Breeding Systems in Diploid and Polyploid Hawthorns (Crataegus): Evidence from Experimental Pollinations of C. monogyna, C. subsphaerica, and Natural Hybrids

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
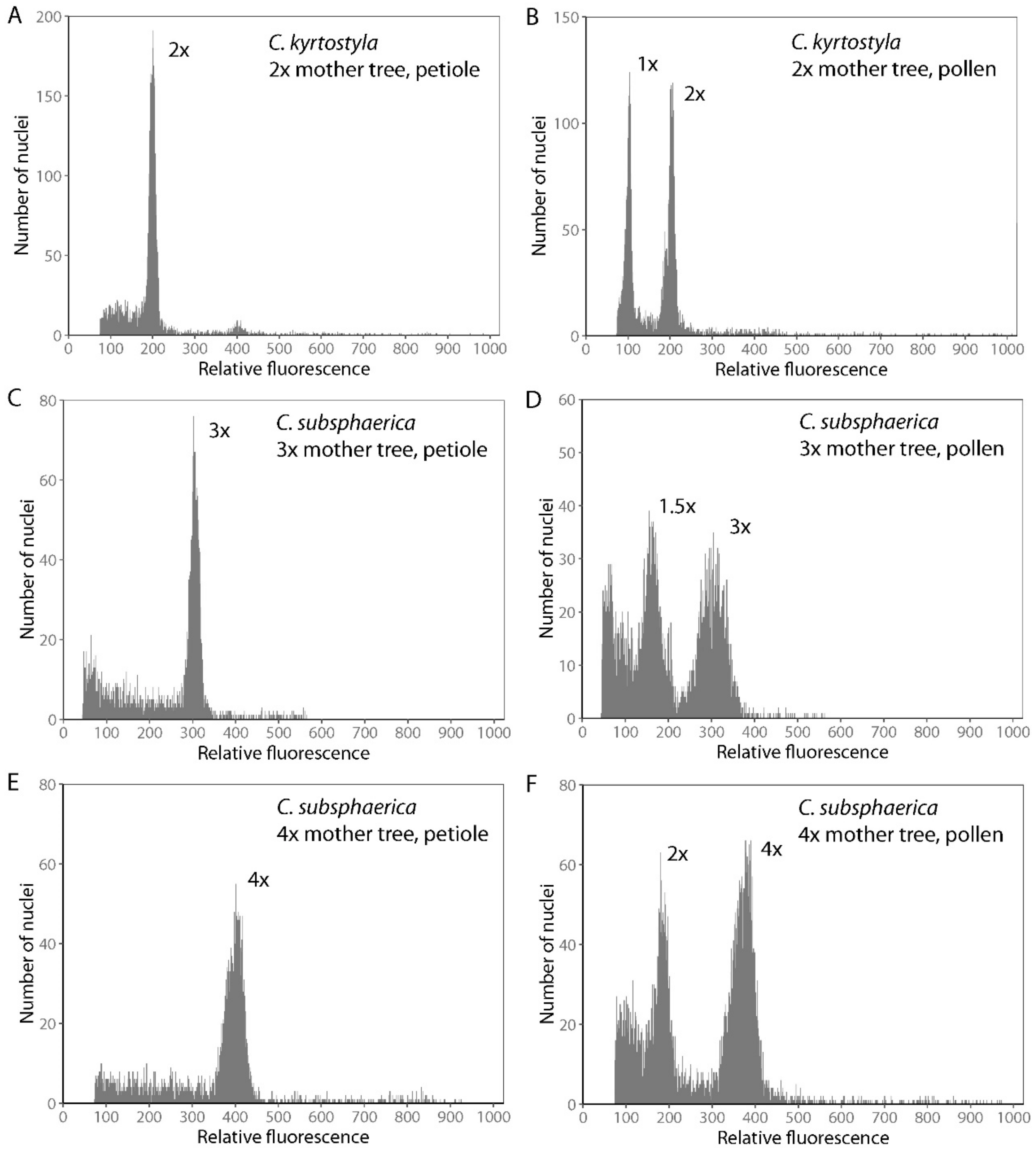
2.2. Determination of Ploidy Level of Crataegus Mother Trees and Their Pollen
2.3. Pollination Experiments
2.4. The Flow Cytometric Seed Screen
3. Results and Discussion
3.1. Cytogenetic Screening of Mother Trees and Their Progeny
3.2. The Flow Cytometric Screen of Crataegus Pollen
3.3. Pollination Experiments
3.4. Seed-Set Analysis and Reproductive Modes in Diploid Crataegus Mother Trees
3.5. Seed-Set Analysis and Reproductive Modes in Triploid Crataegus Mother Trees
3.6. Seed-Set Analysis and Reproductive Modes in Tetraploid Crataegus Mother Trees
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rieseberg, L.H.; Willis, J.H. Plant speciation. Science 2007, 317, 910–914. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J.; Flagel, L.E.; Paterson, A.H.; Rapp, R.A.; Soltis, D.E.; Soltis, P.S.; Wendel, J.F. Evolutionary genetics of genome merger and doubling in plants. Annu. Rev. Genet. 2008, 42, 443–461. [Google Scholar] [CrossRef] [PubMed]
- Soltis, P.S.; Soltis, D.E. The role of hybridization in plant speciation. Annu. Rev. Plant. Biol. 2009, 60, 561–588. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Wickett, N.J.; Ayyampalayam, S.; Chanderbali, A.S.; Landherr, L.; Ralph, P.E.; Tomsho, L.P.; Hu, Y.; Liang, H.; Soltis, P.S.; et al. Ancestral polyploidy in seed plants and angiosperms. Nature 2011, 473, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Husband, B.C.; Baldwin, S.J.; Suda, J. The incidence of polyploidy in natural plant populations: Major patterns and evolutionary processes. In Plant Genome Diversity: Physical Structure, Behaviour and Evolution of Plant Genomes; Leitch, I.J., Greilhuber, J., Doležel, J., Wendel, J.F., Eds.; Springer: Wien, Austria, 2013; Volume 2, pp. 255–276. [Google Scholar]
- Madlung, A. Polyploidy and its effect on evolutionary success: Old questions revisited with new tools. Heredity 2013, 110, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Alix, K.; Gérard, P.R.; Schwarzacher, T.; Heslop-Harrison, J.S.P. Polyploidy and interspecific hybridization: Partners for adaptation, speciation and evolution in plants. Ann. Bot. 2017, 120, 183–194. [Google Scholar] [CrossRef]
- Šarhanová, P.; Sharbel, T.F.; Sochor, M.; Vašut, R.J.; Dancák, M.; Trávnícek, B. Hybridization drives evolution in Rubus subgenus Rubus: Evidence from microsatellite markers. Ann. Bot. 2017, 120, 317–328. [Google Scholar] [CrossRef]
- Weiss-Schneeweiss, H.; Emadzade, K.; Jang, T.S.; Schneeweiss, G.M. Evolutionary consequences, constraints and potential of polyploidy in plants. Cytogenet. Genome Res. 2013, 140, 137–150. [Google Scholar] [CrossRef]
- Talent, N.; Dickinson, T.A. Endosperm formation in aposporous Crataegus (Rosaceae, Spiraeoideae, tribe Pyreae): Parallels to Ranunculaceae and Poaceae. New Phytol. 2007, 173, 231–249. [Google Scholar] [CrossRef]
- Talent, N.; Dickinson, T.A. The potential for ploidy level increases and decreases in Crataegus (Rosaceae, Spiraeoideae, tribe Pyreae). Can. J. Bot. 2007, 85, 570–584. [Google Scholar] [CrossRef]
- Whitton, J.; Sears, C.J.; Baack, E.J.; Otto, S.P. The dynamic nature of apomixis in the angiosperms. Int. J. Plant Sci. 2008, 169, 169–182. [Google Scholar] [CrossRef]
- Hörandl, E. A combinational theory for maintenance of sex. Heredity 2009, 103, 445–457. [Google Scholar] [CrossRef] [PubMed]
- Hojsgaard, D.; Greilhuber, J.; Pellino, M.; Paun, O.; Sharbel, T.F.; Hörandl, E. Emergence of apospory and bypass of meiosis via apomixis after sexual hybridisation and polyploidisation. New Phytol. 2014, 204, 1000–1012. [Google Scholar] [CrossRef] [PubMed]
- Hörandl, E. The complex causality of geographical parthenogenesis. New Phytol. 2006, 171, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Cosendai, A.C.; Hörandl, E. Cytotype stability, facultative apomixis and geographical parthenogenesis in Ranunculus kuepferi (Ranunculaceae). Ann. Bot. 2010, 105, 457–470. [Google Scholar] [CrossRef]
- Marhold, K.; Lihová, J. Polyploidy, hybridization and reticulate evolution: Lessons from the Brassicaceae. Plant Syst. Evol. 2006, 259, 143–174. [Google Scholar] [CrossRef]
- Dickinson, T.A.; Lo, E.; Talent, N. Polyploidy, reproductive biology, and Rosaceae: Understanding evolution and making classifications. Plant Syst. Evol. 2007, 266, 59–78. [Google Scholar] [CrossRef]
- Alix, K.; Joets, J.; Ryder, C.D.; Moore, J.; Barker, G.C.; Bailey, J.P.; King, G.J.; Pat Heslop-Harrison, J.S. The CACTA transposon Bot1 played a major role in Brassica genome divergence and gene proliferation. Plant J. 2008, 56, 1030–1044. [Google Scholar] [CrossRef]
- Husband, B.C.; Ozimec, B.; Martin, S.L.; Pollock, L. Mating consequences of polyploid evolution in flowering plants: Current trends and insights from synthetic polyploids. Int. J. Plant Sci. 2008, 169, 195–206. [Google Scholar] [CrossRef]
- Robertson, K.; Goldberg, E.E.; Igić, B. Comparative evidence for the correlated evolution of polyploidy and self-compatibility in Solanaceae. Evolution 2011, 65, 139–155. [Google Scholar] [CrossRef]
- Ludwig, S.; Robertson, A.; Rich, T.C.G.; Djordević, M.; Cerović, R.; Houston, L.; Harris, S.A.; Hiscock, S.J. Breeding systems, hybridization and continuing evolution in Avon Gorge Sorbus. Ann. Bot. 2013, 111, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Asker, S.E.; Jerling, L. Apomixis in Plants, 1st ed.; CRC Press: Boca Raton, FL, USA, 1992; p. 320. [Google Scholar]
- Simon, J.C.; Delmotte, F.; Rispe, C.; Crease, T. Phylogenetic relationships between parthenogens and their sexual relatives: The possible routes to parthenogenesis in animals. Biol. J. Linn. Soc. 2003, 79, 151–163. [Google Scholar] [CrossRef]
- Šarhanová, P.; Vašut, R.J.; Dančák, M.; Bureš, P.; Trávníček, B. New insights into the variability of reproduction modes in European populations of Rubus subgen. Rubus: How sexual are the polyploid brambles? Sex. Plant Reprod. 2012, 25, 319–335. [Google Scholar] [CrossRef] [PubMed]
- Lovell, J.T.; Aliyu, O.M.; Mau, M.; Schranz, M.E.; Koch, M.; Kiefer, C.; Song, B.-H.; Mitchell-Olds, T.; Sharbel, T.F. On the origin and evolution of apomixis in Boechera. Plant Reprod. 2013, 26, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Stebbins, G.L. Variation and Evolution in Plants; Columbia University Press: New York, NY, USA, 1950. [Google Scholar]
- Hörandl, E. Geographical parthenogenesis: Opportunities for asexuality. In Lost Sex; Schön, I., Martens, K., Van Dijk, P.J., Eds.; Springer: Heidelberg, Germany, 2009; pp. 161–186. [Google Scholar]
- Hörandl, E. The classification of asexual organisms: Old myths, new facts, and a novel pluralistic approach. Taxon 2018, 67, 1066–1081. [Google Scholar] [CrossRef]
- Smith, J.M. The Evolution of Sex; Cambridge University Press: Cambridge, NY, USA, 1978. [Google Scholar]
- Nogler, G.A. Gametophytic apomixis. In Embryology of Angiosperms; Johri, B.M., Ed.; Springer: Berlin/Heidelberg, Germany, 1984; pp. 475–518. [Google Scholar]
- Coyne, J.A.; Orr, H.A. Speciation; Sinauer Associates, Inc.: Sunderland, MA, USA, 2004. [Google Scholar]
- Grant, V. Plant Speciation, 2nd ed.; Columbia University Press: New York, NY, USA, 1981. [Google Scholar]
- Evans, R.; Campbell, C.S. The origin of the apple subfamily (Rosaceae: Maloideae) is clarified by DNA sequence data from duplicated GBSSI genes. Am. J. Bot. 2002, 89, 1478–1484. [Google Scholar] [CrossRef]
- Kalkman, C. Rosaceae. In The Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2004; Volume 6, pp. 343–386. [Google Scholar]
- Campbell, C.S.; Evans, R.C.; Morgan, D.R.; Dickinson, T.A.; Arsenault, M.P. Phylogeny of subtribe Pyrinae (formerly the Maloideae, Rosaceae): Limited resolution of a complex evolutionary history. Plant Syst. Evol. 2007, 266, 119–145. [Google Scholar] [CrossRef]
- Potter, D.; Eriksson, T.; Evans, R.C.; Oh, S.; Smedmark, J.E.E.; Morgan, D.R.; Kerr, M.; Robertson, K.R.; Arsenault, M.; Dickinson, T.A.; et al. Phylogeny and classification of Rosaceae. Plant Syst. Evol. 2007, 266, 5–43. [Google Scholar] [CrossRef]
- Reveal, J.L. Newly required infrafamilial names mandated by changes in the Code of Nomenclature for Algae, Fungi, and Plants. Phytoneuron 2012, 33, 1–32. [Google Scholar]
- Bailey, L.H. The Standard Cyclopedia of Horticulture, 2nd ed.; The Macmillan Company: New York, NY, USA, 1963. [Google Scholar]
- Phipps, J.B.; Robertson, K.R.; Smith, P.G.; Roher, J.R. A checklist of the subfamily Maloideae (Rosaceae). Can. J. Bot. 1990, 68, 2209–2269. [Google Scholar] [CrossRef]
- Christensen, K.I. Revision of Crataegus sect. Crataegus and nothosect. Crataeguineae (Rosaceae-Maloideae) in the Old World. Syst. Bot. Monogr. 1992, 35, 1–199. [Google Scholar] [CrossRef]
- Fineschi, S.; Salvini, D.; Turchini, D.; Pastorelli, R.; Vendramin, G.G. Crataegus monogyna Jacq. and C. laevigata (Poir.) DC. (Rosaceae, Maloideae) display low level of genetic diversity assessed by chloroplast markers. Plant Syst. Evol. 2005, 250, 187–196. [Google Scholar] [CrossRef]
- Depypere, L.; Vander Mijnsbrugge, K.; De Cock, K.; Verschelde, P.; Quataert, P.; Van Slycken, J.; Goetghebeur, P. Indigenous species of Crataegus (Rosaceae-Maloideae) in Flanders (Belgium). An explorative morphometric study. Belg. J. Bot. 2006, 139, 139–152. [Google Scholar] [CrossRef]
- Camp, W.H. The Crataegus problem. Castanea 1942, 7, 51–55. [Google Scholar]
- Zarrei, M.; Stefanović, S.; Dickinson, T.A. Reticulate evolution in North American black-fruited hawthorns (Crataegus section Douglasia; Rosaceae): Evidence from nuclear ITS2 and plastid sequences. Ann. Bot. 2014, 114, 253–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baranec, T. Crataegus L. In Flóra Slovenska IV/3; Bertová, L., Ed.; Veda: Bratislava, Slovakia, 1992; pp. 465–491. [Google Scholar]
- Li, L.-Z.; Gao, P.-Y.; Song, S.-J.; Yuan, Y.-Q.; Liu, C.-T.; Huang, X.-X.; Liu, Q.-B. Monoterpenes and flavones from the leaves of Crataegus pinnatifida with anticoagulant activities. J. Funct. Foods 2015, 12, 237–245. [Google Scholar] [CrossRef]
- Qiao, A.; Wang, Y.; Xiang, L.; Zhang, Z.; He, X. Novel triterpenoids isolated from hawthorn berries functioned as antioxidant and antiproliferative activities. J. Funct. Foods 2015, 13, 308–313. [Google Scholar] [CrossRef]
- Clapham, A.R.; Tutin, T.G.; Moore, D.M. Flora of the British Isles; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Lo, E.Y.Y.; Stefanović, S.; Ritland, K.; Dickinson, T.A. Fine-scale comparisons of genetic variability in seed families of asexually and sexually reproducing Crataegus (Hawthorn; Rosaceae). Am. J. Bot. 2010, 97, 1014–1024. [Google Scholar] [CrossRef] [Green Version]
- Lewis, D. Competition and dominance of incompatibility alleles in diploid pollen. Heredity 1947, 1, 85–108. [Google Scholar] [CrossRef]
- De Nettancourt, D. Incompatibility in angiosperms. Sex. Plant Reprod. 1977, 10, 185–199. [Google Scholar] [CrossRef]
- Raspé, O.; Kohn, J.R. S-allele diversity in Sorbus aucuparia and Crataegus monogyna (Rosaceae: Maloideae). Heredity 2002, 88, 458–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erdelská, O.; Švubová, R.; Mártonfiová, L.; Lux, A. Embryológia krytosemenných rastlín; Veda: Bratislava, Slovakia, 2017; p. 100. [Google Scholar]
- Talent, N.; Dickinson, T.A. Polyploidy in Crataegus and Mespilus (Rosaceae, Maloideae): Evolutionary inferences from flow cytometry of nuclear DNA amounts. Can. J. Bot. 2005, 83, 1268–1304. [Google Scholar] [CrossRef] [Green Version]
- Maheshwari, P. An Introduction to the Embryology of Angiosperms; McGraw-Hill Book Company, Inc.: New York, NY, USA, 1950; pp. 87–95. [Google Scholar]
- Czapik, R. Problems of apomictic reproduction in the families Compositae and Rosaceae. Folia Geobot. Phytotax. 1996, 31, 381–387. [Google Scholar] [CrossRef]
- Muniyamma, M.; Phipps, J.B. Cytological proof of apomixis in Crataegus (Rosaceae). Am. J. Bot. 1979, 66, 149–155. [Google Scholar] [CrossRef]
- Smith, P.G.; Phipps, J.B. Studies in Crataegus (Rosaceae, Maloideae). XIX. Breeding behaviour in Ontario Crataegus series Rotundifoliae. Can. J. Bot. 1988, 66, 1914–1923. [Google Scholar] [CrossRef]
- Scott, R.J. Polyspermy in apomictic Crataegus: Yes and no. New Phytol. 2007, 173, 227–229. [Google Scholar] [CrossRef] [Green Version]
- Scott, R.J.; Spielman, M.; Bailey, J.; Dickinson, H.G. Parent-of-origin effects on seed development in Arabidopsis thaliana. Development 1998, 125, 3329–3341. [Google Scholar]
- Kradolfer, D.; Hennig, L.; Köhler, C. Increased maternal genome dosage bypasses the requirement of the FIS Polycomb Repressive Complex 2 in Arabidopsis seed development. PLoS Genet. 2013, 9, e1003163. [Google Scholar] [CrossRef] [Green Version]
- Savidan, Y.H. Apomixis: Genetics and breeding. Plant Breed. Rev. 2000, 18, 13–86. [Google Scholar] [CrossRef]
- Lo, E.Y.Y.; Stefanović, S.; Dickinson, T.A. Population genetic structure of diploid sexual and polyploid apomicts of Crataegus (hawthorns; Rosaceae) in the Pacific Northwest. Mol. Ecol. 2009, 18, 1145–1160. [Google Scholar] [CrossRef]
- Lo, E.Y.Y.; Stefanović, S.; Dickinson, T.A. Reconstructing reticulation history in a phylogenetic framework and the potential of allopatric speciation driven by polyploidy in an agamic complex in Crataegus (Rosaceae). Evolution 2010, 64, 3593–3608. [Google Scholar] [CrossRef] [PubMed]
- Lo, E.Y.Y.; Stefanović, S.; Dickinson, T.A. Geographical parthenogenesis in Pacific Northwest hawthorns (Crataegus; Rosaceae). Botany 2013, 91, 107–116. [Google Scholar] [CrossRef]
- McGoey, B.V.; Chau, K.; Dickinson, T.A. Stomata size in relation to ploidy level in North American hawthorns (Crataegus, Rosaceae). Madroño 2014, 61, 177–193. [Google Scholar] [CrossRef]
- Coughlan, J.M.; Han, S.; Stefanović, S.; Dickinson, T.A. Widespread generalist clones are associated with range and niche expansion in allopolyploids of Pacific Northwest hawthorns (Crataegus L.). Mol. Ecol. 2017, 26, 5484–5499. [Google Scholar] [CrossRef] [PubMed]
- Brown, H.B. The genus Crataegus with some theories of the origin of its species. Bull. Torrey Bot. Club 1910, 37, 251–260. [Google Scholar] [CrossRef]
- Love, R.; Feigen, M. Interspecific hybridization between native and naturalized Crataegus (Rosaceae) in western Oregon. Madroño 1978, 25, 211–217. [Google Scholar]
- Byatt, J.I. Hybridization between Crataegus monogyna Jacq. and C. laevigata (Poiret) DC. in southeastern England. Watsonia 1975, 10, 253–264. [Google Scholar]
- Christensen, K.I. A biometric study of some hybridizing Crataegus populations in Denmark. Nord. J. Bot. 1983, 2, 537–548. [Google Scholar] [CrossRef]
- Christensen, K.I. A reanalysis of the status of Crataegus eremitagensis, C. raavadensis and C. schumacheri (Rosaceae). Symb. Bot. Ups. 1996, 31, 211–220. [Google Scholar]
- Christensen, K.I.; Zarrei, M.; Kuzmina, M.; Talent, N.; Lin, C.; Dickinson, T.A. Crataegus ×ninae-celottiae and C. ×cogswellii (Rosaceae, Maleae), two spontaneously formed intersectional nothospecies. PhytoKeys 2014, 36, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Ptak, K. Cyto-embryological investigations on the Polish representatives of the genus Crataegus L. I. Chromosome numbers; embryology of diploid and tetraploid species. Acta Biol. Cracov. Ser. Bot. 1986, 28, 107–122. [Google Scholar]
- Ptak, K. Cyto-embryological investigations on the Polish representatives of the genus Crataegus L. II. Embryology of triploid species. Acta Biol. Cracov. Ser. Bot. 1989, 31, 97–112. [Google Scholar]
- Loureiro, J.; Rodriguez, E.; Doležel, J.; Santos, C. Two new nuclear isolation buffers for plant DNA flow cytometry: A test with 37 species. Ann. Bot. 2007, 100, 875–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doležel, J.; Sgorbati, S.; Lucretti, S. Comparison of three DNA fluorochromes for flow cytometric estimation of nuclear DNA content in plants. Physiol. Plant. 1992, 85, 625–631. [Google Scholar] [CrossRef]
- Galbraith, D.W.; Harkins, K.R.; Maddox, J.M.; Ayres, N.M.; Sharma, D.P.; Firoozabady, E. Rapid flow cytometric analysis of the cell cycle in intact plant tissues. Science 1983, 220, 1049–1051. [Google Scholar] [CrossRef]
- Kron, P.; Husband, B.C. Using flow cytometry to estimate pollen DNA content: Improved methodology and applications. Ann. Bot. 2012, 110, 1067–1078. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, T.A.; Phipps, J.B. Studies in Crataegus (Rosaceae: Maloideae). XIV. The breeding system of Crataegus crus-galli sensu lato in Ontario. Am. J. Bot. 1986, 73, 116–130. [Google Scholar] [CrossRef]
- Dobeš, C.; Lückl, A.; Hülber, K.; Paule, J. Prospects and limits of the flow cytometric seed screen-insights from Potentilla sensu lato (Potentilleae, Rosaceae). New Phytol. 2013, 198, 605–616. [Google Scholar] [CrossRef] [Green Version]
- Matzk, F.; Meister, A.; Schubert, I. An efficient screen for reproductive pathways using mature seeds of monocots and dicots. Plant J. 2000, 21, 97–108. [Google Scholar] [CrossRef]
- Kolarčik, V.; Kocová, V.; Vašková, D. Flow cytometric seed screen data are consistent with models of chromosome inheritance in asymmetrically compensating allopolyploids. Cytometry A 2018, 93, 737–748. [Google Scholar] [CrossRef] [Green Version]
- Temsch, E.M.; Greilhuber, J.; Krisai, R. Genome size in liverworts. Preslia 2010, 82, 63–80. [Google Scholar]
- Krahulcová, A.; Rotreklová, O. Use of flow cytometry in research on apomictic plants. Preslia 2010, 82, 23–39. [Google Scholar]
- Brewbaker, J.L. The distribution and phylogenetic significance of binucleate and trinucleate pollen grains in the Angiosperms. Am. J. Bot. 1967, 54, 1069–1083. [Google Scholar] [CrossRef]
- Bino, R.J.; Van Tuyl, J.M.; De Vries, J.N. Flow cytometric determination of relative nuclear DNA contents in bicellulate and tricellulate pollen. Ann. Bot. 1990, 65, 3–8. [Google Scholar] [CrossRef]
- Lepší, M.; Koutecký, P.; Nosková, J.; Lepší, P.; Urfus, T.; Rich, T.C.G. Versatility of reproductive modes and ploidy level interactions in Sorbus s.l. (Malinae, Rosaceae). Bot. J. Linn. Soc. 2019, 191, 502–522. [Google Scholar] [CrossRef]
- Dickinson, T.A. Crataegu Crus-Galli L. Sensu Lato in Southern Ontario: Phenotypic Variation and Variability in Relation to Reproductive Behaviour. Ph.D. Thesis, University of Western Ontario, London, ON, Canada, 1983. [Google Scholar]
- Celotti, N. The Pollen Tube Pathway and Obturator in Hawthorn Sexual Reproduction. Ph.D. Thesis, Queen’s University, Kingston, ON, Canada, 1995. [Google Scholar]
- Purich, M.A. Characterizing Hybridization between Native and Non-Native Crataegus Species. Master’s Thesis, University of Toronto, Toronto, ON, Canada, 2005. [Google Scholar]
- Luu, D.T.; Qin, X.K.; Morse, D.; Cappadocia, M. S-RNase uptake by compatible pollen tubes in gametophytic self-incompatibility. Nature 2000, 407, 649–651. [Google Scholar] [CrossRef]
- Delgado, L.; Galdeano, F.; Sartor, M.E.; Quarin, C.L.; Espinoza, F.; Ortiz, J.P. Analysis of variation for apomictic reproduction in diploid Paspalum rufum. Ann. Bot. 2014, 113, 1211–1218. [Google Scholar] [CrossRef]
- Böcher, T.W. Cytological and embryological studies in the amphi-apomictic Arabis holboellii complex. Kongel. Danske Vidensk.-Selskab. Biol. Skr. 1951, 6, 1–59. [Google Scholar]
- Böcher, T.W. Experimental taxonomical studies in the Arabis holboellii complex. Sven. Bot. Tidskr. 1954, 48, 31–44. [Google Scholar]
- Sharbel, T.F.; Mitchell-Olds, T. Recurrent polyploid origins and chloroplast phylogeography in the Arabis holboellii complex (Brassicaceae). Heredity 2001, 87, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Burgess, M.B.; Cushman, K.R.; Doucette, E.T.; Talent, N.; Frye, C.T.; Campbell, C.S. Effects of apomixis and polyploidy on diversification and geographic distribution in Amelanchier (Rosaceae). Am. J. Bot. 2014, 101, 1375–1387. [Google Scholar] [CrossRef] [PubMed]
- Sochor, M.; Trávníček, B. Melting pot of diversity: First insights into evolutionary patterns of the Colchic bramble flora (Rubus subgenus Rubus, Rosaceae). Bot. J. Linn. Soc. 2016, 181, 610–620. [Google Scholar] [CrossRef] [Green Version]
- Hajrudinović, A.; Siljak-Yakovlev, S.; Brown, S.C.; Pustahija, F.; Bourge, M.; Ballian, D.; Bogunić, F. When sexual meets apomict: Genome size, ploidy level and reproductive mode variation of Sorbus aria s.l. and S. austriaca (Rosaceae) in Bosnia and Herzegovina. Ann. Bot. 2015, 116, 301–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramsey, J.; Schemske, D.W. Pathways, mechanisms, and rates of polyploid formation in flowering plants. Annu. Rev. Ecol. Syst. 1998, 29, 467–501. [Google Scholar] [CrossRef] [Green Version]
- Singh, R. Plant Cytogenetics; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Henry, I.M.; Dilkes, B.P.; Young, K.; Watson, B.; Wu, H.; Comai, L. Aneuploidy and genetic variation in the Arabidopsis thaliana triploid response. Genetics 2005, 170, 1979–1988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramsey, J.; Schemske, D.W. Neopolyploidy in flowering plants. Annu. Rev. Ecol. Syst. 2002, 33, 589–639. [Google Scholar] [CrossRef] [Green Version]
- Henry, I.M.; Dilkes, B.P.; Comai, L. Molecular karyotyping and aneuploidy detection in Arabidopsis thaliana using quantitative fluorescent polymerase chain reaction. Plant J. 2006, 48, 307–319. [Google Scholar] [CrossRef]
- Henry, I.M.; Dilkes, B.P.; Tyagi, A.P.; Lin, H.-Y.; Comai, L. Dosage and parent-of-origin effects shaping aneuploid swarms in A. thaliana. Heredity 2009, 103, 458–468. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Huo, B.; Liu, W.; Li, D.; Liao, L. Abnormal meiosis in an intersectional allotriploid of Populus L. and segregation of ploidy levels in 2x × 3x progeny. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [Green Version]
- De Storme, N.; Mason, A. Plant speciation through chromosome instability and ploidy change: Cellular mechanisms, molecular factors and evolutionary relevance. Curr. Plant Biol. 2014, 1, 10–33. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, T.A. Sex and Rosaceae apomicts. Taxon 2018, 67, 1093–1107. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, T.A.; Belaoussoff, S.; Love, R.M.; Muniyamma, M. North American black-fruited hawthorns: I. Variation in floral construction, breeding system correlates, and their possible evolutionary significance in Crataegus sect. Douglasii Loudon. Folia Geobot. Phytotax. 1996, 31, 355–371. [Google Scholar] [CrossRef]
- Muniyamma, M.; Phipps, J.B. Studies in Crataegus. XI. Further cytological evidence for the occurrence of apomixis in North American hawthorns. Can. J. Bot. 1984, 62, 2316–2324. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality, Date of Experiments, Population Codes, Cytotype Diversity [%] | Taxon | No. Trees | FCSS No. Seeds | |
|---|---|---|---|---|
| Contr. Poll. | Open Poll. | |||
| Slovakia, Prešov, south-eastern margin of village Hermanovce, 49°02′31.7″, 21°01′15.3″; 30.5.2017–31.5.2017; GOCAL; 2x–0.00, 3x–34.04, 4x–65.96 (n = 47) | C. kyrtostyla (2x) | 2 | 4 | 11 |
| Slovakia, Prešov, north-eastern margin of village Hermanovce, 49°03′06.8″, 21°00′22.5″; 30.5.2017-31.5.2017; VYH, PS-VYH; 2x–30.00, 3x–20.00, 4x–50.00 (n = 10) | C. subsphaerica (3x) | 1 | 7 | 5 |
| C. laevigata × C. subsphaerica (3x) | 2 | 0 | 8 | |
| C. subsphaerica (4x) | 5 | 21 | 30 | |
| Slovakia, Košice, Botanical garden of Pavol Jozef Šafárik University, 48°44′09.3″, 21°14′08.6″; 27.4.2017–2.5.2017; BOTZ; 2x–88.89, 3x–11.11, 4x–0.00 (n = 9) | C. monogyna (2x) | 2 | 49 | 41 |
| C. kyrtostyla (2x) | 1 | 2 | 20 | |
| C. subsphaerica (3x) | 1 | 12 | 22 | |
| TOTAL | 14 | 95 | 137 | |
| Type of Pollination/Ploidy Level ♀ × ♂ | Number of Seeds (Number of Pollinations) | Seed Origin | Embryo Ploidy | Endosperm Ploidy | Egg Ploidy | Central Cell Ploidy | Pollen Ploidy (Sperm Cell) | Embryo Origin (egg Cell + Sperm Cell) | Endosperm Origin (Central Cell + Sperm cell) | |
|---|---|---|---|---|---|---|---|---|---|---|
| 2x (intact flowers bagged) | 1 (475) | Sexual | 2x | 3x | 1x | 2x | 1x | (1x) + 1x | (2x) + 1x | |
| 2x × 2x (self-pollination) | 1 (131) | Sexual | 2x | 3x | 1x | 2x | 1x | (1x) + 1x | (2x) + 1x | |
| 2x × 2x (cross-pollination) | 52 (185) | Sexual | 2x | 3x | 1x | 2x | 1x | (1x) + 1x | (2x) + 1x | |
| 2x (open pollination) | 72 | Sexual | 2x | 3x | 1x | 2x | 1x | (1x) + 1x | (2x) + 1x | |
| 2x × 3x (cross-pollination) | 1 (40) | Sexual | 2x | 3x | 1x | 2x | 1x | (1x) + 1x | (2x) + 1x | |
| 3x (intact flowers bagged) | 0 (221) | / | / | / | / | / | / | / | / | |
| 3x × 3x (self-pollination) | 0 (61) | / | / | / | / | / | / | / | / | |
| 3x × 3x (cross-pollination) | 2 (38) | Apomictic | 3x | 9x | 3x | 6x | 1.5x | (3x) + 0 | (6x) + 3x or (6x) + 2×1.5x | |
| 3x (open pollination) | 1 | Apomictica | 3x | 5x | 3x | 3x | 1x | (3x) + 0 | (3x) + 2×1x | |
| 2 | Apomictic | 3x | 7x | 3x | 6x | 1x | (3x) + 0 | (6x) + 1x | ||
| 22 | Apomictic | 3x | 8x | 3x | 6x | 2x | (3x) + 0 | (6x) + 2x | ||
| 8 | Apomictic | 3x | 9x | 3x | 6x | 1.5x | (3x) + 0 | (6x) + 1×3x or (6x) + 2×1.5x | ||
| 2 | Apomictic | 3x | 10x | 3x | 6x | 2x | (3x) + 0 | (6x) + 2×2x | ||
| 3x × 2x (cross-pollination) | 1 | Apomictic | 3x | 6x | 3x | 6x | / | (3x) + 0 | (6x) + 0 | |
| 1 | Apomictic | 3x | 7x | 3x | 6x | 1x | (3x) + 0 | (6x) + 1x | ||
| 12 | (118) | Apomictic | 3x | 8x | 3x | 6x | 1x | (3x) + 0 | (6x) + 1×2x or (6x) + 2×1x | |
| 2 | Sexualb | ~3.7x | ~5x | ~1.7x | ~3.4x | ~2x | ~(1.7x) + ~2x | ~(3.4x) + ~2x | ||
| 1 | Sexual, BIII | 4x | / | 3x | / | / | (3x) + 1x | / | ||
| 4x (intact flowers bagged) | 2 | (101) | Apomictic | 4x | 10x | 4x | 8x | 2x | (4x) + 0 | (8x) + 2x |
| 19 | Apomictic | 4x | 12x | 4x | 8x | 2x | (4x) + 0 | (8x) + 1×4x or (8x) + 2×2x | ||
| 4x (open pollination) | 1 | Apomictic, haploid parthenogenesis | 2x | 8x | 2x | 4x | 2x | (2x) + 0 | (4x) + 1×4x or (4x) + 2×2x | |
| 1 | Sexual | 4x | 6x | 2x | 4x | 2x | (2x) + 2x | (4x) + 2x | ||
| 1 | Sexual, BIII | 6x | 10x | 4x | 8x | 2x | (4x) + 2x | (8x) + 2x | ||
| 4 | Apomictic | 4x | 10x | 4x | 8x | 2x | (4x) + 0 | (8x) + 2x | ||
| 1 | Apomictic | 4x | 11x | 4x | 8x | 3x | (4x) + 0 | (8x) + 3x | ||
| 18 | Apomictic | 4x | 12x | 4x | 8x | 2x | (4x) + 0 | (8x) + 1×4x or (8x) + 2×2x | ||
| 3 | Apomictic | 4x | 14x | 4x | 12x | 2x | (4x) + 0 | (12x) + 2x | ||
| 1 | Apomictic | 4x | 16x | 4x | 12x | 2x | (4x) + 0 | (12x) + 1×4x or (12x) + 2×2x | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vašková, D.; Kolarčik, V. Breeding Systems in Diploid and Polyploid Hawthorns (Crataegus): Evidence from Experimental Pollinations of C. monogyna, C. subsphaerica, and Natural Hybrids. Forests 2019, 10, 1059. https://doi.org/10.3390/f10121059
Vašková D, Kolarčik V. Breeding Systems in Diploid and Polyploid Hawthorns (Crataegus): Evidence from Experimental Pollinations of C. monogyna, C. subsphaerica, and Natural Hybrids. Forests. 2019; 10(12):1059. https://doi.org/10.3390/f10121059
Chicago/Turabian StyleVašková, Dominika, and Vladislav Kolarčik. 2019. "Breeding Systems in Diploid and Polyploid Hawthorns (Crataegus): Evidence from Experimental Pollinations of C. monogyna, C. subsphaerica, and Natural Hybrids" Forests 10, no. 12: 1059. https://doi.org/10.3390/f10121059