Valenciana and Cataluña being lower following this order. In general terms, the Phytophthora diversity is highest in non-declining Q. ilex than in declining trees. The implementation of molecular tools in Phytophthora forests monitoring, complement and help to overcome the limitations of traditional methods, being useful to improve the knowledge about the real composition of the species present in these ecosystems.
Author Contributions
B.M.-S. conducted field sampling, performed experimental work and data analysis, discussed the results and wrote the paper. D.G. performed the data analysis. M.B. participated in the data analyses, discussed the results and revised the manuscript. P.A.C. designed the study, participated in the experimental work and revised the manuscript.
Acknowledgments
We would like to thank M. León from the Instituto Agroforestal Mediterráneo-UPV (Spain) for its technical assistance. This research was supported by funding from the project AGL2011-30438-C02-01 (Ministerio de Economía y Competitividad, Spain) and Euphresco [Instituto Nacional de Investigación y Tecnología Agraria y Agroalimentaria (EUPHESCO-CEP: “Current and Emerging Phytophthoras: Research Supporting Risk Assesssment and Risk Management”)].
Conflicts of Interest
The authors declare no conflict of interest.
References
- Erwin, D.C.; Ribeiro, O.K. Phytophthora Diseases Worldwide; APS Press: St. Paul, MN, USA, 1996; p. 562. ISBN 0-89054-212-0. [Google Scholar]
- Mideros, M.F.; Turissini, D.A.; Guayazán, N.; Ibarra-Avila, H.; Danies, G.; Cárdenas, M.; Lagos, L.E.; Myers, K.; Tabima, J.; Goss, E.M.; et al. Phytophthora betacei, a new species within Phytophthora clade 1c causing late blight on Solanum betaceum in Colombia. Persoonia 2018, 41, 39–55. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, É.D.; Duceppe, M.O.; Bérubé, J.A.; Kimoto, T.; Lemieux, C.; Bilodeau, G.J. Screening for exotic forest pathogens to increase survey capacity using metagenomics. Phytopathology 2018, 108, 1509–1521. [Google Scholar] [CrossRef] [PubMed]
- Shearer, B.L.; Tippett, J.T. Jarrah Dieback: The Dynamics and Management of Phytophthora Cinnamomi in the Jarrah (Eucalyptus Marginata) Forests of South-Western Australia; Lewis, M., Ed.; Department of Conservation and Land Management: Como, WA, Australia, 1989; ISSN 1032-8106.
- Brasier, C.M. Oak tree mortality in Iberia. Nature 1992, 360, 539. [Google Scholar] [CrossRef]
- Brasier, C.M. Phytophthora cinnamomi as a contributory factor on European oak declines. In Recent Advances in Studies on Oak Decline, Proceedings of the International Congress, Brindisi, Italy, 13–18 September 1992; Luisi, N., Lerario, P., Vannini, A., Eds.; Università degli Studi: Bari, Italy, 1992; pp. 49–58. [Google Scholar]
- Jung, T.; Blaschke, H.; Neumann, P. Isolation, identification and pathogenicity of Phytophthora species from declining oak stands. Eur. J. Forest Pathol. 1996, 26, 253–272. [Google Scholar] [CrossRef]
- Jung, T.; Cooke, D.E.L.; Blaschke, H.; Duncan, J.M.; Oßwald, W. Phytophthora quercina sp. nov., causing root rot of European oaks. Mycol. Res. 1999, 103, 785–798. [Google Scholar] [CrossRef]
- Jung, T.; Blaschke, H.; Oßwald, W. Involvement of soilborne Phytophthora species in Central European oak decline and the effect of site factors on the disease. Plant Pathol. 2000, 49, 706–718. [Google Scholar] [CrossRef]
- Jung, T.; Hansen, E.M.; Winton, L.; Oßwald, W.; Delatour, C. Three new species of Phytophthora from European oak forests. Mycol. Res. 2002, 106, 397–411. [Google Scholar] [CrossRef]
- Jung, T.; Nechwatal, J.; Cooke, D.E.L.; Hartmann, G.; Blaschke, M.; Oßwald, W.; Duncan, J.M.; Delatour, C. Phytophthora pseudosyringae sp. nov., a new species causing root and collar rot of deciduous tree species in Europe. Mycol. Res. 2003, 107, 772–789. [Google Scholar] [CrossRef]
- Jung, T.; Hudler, G.W.; Jensen-Tracy, S.L.; Griffiths, H.M.; Fleischmann, F.; Oßwald, W. Involvement of Phytophthora species in the decline of European beech in Europe and the USA. Mycologist 2005, 19, 159–166. [Google Scholar] [CrossRef]
- Jung, T.; Jung Horta, M.; Cacciola, S.O.; Cech, T.; Bakonyi, J.; Seress, D. Multiple new cryptic pathogenic Phytophthora species from Fagaceae forests in Austria, Italy and Portugal. IMA Fungus 2017, 8, 219–244. [Google Scholar] [CrossRef] [Green Version]
- Robin, C.; Desprez-Loustau, M.L.; Capron, G.; Delatour, C. First record of Phytophthora cinnamomi on cork and holm oaks in France and evidence of pathogenicity. Ann. Scie. Forest. 1998, 55, 869–883. [Google Scholar] [CrossRef]
- Hansen, E.; Delatour, C. Phytophthora species in oak forests of north-east France. Ann. For. Sci. 1999, 56, 539–547. [Google Scholar] [CrossRef]
- Vettraino, A.M.; Barzanti, G.P.; Bianco, M.C.; Ragazzi, A.; Capretti, P.; Paoletti, E.; Vannini, A. Occurrence of Phytophthora species in oak stands in Italy and their association with declining oak trees. For. Pathol. 2002, 32, 19–28. [Google Scholar] [CrossRef]
- Vettraino, A.M.; Morel, O.; Perlerou, C.; Robin, C.; Diamandis, S.; Vannini, A. Occurrence and distribution of Phytophthora species in European chestnut stands, and their association with Ink Disease and crown decline. Eur. J. Plant Pathol. 2005, 111, 169–180. [Google Scholar] [CrossRef]
- Rizzo, D.M.; Garbelotto, M.; Davidson, J.M.; Slaughter, G.W.; Koike, S.T. Phytophthora ramorum as the cause of extensive mortality of Quercus spp. and Lithocarpus densiflorus in California. Plant Dis. 2002, 86, 205–214. [Google Scholar] [CrossRef]
- Rizzo, D.M.; Garbelotto, M. Sudden Oak Death: Endangering California and Oregon Forest Ecosystems. Front. Ecol. Environ. 2003, 1, 197–204. [Google Scholar] [CrossRef]
- Balci, Y.; Halmschlager, E. Incidence of Phytophthora species in oak forests in Austria and their possible involvement in oak decline. For. Pathol. 2003, 33, 157–174. [Google Scholar] [CrossRef]
- Balci, Y.; Halmschlager, E. Phytophthora species in oak ecosystems in Turkey and their association with declining oak trees. Plant Pathol. 2003, 52, 694–702. [Google Scholar] [CrossRef]
- Balci, Y.; Balci, S.; Eggers, J.; MacDonald, W.L.; Juzwik, J.; Long, R.P.; Gottschalk, K.W. Phytophthora species associated with forest soils in Eastern and North-central U.S. oak ecosystems. Plant Dis. 2007, 91, 705–710. [Google Scholar] [CrossRef]
- Balci, Y.; Balci, S.; MacDonald, W.L.; Gottschalk, K.W. Relative susceptibility of oaks to seven species of Phytophthora isolated from oak forest soils. For. Pathol. 2008, 38, 394–409. [Google Scholar] [CrossRef]
- Balci, Y.; Balci, S.; Blair, J.E.; Park, S.Y.; Kang, S.; MacDonald, W.L. Phytophthora quercetorum sp. nov., a novel species isolated from eastern and north-central U.S. oak forest soils. Mycol. Res. 2008, 112, 906–916. [Google Scholar] [CrossRef] [PubMed]
- Vannini, A.; Vettraino, A.M. Phytophthora cambivora . For. Phytophthoras 2011, 1. [Google Scholar] [CrossRef]
- Pérez-Sierra, A.; López-García, C.; León, M.; García-Jiménez, J.; Abad-Campos, P.; Jung, T. Previously unrecorded low-temperature Phytophthora species associated with Quercus decline in a Mediterranean forest in eastern Spain. For. Pathol. 2013, 43, 331–339. [Google Scholar] [CrossRef]
- Brasier, C.M. Phytophthora cinnamomi and oak decline in southern Europe-Environmental constraints including climate change. Ann. Sci. For. 1996, 53, 347–358. [Google Scholar] [CrossRef]
- Jung, T.; Orlikowski, L.; Henricot, B.; Abad-Campos, P.; Aday, A.G.; Aguín Casal, O.; Bakonyi, J.; Cacciola, S.O.; Cech, T.; et al. Widespread Phytophthora infestations in European nurseries put forest, semi-natural and horticultural ecosystems at high risk of Phytophthora diseases. For. Pathol. 2016, 46, 134–163. [Google Scholar] [CrossRef]
- Vannini, A.; Bruni, N.; Tomassini, A.; Franceschini, S.; Vettraino, A.M. Pyrosequencing of environmental soil samples reveals biodiversity of the Phytophthora resident community in chestnut forests. FEMS Microbiol. Ecol. 2013, 85, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Jankowiak, R.; Stępniewska, H.; Bilański, P.; Kolařík, M. Occurrence of Phytophthora plurivora and other Phytophthora species in oak forests of southern Poland and their association with site conditions and the health status of trees. Folia Microbiol. 2014, 59, 531. [Google Scholar] [CrossRef]
- Scanu, B.; Linaldeddu, B.T.; Deidda, A.; Jung, T. Diversity of Phytophthora species from declining Mediterranean maquis vegetation, including two new species, Phytophthora crassamura and P. ornamentata sp. nov. PLoS ONE 2015, 10, e0143234. [Google Scholar] [CrossRef]
- Corcobado, T.; Miranda-Torres, J.J.; Martín-García, J.; Jung, T.; Solla, A. Early survival of Quercus ilex subspecies from different populations after infections and co-infections by multiple Phytophthora species. Plant Pathol. 2017, 66, 792–804. [Google Scholar] [CrossRef]
- Corcobado, T.; Cubera, E.; Pérez-Sierra, A.; Jung, T.; Solla, A. First report of Phytophthora gonapodyides involved in the decline of Quercus ilex in xeric conditions in Spain. New Dis. Rep. 2010, 22, 33. [Google Scholar] [CrossRef]
- Hansen, E.M.; Reeser, P.W.; Sutton, W. Phytophthora beyond agriculture. Annu. Rev. Phytopathol. 2012, 50, 359–378. [Google Scholar] [CrossRef] [PubMed]
- Català, S.; Pérez-Sierra, A.; Abad-Campos, P. The use of genus-specific amplicon pyrosequencing to assess Phytophthora species diversity using eDNA from soil and water in northern Spain. PLoS ONE 2015, 10, e0119311. [Google Scholar] [CrossRef] [PubMed]
- Jung, T.; La Spada, F.; Pane, A.; Aloi, F.; Evoli, M.; Horta Jung, M.; Scanu, B.; Faedda, R.; Rizza, C.; Puglisi, I.; et al. Diversity and Distribution of Phytophthora Species in Protected Natural Areas in Sicily. Forests 2019, 10, 259. [Google Scholar] [CrossRef]
- Jung, T.; Pérez-Sierra, A.; Durán, A.; Jung, M.H.; Balci, Y.; Scanu, B. Canker and decline diseases caused by soil- and airborne Phytophthora species in forests and woodlands. Persoonia 2018, 40, 182–220. [Google Scholar] [CrossRef]
- Brasier, C.M. The biosecurity threat to the UK and global environment from international trade in plants. Plant Pathol. 2008, 57, 792–808. [Google Scholar] [CrossRef]
- O’Brien, P.; Williams, N.; Hardy, G. Detecting Phytophthora. Crit. Rev. Microbiol. 2009, 35, 169–181. [Google Scholar] [CrossRef]
- Berlanas, C.; Berbegal, M.; Elena, G.; Laidani, M.; Cibriain, J.F.; Sagües, A.; Gramaje, D. The Fungal and Bacterial Rhizosphere Microbiome Associated with Grapevine Rootstock Genotypes in Mature and Young Vineyards. Front. Microbiol. 2019, 10, 1142. [Google Scholar] [CrossRef]
- Taberlet, P.; Coissac, E.; Hajibabaei, M.; Rieseberg, L.H. Environmental DNA. Mol. Ecol. 2012, 21, 1789–1793. [Google Scholar] [CrossRef]
- Oulas, A.; Pavloudi, C.; Polymenakou, P.; Pavlopoulos, G.A.; Papanikolaou, N.; Kotoulas, G.; Arvanitidis, C.; Iliopoulos, L. Metagenomics: Tools and Insights for Analyzing Next-Generation Sequencing Data Derived from Biodiversity Studies. Bioinform. Biol. Insights 2015, 9, BBI-S12462. [Google Scholar] [CrossRef]
- Vettraino, A.M.; Bonats, P.; Tomassini, A.; Bruni, N.; Vannini, A. Pyrosequencing as a tool for detection of K species: Error rate and risk of false MOTUs. Lett. Appl. Microbiol. 2012, 55, 390–396. [Google Scholar] [CrossRef]
- Català, S.; Berbegal, M.; Pérez-Sierra, A.; Abad-Campos, P. Metabarcoding and development of new real-time specific assays reveal Phytophthora species diversity in Holm Oak forests in eastern Spain. Plant Pathol. 2017, 66, 115–123. [Google Scholar] [CrossRef]
- Prigigallo, M.I.; Abdelfattah, A.; Cacciola, S.O.; Faedda, R.; Sanzani, S.M.; Cooke, D.E.L.; Schena, L. Metabarcoding analysis of Phytophthora diversity using genus-specific primers and 454 pyrosequencing. Phytopathology 2016, 106, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Scibetta, S.; Schena, L.; Chimento, A.; Cacciola, S.O.; Cooke, D.E.L. A molecular method to assess Phytophthora diversity in environmental samples. J. Microbiol. Methods 2012, 88, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Burgess, T.I.; McDougall, K.L.; Scott, P.M.; Hardy, G.E.; Garnas, J. Predictors of Phytophthora diversity and community composition in natural areas across diverse Australian ecoregions. Ecography 2018, 42, 565–577. [Google Scholar] [CrossRef]
- Mora-Sala, B.; Berbegal, M.; Abad-Campos, P. The use of qPCR reveals a high frequency of Phytophthora quercina in two Spanish holm oak areas. Forests 2018, 9, 697. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Scheaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSIBLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Park, J.; Park, B.; Veeraraghavan, N.; Jung, K.; Lee, Y.H.; Rossman, A.; Farr, D.; Cline, E.; Grünwald, N.J.; Luster, D.; et al. Phytophthora database: A forensic database supporting the identification and monitoring of Phytophthora. Plant Dis. 2008, 92, 966–972. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Glynou, K.; Nam, B.; Thines, M.; Maciá-Vicente, J.G. Facultative root-colonizing fungi dominate endophytic assemblages in roots of nonmycorrhizal Microthlaspi species. New Phytol. 2018, 217, 1190–1202. [Google Scholar] [CrossRef] [PubMed]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef]
- Ruiz Gómez, F.J.; Navarro-Cerrillo, R.M.; Pérez deLuque, A.; Oβwald, W.; Vannini, A.; Morales-Rodríguez, C. Assessment of functional and structural changes of soil fungal and oomycete communities in holm oak declined dehesas through metabarcoding analysis. Sci. Rep. 2019, 9, 5315. [Google Scholar] [CrossRef] [PubMed]
- Redondo, M.A.; Boberg, J.; Stenlid, J.; Oliva, J. Contrasting distribution patterns between aquatic and terrestrial Phytophthora species along a climatic gradient are linked to functional traits. ISME J. 2018, 12, 2967–2980. [Google Scholar] [CrossRef] [PubMed]
- Brasier, C.M.; Robredo, F.; Ferraz, J.F.P. Evidence for Phytophthora cinnamomi involvement in Iberian oak decline. Plant Pathol. 1993, 42, 140–145. [Google Scholar] [CrossRef]
- Gallego, F.J.; Perez de Algaba, A.; Fernandez-Escobar, R. Etiology of oak decline in Spain. Eur. J. For. Pathol. 1999, 29, 17–27. [Google Scholar] [CrossRef]
- Sánchez, M.E.; Caetano, P.; Ferraz, J.; Trapero, A. Phytophtora disease of Quercus ilex in south-western Spain. For. Pathol. 2002, 32, 5–18. [Google Scholar] [CrossRef]
- Camilo-Alves, C.S.P.; da Clara, M.I.E.; de Almeida Ribeiro, N.M.C. Decline of Mediterranean oak trees and its association with Phytophthora cinnamomi: A review. Eur. J. Forest Res. 2013, 132, 411–432. [Google Scholar] [CrossRef]
- Serrano, M.S.; de Vita, P.; Fernández-Rebollo, P.; Sánchez, M.E. Calcium fertilizers induce soil suppressiveness to Phytophthora cinnamomi root rot of Quercus ilex. Eur. J. Plant Pathol. 2012, 132, 271–279. [Google Scholar] [CrossRef]
- Mora-Sala, B.; Abad-Campos, P.; Berbegal, M. Response of Quercus ilex seedlings to Phytophthora spp. root infection in a soil infestation test. Eur. J. Plant Pathol. 2018, 154, 2115–2225. [Google Scholar] [CrossRef]
- Jung, T.; Burgess, T.I. Re-evaluation of Phytophthora citricola isolates from multiple woody hosts in Europe and North America reveals a new species, Phytophthora plurivora sp. nov. Persoonia 2009, 22, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Ioos, R.; Laugustin, L.; Rose, S.; Tourvieille, J.; Tourvieille de Labrouhe, D. Development of a PCR test to detect the downy mildew causal agent Plasmopara halstedii in sunflower seeds. Plant Pathol. 2007, 56, 209–218. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, X.J.; Chen, C.Q.; Huang, L.L.; Kang, Z.S. A PCR-based assay for detection of Puccinia striiformis f. sp. tritici in wheat. Plant Dis. 2007, 91, 1669–1674. [Google Scholar] [CrossRef] [PubMed]
- Alaei, H.; Baeyen, S.; Maes, M.; Höfte, M.; Heungens, K. Molecular detection of Puccinia horiana in Chrysanthemum × morifolium through conventional and real-time PCR. J. Microbiol. Methods 2009, 76, 136–145. [Google Scholar] [CrossRef]
- Mrázková, M.; Černý, K.; Tomosovsky, M.; Strnadová, V.; Gregorová, B.; Holub, V.; Panek, M.; Havrdová, L.; Hejná, M. Occurrence of Phytophthora multivora and Phytophthora plurivora in the Czech Republic. Plant Prot. Sci. 2013, 49, 155–164. [Google Scholar] [CrossRef]
Figure 1.
Relative abundance of Phytophthora species detected in the three studied regions from 2013 to 2015 showing more than 1% relative abundance of all reads. Species representing less than 1% of the total reads are grouped in “others”. (A): According to the factor region; Extremadura, Comunidad Valenciana (ComValenciana) and Cataluña (Catalunya). (B): According to the factor type of material; soils, roots and baitings. Results of the survey performed in Extremadura in 2012 are excluded in this analysis, since only soil samples were included.
Figure 1.
Relative abundance of Phytophthora species detected in the three studied regions from 2013 to 2015 showing more than 1% relative abundance of all reads. Species representing less than 1% of the total reads are grouped in “others”. (A): According to the factor region; Extremadura, Comunidad Valenciana (ComValenciana) and Cataluña (Catalunya). (B): According to the factor type of material; soils, roots and baitings. Results of the survey performed in Extremadura in 2012 are excluded in this analysis, since only soil samples were included.

Figure 2.
Relative abundance of Phytophthora species showing more than 1% relative abundance of all reads in Extremadura region according to the factor symptomatology. Species representing less than 1% of the total reads are grouped in others. (A): During the surveys conducted in 2012 and 2013. (B): Survey conducted in 2012. (C): Survey conducted in 2013.
Figure 2.
Relative abundance of Phytophthora species showing more than 1% relative abundance of all reads in Extremadura region according to the factor symptomatology. Species representing less than 1% of the total reads are grouped in others. (A): During the surveys conducted in 2012 and 2013. (B): Survey conducted in 2012. (C): Survey conducted in 2013.

Figure 3.
Boxplot showing alpha diversity measures of the Phytophthora diversity according to the factor region: Extremadura, Comunidad Valenciana (ComValenciana) and Cataluña (Catalonia). (A): Chao 1 estimator (p value < 0.05). (B): Shannon index (p value < 0.01).
Figure 3.
Boxplot showing alpha diversity measures of the Phytophthora diversity according to the factor region: Extremadura, Comunidad Valenciana (ComValenciana) and Cataluña (Catalonia). (A): Chao 1 estimator (p value < 0.05). (B): Shannon index (p value < 0.01).

Figure 4.
Principal Coordinate Analysis (PCoA) based on Bray Curtis dissimilarity metrics, showing the distance in the Phytophthora spp. composition according to the different factors studied. (A): Regions surveyed; Extremadura, Comunidad Valenciana (ComValenciana) and Cataluña (Catalunya). (B): Type of material used for the detection; roots, soils and baitings. (C): Symptomatology of the holm oaks sampled: declining and non-declining.
Figure 4.
Principal Coordinate Analysis (PCoA) based on Bray Curtis dissimilarity metrics, showing the distance in the Phytophthora spp. composition according to the different factors studied. (A): Regions surveyed; Extremadura, Comunidad Valenciana (ComValenciana) and Cataluña (Catalunya). (B): Type of material used for the detection; roots, soils and baitings. (C): Symptomatology of the holm oaks sampled: declining and non-declining.

Figure 5.
Venn diagram showing the overlap of operational taxonomic units (OTUs) identified in the Phytophthora population present in the holm oaks surveyed in the study according to the different factors studied. (A): Regions surveyed; Extremadura, Comunidad Valenciana (ComValenciana) and Cataluña (Catalunya), (B): Type of material used for the detection; roots, soils and baitings. (C): Symptomatology of the holm oaks sampled: declining and non-declining.
Figure 5.
Venn diagram showing the overlap of operational taxonomic units (OTUs) identified in the Phytophthora population present in the holm oaks surveyed in the study according to the different factors studied. (A): Regions surveyed; Extremadura, Comunidad Valenciana (ComValenciana) and Cataluña (Catalunya), (B): Type of material used for the detection; roots, soils and baitings. (C): Symptomatology of the holm oaks sampled: declining and non-declining.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Phytophthora phylotypes detected by next generation sequencing (NGS), based on the Internal Transcribed Spacer (ITS1) region.
Table 1.
Phytophthora phylotypes detected by next generation sequencing (NGS), based on the Internal Transcribed Spacer (ITS1) region.
| Phylotypes | Clade | Spanish Regions | |||||
|---|---|---|---|---|---|---|---|
| Extremadura | Cataluña | Comunidad Valenciana | |||||
| Font Roja | Hunde | Pina | Alcublas | ||||
| BAL | 1 | ✓ | ✓ | ✓ | |||
| BIL | 6 | ✓ | |||||
| BOT | 2 | ✓ | |||||
| CAM | 7 | ✓ | |||||
| CHL | 6 | ✓ | ✓ | ✓ | ✓ | ||
| CIN | 7 | ✓ | ✓ | ||||
| CIP | 2 | ✓ | ✓ | ✓ | |||
| CLA | 1 | ✓ | |||||
| CRY | 8 | ✓ | |||||
| GAL | 10 | ✓ | |||||
| GEM | 6 | ✓ | |||||
| GON | 6 | ✓ | ✓ | ✓ | |||
| HYD | 9 | ✓ | ✓ | ||||
| INS | 9 | ✓ | ✓ | ||||
| KEL | 8 | ✓ | ✓ | ||||
| LAC | 6 | ✓ | ✓ | ✓ | ✓ | ||
| LCT | ✓ | ||||||
| LAG | 9 | ✓ | ✓ | ||||
| MEA | 2 | ✓ | ✓ | ✓ | |||
| MEG | 6 | ✓ | ✓ | ||||
| MUL | 2 | ✓ | |||||
| PAS | 9 | ✓ | |||||
| PLU | 2 | ✓ | ✓ | ✓ | ✓ | ||
| PSC | 8 | ✓ | ✓ | ✓ | ✓ | ✓ | |
| PSY | 3 | ✓ | ✓ | ✓ | ✓ | ✓ | |
| QUE | 12 | ✓ | ✓ | ✓ | ✓ | ✓ | |
| RIP | 6 | ✓ | |||||
| ROS | 6 | ✓ | |||||
| TEN | 1 | ✓ | |||||
| ULIG-EUR | 7 | ✓ | ✓ | ||||
| SP.1 | 6 | ✓ | |||||
| SP.2 | 7 | ✓ | |||||
| SP.3 | 8 | ✓ | |||||
| SP.4 | 1 | ✓ | |||||
| SP.5 | 7 | ✓ | |||||
| SP.6 | 3 | ✓ | |||||
| SP.7 | 1 | ✓ | |||||
BAL, P. taxon ballota; BIL, P. bilorbang; BOT, P. botryosa; CAM, P. cambivora; CHL, P. chlamydospora; CIN, P. cinnamomi; CIP, P. citrophthora; CLA, P. clandestina; CRY, P. cryptogea; GAL, P. gallica; GEM, P. gemini; GON, P. gonapodyides; HYD, P. hydropathica; INS, P. insolita; KEL, P. sp. kelmania; LAC, P. lacustris; LCT, P. lactucae; LAG, P. lagoariana; MEA, P. meadii; MEG, P. megasperma; MUL, P. multivora; PAS, P. sp. palustris; PLU, P. plurivora; PSC, P. pseudocryptogea; PSY, P. psychrophila; QUE, P. quercina; RIP, P. riparia; ROS, P. rosacearum; TEN, P. tentaculata; ULIG-EUR, P. uliginosa-P. europaea; SP.1-SP.7, new phylotypes found not identified to the species level.
Table 2.
Analysis of variance (ANOVA) table for the alpha diversity of Phytophthora species detected in the study.
Table 2.
Analysis of variance (ANOVA) table for the alpha diversity of Phytophthora species detected in the study.
| α Diversity | |||
|---|---|---|---|
| Region | Material | Region × Material | |
| Chao 1 estimator | F2,245 = 10.8 p ≤ 0.01 | F2,196 = 1.9 p = 0.091 | F4,188 = 5.5 p ≤ 0.01 |
| Shannon index | F2,245 = 7.1 p ≤ 0.01 | F2,196 = 2.3 p = 0.033 | F4,188 = 4.7 p ≤ 0.01 |
Table 3.
Analysis of variance (ANOVA) tables for the alpha diversity of Phytophthora species detected in Extremadura dehesas during the surveys conducted in 2012 and 2013.
Table 3.
Analysis of variance (ANOVA) tables for the alpha diversity of Phytophthora species detected in Extremadura dehesas during the surveys conducted in 2012 and 2013.
| α Diversity | ||||||
|---|---|---|---|---|---|---|
| Survey | Chao 1 Estimator | Shannon Index | ||||
| Material | Symptoms | Material × Symptoms | Material | Symptoms | Material × Symptoms | |
| 2012 | ndt | F1,47= 2.39 p = 0.0218 | ndt | ndt | F1,47= 1.54 p = 0.1303 | ndt |
| 2013 | F2,126 = 6.03 p = 0.0029 | F1,127 = 1.23 p = 0.2199 | F2,123 = 2.7 p = 0.023 | F2,126 = 4.95 p = 0.0081 | F1,127 = 0.83 p = 0.4085 | F2,123 = 2.4 p = 0.041 |
ndt = not determined.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mora-Sala, B.; Gramaje, D.; Abad-Campos, P.; Berbegal, M. Diversity of Phytophthora Species Associated with Quercus ilex L. in Three Spanish Regions Evaluated by NGS. Forests 2019, 10, 979. https://doi.org/10.3390/f10110979
AMA Style
Mora-Sala B, Gramaje D, Abad-Campos P, Berbegal M. Diversity of Phytophthora Species Associated with Quercus ilex L. in Three Spanish Regions Evaluated by NGS. Forests. 2019; 10(11):979. https://doi.org/10.3390/f10110979
Chicago/Turabian StyleMora-Sala, Beatriz, David Gramaje, Paloma Abad-Campos, and Mónica Berbegal. 2019. "Diversity of Phytophthora Species Associated with Quercus ilex L. in Three Spanish Regions Evaluated by NGS" Forests 10, no. 11: 979. https://doi.org/10.3390/f10110979
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.